Introduction
Proliferative vitreoretinopathy (PVR) is a
complication of retinal detachment (RD) and is the primary cause of
surgical failure following RD treatment (1). PVR is characterized by the formation
of fibrotic tissue on the detached retina, which hinders retinal
reattachment and can potentially cause blindness (2). Surgical removal of the fibrotic
membranes by retinal repair is the primary treatment for PVR. The
pathology of the fibrotic membrane reflects an excessive wound
healing reaction characterized by cell proliferation, migration,
extracellular matrix (ECM) production and remodeling (3). For these reasons, the identification
of novel treatments to improve the available therapeutic options is
essential for managing this disease.
Retinal pigment epithelial (RPE) cells are a major
component of the fibrotic membrane; these cells transform into
fibroblast-like cells through epithelial-to-mesenchymal transition
(EMT) (4). Under normal, healthy
conditions, RPE cells form a monolayer comprising the blood-retina
barrier between the retina and choroid (5). During EMT, RPE cells lose their
epithelial characteristics, including unique cell-to-cell contact,
and obtain mesenchymal properties (6). In addition, the RPE cells
proliferate, migrate towards the vitreous body or intra-retinal
layers, and produce ECM, leading to the formation of fibrotic
membranes (7). However, the
specific molecular mechanisms mediating these processes remain to
be fully elucidated, and an effective therapeutic approach
targeting RPE cells has not been developed.
NADPH oxidases (NOXs) are a family of
heme-containing proteins, the main function of which is the
production of reactive oxygen species. A total of seven NOX
proteins have been identified, including NOX1-5 and DUOX1-2. These
enzymes are widely expressed in numerous tissues and have various
functions, including roles in cell signaling, regulation of gene
expression, cell death, cell differentiation, and growth (8). The expression and activity of these
enzymes are regulated by growth factors, including angiotensin II,
platelet-derived growth factor and transforming growth factor
(TGF)-β (9). Among these, TGF-β
initiates the upregulation of other PVR-inducing factors. The major
TGF-β isoform in the posterior segment of the eye, TGF-β2, is
overexpressed in the vitreous and proliferative membranes of
patients with PVR (10,11).
A previous study showed that RPE cells express NOX1,
NOX2 and NOX4 under normal physiological conditions (12). NOX4 is a 578-amino acid
flavocytochrome protein with six transmembrane domains, which has
been shown to modulate epithelial cell cytoskeletal alterations,
smooth muscle cell differentiation, pulmonary fibroblast
proliferation, and liver fibrosis (13-16). NOXs are involved in the
development and progression of a wide spectrum of diseases and
represent a significant therapeutic target. However, the majority
of NOX inhibitors have not demonstrated sufficient clinical
efficacy. For example, apocynin affects other events, including the
induction of transcription factor activator protein-1 (17) and the formation of thromboxane A2,
which is independent of NOXs (18). In addition, diphenyleneiodonium is
not specific for NOXs as it inhibits all flavoenzymes, including
xanthine oxidase and cytochrome P450 (19).
Accordingly, the present study aimed to investigate
the effects of a novel NOX inhibitor, VAS2870, which is the only
confirmed low-molecular-weight pharmacological inhibitor of NOX, on
the TGF-β-dependent expression of NOX4, migration, EMT and cell
proliferation in RPE cells. The findings may provide important
insights into the potential application of NOX inhibitors for the
treatment and prevention of PVR.
Materials and methods
RPE cell culture and treatment
The ARPE-19 human RPE cells were obtained from the
American Type Culture Collection (Manassas, VA, USA). The ARPE-19
line is an immortalized cell line, which spontaneously arose from
cultures of human RPE. The cells were cultured in Dulbecco's
modified Eagle's medium (DMEM)/F12 containing 10% fetal bovine
serum (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA).
The cells were cultured to 70% confluence at 37°C in a humidified
atmosphere containing 5% CO2, and the medium was
replaced every 2-3 days. Subsequently, the cells were disaggregated
with 0.25% trypsin-0.02% ethylenediaminetetraacetic acid solution
and were passaged every 3-5 days. The cells were randomly divided
into five groups: Control, 5 ng/ml TGF-β2, TGF-β2+1 µM/ml
VAS2870, TGF-β2+3 µM/ml VAS2870, and TGF-β2+5 µM/ml
VAS2870. TGF-β2 and VAS2870 were purchased from Sigma-Aldrich;
Merck KGaA (Darmstadt, Germany).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was isolated using an RNA extraction kit
(Fastagen Biotech, Shanghai, China) and reverse transcribed into
complementary DNA (cDNA) using a PrimerScript First Strand cDNA
Synthesis kit (Takara Biotechnology Co., Ltd., Shiga, Japan)
according to the manufacturer's protocol. The quality of the RNA
samples was controlled by measuring the absorbance at A260/280; an
absorption between 1.8 and 2.1 indicated that the RNA was of high
quality. For PCR amplification, a MyTaq Red DNA Polymerase
(Bioline, London, England) kit was used according to the
manufacturer's protocol (10 µl 5xMyTaq Red Reaction buffer,
forward and reverse primers (20 µM), each 1 µl, 1
µl MyTaq Red DNA polymerase, 10 ng cDNA and up to 50
µl ddH2O). The primers used for detection of mRNA
expression of NOX1-4 and GAPDH were based on
previously published sequences (12). For quantitative detection of
NOX1-4 mRNAs, the following sequence-specific primers were
used: NOX1, forward 5′-TTC ACC AAT TCC CAG GAT TGA AGT GGA
TGG TC-3′ and reverse 5′-GAC CTG TCA CGA TTG CAT GGC CTT GTC AA-3′;
NOX2, forward 5′-AGA GGG TTG GAG GTG GAG AAT T-3′ and
reverse 5′-GCA CAA GGA GCA GGA CTA GAT GA-3′; NOX4, forward
5′-CTG GAG GAG CTG GCT CGC CAA CGA AG-3′ and reverse 5′-GTG ATC ATG
AGG AAT AGC ACC ACC ACC ATG CAG-3′; GAPDH, forward 5′-UUC
UCC GAA CGU GUC ACG UTT-3′ and reverse 5′-ACG UGA CAC GUU CGG AGA
ATT-3′. The PCR was performed as follows: 30 sec at 95°C, followed
by 40 cycles of 5 sec at 95°C, 20 sec at 60°C and 20 sec at 72°C.
The amplified DNA was visualized with SYBR Safe DNA Gel staining
(diluted to 1:10,000; Invitrogen; Thermo Fisher Scientific, Inc.)
on 3% agarose TAE gels run at 100 V for 30 min. Positive control
tissues for each NOX homolog were also examined (human NOX1, NOX2
and NOX4). It was not possible to validate the expression of NOX3
as it is predominantly expressed in the inner ear. The gene
expression levels were normalized to the endogenous control GAPDH,
and fold changes were calculated using the 2-ΔΔCq method
(20). All PCR reactions were
performed at least in triplicate.
Cell Counting Kit-8 (CCK-8) assays
Cell viability was examined using CCK-8 assays (7Sea
Pharmatech, Co., Ltd., Shanghai, China). VAS2870 (Sigma-Aldrich;
Merck KGaA) was diluted in dimethylsulfoxide (DMSO; Sigma-Aldrich;
Merck KGaA) to different concentrations. The RPE cells were plated
in 96-well plates (5×103 cells/well) with 100-µl
complete culture medium. The cells were then treated with 0.1, 0.5,
1, 5 or 10 µM/ml VAS2870 (dissolved in DMSO). After 24, 48
or 72 h, 10 µl CCK-8 solution was added to each well, and
the plates were incubated for 2 h at 37°C. The absorbance was
determined at 450 nm with a microplate reader (Epoch; Bio-Tek
Instruments Inc., Winooski, VT, USA).
Wound healing assays
The RPE cells were seeded in 6-well cell culture
plates at a concentration of 5×105 cells/well, grown to
almost 100% confluence as a monolayer and then incubated with
serum-free DMEM. Wounds were gently introduced in the center of the
well using a sterile 200-µl pipette tip, following which the
plates were washed with phosphate-buffered saline (PBS) to remove
cell debris. TGF-β2 (5 ng/ml) and VA2870 (5 µM) were then
added to the cells. Images of marked regions along the wound region
were captured using an inverted phase contrast microscope (Leica
DMI3000B; Leica Microsystems GmbH, Asslar, Germany) immediately
following creation of the wound. The distance migrated by the RPE
cells into the wounded region was determined by collecting images
every 24 h.
Cell cycle analysis
The RPE cells were seeded in 25-cm2
culture flasks at a density of 1×106 cells/flask. The
cells were starved overnight and divided into four groups (control
group, and 1, 3, and 5 µM VAS2870 groups). Following
incubation at 37°C for 24 h, the cells were collected and fixed in
70% ice-cold ethanol at 4°C overnight. The cells were then washed
twice with PBS, resuspended in 500-µl PBS containing 100
µg/ml propidium iodide (Sigma-Aldrich; Merck KGaA) and 100
µg/ml RNase A (Sigma-Aldrich; Merck KGaA), and incubated in
the dark for 20-40 min. Finally, cell cycle distribution was
detected by flow cytometry at -24°C, and the cells were analyzed
using a FACSCalibur with FACSort CellQuest v6.0 software (BD
Biosciences, Franklin Lakes, NJ, USA).
Immunofluorescence staining
The cells grown on 8×8 mm2
poly-L-lysine-coated coverslips in 24-well plates were starved
overnight and then treated with 5 ng/ml TGF-β2 and 5 µM
VAS2870. After 24 h, the cells were fixed with 4% paraformaldehyde
for 30 min and washed three times with PBS. The cells were then
permeabilized with 0.3% Triton X-100 for 20 min at room
temperature, and slides were blocked in goat serum (Boster
Biological Technology, Pleasanton, CA, USA) for 1 h, followed by
overnight incubation at 4°C with the following primary antibodies:
Monoclonal rabbit anti-human α-smooth muscle actin (SMA; 1:200;
cat. no. ab124964; Abcam, Cambridge, MA, USA) and monoclonal rabbit
anti-human E-cadherin (1:200; cat. no. ab53226; Abcam). Following
washing with PBS extensively, the cells were exposed to the
fluorescent secondary antibodies, goat anti-rabbit Cy3 (1:10,000;
cat. no. CW0114S) and goat anti-rabbit FITC (1:10,000; cat. no.
CW0159S) from CWBio (Beijing, China), in the dark for 30 min. The
coverslips were washed again, mounted with DAPI for 5 min, and
observed under a fluorescence microscope (DP71; Olympus, Tokyo,
Japan).
Western blot analysis
The cells were lysed in RIPA buffer (Sigma-Aldrich;
Merck KGaA) supplemented with 1% protease inhibitor cocktail (Roche
Diagnostics, Basel, Switzerland). The proteins (20 µg per
lane) were separated using sodium dodecyl sulfate polyacrylamide
gel electrophoresis (8-10% gel) and transferred onto polyvinylidene
difluoride membranes. Protein concentration was determined using
the BCA kit (Beyotime Institute of Biotechnology, Haimen, China).
The membranes were blocked in tris-buffered saline with 1% Tween-20
(TBST) containing 8% skim milk at room temperature for 2 h, and
incubated with the following primary antibodies: Monoclonal rabbit
anti-human cyclin D1 (1:20,000; cat. no. ab134175), monoclonal
anti-E-cadherin (1:1,000; cat. no. ab40772), monoclonal anti-NOX4
(1:1,000; cat. no. ab32419) and monoclonal anti-α-SMA (1:1,000;
cat. no. ab124964), all from Abcam, in a sealed bag overnight at
4°C and the membranes were then washed three times for 10 min each
with TBS. The membranes were then incubated with horseradish
peroxidase (HRP)-conjugated secondary antibodies (1:100,000; goat
polyclonal anti-rabbit secondary antibody (IgG-HRP; cat. no.
ab6721; Abcam) for 2 h at room temperature, washed three times with
TBST for 15 min each, and subjected to chemiluminescence detection
with ECL substrate solution (Genshare Biological, Shaanxi, China)
for 2 min. Subsequently, the membranes were placed in a LAS-3000
FujiFilm intelligent dark box. The illuminated bands were detected,
and images were captured using Image reader LAS-3000 software 8.0
(Fujifilm Holdings Corporation, Tokyo, Japan). Three independent
experiments were performed.
Statistical analysis
All data are presented as the mean ± standard
deviation. Statistical analyses were performed using SPSS version
18.0 for Windows (SPSS, Inc., Chicago, IL, USA). Statistical
comparisons were performed using Student's t-test or two-way
analysis of variance followed by ANOVA analysis (followed by
Tukey's post hoc test) for multiple comparisons. P<0.05 was
considered to indicate a statistically significant difference.
Results
VAS2870 inhibits the expression of NOX4
in TGF-β2-treated RPE cells
The effects of VAS2870 on the expression of NOX4
following TGF-β2-induced EMT in ARPE-19 cells were determined by
western blot analysis. Specifically, the expression of NOX4 was
assessed at different time points (0, 4, 6, 8 and 12 h) following
TGF-β2 (5 ng/ml) stimulation. The expression of NOX4 was initially
induced at 4 h following TGF-β treatment and expression was most
pronounced at 8 h (Fig. 1A). In
addition, in the RPE cells at 8 h, the mRNA levels of NOX4
were highest, compared with the expression of NOX1 and
NOX2 (Fig. 1B). When
treated with different concentrations of TGF-β2, the protein
expression of NOX4 was significantly increased, compared with the
levels in the normal control group (Fig. 1C). In addition, the upregulation
of NOX4 induced by TGF-β2 was significantly inhibited by VAS2870
(Fig. 1D). These results
suggested that VAS2870 sufficiently inhibited the TGF-β- dependent
expression of NOX in a dose-dependent manner.
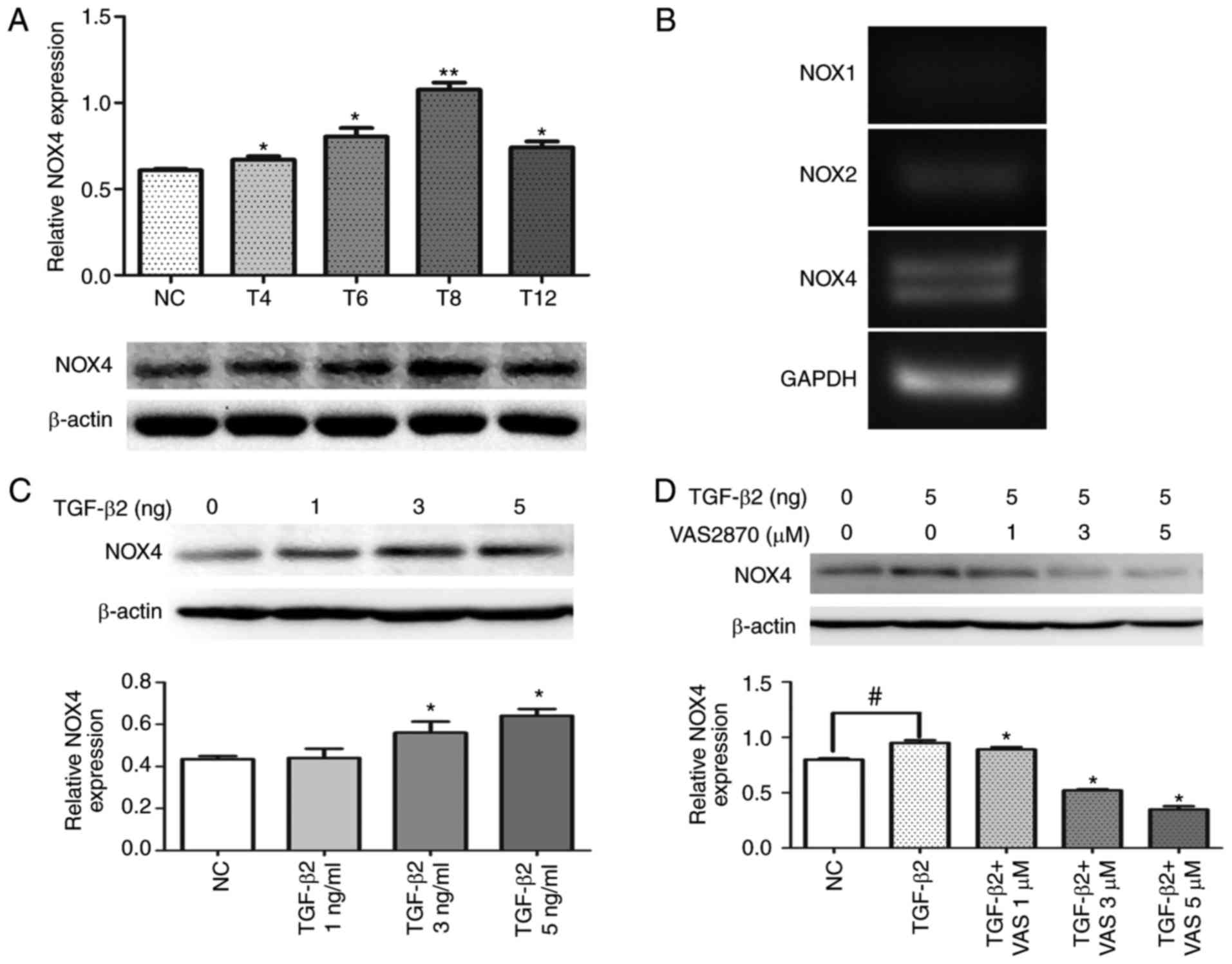 | Figure 1VAS2870 suppresses the TGF-β2-induced
activated expression of NOX4. (A) Representative western blots
depicting the time-course (4, 6, 8 and 12 h) of the expression of
NOX4 following TGF-β2 treatment, compared with that in control
explants. *P<0.05 and **P<0.01, vs. NC
group. (B) Representative results of reverse
transcription-quantitative PCR analysis of NOX1, NOX2
and NOX4 mRNA transcripts after 8 h. For each lane, the PCR
products shown correspond to the expected base pair lengths
(NOX1, 62 bp; NOX2, 45 bp; NOX4, 59 bp;
GAPDH, 42 bp). (C) Changes in the expression of NOX4 in the
presence of 1, 3 and 5 ng/ml TGF-β2 for 24 h.
*P<0.05, vs. NC group. (D) Changes in the expression
of NOX4 in the presence of VAS2870 (1, 3 or 5 µM) with
TGF-β2 (5 ng/ml) for 24 h. *P<0.05, vs. TGF-β group;
#P<0.05, vs. NC group. TGF-β2, transforming growth
factor-β2; NOX4, NADPH oxidase 4; PCR, polymerase chain reaction;
NC, normal control. |
Effects of VAS2870 on RPE cell
viability
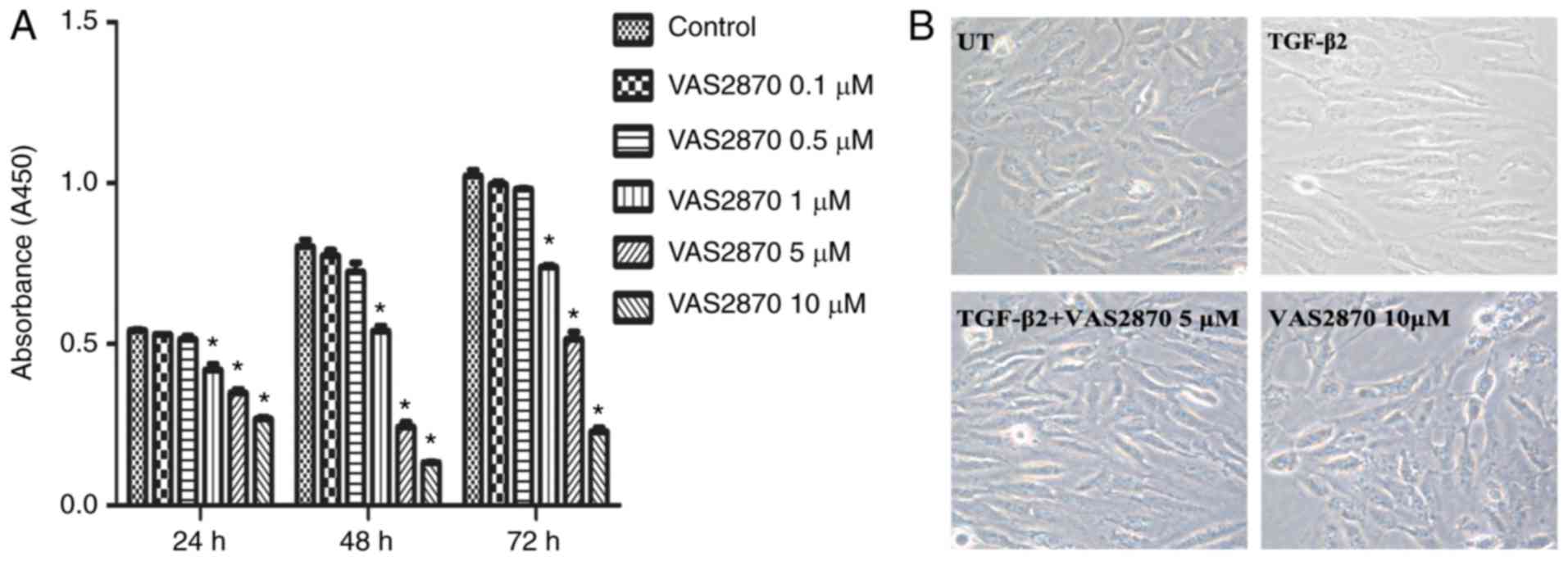
The effects of VAS2870 on RPE cell viability were
examined using different concentrations of VAS2870 for 24, 48 and
72 h. As indicated in Fig. 2,
VAS2870 treatment resulted in significant inhibitory effects on
cell growth in a concentration-dependent manner (Fig. 2A). The viability of cells treated
with 10 µM VAS2870 was significantly lower, compared with
that in cells treated with 1 and 5 µM VAS2870 at all three
time points. To determine whether the growth inhibitory effect was
due to VAS2870-induced cell death, the morphology of cells was
evaluated. The results revealed that cells in the normal group were
spindle-shaped with oval nuclei and visible black particles in the
cytoplasm. By contrast, in the TGF-β2 group, the cells were
elongated, whereas 5 µM VAS2870 did not affect cell
survival. However, treatment with 10 µM VAS2870 for 24 h
resulted in an increased number of dead cells, which had lost their
original spindle shape and in which cytochrome had diffused into
the extracellular matrix (Fig.
2B). These results indicated that the growth inhibitory effects
of low concentrations of VAS2870 (1 and 5 µM) were
predominantly caused by the suppression of cell proliferation,
whereas a higher concentration (10 µM) resulted in cell
death. Therefore, 1 and 5 µM VAS2870 was used for subsequent
experiments.
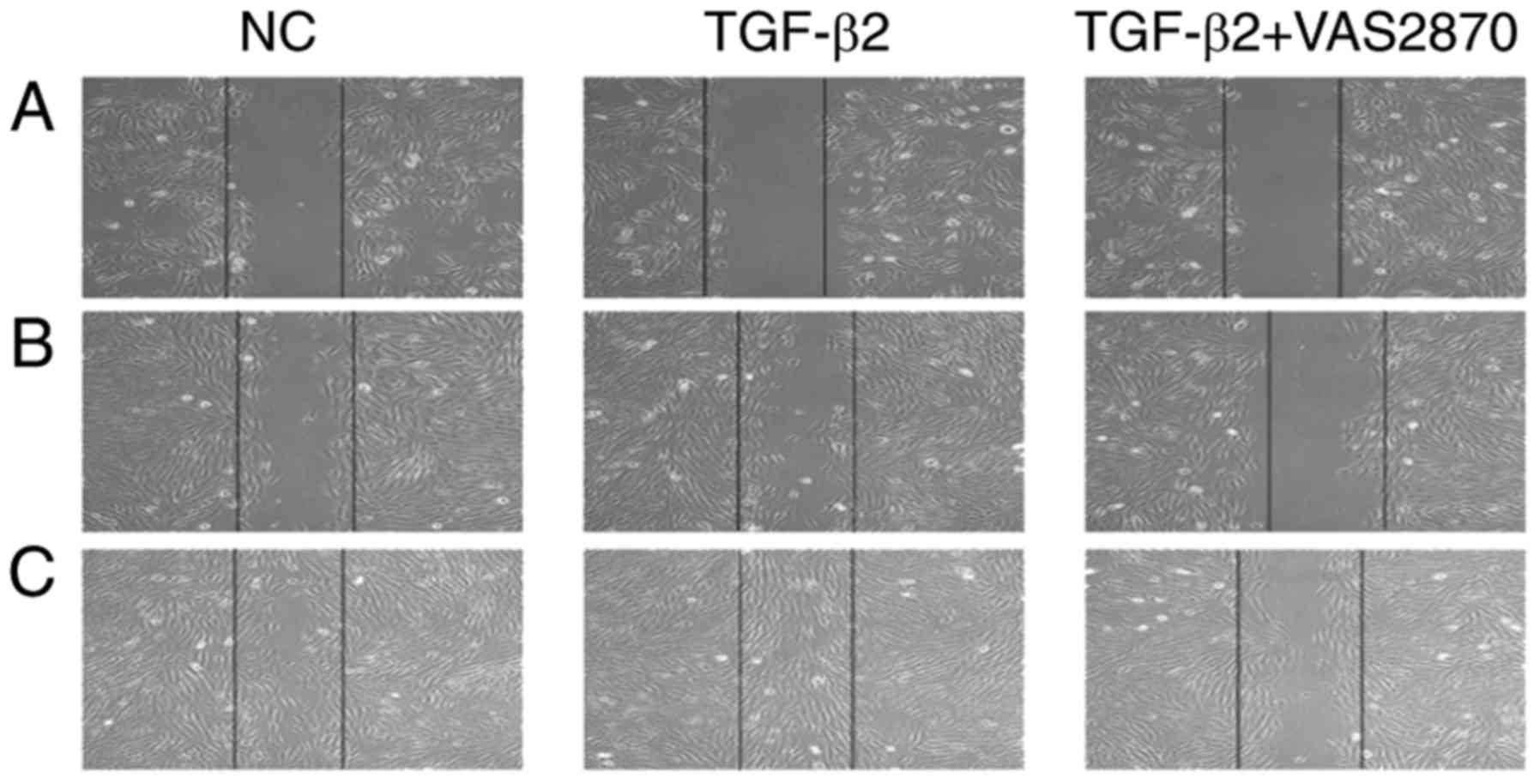
VAS2870 inhibits TGFβ2-induced cell
migration
TGFβ2-mediated cell migration in RPE cells has been
implicated in the development of PVR (21). Therefore, the present study
examined whether VAS2870 suppressed this event. As shown in
Fig. 3A, TGFβ2 significantly
enhanced cell migration, compared with that in the control group.
The distance covered by cells treated with VAS2870 (5 µM)
was shorter than that in the TGF-β2-treated group at 24 h (Fig. 3B). By 48 h, the wound had healed
in the TGFβ2 group; however, at the same time point, a gap there
remained in the VAS2870 group (Fig.
3C). These data demonstrated that pretreatment with VAS2870
inhibit TGF-β-dependent migration at a concentration of 5
µM.
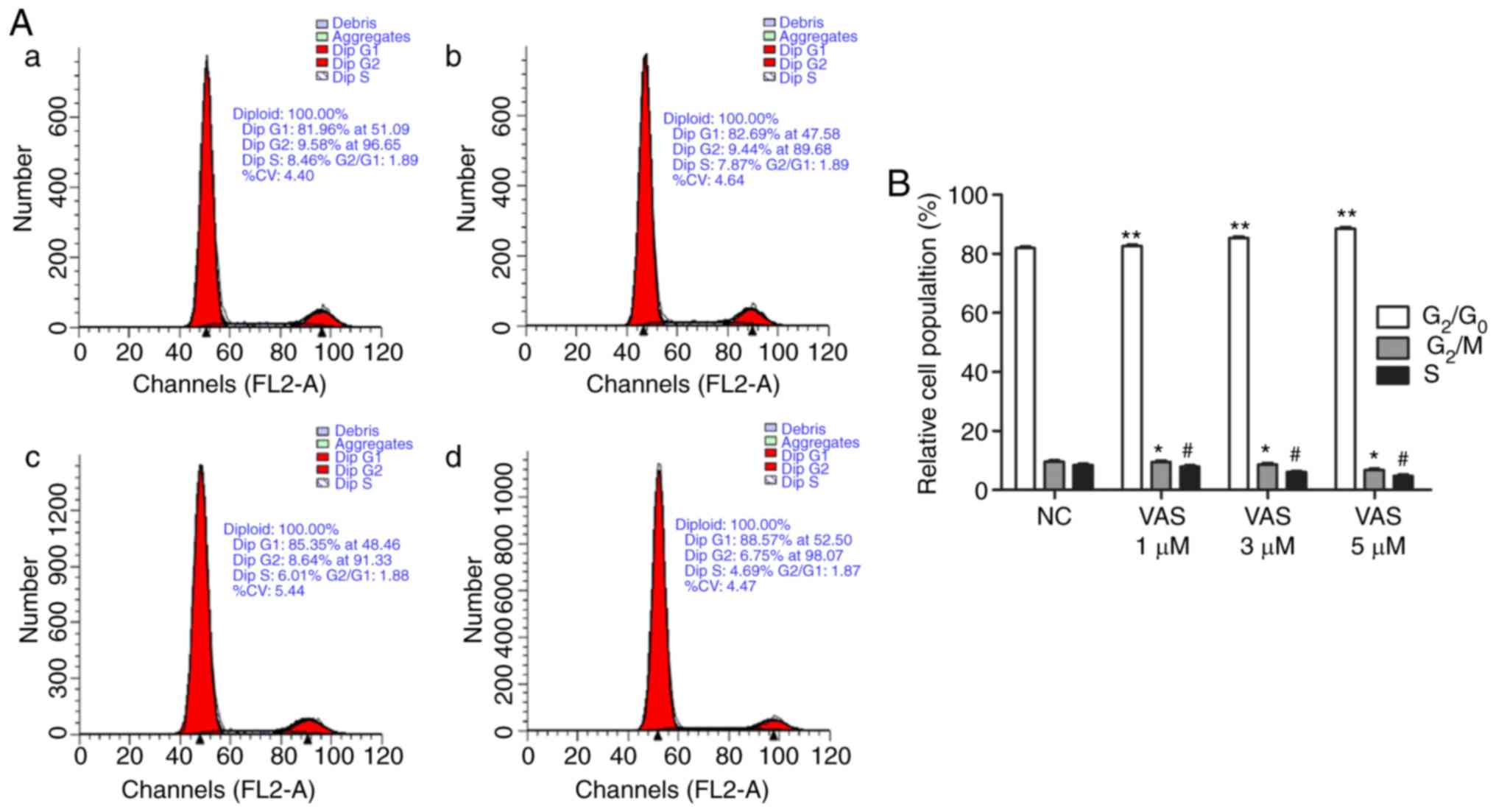
VAS2870 inhibits the proliferation of RPE
cells by inducing G1-phase cell cycle arrest
The present study also evaluated the effects of
VAS2870 on cell cycle arrest (Fig.
4A). Upon culture with different concentrations of VAS2870 (0,
1, 3 and 5 µM) for 24 h, an accumulation of RPE cells in the
G0/G1 phase and a decrease in the number of
cells in the S and G2/M phases were noted compared with
those in the normal control group (Fig. 4B). These findings indicated that
VAS2870 inhibited the proliferation of RPE cells via G1
phase cell cycle arrest.
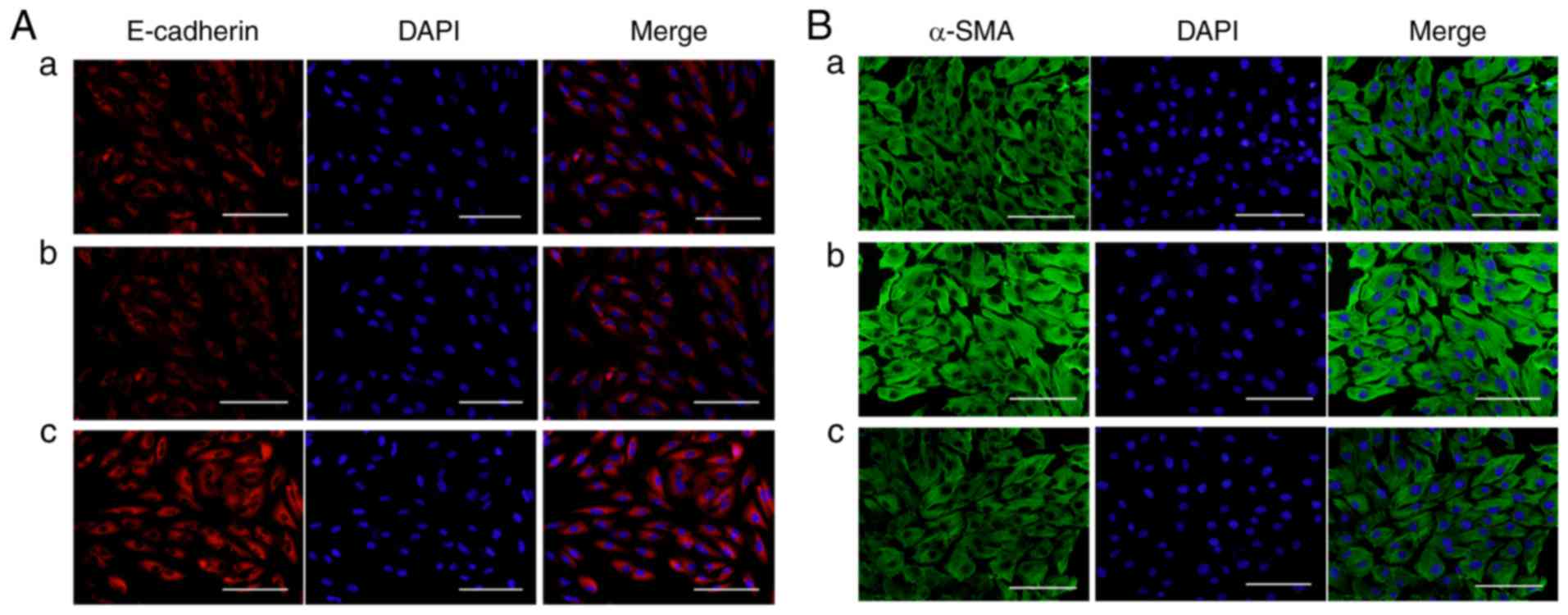
VAS2870 prevents TGF-β2-induced EMT in
RPE cells
To investigate whether VAS2870 affects EMT in RPE
cells, the present study examined the expression of E-cadherin (an
epithelial marker) and α-SMA (a mesenchymal marker) following
treatment with VAS2870. The expression and distribution of
E-cadherin and α-SMA in RPE cells was also detected by
immunofluorescence staining. Under physiological conditions,
E-cadherin was expressed in the cell membrane (Fig. 5Aa) whereas, in the presence of
TGF-β2, staining for E-cadherin in the VAS2870 group was more
marked, compared with that in the TGF-β2 group (Fig. 5Ab and c). Under physiological
conditions, α-SMA was localized to the cytoplasm in RPE cells
(Fig. 5Ba), whereas staining for
α-SMA was diffuse and enhanced in the TGF-β2 group (Fig. 5Bb), compared with that in the
VAS2870 group (Fig. 5Bc).
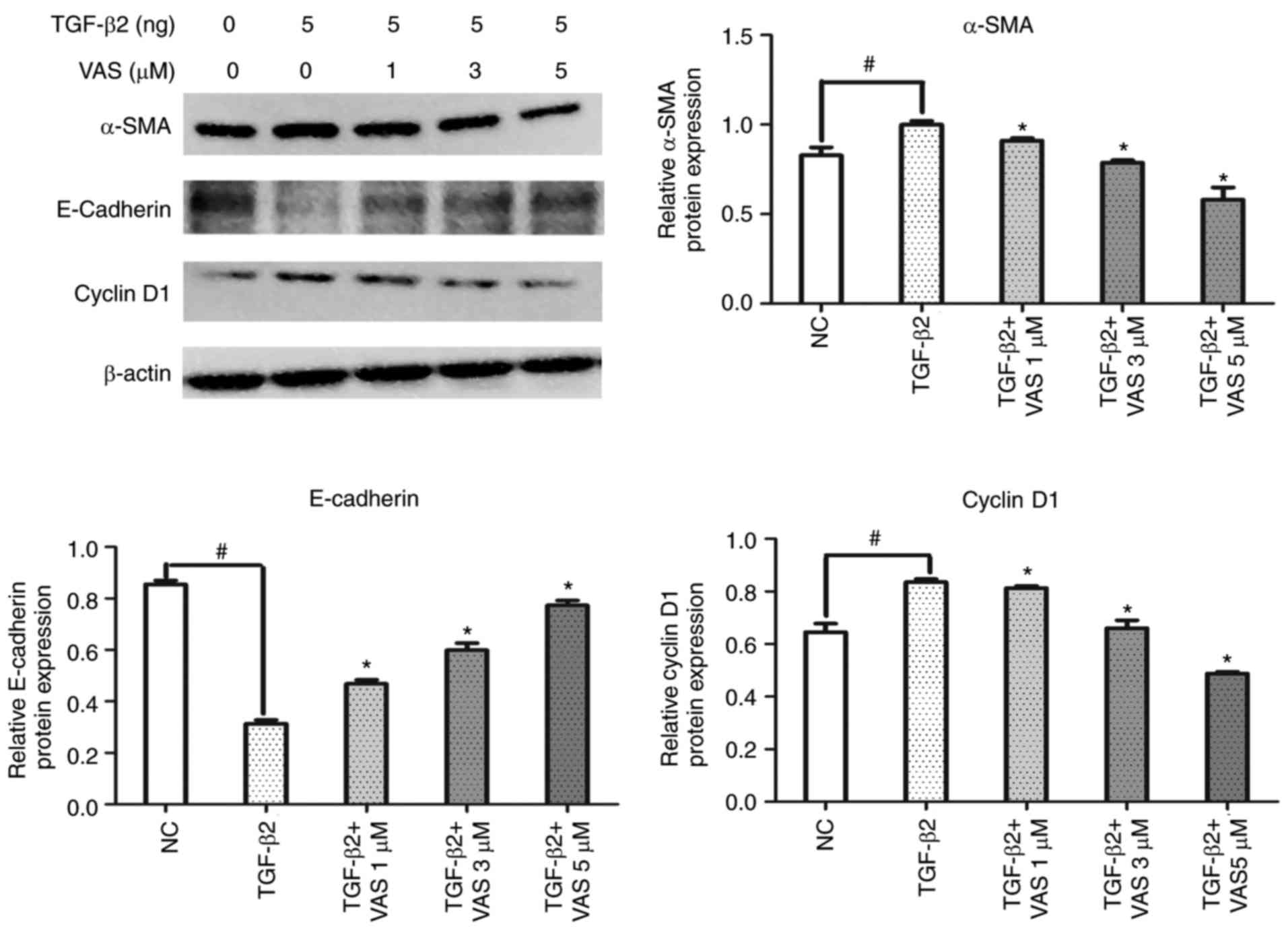
The results of western blot analysis revealed that
the expression of α-SMA was significantly upregulated and that of
E-cadherin was downregulated following treatment with TGF-β2 for 24
h, compared with levels in the control group (P<0.05; Fig. 6), indicating that TGF-β2 promoted
the differentiation of RPE cells. Of note, the differentiation
capacity was reduced by VAS2870; treatment significantly reduced
the expression of α-SMA, compared with that in the TGF-β2 group,
but increased the expression of E-cadherin in a
concentration-dependent manner (P<0.05; Fig. 6). Therefore, these data showed
that the NOX inhibitor VAS2870 significantly attenuated
TGF-β2-induced EMT in RPE cells.
To investigate the mechanism associated with cell
cycle arrest, the present study examined the effects of the
pharmacological inhibition of NOX on the expression of cell
cycle-regulated proteins by western blot analysis. As expected,
VAS2870 significantly decreased the expression of cyclin D1 in a
concentration-dependent manner (P<0.05; Fig. 6). Taken together, these data
suggested that VAS2870 suppressed the proliferation of RPE cells by
inducing G1 phase cell cycle arrest, which occurred
through the inhibition of cyclin D1.
In conclusion, the data obtained demonstrated that
NOXs are critical for TGF-β-dependent EMT and migration in RPE
cells and modulate signaling pathways, which promote proliferation
in these cells.
Discussion
In the present study, the effects of the novel NOX
inhibitor VAS2870 on the expression of NOX4 and the functional
effects mediated by growth factors in RPE cells were determined for
the first time, to the best of our knowledge. It was demonstrated
that the mRNA expression of NOX4 was highest among the NOX
homologs analyzed (NOX1 and NOX2) in RPE cells, and
that the protein expression of NOX4 was increased significantly
upon TGF-β2 stimulation in these cells. It was also shown that the
pan-NOX inhibitor VAS2870 markedly suppressed the TGF-β2-induced
expression of NOX4, and that RPE cells retained an epithelial-like
phenotype following VAS2870 treatment. In addition, VAS2870
suppressed cell proliferation, and TGF-β-dependent migration and
EMT.
The role of NOX as a signaling molecule in cell
migration has been reported by several groups. In the present
study, it was demonstrated that the NOX inhibitor VAS2870
eliminated TGF-β-dependent migration in RPE cells in a
concentration-dependent manner. These findings are consistent with
a previous report demonstrating that this molecule can inhibit
TGF-β-dependent migration in vascular smooth muscle cells (22). In addition, Boudreau et al
indicated that NOX4 was involved in TGF-β-induced lung and breast
epithelial cell migration (23).
However, the role of TGF-β in cell proliferation
remains to be fully elucidated. Treatment with VAS2870 inhibited
cell cycle progression through the G1 phase, as
determined by flow cytometry. Several studies have shown that other
NOX inhibitors reduce proliferation in various cell types. For
example, in pulmonary vascular cells, hypoxia-induced proliferation
was significantly suppressed by the inhibition of NOX activity
using the NOX inhibitor GKT137831 (24). In addition, NOX4 was shown to
mediate proliferation in response to TGF-β in human pulmonary
artery smooth muscle cells (25).
Notably, the present study showed that VAS2870
reversed TGF-β2-induced EMT in RPE cells. During this process,
expression of the epithelial differentiation marker E-cadherin in
these cells was lost and the expression of mesenchymal marker α-SMA
increased (26,27). E-cadherin, a cell-cell adhesion
molecule, is located at the cell-cell boundary of the normal
epithelium (28) and is
associated with invasion and metastasis in cancer (29). The present study showed that this
protein was upregulated in the membrane of RPE cells, indicating
that it can act as an intercellular adhesion molecule. Consistent
with these findings, angiotensin II-induced hepatocyte EMT was
previously reported to be inhibited by NOX4 small interfering
(si)RNA in hepatocytes (30). In
the lens capsule, VAS2870 was shown to inhibit the expression of
NOX4, which is the only NOX isoform expressed during TGF-β2-induced
EM; those cells which survived following VAS2870 treatment retained
an epithelial-like phenotype, indicating that VAS2870 reversed EMT
through the inhibition of NOX4 (31).
NOXs have been shown to transmit downstream TGF-β
signaling. In skeletal muscle cells, TGF-β induces its own
expression, and the NOX pharmacological inhibitor apocynin has been
shown to inhibit this process (32). By contrast, in liver tumor cells,
VAS2870 effectively attenuates serum-dependent growth and the
phosphorylation of AKT and extracellular signal-regulated kinase
(33). In addition, Boudreau
et al demonstrated that silencing NOX4 prevented cell
migration mediated by the small mothers against decapentaplegic
(Smad) signaling pathway in breast cancer cells (34). In the investigation of ocular
disease, NOX4 was shown to be required for complete activation of
mitogen-activated protein kinase (MAPK) signaling upstream of
reactive oxygen species in human bronchial epithelial cells during
EMT (35). Taken together, these
findings suggest that NOX4 may act as a common mediator of these
pathways and that redox signaling may serve as a nexus between
parallel signaling pathways, including those of Smads and
MAPKs.
In conclusion, the results of the present study
indicated that the pharmacological inhibition of NOX with VAS2870
effectively impaired the proliferation of RPE cells. VAS2870
treatment also attenuated the migration and EMT induced by TGF-β2,
a physiological pro-EMT stimulus. Notably, the effects of VAS2870
on NOX enzymes were not isoform-specific, and the final results may
have been representative of the combined effects of inhibiting all
isoforms expressed in the cells. In RPE cells, other isoforms,
including NOX1 and NOX2, were expressed at low levels. In
particular, NOX4 appeared to be crucial in RPE cells. Therefore,
the pharmacological inhibition of NOX may provide insights into the
development of improved treatments for PVR. However, there were
limitations to the present study. First, only the biological
behaviors of RPE cells following NOX4 inhibition were observed, and
the molecular mechanisms responsible for this process were not
elucidated. Further experiments are required to examine the
efficiency of NOX4 silencing by siRNA, using the same experimental
conditions, and to examine the effects of VAS2870 on other signal
transduction pathways, to fully elucidate the complex interactions
among these pathways. The pharmacological inhibition of NOX may be
a promising therapeutic approach for proliferative retinopathy and
for the prevention of secondary PVR.
Acknowledgments
Not applicable.
References
|
1
|
Ho PC and McMeel JW: Retinal detachment
with proliferative vitreoretinopathy: Surgical results with scleral
buckling, closed vitrectomy, and intravitreous air injection. Br J
Ophthalmol. 69:584–587. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kroll P, Rodrigues EB and Hoerle S:
Pathogenesis and classification of proliferative diabetic
vitreoretinopathy. Ophthalmologica. 221:78–94. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tosi GM, Marigliani D, Romeo N and Toti P:
Disease pathways in proliferative vitreoretinopathy: An ongoing
challenge. J Cell Physiol. 229:1577–1583. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cui JZ, Chiu A, Maberley D, Ma P, Samad A
and Matsubara JA: Stage specificity of novel growth factor
expression during development of proliferative vitreoretinopathy.
Eye. 21:200–208. 2007. View Article : Google Scholar
|
|
5
|
Pastor JC, de la Rúa ER and Martín F:
Proliferative vitreoretinopathy: Risk factors and pathobiology.
Prog Retin Eye Res. 21:127–144. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Grisanti S and Guidry C:
Transdifferentiation of retinal pigment epithelial cells from
epithelial to mesenchymal phenotype. Invest Ophthalmol Vis Sci.
36:391–405. 1995.PubMed/NCBI
|
|
7
|
Pennock S, Haddock LJ, Eliott D, Mukai S
and Kazlauskas A: Is neutralizing vitreal growth factors a viable
strategy to prevent proliferative vitreoretinopathy. Prog Retin Eye
Res. 40:16–34. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lassègue B: Clempus RE. Vascular NAD(P)H
oxidases: Specific features, expression, and regulation. Am J
Physiol Regul. Integr Comp Physiol. 285:R277–R297. 2003. View Article : Google Scholar
|
|
9
|
Brown DI and Griendling KK: Nox proteins
in signal transduction. Free Radic Biol Med. 47:1239–1253. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kita T, Hata Y, Kano K, Miura M, Nakao S,
Noda Y, Shimokawa H and Ishibashi T: Transforming growth
factor-beta2 and connective tissue growth factor in proliferative
vitreoretinal diseases: Possible involvement of hyalocytes and
therapeutic potential of Rho kinase inhibitor. Diabetes.
56:231–238. 2007. View Article : Google Scholar
|
|
11
|
Xu J, Lamouille S and Derynck R:
TGF-beta-induced epithelial to mesenchymal transition. Cell Res.
19:156–172. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qiu Y, Tao L, Lei C, Wang J, Yang P, Li Q
and Lei B: Downregulating p22phox ameliorates inflammatory response
in Angiotensin II-induced oxidative stress by regulating MAPK and
NF-κB pathways in ARPE-19 cells. Sci Rep. 5:143622015. View Article : Google Scholar
|
|
13
|
Kim JH, Park S, Chung H and Oh S: Wnt5a
attenuates the pathogenic effects of the Wnt/beta-catenin pathway
in human retinal pigment epithelial cells via down-regulating
β-catenin and Snail. BMB Rep. 48:525–530. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Michaeloudes C, Sukkar MB, Khorasani NM,
Bhavsar PK and Chung KF: TGF-β regulates Nox4, MnSOD and catalase
expression, and IL-6 release in airway smooth muscle cells. Am J
Physiol Lung Cell Mol Physiol. 300:L295–L304. 2011. View Article : Google Scholar
|
|
15
|
Li S, Tabar SS, Malec V, Eul BG, Klepetko
W, Weissmann N, Grimminger F, Seeger W, Rose F and Hänze J: NOX4
regulates ROS levels under normoxic and hypoxic conditions,
triggers proliferation, and inhibits apoptosis in pulmonary artery
adventitial fibroblasts. Antioxid Redox Signal. 10:1687–1698. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pan X, Dai Y, Li X, Niu N, Li W, Liu F,
Zhao Y and Yu Z: Inhibition of arsenic-induced rat liver injury by
grape seed exact through suppression of NADPH oxidase and
TGF-β/Smad activation. Toxicol Appl Pharmacol. 254:323–331. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lapperre TS, Jimenez LA, Antonicelli F,
Drost EM, Hiemstra PS, Stolk J, MacNee W and Rahman I: Apocynin
increases glutathione synthesis and activates AP-1 in alveolar
epithelial cells. FEBS Lett. 443:235–239. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Engels F, Renirie BF, Hart BA, Labadie RP
and Nijkamp FP: Effects of apocynin, a drug isolated from the roots
of Picrorhiza kurroa, on arachidonic acid metabolism. FEBS Lett.
305:254–256. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Majander A, Finel M and Wikström M:
Diphenyleneiodonium inhibits reduction of iron-sulfur clusters in
the mitochondrial NADH-ubiquinone oxidoreductase (Complex I). J
Biol Chem. 269:21037–21042. 1994.PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods.
25:402–408. 2001. View Article : Google Scholar
|
|
21
|
Ishikawa K, He S, Terasaki H, Nazari H,
Zhang H, Spee C, Kannan R and Hinton DR: Resveratrol inhibits
epithelial-mesenchymal transition of retinal pigment epithelium and
development of proliferative vitreoretinopathy. Sci Rep.
5:163862015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
ten Freyhaus H, Huntgeburth M, Wingler K,
Schnitker J, Bäumer AT, Vantler M, Bekhite MM, Wartenberg M, Sauer
H and Rosenkranz S: Novel Nox inhibitor VAS2870 attenuates
PDGF-dependent smooth muscle cell chemotaxis, but not
proliferation. Cardiovasc Res. 71:331–341. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Boudreau HE, Casterline BW, Burke DJ and
Leto TL: Wild-type and mutant p53 differentially regulate NADPH
oxidase 4 in TGF-β-mediated migration of human lung and breast
epithelial cells. Br J Cancer. 110:2569–2582. 2014. View Article : Google Scholar :
|
|
24
|
Green DE, Murphy TC, Kang BY, Kleinhenz
JM, Szyndralewiez C, Page P, Sutliff RL and Hart CM: The Nox4
inhibitor GKT137831 attenuates hypoxia-induced pulmonary vascular
cell proliferation. Am J Respir Cell Mol Biol. 47:718–726. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Martin-Garrido A, Brown DI, Lyle AN,
Dikalova A, Seidel-Rogol B, Lassègue B, San Martín A and Griendling
KK: NADPH oxidase 4 mediates TGF-β-induced smooth muscle α-actin
via p38MAPK and serum response factor. Free Radic Biol Med.
50:354–362. 2011. View Article : Google Scholar
|
|
26
|
Tobar N, Toyes M, Urra C, Méndez N,
Arancibia R, Smith PC and Martínez J: c-Jun N terminal kinase
modulates NOX-4 derived ROS production and myofibroblasts
differentiation in human breast stromal cells. BMC Cancer.
14:6402014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Oka H, Shiozaki H, Kobayashi K, Inoue M,
Tahara H, Kobayashi T, Takatsuka Y, Matsuyoshi N, Hirano S,
Takeichi M, et al: Expression of E-cadherin cell adhesion molecules
in human breast cancer tissues and its relationship to metastasis.
Cancer Res. 53:1696–1701. 1993.PubMed/NCBI
|
|
28
|
Li H, Wang H, Wang F, Gu Q and Xu X: Snail
involves in the trans forming growth factor β1-mediated
epithelial-mesenchymal tran sition of retinal pigment epithelial
cells. PLoS One. 6:e233222011. View Article : Google Scholar
|
|
29
|
Palma-Nicolás JP and López-Colomé AM:
Trombin induces slug-mediated E-cadherin transcriptional repression
and the parallel up-regulation of N-cadherin by a
transcription-independent mechanism in RPE cells. J Cell Physiol.
228:581–589. 2013. View Article : Google Scholar
|
|
30
|
Zhang LL, Huang S, Ma XX, Zhang WY, Wang
D, Jin SY, Zhang YP, Li Y and Li X: Angiotensin(1-7) attenuated
angiotensin II-induced hepatocyte EMT by inhibiting NOX-derived
H2O2-activated NLRP3 inflammasome/IL-1β/Smad
circuit. Free Radic Biol Med. 97:531–543. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Das SJ, Lovicu FJ and Collinson EJ: Nox4
plays a role in TGF-β-dependent lens epithelial to mesenchymal
transition. Invest Ophthalmol Vis Sci. 57:3665–3673. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Abrigo J, Morales MG, Simon F, Cabrera D,
Di Capua G and Cabello-Verrugio C: Apocynin inhibits the
upregulation of TGF-β1 expression and ROS production induced by
TGF-β in skeletal muscle cells. Phytomedicine. 22:885–893. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sancho P and Fabregat I: The NADPH oxidase
inhibitor VAS2870 impairs cell growth and enhances TGF-β-induced
apoptosis of liver tumor cells. Biochem Pharmacol. 81:917–924.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Boudreau HE, Casterline BW, Rada B,
Korzeniowska A and Leto TL: Nox4 involvement in TGF-beta and
SMAD3-driven induction of the epithelial-to-mesenchymal transition
and migration of breast epithelial cells. Free Radic Biol Med.
53:1489–1499. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ge A, Ma Y, Liu YN, Li YS, Gu H, Zhang JX,
Wang QX, Zeng XN and Huang M: Diosmetin prevents TGF-β1-induced
epithelial-mesenchymal transition via ROS/MAPK signaling pathways.
Life Sci. 153:1–8. 2016. View Article : Google Scholar : PubMed/NCBI
|