Introduction
Acute otitis media is reported as one of the most
common diseases due to viral, or fungal pathogens and bacterial
infection (1,2). The progression, pathophysiology as
well as pathogenesis of acute otitis media are influenced by
various factors, such as pathogenicity of the pathogens, reactive
oxygen species (ROS) generation and inflammatory cytokines
secretion (3,4). Presently, ~10–20% children
experience recurrence and persistence of otitis media with
long-term loss of hearing (5).
Finding effective therapy is urgent for clinical treatment.
Studies before have indicated that a large number of
cytokines or chemokines participate in acute otitis media
pathogenesis (6).
Lipopolysaccharides (LPS) are from gram-negative bacteria, which
has been suggested to induce pro-inflammatory cytokine release,
including tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β)
and IL-6 (7). It has been
reported that TNF-α can regulate the activity and expression of
Toll-like receptors (TLRs), which is also important for TLR
down-stream signaling pathway, including the nuclear factor-κB
(NF-κB) (8). Recently, studies
have suggested an essential function of TLR4 signaling pathway
through MyD88-dependent pathway during the middle ear inflammatory
responding to bacteria infection (9). Further, they are also crucial for
the recovery process from otitis media (10).
Thioredoxin-interacting protein (TXNIP) is evidenced
as a binding partner to the Nod-like receptor protein 3 (NLRP3)
inflammasome, which is involved in activation of ROS-related NLRP3
inflammasome (11). NLRP3
inflammasome is known as a protein complex, consisting of an NLRP3
inflammasome sensor, the adaptor protein ASC and caspase-1
(12). TXNIP/NLRP3 signaling
pathway is well reported to regulate inflammatory response through
regulating NF-κB transcription to induce IL-1β pro-inflammatory
cytokines release (13). ERK1/2,
and p38 MAPK are well introduced for their role in ROS generation
(14). Previously, suppression of
MAPKs was reported to improve otitis media, indicating its role in
otitis media progression.
Fisetin, a natural polyphenol found in various
plants, including vegetables and fruits, has been shown to be able
to defend both inflammation response and oxidative stress in cells,
exhibiting potent anti-inflammatory and anti-oxidative properties
in various conditions (15,16). Analyses of the related signaling
pathways modulated by the polyphenol have indicated that fisetin
has the ability to suppress the activation of NF-κB, ERK1/2, p38
MAPK, and the secretion of pro-inflammatory cytokines, such as
IL-1β, IL-6 and TNF-α in a variety of different cells (17). Thus, we supposed that fisetin may
be of potential value in treating LPS-induced acute otitis media by
inflammation and ROS inhibition. The present study evaluated the
role of fisetin in acute otitis media induced by LPS. We also
investigated the signaling pathways underlying the possible
inflammatory responses and ROS in the disease in mice, as well as
the function of fisetin during the acute otitis media
formation.
Materials and methods
The establishment of animal models
Sixty male, 8-week-old, C57BL/6 mice weighing 18–22
g were purchased from Shanghai Laboratory Animal Research Center
(Shanghai, China). The mice were housed in a constant temperature
of 22±2°C and relative humidity of 60±10% environment under 12 h
light/dark cycles at the Third Affiliated Hospital of Sun Yat-sen
University. The animals were permited free access to water and
chow, and were housed and fed in the specific pathogen-free
facility. The mouse experiments were conducted minimizing animal
suffering following the Guide for the Care and Use of Laboratory
Animals, issued by the National Institutes of Health in 1996. All
animal procedures were done following the guidelines for Care and
Use of Laboratory Animals approved by Sun Yat-sen University. The
acute otitis media mouse model was established through LPS
injection (1.0 mg/ml; Sigma, St. Louis, CA, USA) into the middle
ear of the mice via tympanic membrane in the right ear only for 24
h. The mice (n=15) without LPS treatment served as the control
(Con). Forty-three mice were induced with acute otitis media
successfully. After LPS exposure for 24 h, 36 mice with acute
otitis media were randomly divided into two groups. Eighteen mice
after LPS treatment were given low dose of 10 mg/kg of fisetin
(FisL) dissolved in 5% dimethyl sulfoxide (DMSO) in
phosphate-buffered saline (PBS) by intragastric administration
every day for 10 days, and the other 18 mice after LPS treatment
were given high dose of 20 mg/kg of fisetin (FisH) dissolved in 5%
DMSO in PBS through intragastric administration every day for 10
days. Finally, the mice were sacrificed and the eyeball blood was
collected and centrifuged at 15,000 × g for 20 min prepared for
following research. Middle ear lavage fluids (MELF) were collected,
and then centrifuged at 500 × g for 15 min, and the single-use
aliquots of MELF were stored at −80°C. The cell pellets were then
washed twice for further detections. Middle ear effusions from mice
were harvested with 10 μl saline (×3) first, and then the
middle ears were washed with 200 μl physiologic saline.
Enzyme-linked immunosorbent assay
(ELISA)
Concentrations of cytokines of IL-1β, TNF-α, IL-6
and VEGF in the MEE and serum of mice were determined by the ELISA
with the mouse enzyme immunoassay sets (R&D Systems, Inc.,
Minneapolis, MN, USA) following the manufacturer's instructions.
The samples were performed in duplicate.
Assessment of bioindicators
All the serum stored at −80°C were removed and
maintained in glacial table. SOD activity and MDA levels in serum
and MEE were tested using biochemical kit, Superoxide Dismutase
assay kit and Maleic Dialdehyde assay kit, respectively (Jiancheng
Biotech Co., Ltd., Nanjing, China) following the manufacturer's
instruction.
Real-time quantitative polymerase chain
reaction (RT-qPCR) analysis
Total RNA from the middle ear tissue samples was
isolated using TRIzol (Invitrogen, Carlsbad, CA, USA) following the
manufacturer's instructions. The cDNA was synthesized using
SuperScript II reverse transcriptase (Thermo Fisher Scientific,
Waltham, MA, USA). Quantitative PCR was performed with SYBR-Green
Real-Time PCR Master mix (Thermo Fisher Scientific). Finally, the
quantitative expression data were collected and analyzed by a 7900
Real-time PCR system (Applied Biosystems, Foster City, CA, USA).
Primers were designed to determine endogenous genes and
glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as the
endogenous control. The crossing point of target genes with GAPDH
was calculated using the formula 2-(target gene-GAPDH) and the
relative amounts were quantified. Forward TNF-α, (5′-3′) CTT CGT
ACT ATC ATG TAG GTC GAG and reverse primers, (5′-3′) ATG CTC GGC
ATG TTT GAA CTA GTC; forward IL-1β, (5′-3′) ATG CGG TAG CAA GTT ACG
AC and reverse primers, (5′-3′) GAG GTG GCT GTT GGT ATT GAT;
forward IL-6, (5′-3′) CTA AGG ACG GAG AGT CAG T and reverse
primers, (5′-3′) CTG TCG TCG ATG AGA TTA TG; forward VEGF, (5′-3′)
CCT CCT GGC GCT CTG ATA TGT and reverse, (5′-3′) GTG AGT GTG TAG
GTG TGC GC; forward TXNIP, (5′-3′) CTT GCC GTC ATC TTC CTA CA and
reverse primers, (5′-3′) GCC AAG GTA AAG GAG GCA CT; forward NLRP3,
(5′-3′) GCT CAC TCT TAT CTA TCC CGA and reverse primers, (5′-3′)
ACA AGG CGT AGC AGA GAG CT; forward GAPDH, (5′-3′) AGT CTA CGT GCA
ATC AAC AGA ATG and reverse primers, (5′-3′) CGC CCT AAC AAC ACG
ACA TCC AT.
Western blot assays
The middle ear tissues were harvested and frozen in
liquid nitrogen immediately, and then were stored in −80℃ until
homogenization. Proteins were extracted from the middle ear tissue
samples using T-PER tissue protein extraction reagent kit (Thermo
Fisher Scientific) according to the manufacturer's instructions.
Protein concentrations were determined by BCA protein assay kit
(Thermo Fisher Scientific) following the manufacturer's
instruction, and equal amounts (40 μg) of protein were
loaded per well on a 10% sodium dodecyl sulphatepolyacrylamide gel.
Subsequently, proteins were transferred onto polyvinylidene
difluoride membrane. The resulting membrane was blocked with
Tris-buffered saline containing 0.05% Tween-20 (TBS-T),
supplemented with 5% skim milk (Sigma) at room temperature for 2 h
on a rotary shaker, and followed by TBS-T washing. The specific
primary antibody, diluted in TBST, was incubated with the membrane
at 4°C overnight. Subsequently, the membrane was washed with TBS-T
followed by incubation with the horseradish peroxidase
(HRP)-conjugated secondary antibody at room temperature for 1 h.
The immunoactive proteins were detected by using an enhanced
chemiluminescence western blot detection kit. Western blot bands
were observed using GE Healthcare ECL western blotting analysis
system (GE Healthcare, Logan, UT, USA) and exposed to Kodak X-ray
film. The primary antibodies used in our study are as follows:
rabbit anti-GAPDH (1:1,000; Cell Signaling Technology, Beverly, MA,
USA), anti-TLR-4 (1:1,000), mouse rabbit anti-MyD88 (1:1,000),
rabbit anti-p-IKKα (1:1,000) (all from Abcam, Cambridge, MA, USA),
rabbit anti-p-IκBα (1:1,000), rabbit anti-p-NF-κB (1:1,000), rabbit
anti-NF-κB (1:1,000), rabbit anti-caspase-3 (1:1,000) (all from
Cell Signaling Technology), rabbit anti-PARP (1:1,000; Abcam),
rabbit anti-Bax (1:1,000; Cell Signaling Technology), rabbit
anti-Bad (1:1,000), mouse anti-Bcl-2 (1:1,000), rabbit anti-Bcl-xL
(1:1,000) (all from Abcam), rabbit anti-SOD1 (1:1,000), rabbit
anti-SOD2 (1:1,000) (both from Cell Signaling Technology), rabbit
anti-HO-1 (1:1,000), rabbit anti-Nrf2 (1:1,000), rabbit anti-TXNIP
(1:1,000), rabbit anti-NLRP3 (1:1,000), rabbit anti-ASC (1:1,000)
(all from Abcam), rabbit anti-caspase-1 (1:1,000; Cell Signaling
Technology), rabbit anti-p-ERK1/2 (1:1,000), rabbit anti-ERK1/2
(1:1,000) (both from Abcam), rabbit anti-p38 (1:1,000), and rabbit
anti-p-p38 (1:1,000) (both from Cell Signaling Technology).
Flow cytometry analysis
The cells in MELF were centrifuged at 500 × g for 10
min, the cell pellets were washed twice with cold PBS, and next the
Fc receptors were fine blocked using the Mouse BD Fc Block
purchased from BD Biosciences (Franklin Lakes, NJ, USA). Then, the
cells were stained using specific immune (CD11b; BD Biosciences)
markers on cell surface, followed by the staining parameters: cells
experiencing early apoptosis marked as Annexin V-positive and
propidium iodide-negative, and the cells during late apoptosis
described as Annexin V-positive and propidium iodide-positive.
Immunohistochemical assays
Histopathologic evaluation was performed on mice.
Mouse middle ear tissue samples were fixed with 10% buffered
formalin, imbedded in paraffin, and sliced into 5 μM
sections. After hematoxylin and eosin (H&E) staining,
pathological changes of the tissues were observed under a light
microscope. For fluorescent analysis, the mouse middle ear tissue
samples were carefully isolated and fixed in 4% paraformaldehyde
for 16 h after cold 4% paraformaldehyde perfusion. Then, optimum
cutting temperature (OCT) package tissues were sliced as 20–30
μm sections. The tissues were incubated with primary
antibodies (TXNIP, NLRP3, p-p38 and p-ERK1/2) at 4°C overnight
after deparaffinized and rehydrated. Fluorophore-conjugated
secondary antibodies were treated 1 h at 25°C thermostat. The Alexa
Fluor 488 labeled anti-rabbit or anti-mouse secondary antibodies
(Invitrogen) were used. Sections were subjected to
immunofluorescence staining via epifluorescence microscopy (Sunny
Co., Shanghai, China). Leica TCS SP5 confocal microscope (Leica,
Richmond Hill, ON, Canada) was used to obtain images and carried
out in a blinded manner with respect to treatment groups.
Statistical analysis
Data are expressed as means ± SEM. Treated tissues
and the corresponding controls were compared using GraphPad Prism
(version 6.0; GraphPad Software, La Jolla, CA, USA) by one-way
ANOVA with Dunn's least significant difference tests. Differences
between groups were considered significant at p<0.05.
Results
Fisetin ameliorates the middle ear injury
of mice after LPS exposure
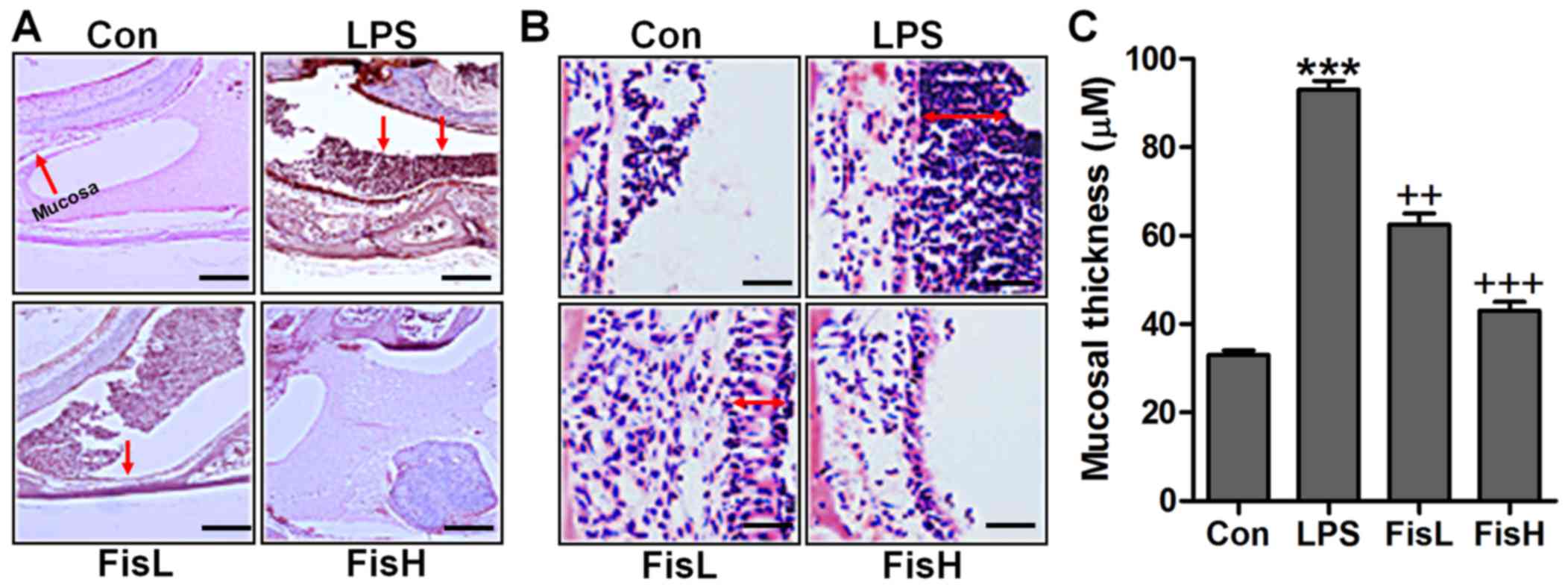
In order to prove the successful establishment of
acute otitis media in the mouse model, the histologic sections and
the mucosa thickness in the mouse middle ear were observed through
H&E staining. As shown in Fig.
1A, the normal control groups showed no signs of inflammatory
response, while in the LPS-treated mice, the cells exhibited brown
color suggesting significant inflammation, which was comparable to
the control group. Of note, fisetin administration apparently
reduced the number of inflammatory cells, mainly including
monocytes, macrophages, leukocyte, neutrophil and eosinophils
(18,19). In addition, the photomicrographs
indicated the mucosa in middle ear of mice with LPS induction. By
assessment, the mucosa in the roof of LPS-treated mice with acute
otitis media was much thicker compared to the control ones, which
was reduced for fisetin treatment at different concentrations
(Fig. 2B and C). The findings
above indicated that LPS indeed induced acute otitis media in mice,
and interestingly, fisetin showed ameliorated effects on
LPS-triggered acute otitis media in the middle ear of mice.
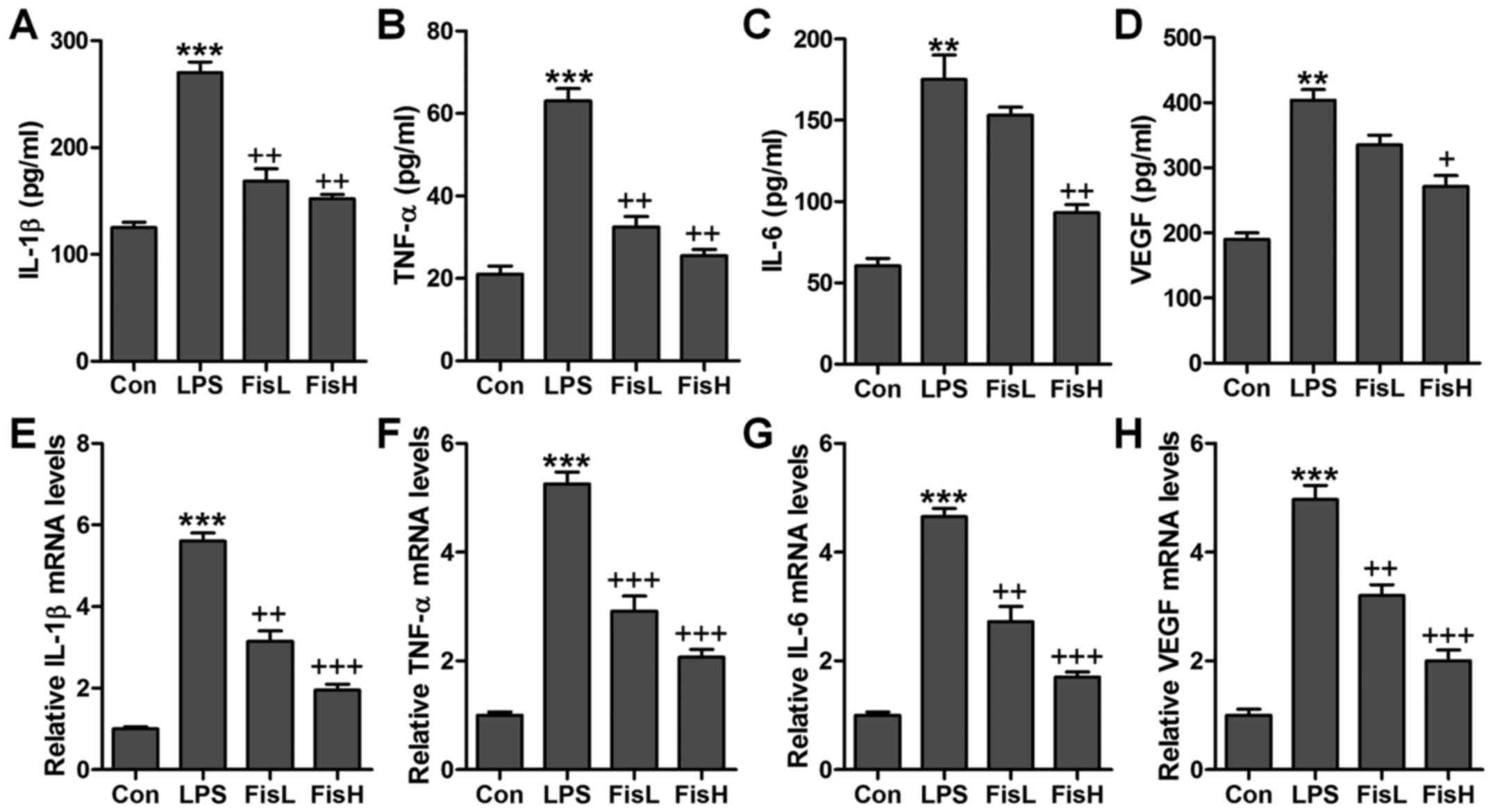
Fisetin reduces pro-inflammatory cytokine
release in LPS-induced mice with acute otitis media
Inflammation response has been suggested to be of
great importance to cause injury in the middle ear under different
conditions, such as bacteria exposure (20). According to the observations
above, the ear injury was induced in mice after LPS exposure, which
is well known to induce inflammation response in various diseases
(21). Thus, here we attempted to
investigate whether fisetin-improved acute otitis media was
associated with inflammation suppression, pro-inflammatory
cytokines were measured in the serum and middle ear tissue samples
obtained from mice after LPS treatment. The pro-inflammatory
cytokines, IL-1β, TNF-α, IL-6 and VEGF, were highly accumulated in
LPS-treated mice, which were reduced for fisetin administration
with significant difference in comparison to the LPS group
(Fig. 2A–D). Further, these
factors in the dissected tissue samples of middle ear were
calculated through RT-qPCR assays. The results illustrated that
IL-1β, TNF-α, IL-6 and VEGF gene abundance occurred in LPS-treated
group in the absence of fisetin. However, fisetin administration
dramatically downregulated IL-1β, TNF-α, IL-6 and VEGF mRNA levels
in a dose-dependent manner (Fig.
2E–H). The data above demonstrated that inflammation response
was induced in the middle ear of mice with LPS treatment, and
fisetin exhibited effective role in reversing pro-inflammatory
cytokine secretion.
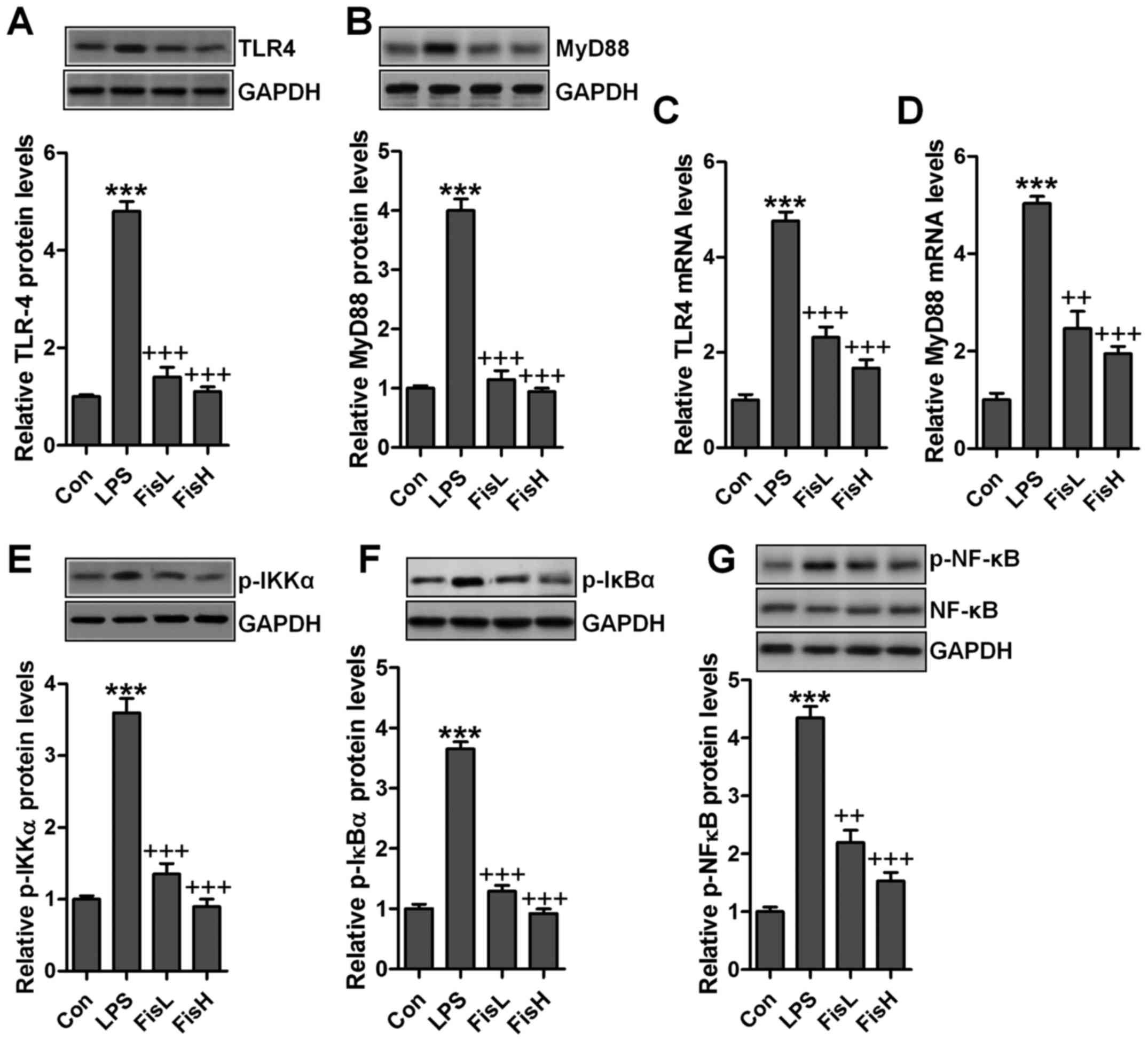
Fisetin ameliorates LPS-induced
inflammation in the middle ear of mice through TLR4/NF-κB signaling
pathway
As our above data indicated, inflammation response
was indeed induced in LPS-treated mice with acute otitis media in
the middle ear. Fisetin was evidenced to be repressive for
pro-inflammatory cytokine release. TLR4/NF-κB signaling pathway is
well reported to be essential for inflammation response through
transcription factor activity regulation, contributing to
pro-inflammatory cytokine secretion (22). Hence, in this regard, we attempted
to explore if the classic TLR4/NF-κB signaling pathway participated
in fisetin-improved acute otitis media in LPS-induced mice. As
shown in Fig. 3A and B, TLR4
protein levels in the middle ear tissue samples were found to be
highly expressed due to LPS treatment by the use of western blot
analysis, leading to the downstream signal MyD88 upregulation.
Notably, fisetin reduced TLR4 and MyD88 protein expression levels,
which was comparable to the LPS group. Also, RT-qPCR assays showed
similar results that fisetin had a suppressive role in TLR4 and
MyD88 activity induced by LPS (Fig.
3C and D). NF-κB phosphorylation is crucial for inflammation
response regulated by TLR4/MyD88 signaling pathway. LPS treatment
considerably increased IKKα and IκBα phosphorylation, which
improved NF-κB phosphorylation (Fig.
3E–G). In contrast, fisetin administration apparently
restrained IKKα, IκBα and NF-κB activity, in line with the results
of pro-inflammatory cytokine alteration mentioned above. The
results suggested that fisetin-improved acute otitis media caused
by LPS was dependent on TLR4/NF-κB signaling pathway
suppression.
Fisetin inhibits apoptosis in the middle
ear of mice treated with LPS
Previous studies indicated that apoptosis was
closely related to acute otitis media induced by bacterial
(23). Thus, the neutrophil
apoptosis may be involved in fisetin against LPS challenging in
acute otitis media. Here, flow cytometry analysis showed that cells
in the MELF obtained from LPS-treated mice underwent significant
apoptosis, while fisetin-treated groups at different concentrations
displayed markedly downregulated rate (Fig. 4A and B). Caspase-3 enhances
apoptotic response via PARP activation (24). Pro-apoptotic members and
anti-apoptotic molecules are in homeostasis under normal condition,
which will be disrupted for different stress exposure, including
LPS. In this study, we found that caspase-3 and PARP cleavage was
highly upregulated for LPS induction, contributing to cell death in
the middle ear of mice (Fig.
4C–E). Additionally, pro-apoptotic signals, Bax and Bad were
also expressed in abundance from the protein levels, which was in
agreement with cleaved caspase-3 and PARP (Fig. 4F and G). On the contrary,
anti-apoptotic members, Bcl-2 and Bcl-xL protein levels were
downregulated due to LPS (Fig. 4H and
I). Fisetin treatment decreased caspase-3 and PARP activation,
accompanied with reduced Bax and Bad, whereas Bcl-2 and Bcl-xL were
obviously increased. Moreover, RT-qPCR assays showed that Bax and
Bad mRNA levels were augmented in LPS group, which was comparable
with the control ones, while being impeded after fisetin treatment
(Fig. 4J and K). Consistent with
preotein alterations, Bcl-2 and Bcl-xL gene levels were reduced for
LPS, and fisetin reversed Bcl-2 and Bcl-xL expression in a
dose-dependent manner (Fig. 4L and
M). Taken together, the data illustrated that fisetin could
improve acute otitis media through apoptosis suppression via
inactivating caspase-3 signaling pathway.
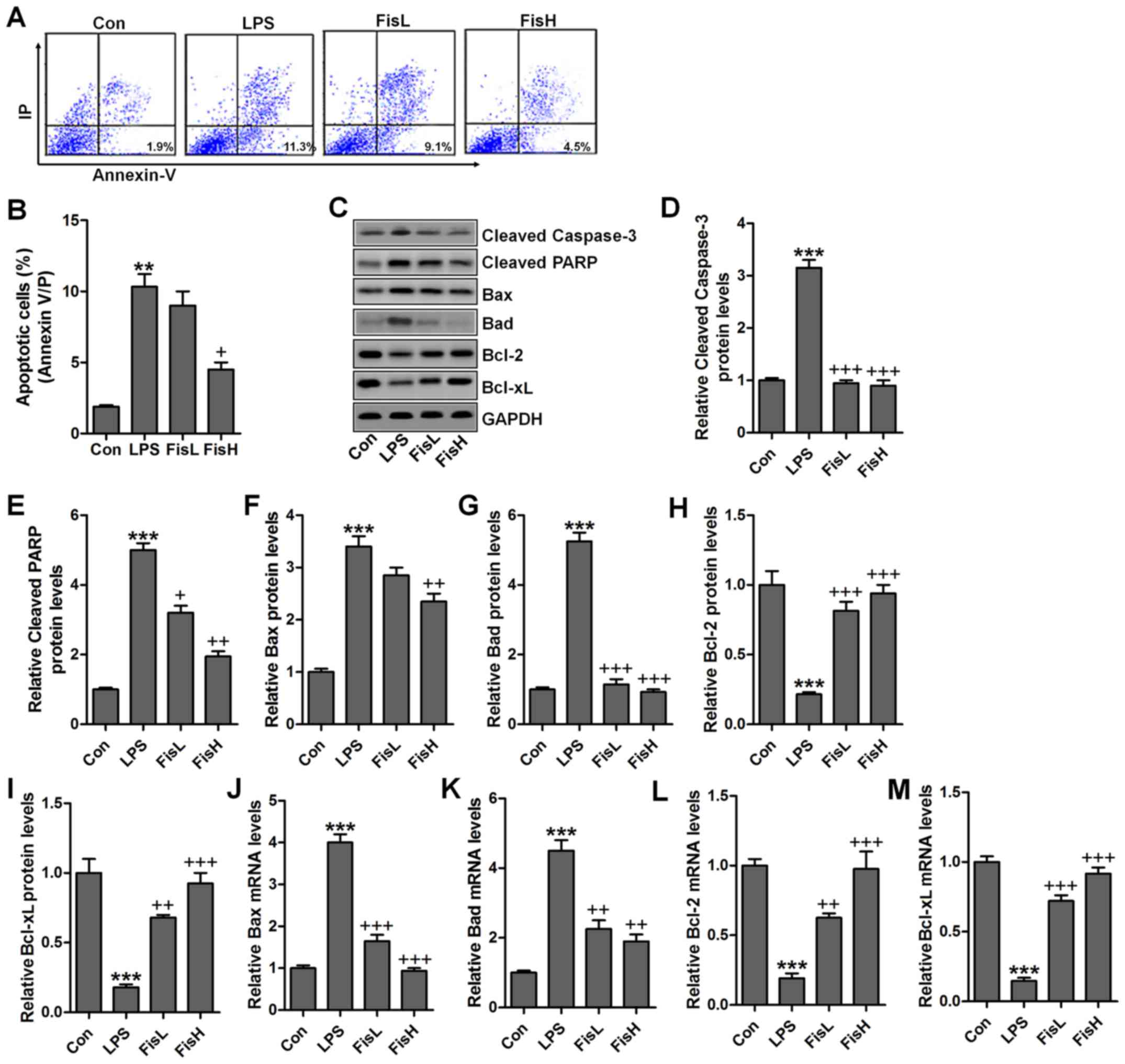 | Figure 4Fisetin inhibits apoptosis in the
middle ear of mice treated with lipopolysaccharides (LPS). (A) Flow
cytometry was used to examine apoptosis in lipopolysaccharide
(LPS)-treated mice with acute otitis media after fisetin
administration at different concentrations. (B) The percentage of
apoptotic cells following flow cytometry analysis was shown. (C)
The representative images of cleaved caspase-3, cleaved PARP, Bax,
Bad, Bcl-2, and Bcl-xL were shown through western blot analysis.
The quantification of (D) cleaved caspase-3, (E) cleaved PARP, (F)
Bax, (G) Bad, (H) Bcl-2, and (I) Bcl-xL is exhibited. RT-qPCR
assays were used to determine (J) Bax, (K) Bad, (L) Bcl-2 and (M)
Bcl-xL gene levels. Data are expressed as the mean ± SEM (n=10).
*p<0.05, **p<0.01 and
***p<0.001 vs. the control (Con) group;
+p<0.05, ++p<0.01 and
+++p<0.001 vs. the LPS group. |
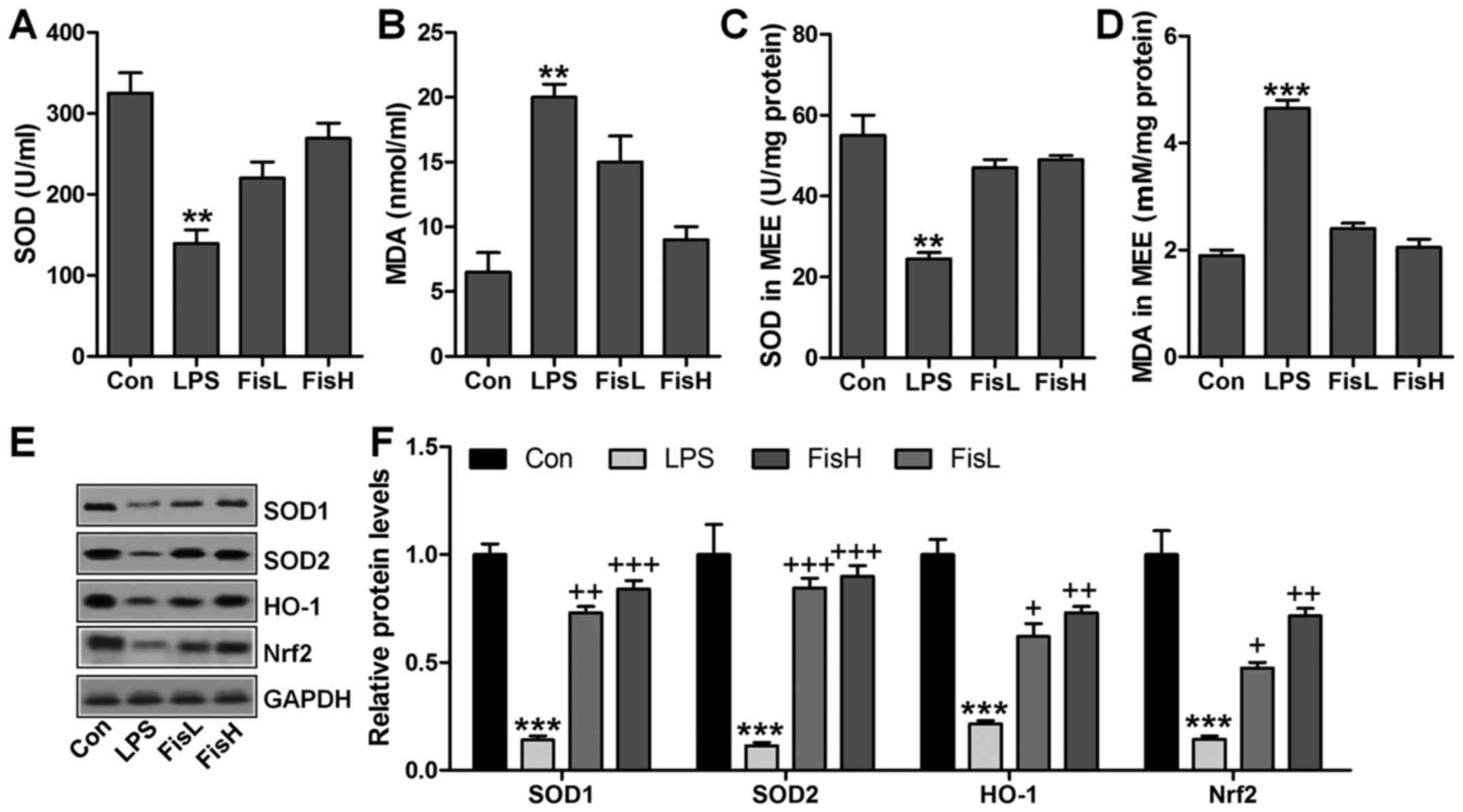
Fisetin upregulates anti-oxidant levels
in LPS-exposed mice with acute otitis media
ROS generation with decreased anti-oxidants is
considered to induce tissue injury or organ dysfunction for various
reasons, including LPS induction (25). In this regard, ROS-related signals
were calculated to reveal that if fisetin-improved acute otitis
media was related to ROS suppression. As shown in Fig. 5A, SOD activity in serum was found
to be decreased for LPS treatment, which recovered highly for
fisetin administration. In contrast, LPS-triggered higher MDA
levels were significantly reduced for fisetin treatment (Fig. 5B). Consistently, in MEE, SOD
activity and MDA levels for LPS challenge were similar to that in
serum, and fisetin reversed expression of the two indicators to
near normal levels (Fig. 5C and
D). For further confirmation, western blot analysis was
performed to determine SOD1, SOD2, HO-1 and Nrf2 protein levels in
the middle ear of mice under different conditions. As shown in
Fig. 5E and F, SOD1, SOD2, HO-1
and Nrf2 protein levels were apparently reduced in LPS
administration, which were highly improved by fisetin treatment, in
a dose-dependent manner. In conclusion, the data here indicated
that ROS production induced by LPS could be decreased in fisetin
treatment to improve acute otitis media in mice.
Fisetin-improved acute otitis media
triggered by LPS is associated with TXNIP and MAPKs signaling
pathways
According to previous studies, TXNIP and MAPKs are
involved in inflammtion and ROS production through NLRP3 and
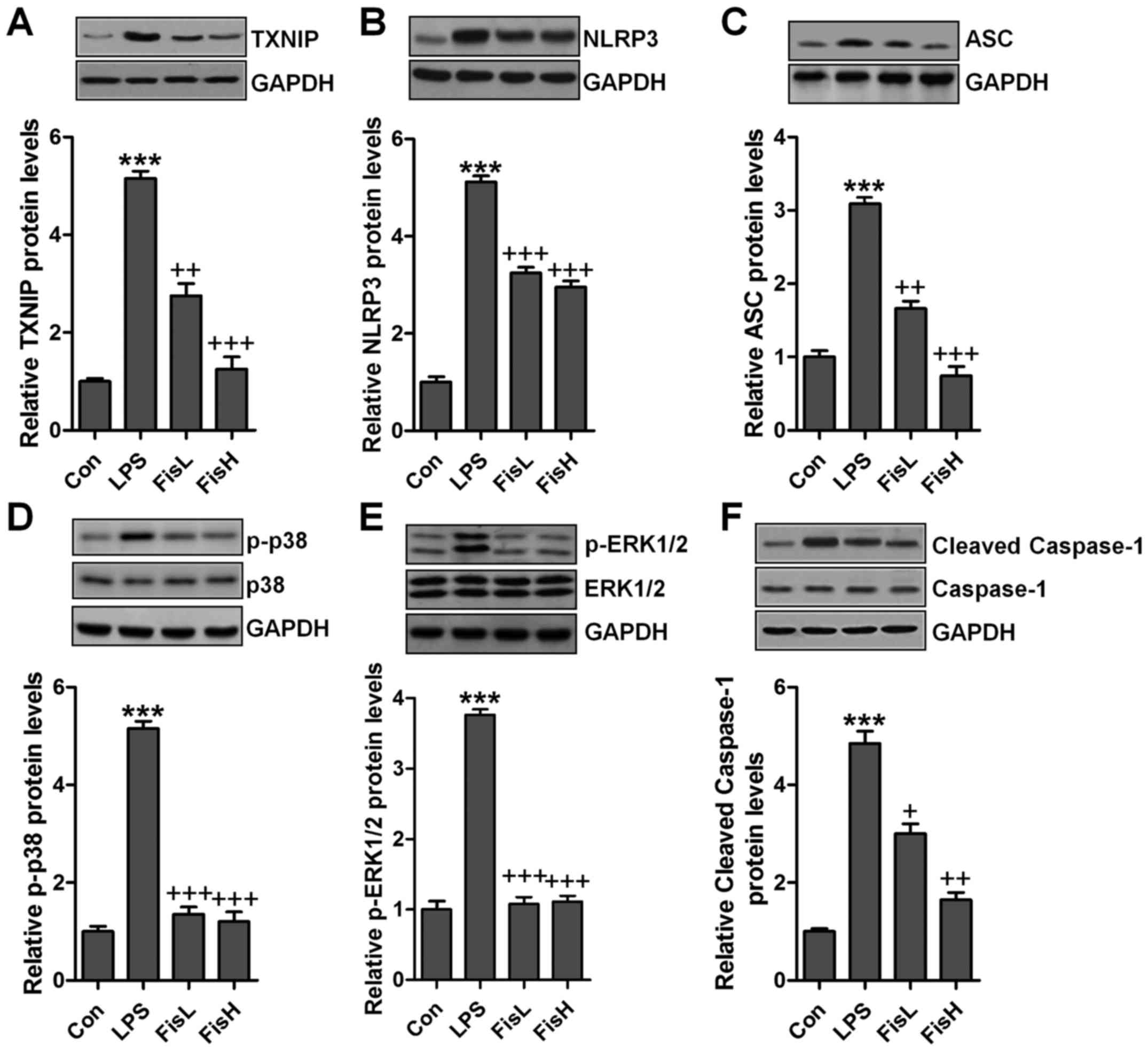
p38/ERK1/2 activation (26,27). Hence, western blot analysis was
used to calculate TXNIP/NLRP3 and MAPKs signaling pathways. As
shown in Fig. 6A and B, TXNIP and
NLRP3 protein expression levels were highly upregulated in LPS
exposure, leading to ROS generation largely. After fisetin
administration, TXNIP and NLRP3 were downregulated. TXNIP/NLRP3
signaling pathway activation is a key to induce ASC expression,
causing caspase-1 cleavage and enhancing NF-κB transcription to
induce pro-inflammatory cytokines secretion (28). Similarly, in this study, we found
that ASC was increased following TXNIP and NLRP3 upregulation in
LPS-treated group, which was reduced by fisetin addition (Fig. 6C). Subsequently, cleaved caspase-1
was also stimulated for LPS, reduced by fisetin administration
(Fig. 6D). Finally, MAPKs, ERK1/2
and p38, phosphorylated levels were measured through western blot
analysis. As shown in Fig. 6E and
F, we found that ERK1/2 and p38 phosphorylated levels were
highly stimulated in LPS-treated group, while in fisetin-treated
groups, both ERK1/2 and p38 phosphorylation were restrained.
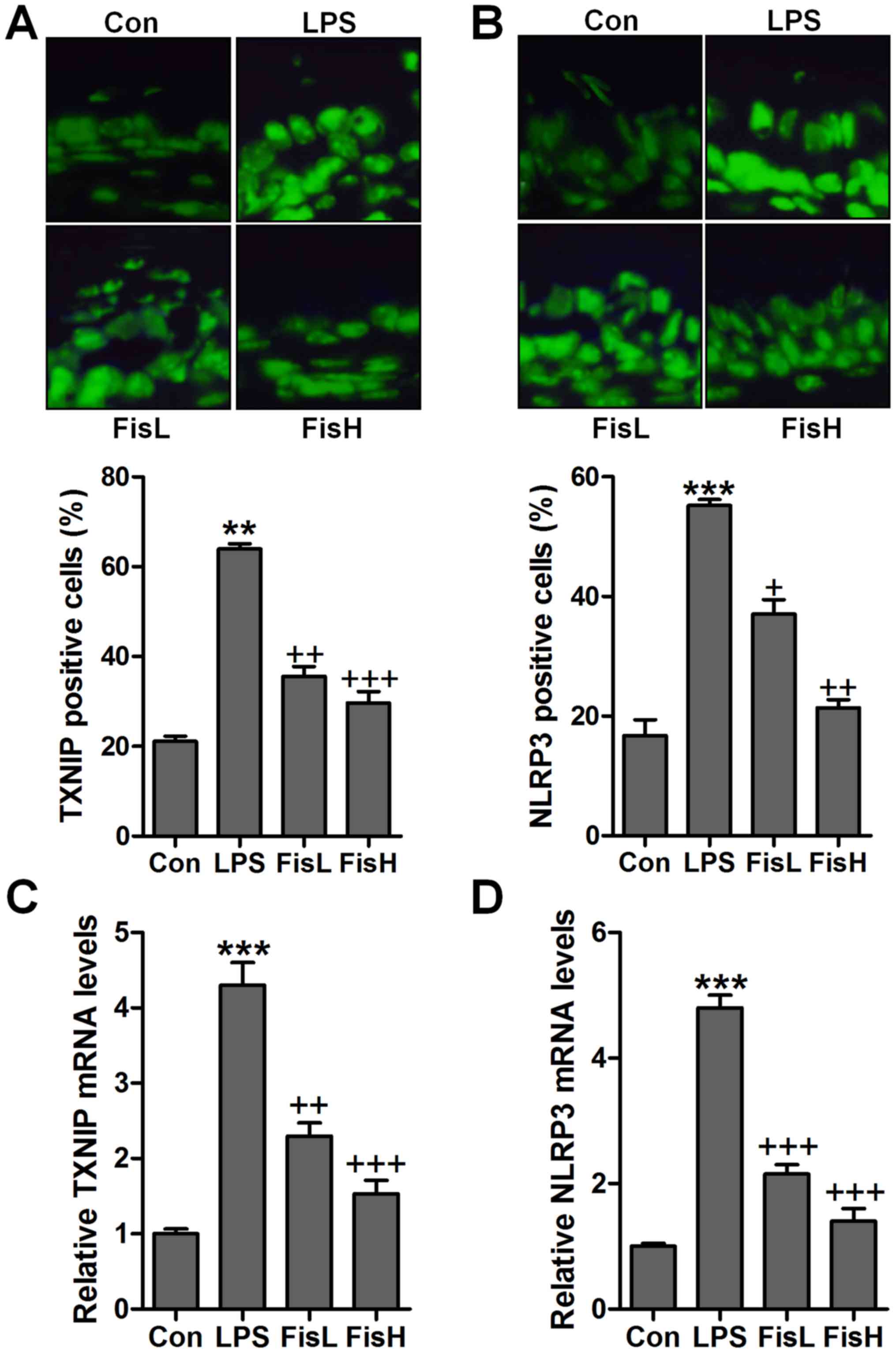
To further prove TXNIP and NLRP3 in acute otitis
media formation, immunofluorescent analysis was used to calculate
TXNIP and NLRP3 alterations in fisetin-treated mice induced by LPS.
Apparently high immunofluorescent intensity was observed in
LPS-treated group, which was weaken for fisetin treatment (Fig. 7A and B). Additionally, RT-qPCR
assays further indicated that TXNIP and NLRP3 mRNA levels were
reduced by LPS, reversed by fisetin and shown in a dose-dependent
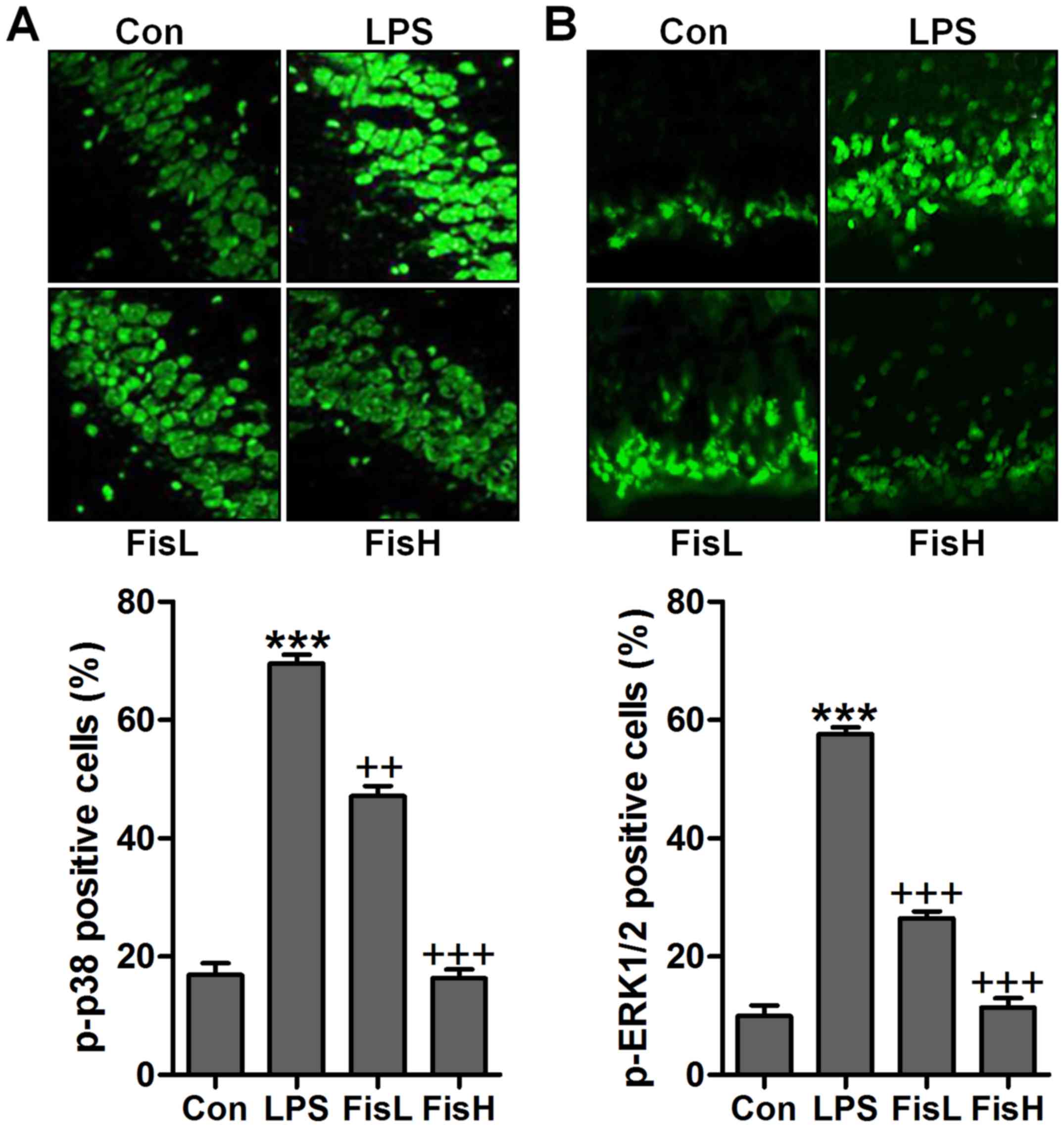
manner (Fig. 7C and D). Moreover,
p-p38 and p-ERK1/2 were expressed with extremely high fluorescent
intensity in the middle ear tissue of mice after LPS exposure,
which was blocked by fisetin administration (Fig. 8). The data indicated that
fisetin-improved acute otitis media induced by LPS was highly
related to TXNIP/NLRP3 and MAPKs signaling pathway repression.
Discussion
Acute otitis media is reported as one of the most
common infectious diseases for children. Acute otitis media could
develop into otitis media chronically, leading to hearing loss
(1,29). Severe complications include
language disability and intellectual impairment (30). However, until now the specific
molecular mechanism of acute otitis media is poorly known, and
finding effective therapeutic strategy is urgently required.
Fisetin has been reported to be effective in anti-inflammation and
anti-oxidation through various signaling pathways (31). A previous study found that acute
otitis media development has a close relationship with inflammatory
response and oxidative stress (32). Moreover, apoptosis is a cellular
process regulating different cell types, leading to cell death
eventually (33).
Apoptosis-induced cell dysfunction has been reported in a variety
of diseases, including acute lung injury, acute renal injury and
even the acute otitis media (34). LPS, as inflammatory reponse
inducer, is considered to be feasible for acute otitis media
establishment in mice (35). In
this study, LPS was used to induce acute otitis media in C57BL/6
mice by inflammation induction, apoptosis accumulation and ROS
generation through TLR4/NF-κB, caspase-3/PARP and TXNIP/NLRP3
signaling pathways. Of note, it was the first time that fisetin was
applied to ameliorate acute otitis media induced by LPS injection
into the middle ear of mice. The thickened mucosa confirmed the
successful establishment of acute otitis media in mice, indicating
the inflammatory severity. Importantly, fisetin showed activity to
reduce the thickened mucosa induced by LPS, which demonstrated that
fisetin could improve LPS-triggered acute otitis media in the
middle ear of mice.
Earlier studies have reported that fisetin
suppresses pro-inflammatory cytokine release is protective against
acute organ or tissue injuries (36,37). The suppressive role of fisetin in
acute otitis media could inhibit LPS-induced injury in mice.
Blocking pro-inflammatory cytokines secretion could be an effective
target for otitis media treatment. In order to investigate the
restraining of pro-inflammatory cytokines in fisetin-treated
groups, TNF-α, IL-1β, IL-6 and VEGF molecules were assessed through
ELISA method and RT-qPCR analysis. We found that pro-inflammatory
cytokines, TNF-α, IL-1β, IL-6 and VEGF were highly expressed in LPS
group, which was in line with the histological study. Fisetin
impeded secretion of these factors in a dose-dependent manner,
illustrating that fisetin could be a potential natural compound for
acute otitis media treatment through inhibiting pro-inflammatory
cytokine release.
TLR4/NF-κB has been well investigated in activating
inflammatory response (38). TLR4
once activated by LPS could stimulate its down-streaming signal
MyD88, leading to IKKα phosphorylation (39). Subsequently, IκBα and NF-κB
complex is disassociated through phosphorylation, contributing to
NF-κB translocation into the nucleus and leading to
pro-inflammatory cytokine release (40). Similarly, in this study, we found
that TLR4/NF-κB signaling pathway was highly activated by LPS
induction, which was suppressed due to fisetin administration. In
addition, TXNIP/NLRP3 is an important regulator for inflammation
response (41). TXNIP could bind
to NLRP3 inflammasome (42). Once
TXNIP/NLRP3 is activated under different conditions, NLRP3 is
ligated with ASC, which in turn combines to pro-caspase-1, leading
to its transformation to the cleaved caspase-1. Cleaved caspase-1
modulates the maturation of pro-inflammatory cytokines, including
IL-1β and IL-18 (43).
Previously, NLRP3 caused renal injury has been reported (44). We found that TXNIP and NLRP3
signaling pathway was upregulated for LPS treatment, followed by
ASC and caspase-1 cleavage increase, which was in line with the
inflammation response.
Apoptotic response is important in regulating
cellular process through inducing cell death (45). Caspase-3 activation induced PARP
cleavage, contributing to apoptosis and causing cell injury
(46). Bcl-2, belonging to
anti-apoptotic family, is known to suppress apoptosis. In contrast,
pro-apoptotic members, Bad and Bax, which also include in Bcl-2
family, are apoptosis inducers (47). Here, we found that apoptosis was
stimulated for LPS, resulting in cell death and acute otitis media
in the middle ear of mice eventually, which was conformed by
caspase-3 and PARP cleavage, as well as increased Bax and Bad. In
contrast, decreased Bcl-2 and Bcl-xL was observed. Fisetin
suppressed apoptosis through caspase-3 and PARP inactivation.
Furthermore, ROS generation is another cellular stress, leading to
cell injury and contributing to inflammation response (48). ROS production is highly related to
anti-oxidants and oxidant accumulation. Previous studies have
suggested that ROS over-production is a key point, causing injuries
under various situations, such as liver, heart and lung injury
(49). Further, acute otitis
media is reported to be associated with ROS levels. MAPKs, p38 and
ERK1/2, are reported to be of great importance in promoting ROS
generation (50). It has been
reported that MAPKs activate the pathological process of various
diseases (51). In this study,
MAPK signaling pathway was activated, accompanied with high
expression of phosphorylated p38 and ERK1/2 after LPS stimulation,
which was reduced by fisetin administration, indicating that
fisetin-improved acute otitis media is related to MAPK
suppression.
Our results indicated that pro-inflammatory cytokine
levels, apoptosis and ROS generation were significantly upregulated
in acute otitis media mouse model induced by LPS. Fisetin
significantly reduced the levels of pro-inflammatory cytokines,
apoptosis and ROS. Therefore, fisetin is a promising therapeutic
strategy for the treatment of acute otitis media.
Acknowledgments
The authors would like to thank all the members of
the Department of Otorhinolaryngology for helping in this work.
References
|
1
|
Morris LM, DeGagne JM, Kempton JB, Hausman
F and Trune DR: Mouse middle ear ion homeostasis channels and
intercellular junctions. PLoS One. 7:e390042012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Darrow DH, Dash N and Derkay CS: Otitis
media: Concepts and controversies. Curr Opin Otolaryngol Head Neck
Surg. 11:416–423. 2003. View Article : Google Scholar
|
|
3
|
Bluestone CD and Klein JO: Physiology,
pathophysiology and pathogenesis. Otitis Media in Infants and
Children (4th edition). Decker BC: 41–42. 2007.
|
|
4
|
Post JC: Direct evidence of bacterial
biofilms in otitis media. Laryngoscope. 111:2083–2094. 2001.
View Article : Google Scholar
|
|
5
|
Topcuoglu N, Keskin F, Ciftci S, Paltura
C, Kulekci M, Ustek D and Kulekci G: Relationship between oral
anaerobic bacteria and otitis media with effusion. Int J Med Sci.
9:256–261. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hebda PA, Piltcher OB, Swarts JD, Alper
CM, Zeevi A and Doyle WJ: Cytokine profiles in a rat model of
otitis media with effusion caused by eustachian tube obstruction
with and without Streptococcus pneumoniae infection. Laryngoscope.
112:1657–1662. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bouchet V, Hood DW, Li J, Brisson JR,
Randle GA, Martin A, Li Z, Goldstein R, Schweda EK, Pelton SI,
Richards JC, et al: Host-derived sialic acid is incorporated into
Haemophilus influenzae lipopolysaccharide and is a major virulence
factor in experimental otitis media(J). Proc Natl Acad Sci USA.
100:8898–8903. 2003. View Article : Google Scholar
|
|
8
|
Gallucci RM, Sugawara T, Yucesoy B,
Berryann K, Simeonova PP, Matheson JM and Luster MI: Interleukin-6
treatment augments cutaneous wound healing in immunosuppressed
mice. J Interferon Cytokine Res. 21:603–609. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Meng Z, Yan C, Deng Q, Gao DF and Niu XL:
Curcumin inhibits LPS-induced inflammation in rat vascular smooth
muscle cells in vitro via ROS-relative TLR4-MAPK/NF-κB pathways.
Acta Pharmacol Sin. 34:901–911. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schwarz JM and Bilbo SD: LPS elicits a
much larger and broader inflammatory response than Escherichia coli
infection within the hippocampus of neonatal rats. Neurosci Lett.
497:110–115. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Niemi K, Teirilä L, Lappalainen J,
Rajamäki K, Baumann MH, Öörni K, Wolff H, Kovanen PT, Matikainen S
and Eklund KK: Serum amyloid A activates the NLRP3 inflammasome via
P2X7 receptor and a cathepsin B-sensitive pathway. J Immunol.
186:6119–6128. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gicquel T, Victoni T, Fautrel A, Robert S,
Gleonnec F, Guezingar M, Couillin I, Catros V, Boichot E and
Lagente V: Involvement of purinergic receptors and NOD-like
receptor-family protein 3-inflammasome pathway in the adenosine
triphosphate-induced cytokine release from macrophages. Clin Exp
Pharmacol Physiol. 41:279–286. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schmitz ML, Bacher S and Kracht M: I kappa
B-independent control of NF-kappa B activity by modulatory
phosphorylations. Trends Biochem Sci. 26:186–190. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hsieh PF, Hou CW, Yao PW, Wu SP, Peng YF,
Shen ML, Lin CH, Chao YY, Chang MH and Jeng KC: Sesamin ameliorates
oxidative stress and mortality in kainic acid-induced status
epilepticus by inhibition of MAPK and COX-2 activation. J
Neuroinflammation. 8:572011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Murtaza I, Adhami VM, Hafeez BB, Saleem M
and Mukhtar H: Fisetin, a natural flavonoid, targets chemoresistant
human pancreatic cancer AsPC-1 cells through DR3-mediated
inhibition of NF-kappaB. Int J Cancer. 125:2465–2473. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li J, Cheng Y, Qu W, Sun Y, Wang Z, Wang H
and Tian B: Fisetin, a dietary flavonoid, induces cell cycle arrest
and apoptosis through activation of p53 and inhibition of NF-kappa
B pathways in bladder cancer cells. Basic Clin Pharmacol Toxicol.
108:84–93. 2011. View Article : Google Scholar
|
|
17
|
Ying TH, Yang SF, Tsai SJ, Hsieh SC, Huang
YC, Bau DT and Hsieh YH: Fisetin induces apoptosis in human
cervical cancer HeLa cells through ERK1/2-mediated activation of
caspase-8-/caspase-3-dependent pathway. Arch Toxicol. 86:263–273.
2012. View Article : Google Scholar
|
|
18
|
Stol K, Diavatopoulos DA, Graamans K,
Engel JA, Melchers WJ, Savelkoul HF, Hays JP, Warris A and Hermans
PW: Inflammation in the middle ear of children with recurrent or
chronic otitis media is associated with bacterial load. Pediatr
Infect Dis J. 31:1128–1134. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tateossian H, Morse S, Parker A, Mburu P,
Warr N, Acevedo-Arozena A, Cheeseman M, Wells S and Brown SD:
Otitis media in the Tgif knockout mouse implicates TGFβ signalling
in chronic middle ear inflammatory disease. Hum Mol Genet.
22:2553–2565. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ferenbach D, Kluth DC and Hughes J:
Inflammatory cells in renal injury and repair. Semin Nephrol.
27:250–259. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Thieringer R, Fenyk-Melody JE, Le Grand
CB, Shelton BA, Detmers PA, Somers EP, Carbin L, Moller DE, Wright
SD and Berger J: Activation of peroxisome proliferator-activated
receptor gamma does not inhibit IL-6 or TNF-alpha responses of
macrophages to lipopolysaccharide in vitro or in vivo. J Immunol.
164:1046–1054. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chávez-Sánchez L, Garza-Reyes MG,
Espinosa-Luna JE, Chávez-Rueda K, Legorreta-Haquet MV and
Blanco-Favela F: The role of TLR2, TLR4 and CD36 in macrophage
activation and foam cell formation in response to oxLDL in humans.
Hum Immunol. 75:322–329. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang W, Zhou A, Zhang X, Xiang Y, Huang Y,
Wang L, Zhang S, Liu Y, Yin Y and He Y: Interleukin 17A promotes
pneumococcal clearance by recruiting neutrophils and inducing
apoptosis through a p38 mitogen-activated protein kinase-dependent
mechanism in acute otitis media. Infect Immun. 82:2368–2377. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Brentnall M, Rodriguez-Menocal L, De
Guevara RL, Cepero E and Boise LH: caspase-9, caspase-3 and
caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell
Biol. 14:322013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Park J, Min JS, Kim B, Chae UB, Yun JW,
Choi MS, Kong IK, Chang KT and Lee DS: Mitochondrial ROS govern the
LPS-induced pro-inflammatory response in microglia cells by
regulating MAPK and NF-κB pathways. Neurosci Lett. 584:191–196.
2015. View Article : Google Scholar
|
|
26
|
Li W, Wu Z, Ma Q, Liu J, Xu Q, Han L, Duan
W, Lv Y, Wang F, Reindl KM, et al: Hyperglycemia regulates
TXNIP/TRX/ROS axis via p38 MAPK and ERK pathways in pancreatic
cancer. Curr Cancer Drug Targets. 14:348–356. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cao G, Jiang N, Hu Y, Zhang Y, Wang G, Yin
M, Ma X, Zhou K, Qi J, Yu B, et al: Ruscogenin attenuates cerebral
ischemia-induced blood-brain barrier dysfunction by suppressing
TXNIP/NLRP3 inflammasome activation and the MAPK pathway. Int J Mol
Sci. 17:14182016. View Article : Google Scholar :
|
|
28
|
Jiang L, Fei D, Gong R, Yang W, Yu W, Pan
S and Zhao M and Zhao M: CORM-2 inhibits TXNIP/NLRP3 inflammasome
pathway in LPS-induced acute lung injury. Inflamm Res. 65:905–915.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chu TG, Cachola DR III, Regal MA, Llamas
AC, Martinez NV and Santos WR: Pneumococcal conjugate vaccine
(Non-Typeable Haemophilus influenzae (NTHi) protein D, diphtheria
or tetanus toxoid conjugates) in prevention of acute otitis media
in children: A Cohort Study. Philipp J Otolaryngol Head Neck Surg.
31:12016.
|
|
30
|
Zhang J, Xu M, Zheng Q, Zhang Y, Ma W and
Zhang Z: Blocking macrophage migration inhibitory factor activity
alleviates mouse acute otitis media in vivo. Immunol Lett.
162:101–108. 2014. View Article : Google Scholar :
|
|
31
|
Yang PM, Tseng HH, Peng CW, Chen WS and
Chiu SJ: Dietary flavonoid fisetin targets caspase-3-deficient
human breast cancer MCF-7 cells by induction of
caspase-7-associated apoptosis and inhibition of autophagy. Int J
Oncol. 40:469–478. 2012.
|
|
32
|
Hafré L, Einarsdottir E, Kentala E,
Hammarén-Malmi S, Bhutta MF, MacArthur CJ, Wilmot B, Casselbrant M,
Conley YP, Weeks DE, et al: Predisposition to childhood otitis
media and genetic polymorphisms within the toll-like receptor 4
(TLR4) locus. PLoS One. 10:e01325512015. View Article : Google Scholar
|
|
33
|
Alladi PA, Roy T, Singh N and Wadhwa S:
Prenatal auditory enrichment with species-specific calls and sitar
music modulates expression of Bcl-2 and Bax to alter programmed
cell death in developing chick auditory nuclei. Int J Dev Neurosci.
23:363–373. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li X, Luo R, Jiang R, Meng X, Wu X, Zhang
S and Hua W: The role of the Hsp90/Akt pathway in myocardial
calpain-induced caspase-3 activation and apoptosis during sepsis.
BMC Cardiovasc Disord. 13:82013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Costa SS, Silva MNL, Rosito LPS and
Selaimen FA: One case, two lessons: An aberrant internal carotid
artery causing acquired cholesteatoma. Braz J Otorhinolaryngol.
80:453–454. 2014.In Portuguese. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ha T, Xia Y, Liu X, Lu C, Liu L, Kelley J,
Kalbfleisch J, Kao RL, Williams DL and Li C: Glucan phosphate
attenuates myocardial HMGB1 translocation in severe sepsis through
inhibiting NF-κB activation. Am J Physiol Heart Circ Physiol.
301:H848–H855. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Singh AK, Jiang Y, Gupta S, Younus M and
Ramzan M: Anti-inflammatory potency of nano-formulated puerarin and
curcumin in rats subjected to the lipopolysaccharide-induced
inflammation. J Med Food. 16:899–911. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lin J, Wang H, Li J, Wang Q, Zhang S, Feng
N, Fan R and Pei J: κ-Opioid receptor stimulation modulates
TLR4/NF-κB signaling in the rat heart subjected to
ischemia-reperfusion. Cytokine. 61:842–848. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ridder DA and Schwaninger M: NF-kappaB
signaling in cerebral ischemia. Neuroscience. 158:995–1006. 2009.
View Article : Google Scholar
|
|
40
|
Fan H, Li L, Zhang X, Liu Y, Yang C, Yang
Y and Yin J: Oxymatrine downregulates TLR4, TLR2, MyD88, and
NF-kappaB and protects rat brains against focal ischemia. Mediators
Inflamm. 2009:7047062009. View Article : Google Scholar
|
|
41
|
Elgort MG, O'Shea JM, Jiang Y and Ayer DE:
Transcriptional and translational downregulation of thioredoxin
interacting protein is required for metabolic reprogramming during
G(1). Genes Cancer. 1:893–907. 2010. View Article : Google Scholar
|
|
42
|
Zhou J, Yu Q and Chng WJ: TXNIP (VDUP-1,
TBP-2): A major redox regulator commonly suppressed in cancer by
epigenetic mechanisms. Int J Biochem Cell Biol. 43:1668–1673. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tschopp J and Schroder K: NLRP3
inflammasome activation: The convergence of multiple signalling
pathways on ROS production? Nat Rev Immunol. 10:210–215. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ye W, Lei Y, Yu M, Xu Y, Cao M, Yu L,
Zhang L, Li P, Bai W and Xu Z: RP3 inflammasome is responsible for
Hantavirus inducing interleukin-1β in THP-1 cells. Int J Mol Med.
35:1633–1640. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Faber AC, Coffee EM, Costa C, Dastur A,
Ebi H, Hata AN, Yeo AT, Edelman EJ, Song Y, Tam AT, et al: mTOR
inhibition specifically sensitizes colorectal cancers with KRAS or
BRAF mutations to BCL-2/BCL-XL inhibition by suppressing MCL-1.
Cancer Discov. 4:42–52. 2014. View Article : Google Scholar :
|
|
46
|
Hata AN, Yeo A, Faber AC, Lifshits E, Chen
Z, Cheng KA, Walton Z, Sarosiek KA, Letai A, Heist RS, et al:
Failure to induce apoptosis via BCL-2 family proteins underlies
lack of efficacy of combined MEK and PI3K inhibitors for
KRAS-mutant lung cancers. Cancer Res. 74:3146–3156. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
He L, Torres-Lockhart K, Forster N,
Ramakrishnan S, Greninger P, Garnett MJ, McDermott U, Rothenberg
SM, Benes CH and Ellisen LW: Mcl-1 and FBW7 control a dominant
survival pathway underlying HDAC and Bcl-2 inhibitor synergy in
squamous cell carcinoma. Cancer Discov. 3:324–337. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Haroon ZA, Raleigh JA, Greenberg CS and
Dewhirst MW: Early wound healing exhibits cytokine surge without
evidence of hypoxia. Ann Surg. 231:137–147. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Indran IR, Hande MP and Pervaiz S: hTERT
overexpression alleviates intracellular ROS production, improves
mitochondrial function, and inhibits ROS-mediated apoptosis in
cancer cells. Cancer Res. 71:266–276. 2011. View Article : Google Scholar
|
|
50
|
Genestra M: Oxyl radicals, redox-sensitive
signalling cascades and antioxidants. Cell Signal. 19:1807–1819.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hsin YH, Chen CF, Huang S, Shih TS, Lai PS
and Chueh PJ: The apoptotic effect of nanosilver is mediated by a
ROS- and JNK-dependent mechanism involving the mitochondrial
pathway in NIH3T3 cells. Toxicol Lett. 179:130–139. 2008.
View Article : Google Scholar : PubMed/NCBI
|