Introduction
Peripheral nerve injury is a common disorder of the
nervous system and affects ~13 to 22 individuals per 100,000 each
year, which has become a severe public health issue worldwide
(1). Furthermore, the amount of
patients suffering from peripheral nerve injuries is expected to
increase. It is generally recognized that the peripheral nervous
system is different from the central nervous system in that nerve
regeneration will be activated if peripheral neurons are damaged
(2). However, even with the
instinctive regeneration capability of peripheral neurons,
efficient therapy for peripheral nerve injury remains unavailable.
Thus, it is necessary to develop novel and effective therapies to
promote axon regeneration after peripheral nerve injury. In order
to fulfill such a purpose, the comprehensive investigation of the
changes in lesion neurons and identification of the biological
molecules that are responsible for neuronal survival and axonal
regeneration demands a prompt solution.
The intrinsic regenerative capacity of a peripheral
nerve is always activated after injury to that nerve. Generally,
this regeneration process depends on axonal degeneration,
phagocytosis of axonal debris, and secretion of trophic factors and
cytokines by Schwann cells (SCs) (3). Among all these factors important to
the regeneration of peripheral neurons, SCs, originating from the
neural crest, serve a determinant role in the intrinsic
regenerative capacity of the peripheral nervous system due to its
high abundance and potency in repairing the lesion sites in this
location (2). In previous
studies, it has been shown that SCs can be employed for transplants
in the spinal cord for nerve repair (4,5).
Moreover, SCs are able to maintain the peripheral nerve fibers.
Malfunctions of SCs are reported to be associated with peripheral
nerve injuries and various neuropathy conditions, including
hereditary, metabolic and inflammatory conditions (6,7).
Currently, skin-derived precursors (SKPs) are an excellent resource
for the isolation of SCs that can be utilized as outstanding cell
models suited for investigation of the mechanism involved in nerve
regeneration. Additionally, SKP-derived SCs (SKP-SCs) have been
explored for their potential application in the scenario of
peripheral nerve injury (8). With
the exception of the application of SKP-SCs alone, only
concatenated application of the acellular nerve allograft (ANA)
injection with SKP-SC has been presented as a better modality for
nerve repair compared with intrinsic regeneration (9,10).
In addition, in our previous studies, it was found that injection
of heregulin-1β, a type of pleiotropic growth factor or neuregulin,
further promotes the outcome of reinnervation by the ANA with
SKP-SC therapy via regulation of neuronal and glial development
(11,12). However, even though the treatment
of peripheral nerve injuries with heregulin-1β achieves a notable
outcome, the underlying mechanism through which heregulin-1β
facilitates ANA with SKP-SC therapy remains unclear.
Recently, multiple studies have suggested that the
mechanism asso ciated with injuries to the peripheral nerves
depends on the function of various non-coding RNAs (13–16). Among which, long non-coding RNAs
(lncRNAs) have been shown to be involved in various biological
processes and disease pathologies (17). Considering the promising effect of
heregulin-1β in treating peripheral nerve injures and the multiple
functions of lncRNA, it is reasonable to investigate the
association between these two factors in a peripheral nerve
injuries model. Furthermore, although a number of lncRNAs have been
identified to serve key roles in gene transcription and translation
in mammalian cells (18), few
lncRNAs that may be associated with the repair of peripheral nerve
injuries have been found.
Therefore, in the present study, the association
between lncRNA expression profiles and nerve repair was detected
using microarray assays in a well-built rat model with sciatic
nerve injury. The ANA injection with SKP-SC and the ANA injection
with SKP-SC plus heregulin-1β were used to treat the sciatic nerve
injury. Moreover, the specific functions of lncRNA BC088327 in
peripheral nerve injuries were further assessed using cell
viability, cell cycle and apoptosis assays in a hypoxic SC model.
The findings outlined in the present study demonstrated that the
expression profile of lncRNAs was closely associated with the
presence of heregulin-1β, and that lncRNA BC088327 may play a
synergistic role with heregulin-1β in repairing peripheral nerve
injury.
Materials and methods
Cell isolation and culture
SKPs were generated from the dermis of postnatal day
Wistar rats and cultured routinely according to a protocol
described by previous studies (6,19).
The isolated SKPs were identified with the staining of nestin and
fibronectin on the cell surface. Cultured SKP-SCs were fixed in
ice-cold absolute methanol for 20 min and blocked with 1% goat
serum for 1 h. Cells were then incubated with mouse monoclonal
anti-Nestin (1:100; cat. no. ab6142; Abcam, San Francisco, CA,
USA), rabbit polyclonal anti-fibronectin (1:100; cat. no. ab2413;
Abcam) at 4°C overnight and visualized with anti-rabbit IgG (H+L)
F(ab′)2 Fragment (Alexa Fluor 488 conjugate) (1:500; cat. no. 4412;
Cell Signaling Technology, Danvers, MA, USA) and anti-mouse IgG
(H+L) F(ab′)2 Fragment (Alexa Fluor 488 conjugate) (1:500; cat. no.
4408; Cell Signaling Technology). SCs were induced by incubating
SKPs on poly-D-lysine-laminin-coated culture dishes (Corning, Inc.,
Corning, NY, USA) in Dulbecco's modified Eagle's medium (DMEM)/F12,
with 4 mM forksolin, 10 ng/ml heregulin-1β (cat. no. 4711-50;
Biovision, Inc., Milpitas, CA, USA) and 1% N2 supplement
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) for 1
week. Thereafter, cells appearing to have bipolar SC morphology
under phase-contrast microscopy were isolated with cloning
cylinders and expanded in the same medium until >95% purity was
achieved (6), and then identified
by p75NGFR and S100-β staining, as previously described (12).
ANA procedure
A total of 20 adult male SD rats (weight, 200–250 g;
Shanghai SLAC Laboratory Animal Co., Ltd., Shanghai, China) were
anesthetized with chloral hydrate (350 mg/kg) and subjected to
sciatic nerve transection in order to mimic peripheral nerve
injury, as previously described (12). Rats were maintained under SPF
conditions including a 12 h light/12 h dark cycle, temperature of
20–22°C, and sterilized water and food. The rats were then treated
with ANAs, as described in a previous study (12). All surgical procedures were
performed bilaterally. Following the skin incision and a gluteal
splitting incision, the sciatic nerve and its branches were exposed
bilaterally. With the aid of a surgical microscope, the sciatic
nerve (20 mm) was severed and removed closed to the obturator
tendon in the mid-thigh. To prepare the ANA, the 20-mm sciatic
nerve was rinsed with Hank's balanced salt solution (HBSS) and
frozen in liquid nitrogen for 2 min following a 2-min rewarming
period twice, and then stored in liquid nitrogen. At 1 day prior to
transplantation, the ANA was incubated with 2 U/ml chondroitinase
ABC or phosphate-buffered saline (PBS) for 16 h at 37°C, then
washed 2 times with HBSS and stored at 0°C.
Animal care and surgery
The study was approved by the Institutional Animal
Care and Use Committee of Fudan University (Shanghai, China). Rats
were maintained under SPF conditions including a 12 h light/12 h
dark cycle, temperature of 20–22°C, and sterilized water and food.
Recipient rats were divided into the ANA+SKP-SC-treated group
(control group, n=8) and the ANA+SKP-SC+heregulin-1β-treated group
(experimental group, n=8), and underwent exposure of the right
sciatic nerve. The recipient's nerve was transected at 5 mm
proximal to the sciatic trifurcation and 15 mm of nerve was removed
under a surgical microscope, as described in our previous study
(12). For the two groups, 3 ml
of 2×106 cells/ml SKP-SCs were injected into the ANA at
the proximal end and at the distal end 3 mm from the anastomosis,
respectively, using a micro-injector under an SXP-10 microscope at
×10 magnification (Shanghai Medical Equipment Works Co., Ltd.,
Shanghai, China). Additionally, for the experimental group, 500
ng/ml heregulin-1β (3 ml) was injected into the ANA. At the end of
the 8th week, the sciatic nerve was harvested en bloc ~3 mm
proximal and ~4 mm distal to the interposed graft. The nerves were
stored at liquid nitrogen and transferred to a −80°C fridge for the
downstream application.
Microarray assays for differentially
expressed lncRNA and mRNA screening
The collected tissues were treated with TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.) and the total RNAs
were purified following the manufacturer's instructions. The
integrity of the purified RNAs was checked with NanoDrop 2000
(Thermo Fisher Scientific, Inc.). The qualified total RNA was
further purified by a RNeasy micro kit and RNase-free DNase set
(both Qiagen GmbH, Hilden, Germany). Next, the total RNA was
amplified, labeled and purified using the Affymetrix WT Amplication
kit and the GeneChip WT Terminal Labeling kit (both Affymetrix;
Thermo Fisher Scientific, Inc.) following the manufacturer's
instructions. Each labeled cRNA (1 µg) was fragmented by
adding 5 µl 10X blocking agent and 1 µl of 25X
fragmentation buffer, and then the mixture was heated at 60°C for
30 min. Finally, 25 µl 2X GE hybridization buffer was added
to dilute the labeled cRNA. Hybridization solution (50 µl)
was dispensed into the gasket slide and assembled into the lncRNA
expression microarray slide. The slides were incubated for 17 h at
65°C in an Agilent hybridization oven (Agilent Technologies, Inc.,
Santa Clara, CA, USA). The hybridized arrays were washed with 1X
washing buffer (0.05 mol/l Tris/HCl, 0.15 mol/l NaCl, 0.05% Tween
20, pH 7.6), and scanned using the Agilent DNA microarray scanner
(catalog no. G2505C; Agilent Technologies, Inc.).
Agilent Feature Extraction software (version
11.0.1.1) was used to analyze acquired array images. Quantile
normalization and subsequent data processing were performed using
the GeneSpring GX v12.1 software package (Agilent Technologies,
Inc.). After quantile normalization of the raw data, lncRNAs and
mRNAs with at least 1 out of 2 samples flagged as 'Present' or
'Marginal' ('All Targets Value') were chosen for further data
analysis. Differentially expressed lncRNAs and mRNAs between the
two groups were identified through fold-change filtering: Absolute
fold-change ≥2, ANA+SKP-SC+heregulin-1β-treated group vs.
ANA+SKP-SC-treated group. Hierarchical clustering was performed
using homemade scripts.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis for the differentially
expressed lncRNAs
Total RNA was reverse-transcribed into cDNA using
the Reverse Transcription kit (Roche Diagnostics GmbH, Mannheim,
Germany). qPCR was performed on an Applied Biosystems 7500
instrument using a 20-µl reaction volume containing 10
µl of 2X light cycler 480 SYBR-Green I Master (Roche
Diagnostics GmbH). Reactions were incubated in a 96-well optical
plate at 95°C for 10 min, followed by 45 cycles of 95°C for 15 sec
and 60°C for 60 sec. Each sample was represented by at least three
replicates for analysis. At the end of the PCR cycles, melting
curve analysis was conducted to validate the PCR products. The
primers for the specific amplification are listed in Table I. GAPDH was used as an internal
control (20).
 | Table IPrimers used in qauntitative
polymerase chain reaction analysis. |
Table I
Primers used in qauntitative
polymerase chain reaction analysis.
| Seqnames | Forward primer | Reverse primer |
|---|
| BC086339 |
CAGCGGGAGGCTCATACTACAT |
GGCTCCACAGCAGGAGACAA |
| XR_007612 |
ATGTTCCAGATCCCGTACCA |
TCTTCTAGTGTTTCCCTAAGTTTC |
| uc.19+ |
AATCGCATTACTGCCCTTG |
CTTGGGCTCTTAGCGTCTTA |
| DQ203294 |
AGAATCGGCACGAGGACCAA |
TGAAGCCAGGCATACTGAAC |
| Bc088327 |
ATGAATGTGGAAATCGGAAAC |
TTGGTAGGCTTGTCAGGGTC |
| uc.184+ |
TTCCAACAGTGAACATTTTTAGG |
GAGATATCCCGTCAGTGAAAAG |
| uc.75− |
CCAAAACACAATAAATGCTCTTCT |
TTTATGTGGCTCATTTTATATTTCC |
| S56464 |
ACCTCTTAGTGGGTTATTGGC |
TGGCTAACATCCTGGGTGAG |
| uc.366− |
CCGTCAAGTCTGCAAGGTCTC |
GGCAAGTCATTTATTATGGGTTCT |
| uc. 173+ |
TAATAAATGACTTGCCTCCTCG |
ATGCAGCAATAACTTAGTTCACAC |
| uc.10− |
TGAGTGTAGAGGAGCAGAGGC |
AAGGTACTGCGTGCATGAATA |
| uc.456− |
AAGATGACTGCCTTTCCTGAT |
TGTCTAACCCTTGGAATCCTC |
| uc.128− |
TACCAGTCCTGTCGGTTGC |
CCACAGAGCGGAAGTTGAT |
| D26496 |
GGAGCACGGCTATCACAGA |
CTAGTGAATCAAGTGGAGGGAG |
| uc.298− |
TTTAACAGTGGTTTTTCTTTTTTG |
CAGAAGAAAACTGTTGGGTGG |
| uc.28− |
TGAACAGGAATTAAGAATAAAGCTG |
GCGGGAACTACTCTGACAACA |
| uc.325− |
TGAGGCAGCGCATCGTG |
TGTCACAGGGCCGCACA |
| uc.299− |
CAGCAATCTTACCATGAGGTGT |
CTTTCCCGAAGGGTTATGTTAC |
| uc.66+ |
TAATAATATGGGATGACTGGGG |
TTGGTTGTTTTCTGTTTTGTGAT |
| uc.80+ |
TAATAAAAAGGAATCCATAAAAAAT |
ATAATTACAACTCAGGTTACGCTT |
| GAPDH |
TGTTCGTCATGGGTGTGAA |
ATGGCATGGACTGTGGTCAT |
Cell transfection
siRNA (siRNA, 5′-AAUAAAAUUGUAAAAAAGGAA-3′; NC,
5′-ACGUGACACGUUCGGAGAATT-3′) was provided by Sangon Biotech
(Shanghai, China). To induce hypoxic conditions in vitro,
SCs previously incubated with heregulin-1β were cultured at 37°C in
a humidified incubator with a mixture of 1% O2, 5%
CO2 and 94% N2 for 12, 24 or 36 h. Subsequent
to 72 h of incubation, the SCs were transferred to new DMEM and
transfected with siRNA targeted at lncRNA BC088327 (final
concentration, 100 nM) with Lipofectamine 2000®
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. After transfection for 48 h, the cells
were harvested for cell viability, cell cycle and cell apoptosis
assays.
Cell viability detection by Cell Counting
Kit-8 (CCK-8) and EdU assay
The negative control or lncRNA BC088327
siRNA-transfected SCs were plated in separate 96-well plates (3
wells/group) with DMEM for 72 h. For the CCK-8 assay, 10 µl
CCK-8 (Dojindo Molecular Technologies, Inc., Kumamoto, Japan) was
then added to randomly selected wells in each group every 24 h
(total time, 72 h), and the viability of the cells was measured at
490 nm using an ELISA reader (BioTek Instruments, Inc., Winooski,
VT, USA) according to the manufacturer's instructions. Each
time-point was represented by at least three replicates. Upon
completion of cell culture, the cell viabilities in different
groups were further confirmed using the Click-iT EdU kit (Molecular
Probes; Thermo Fisher Scientific, Inc.). The results were detected
by flow cytometry, described as follows.
Cell cycle assays
The SCs (1.0×106/well) were plated in
6-well plates with DMSO (10 µM) for 48 h. Prior to the cell
cycle assay, the cells were washed twice with ice-cold PBS and
fixed in 70% ethanol at 4°C overnight. For RNA removal, the cells
were be incubated with 10 mg/ml RNase A (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) at 37°C for 30 min, and then stained with 50
mg/ml propidium iodide (PI; Sigma-Aldrich; Merck KGaA). Cell cycle
distribution was assessed by flow cytometry (FC500; Beckman
Coulter, Inc., Miami, FL, USA). Each group was represented by at
least three replicates.
Quantification of apoptosis by flow
cytometry
Apoptosis was assessed using Annexin V/fluorescein
isothiocyanate (FITC)-PI measurement. The cells (1x105)
were washed twice with PBS (pH 7.4) and resuspended in staining
buffer containing 1 µg/ml PI and 0.025 µg/ml Annexin
V-FITC (cat. no. 556547; BD Biosciences, Franklin Lakes, NJ, USA).
Double-labeling was performed at room temperature for 10 min in the
dark prior to the flow cytometry analysis. The stained cells were
then analyzed with a flow cytometer (FC500; Beckman Coulter, Inc.).
Each group were represented by at least three replicates.
Statistical analysis
Data are expressed as the mean ± standard deviation
and were analyzed using SPSS 13.0 software (SPSS, Inc., Chicago,
IL, USA). An analysis of variance and a least significant
difference post hoc test were performed, with P<0.05 indicating
a statistically significant difference.
Results
Differentially expressed lncRNAs screened
by microarray assay
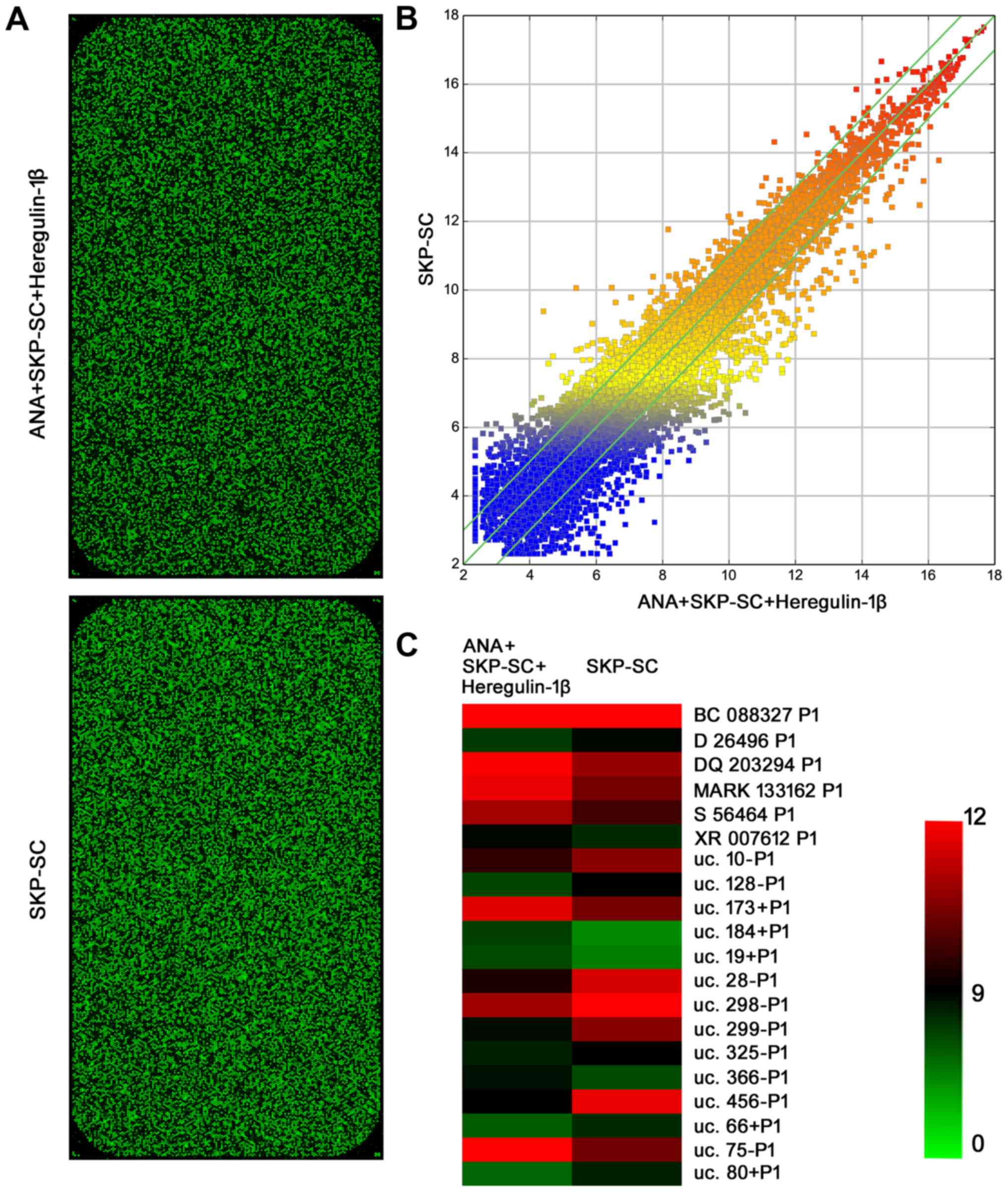
The results of the lncRNA microarray showed that a
total of 8,124 lncRNAs were detected in the rat model, of which,
805 lncRNAs were significantly differentially expressed (absolute
fold-change ≥2, ANA+SKP-SC+heregulin-1β-treated group vs.
ANA+SKP-SC-treated group). It was found that 323 lncRNAs were
upregulated and 482 lncRNAs were downregulated. The hierarchical
cluster analysis of the lncRNA microarray data is shown in Fig. 1. The mRNA microarray identified a
total of 13,112 mRNAs, in which 1,293 lncRNAs were significantly
differentially expressed (absolute fold-change ≥2,
ANA+SKP-SC+heregulin-1β-treated group vs. ANA+SKP-SC-treated
group), including 603 upregulated lncRNAs and 690 downregulated
lncRNAs (Table II). Based on the
fold-change data, 20 lncRNAs with an upregulated fold-change >5
and a downregulated fold-change >10 were screened by RT-qPCR
analysis (Table III).
| Table IISummary of differentially expressed
lncRNAs and mRNAs between treatment and control groups. |
Table II
Summary of differentially expressed
lncRNAs and mRNAs between treatment and control groups.
| Probe class | Differentially
expressed | Upregulated | Downregulated |
|---|
| lncRNA | 805 | 323 | 482 |
| mRNA | 1293 | 603 | 690 |
| Table IIIlncRNAs with significant differential
expression. |
Table III
lncRNAs with significant differential
expression.
| lncRNA | Relative
expression | Fold-change |
|---|
| BC086339 | Upregulated | 5.9482925 |
| XR_007612 | Upregulated | 5.9529158 |
| uc.19+ | Upregulated | 6.2218258 |
| DQ203294 | Upregulated | 6.7717983 |
| BC088327 | Upregulated | 8.9564234 |
| uc.184+ | Upregulated | 7.7992154 |
| uc.75− | Upregulated | 8.860086 |
| S56464 | Upregulated | 8.8825432 |
| uc.366− | Upregulated | 9.6759103 |
| uc.173+ | Upregulated | 12.2055043 |
| uc.10− | Downregulated | 11.5705296 |
| uc.456− | Downregulated | 12.0019673 |
| uc.128− | Downregulated | 12.614861 |
| D26496 | Downregulated | 13.01382 |
| uc.298− | Downregulated | 13.4702367 |
| uc.28− | Downregulated | 13.8457471 |
| uc.325− | Downregulated | 13.8457471 |
| uc.299− | Downregulated | 13.8457471 |
| uc.66+ | Downregulated | 13.8457471 |
| uc.80+ | Downregulated | 15.2508305 |
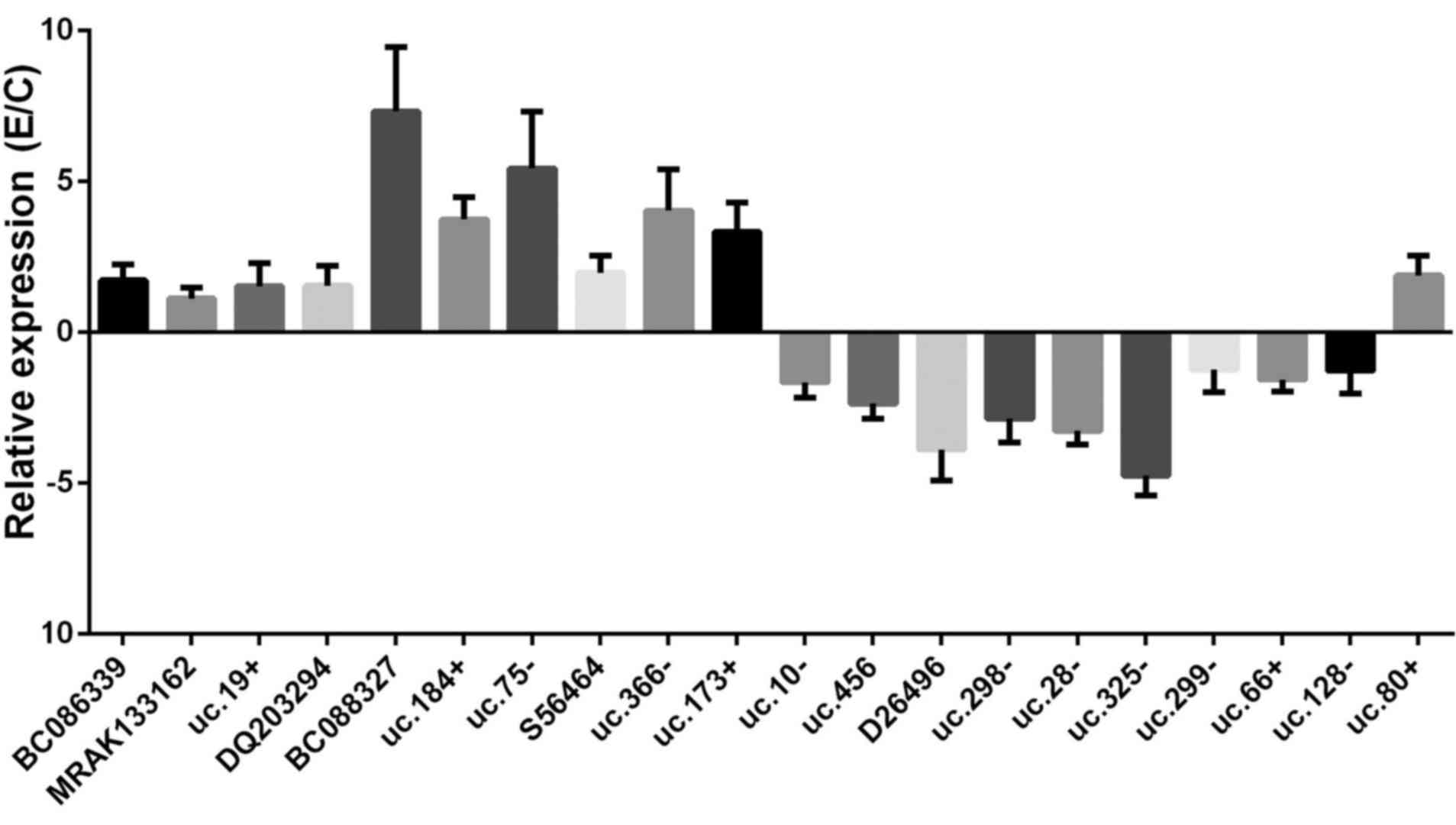
RT-qPCR analysis of the expression of the
lncRNAs
Following the preliminary analysis of the results
from the lncRNA profile, a total of 20 lncRNAs were analyzed by the
RT-qPCR assay (Fig. 2). The
results were normalized to a GAPDH control. The lncRNAs presented
with similar expression tendencies as those in the microarray
assay. Among them, the BC088327 and uc.75 exhibited the greatest
change in expression compared with the other lncRNAs, and the
extent of D26496 and uc.325 down-regulation was markedly greater
than that of the other lncRNAs.
BC088327 serves important roles in nerve
cell proliferation
To further study the functions of the differentially
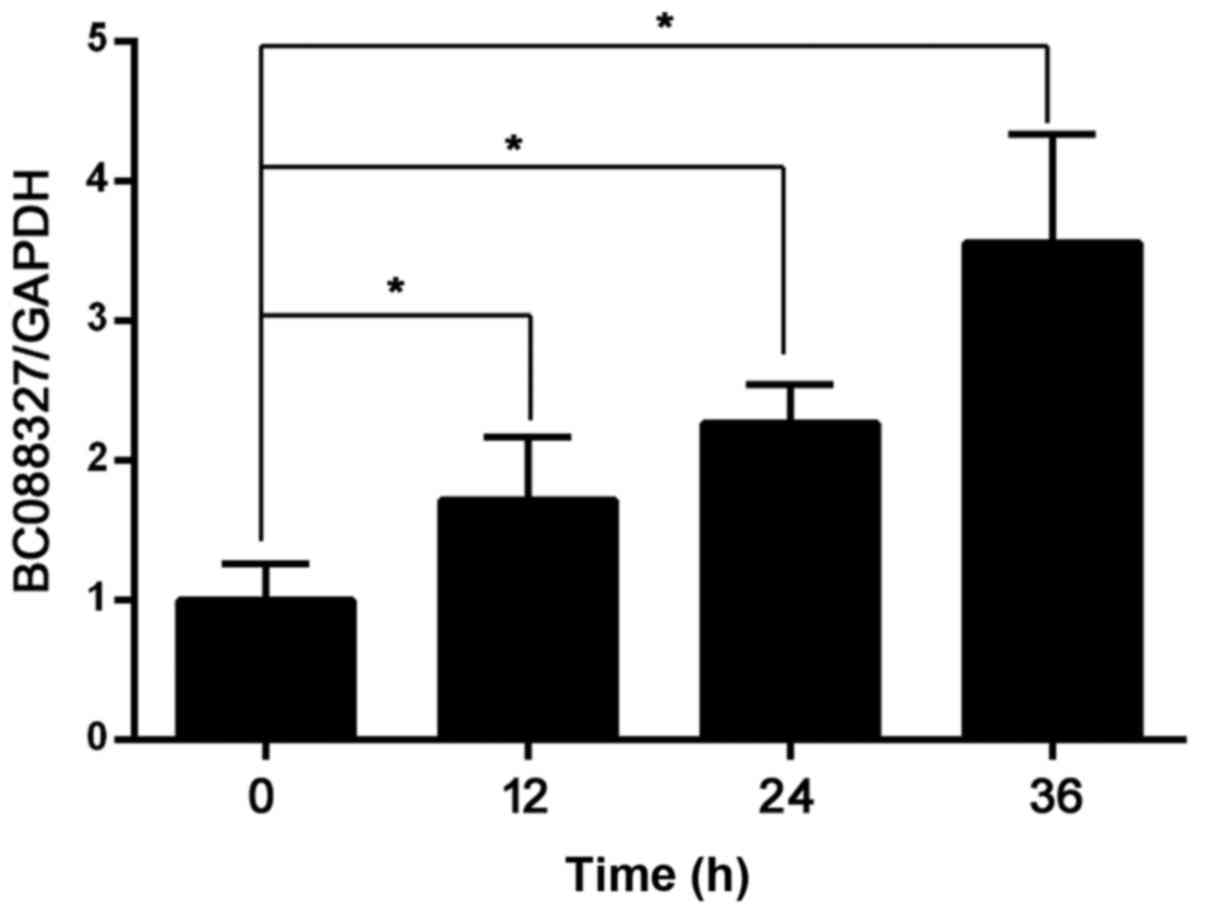
expressed lncRNAs, a hypoxia-induced SC model was induced. The
results of RT-qPCR showed an increased expression level of lncRNA
BC088327 with hypoxic exposure time in the presence of heregulin-1β
(Fig. 3), while other lncRNAs did
not response to hypoxia in the presence of heregulin-1β (data not
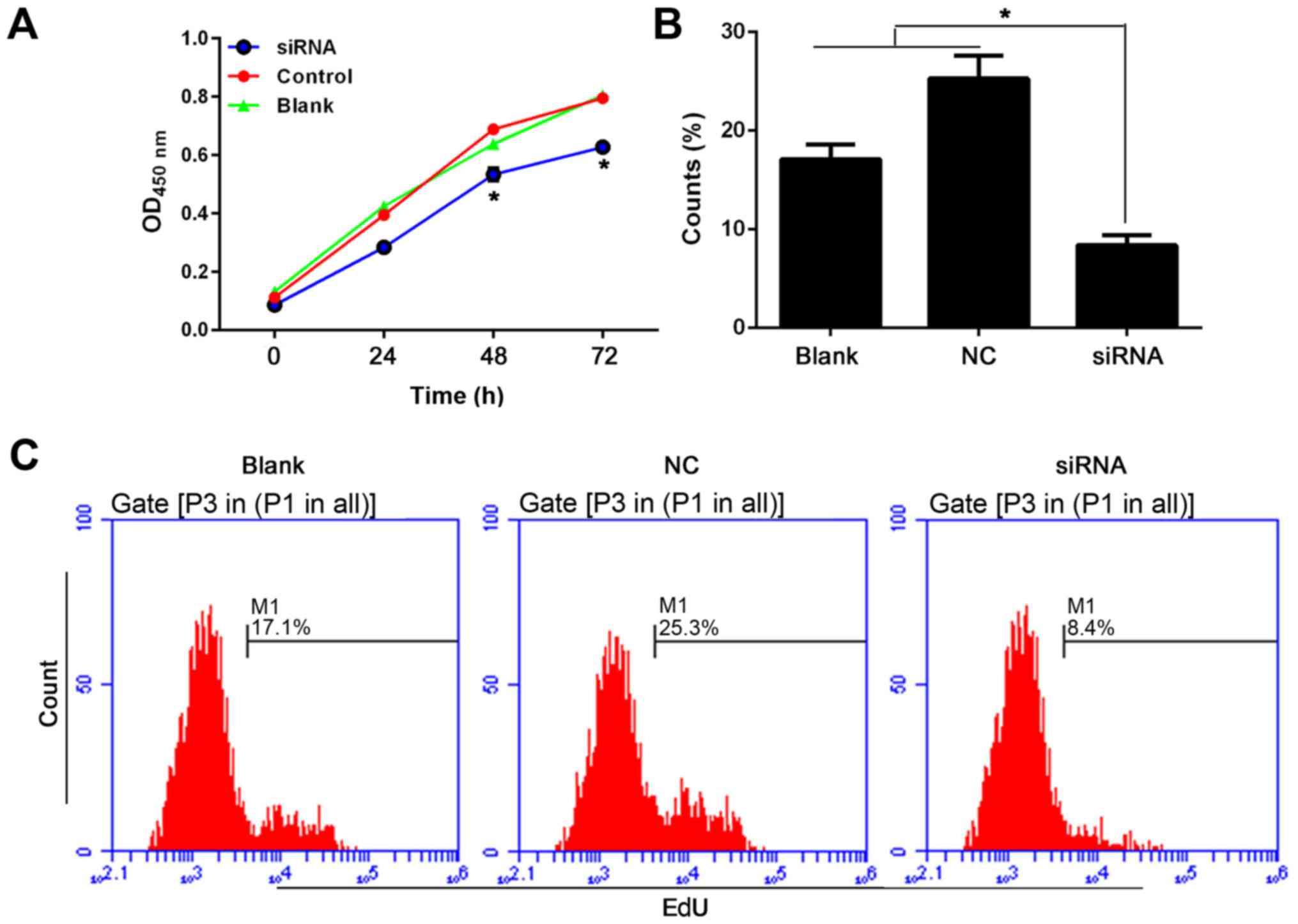
shown). For detecting the role of BC088327 in cell viability, siRNA
that targeted BC088327 was designed and transfected into the SCs,
and then the CCK-8 stain assay was performed (Fig. 4A). Together with EdU staining,
these results showed that transfection with siRNA significantly
suppressed the cell viability, indicating that upregulation of
BC088327 may be associated with the activation of cellular
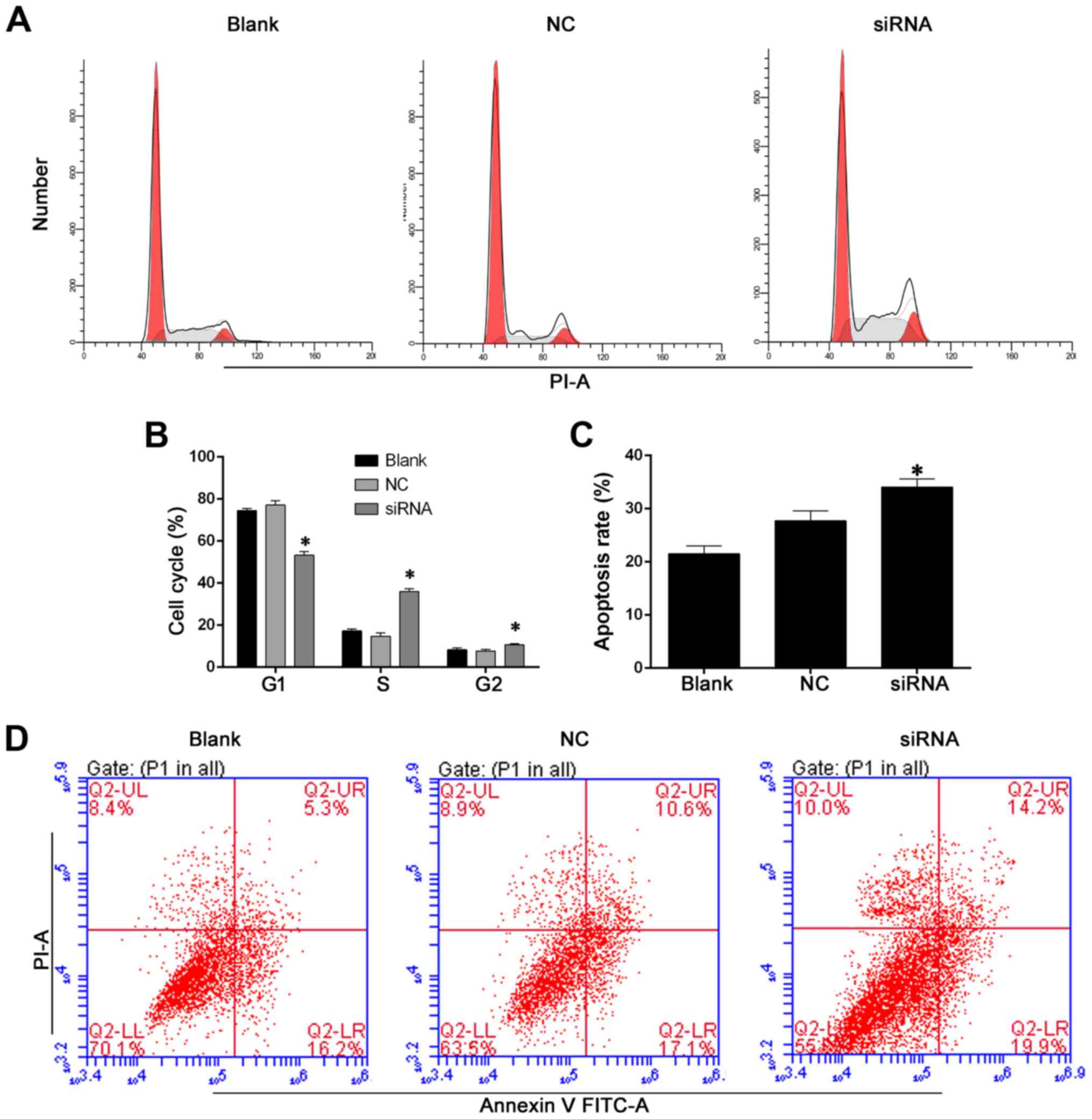
proliferation in the presence of heregulin-1β (Fig. 4B and C). Cell cycle analysis was
also performed using a flow cytometry assay (Fig. 5A and B). Consistent with the CCK-8
assay, knockdown of BC088327 increased the cell proportion
distributed in the S and G2/M phases in hypoxic SCs in the presence
of heregulin-1β. Moreover, the apoptotic rate of cells transfected
with BC088327 siRNA were significant augmented compared with that
in the blank and NC groups (Fig. 5C
and D) (P<0.05). Taken together, these results suggested
that the function of BC088327 was likely to be synergic with
heregulin-1β, which promoted peripheral nerve repair through
induction of cell proliferation.
Discussion
As molecules that are involved in various processes
associated with diseases, oncogenesis and metabolism (21), lncRNAs have received increasing
attention over the past decade. However, even with the emerging
focus on lncRNAs, the mechanisms behind their functions remain
poorly explored (22,23). Currently, information on the
function of lncRNAs mainly focuses on their roles in the regulation
of gene expression at multiple levels, including epigenetic
modification, and transcriptional and post-transcriptional
regulation (24,25). The involvement of lncRNAs in the
regulation of peripheral nerve injury and the repair process, and
the role of lncRNAs in the nervous system have been studied,
however, the molecular mechanism was not clear (26,27).
In the present study, multiple lncRNAs were shown to
be differentially expressed in rats with peripheral nerve injuries
that were treated with ANA plus SKPs-SCs and heregulin-1β when
compared with the control group (ANA plus SKPs-SCs). Among all the
differentially expressed lncRNAs, BC088327 was upregulated to the
largest extent, and knockdown of the lncRNA induced cell apoptosis
and cell cycle arrest in hypoxic SCs with the presence of
heregulin-1β. It is well known that heregulin-1β is a member of the
neuregulin family, which are polypeptide ligands of ErbB tyrosine
kinase receptors. Members of this family serve essential roles in
the transmission of growth and differentiation signals in neuronal
development (11). Since soluble
heregulin-1β can specifically activate mitogen-activated protein
kinase and protein kinase B pathways, it is implied to regulate the
development, growth and repair of the tissues (28–30). In our previous study (12), it was demonstrated that
heregulin-1β promotes the outcome of reinnervation by ANA+SKP-SC
therapy. In the present study, it was found that lncRNA BC088327
was upregulated in the SKPs-SC plus ANA+heregulin-1β group.
Moreover, in hypoxic SCs, silencing of BC088327 is likely to play a
synergic role with heregulin-1β, as knockdown of BC088327
suppressed the cell proliferation through the induction of cell
cycle arrest and cell apoptosis. However, the underlying mechanism
involved in the function of BC088327 may be complicated. Based on a
microarray assay of the mRNA profile, in the experimental group,
the cytokines involving inflammatory signal molecules, including
interleukin (IL)-1β, tumor necrosis factor-α and IL-6, were
increased (all fold-changes >2). All these cytokines can
contribute to the activation of downstream cell apoptosis (21,31,32). Nevertheless, solid evidence in the
present study also showed that extracellular signal-regulated
kinase (ERK) and c-Myc were upregulated in the experimental group
(all fold-changes >2). Contrary to the effect of cytokines,
c-Myc is an important mediator of the cell cycle, and increased ERK
and c-Myc levels will lead to cell proliferation and
differentiation that is consistent with the functions of
heregulin-1β (33,34). Although the present results
indicated a synergic interaction between BC088327 and heregulin-1β,
the exact mechanism through which BC088327 exerted its protective
function on peripheral nerves requires further comprehensive
investigation.
With the exception of BC088327, certain other
differentially expressed lncRNAs may also be associated with
neuronal repair, including lncRNAs that have been previously
reported to be associated with cell proliferation or
differentiation in other models (35,36). For example, the upregulated lncRNA
uc.173 induced by heregulin-1β is able to inhibit the neuronal
apoptosis in the hippocampus tissue model (37). In the study by Yao et al,
it was indicated that silencing of uc.217 expression could
significantly promote neurite outgrowth in cultured dorsal root
ganglion neurons (38). However,
the functions of the majority of the differentially expressed
lncRNAs detected in the present study remain unknown. Further
studies on these lncRNAs and their downstream effectors are
required to unveil the structure of the regulation network of
lncRNAs.
In conclusion, the differentially expressed lncRNA
BC088327 serves an important role in regulating cell proliferation
during the process of nerve repair. Considering its synergic action
with hegerulin-1β, lncRNA BC088327 can be either employed as a
biomarker for detecting the extent of the nerve injury or as a
novel therapeutic target for promotion of nerve repair. For such
purposes, a better understanding of the functions of lncRNAs, such
as the regulatory network and mechanism involved in their effect,
should be further expanded upon in the future.
Acknowledgments
This study was supported by funds from the Shanghai
Municipal Commission of Health and Family Planning (grant no.
2014-399).
Abbreviations:
|
lncRNAs
|
long non-coding RNAs
|
|
SCs
|
Schwann cells
|
|
SKPs
|
Schwann cells isolated from
skin-derived precursors
|
|
ANA
|
acellular nerve allograft
|
Notes
[1] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Cattin AL and Lloyd AC: The multicellular
complexity of peripheral nerve regeneration. Curr Opin Neurobiol.
39:38–46. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Trehan SK, Model Z and Lee SK: Nerve
repair and nerve grafting. Hand Clin. 32:119–125. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Giger RJ, Hollis ER II and Tuszynski MH:
Guidance molecules in axon regeneration. Cold Spring Harb Perspect
Biol. 2:a0018672010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guest J, Santamaria AJ and Benavides FD:
Clinical translation of autologous Schwann cell transplantation for
the treatment of spinal cord injury. Curr Opin Organ Transplant.
18:682–689. 2013.PubMed/NCBI
|
|
5
|
Tabakow P, Raisman G, Fortuna W, Czyz M,
Huber J, Li D, Szewczyk P, Okurowski S, Miedzybrodzki R, Czapiga B,
et al: Functional regeneration of supraspinal connections in a
patient with transected spinal cord following transplantation of
bulbar olfactory ensheathing cells with peripheral nerve bridging.
Cell Transplant. 23:1631–1655. 2014. View Article : Google Scholar
|
|
6
|
Walsh SK, Kumar R, Grochmal JK, Kemp SW,
Forden J and Midha R: Fate of stem cell transplants in peripheral
nerves. Stem Cell Res (Amst). 8:226–238. 2012. View Article : Google Scholar
|
|
7
|
Guertin AD, Zhang DP, Mak KS, Alberta JA
and Kim HA: Microanatomy of axon/glial signaling during Wallerian
degeneration. J Neurosci. 25:3478–3487. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Walsh S and Midha R: Practical
considerations concerning the use of stem cells for peripheral
nerve repair. Neurosurg Focus. 26:E22009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu GB, Cheng YX, Feng YK, Pang CJ, Li Q,
Wang Y, Jia H and Tong XJ: Adipose-derived stem cells promote
peripheral nerve repair. Arch Med Sci. 7:592–596. 2011. View Article : Google Scholar
|
|
10
|
Khuong HT, Kumar R, Senjaya F, Grochmal J,
Ivanovic A, Shakhbazau A, Forden J, Webb A, Biernaskie J and Midha
R: Skin derived precursor Schwann cells improve behavioral recovery
for acute and delayed nerve repair. Exp Neurol. 254:168–179. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mei L and Xiong WC: Neuregulin 1 in neural
development, synaptic plasticity and schizophrenia. Nat Rev
Neurosci. 9:437–452. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang H, Wu J, Zhang X, Ding L and Zeng Q:
Study of synergistic role of allogenic skin-derived precursor
differentiated Schwann cells and heregulin-1β in nerve regeneration
with an acellular nerve allograft. Neurochem Int. 97:146–153. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Verrier JD, Semple-Rowland S, Madorsky I,
Papin JE and Notterpek L: Reduction of Dicer impairs Schwann cell
differentiation and myelination. J Neurosci Res. 88:2558–2568.
2010.PubMed/NCBI
|
|
14
|
Arthur-Farraj PJ, Morgan CC, Adamowicz M,
Gomez-Sanchez JA, Fazal SV, Beucher A, Razzaghi B, Mirsky R, Jessen
KR and Aitman TJ: Changes in the coding and non-coding
transcriptome and DNA methylome that define the schwann cell repair
phenotype after nerve injury. Cell Rep. 20:2719–2734. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pan B, Zhou HX, Liu Y, Yan JY, Wang Y, Yao
X, Deng YQ, Chen SY, Lu L, Wei ZJ, et al: Time-dependent
differential expression of long non-coding RNAs following
peripheral nerve injury. Int J Mol Med. 39:1381–1392. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou S, Ding F and Gu X: Non-coding RNAs
as emerging regulators of neural injury responses and regeneration.
Neurosci Bull. 32:253–264. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schmitt AM and Chang HY: Long noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rühle F and Stoll M: Long non-coding RNA
databases in cardiovascular research. Genomics Proteomics
Bioinformatics. 14:191–199. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Toma JG, Akhavan M, Fernandes KJ,
Barnabé-Heider F, Sadikot A, Kaplan DR and Miller FD: Isolation of
multipotent adult stem cells from the dermis of mammalian skin. Nat
Cell Biol. 3:778–784. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Method. 25:402–408. 2001.
View Article : Google Scholar
|
|
21
|
Takayama K and Inoue S: The emerging role
of noncoding RNA in prostate cancer progression and its implication
on diagnosis and treatment. Brief Funct Genomics. 15:257–265. 2016.
View Article : Google Scholar
|
|
22
|
Geisler S and Coller J: RNA in unexpected
places: long non-coding RNA functions in diverse cellular contexts.
Nat Rev Mol Cell Biol. 14:699–712. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang L, Froberg JE and Lee JT: Long
noncoding RNAs: fresh perspectives into the RNA world. Trends
Biochem Sci. 39:35–43. 2014. View Article : Google Scholar :
|
|
24
|
Wilusz JE, Sunwoo H and Spector DL: Long
noncoding RNAs: functional surprises from the RNA world. Genes Dev.
23:1494–1504. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Taft RJ, Pang KC, Mercer TR, Dinger M and
Mattick JS: Non-coding RNAs: regulators of disease. J Pathol.
220:126–139. 2010. View Article : Google Scholar
|
|
26
|
Knauss JL and Sun T: Regulatory mechanisms
of long noncoding RNAs in vertebrate central nervous system
development and function. Neuroscience. 235:200–214. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ng SY, Bogu GK, Soh BS and Stanton LW: The
long noncoding RNA RMST interacts with SOX2 to regulate
neurogenesis. Mol Cell. 51:349–359. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang SH, Sharrocks AD and Whitmarsh AJ:
MAP kinase signalling cascades and transcriptional regulation.
Gene. 513:1–13. 2013. View Article : Google Scholar
|
|
29
|
Gu Y, Xue C, Zhu J, Sun H, Ding F, Cao Z
and Gu X: Basic fibroblast growth factor (bFGF) facilitates
differentiation of adult dorsal root ganglia-derived neural stem
cells toward Schwann cells by binding to FGFR-1 through MAPK/ERK
activation. J Mol Neurosci. 52:538–551. 2014. View Article : Google Scholar
|
|
30
|
Awasthi S and Hamburger AW: Heregulin
negatively regulates transcription of ErbB2/3 receptors via an
AKT-mediated pathway. J Cell Physiol. 229:1831–1841. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gierut JJ, Wood LB, Lau KS, Lin YJ,
Genetti C, Samatar AA, Lauffenburger DA and Haigis KM:
Network-level effects of kinase inhibitors modulate TNF-α-induced
apoptosis in the intestinal epithelium. Sci Signal. 8:ra1292015.
View Article : Google Scholar
|
|
32
|
Gurung P and Kanneganti TD: Novel roles
for caspase-8 in IL-1β and inflammasome regulation. Am J Pathol.
185:17–25. 2015. View Article : Google Scholar :
|
|
33
|
Bueno MJ and Malumbres M: MicroRNAs and
the cell cycle. Biochim Biophys Acta. 1812:592–601. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen Y, Lin C, Liu Y and Jiang Y: HMGB1
promotes HCC progression partly by downregulating p21 via ERK/c-Myc
pathway and upregulating MMP-2. Tumour Biol. 37:4399–4408. 2016.
View Article : Google Scholar :
|
|
35
|
Chen SX, Yin JF, Lin BC, Su HF, Zheng Z,
Xie CY and Fei ZH: Upregulated expression of long noncoding RNA
SNHG15 promotes cell proliferation and invasion through regulates
MMP2/MMP9 in patients with GC. Tumour Biol. 37:6801–6812. 2016.
View Article : Google Scholar
|
|
36
|
Zhang J, Yao T, Wang Y, Yu J, Liu Y and
Lin Z: Long noncoding RNA MEG3 is downregulated in cervical cancer
and affects cell proliferation and apoptosis by regulating miR-21.
Cancer Biol Ther. 17:104–113. 2016. View Article : Google Scholar :
|
|
37
|
Nan A, Zhou X, Chen L, Liu M, Zhang N,
Zhang L, Luo Y, Liu Z, Dai L and Jiang Y: A transcribed
ultraconserved noncoding RNA, Uc.173, is a key molecule for the
inhibition of lead-induced neuronal apoptosis. Oncotarget.
7:112–124. 2016. View Article : Google Scholar :
|
|
38
|
Yao C, Wang J, Zhang H, Zhou S, Qian T,
Ding F, Gu X and Yu B: Long non-coding RNA uc.217 regulates neurite
outgrowth in dorsal root ganglion neurons following peripheral
nerve injury. Eur J Neurosci. 42:1718–1725. 2015. View Article : Google Scholar
|