Introduction
Chronic rhinosinusitis (CRS) can be divided into two
subtypes: CRS without nasal polyps (CRSsNP) and CRS with nasal
polyps (CRSwNP). CRSwNP is a heterogeneous disease with an unclear
pathophysiology (1). CRSwNP is
subdivided into different endotypes by inflammatory markers and/or
cells that serve a role in the disease (1). One theory about the maintenance of
the inflammatory reaction is a variation of T cells within the
polypoid tissue (2). In a
previous study (2), cluster of
differentiation (CD)4+ and CD8+ T cell
subsets were characterized by multicolor flow cytometry, which
revealed a predominance of CD8+ T cells in nasal polyps
compared with the peripheral blood mononuclear cells (PBMCs) in
patients with CRSwNP. There was a significantly higher amount of
CD8+ T cells compared with CD4+ T cells in
nasal polyps, whereas there were significantly more CD4+
T cells compared with CD8+ T cells in the PBMCs
(2). These data suggest a local
regulation of the immune response within nasal polyps. Furthermore,
both CD4+ and CD8+ T cells were able to
differentiate into an effector memory phenotype. It was postulated
that variations in regulatory T cells are responsible for a number
of autoimmune diseases (3). A
previous study reported a significant increase in activated
regulatory T cells (Treg) in polypoid tissue compared
with the PBMCs in patients with CRSwNP (2). Specific triggers, including fungal
colonization (4–6) or Staphylococcal superantigens
(7,8) influence T cell recruitment in CRSwNP
and may also influence T cell subset composition within the polyps.
Additionally, local changes in B cell subpopulations in the nasal
polyps compared with PBMCs has been reported and underlines the
role of lymphocytes in this disease (9).
Memory T cells represent the main subset of
CD4+ and CD8+ T cells in polypoid tissue in
patients with CRSwNP (2). These
memory T cells can be classified into two subsets based on the
expression of homing receptors, including C-C chemokine receptor 7
(CCR7) (10), into
CCR7+ central memory T cells (Tcm) and
CCR7-effector memory T cells (Tem). After antigen
presentation and differentiation into Tem, T cells
migrate towards non-lymphoid tissue (NLT). It was previously
assumed that these T cells recirculate into the PBMCs; however,
immunological studies have reported persistent populations of
tissue-resident memory T cells (Trm) in NLT (11). These T cells may be identified by
a high expression of CD69 and a downregulation of the sphingosine-1
phosphate receptor 1 (S1PR1). S1PR1 is required for naive T cells
to circulate and exit the thymus and peripheral lymphoid organs
(11). S1PR1 downregulation is an
essential marker for Trm (12). In contrast, CD69 upregulation is a
major signal for the persistence of Trm in NLT (13). Both signals are necessary for the
persistence of Trm (14) in the local tissue. Chemoattractant
receptors, including CCR7, also serve a role in the egress of T
cells from NLT (15), therefore a
downregulation in CCR7 is a sign that T cells persist in the tissue
and do not recirculate. These Trm cells are described as
being more potent in protection against local infections compared
with memory T cells residing elsewhere (16). Furthermore, Trm have
been reported to serve a role in drug hypersensitivity reactions
(17).
The aim of the present study was to quantify the
number of CD4+ and CD8+ Trm cells
in the nasal polyps compared with PBMCs in patients with CRSwNP and
to determine whether there were differences between these
subpopulations.
Materials and methods
Ethical approval
The study was approved by the Ethics Board of the
Medical Faculty, Julius-Maximilian-University, Wuerzburg, Germany.
Ethics approval and written informed consent was obtained from all
patients.
Preparation of human lymphocytes
A total of 10 ml of heparinized blood samples were
obtained intraoperatively by venous puncture from 10 patients
undergoing paranasal sinus surgery between July and October 2017.
All patients received intranasal topical steroids (mometasone
furoate nasal spray, 50 μg/puff) prior to surgery. Patients
were recruited from the Department of Otorhinolaryngology, Plastic,
Aesthetic and Reconstructive Head and Neck Surgery of the
University of Wuerzburg (Wuerzburg, Germany). Patients with
Churg-Strauss syndrome, primary ciliary dyskinesia or cystic
fibrosis were excluded. Patient characteristics are summarized in
Table I. Lymphocytes were
separated by density-gradient centrifugation for 10 min at 1,000 ×
g at room temperature with equal amounts of Ficoll (Biochrom GmbH,
Berlin, Germany), using a membrane-containing 10 ml cell tube
(Greiner Bio-One, Kremsmünster, Austria). Tubes were washed twice
with PBS and the cell number and viability were determined using a
Cell Counter + Analyzer System (CASY TT; Innovatis Technologies,
Inc., Fairfax, VA, USA) according to the manufacturer's protocol.
Following centrifugation at 500 × g at 20°C for 5 min, the cells
were frozen at −80°C with 1 ml freezing medium, which contained 10
parts of fetal bovine serum (Linaris Biologische Produkte GmbH,
Dossenheim, Germany) and one part dimethylsulfoxide.
 | Table IBaseline characteristics of the study
group. |
Table I
Baseline characteristics of the study
group.
| Clinical
feature | Study group
(n=10) |
|---|
| Age, years
(standard deviation) | 44 (9.48) |
| Sex,
female/male | 2/8 |
| Previous surgery, n
(%) | 2 (20) |
| Eosinophilic
polyps, n (%) | 7 (70) |
| Allergy, n (%) | 4 (40) |
| Samter's triad, n
(%) | 1 (10) |
Preparation of tissue samples
All tissue samples were collected intraoperatively
from 10 patients undergoing regular paranasal sinus surgery due to
CRSwNP. Additionally, nasal mucosa was collected from 3 patients
diagnosed with CRSsNP undergoing paranasal sinus surgery between
August and September 2016 at the local university. Exclusion
criteria were as above. All patients were female and mean age was
45.33±17.44. The polyps and nasal mucosa samples were cut into
small fragments and mashed through a cell strainer (Greiner
Bio-One) from 100 to 40 μm in PBS. Tissues were washed twice
in PBS and the cell number and viability were determined using a
CASY TT system according to the manufacturer's protocol. Following
centrifugation for 5 min at, 1,600 rpm, cells were frozen at −80°C
in 1 ml freezing medium.
Fluorescence-activated cell sorting
The following antibodies were used: Anti-CD45
Pacific Orange (1:300; MHCD4530; Thermo Fisher Scientific Inc.,
Waltham, MA, USA), anti-CD3 phycoerythrin (PE)-Cy7 (1:300; 300420;)
anti-CD4 Pacific Blue (1:50, Nr. 300521), anti-CD8a Alexa 700
(1:50, 301028) anti-CD45RA peridinin chlorophyll protein
complex-Cy5.5 (1:50; 304122), anti-CCR7 Alexa488 (1:80; 353206),
anti-CD69 Alexa 488 (1:50, Nr. 310916), anti CD69 allophycocyanin
(APC; 1:50; 310909), anti-CD4 fluorescein isothiocyanate (1:40;
300506), anti-FoxP3 Pacific Blue (1:25; 320216) anti-CD52 (CTLA-4)
PE (1:400;349906; all BioLegend, Inc., San Diego, CA, USA) and
anti-S1PR1 eFluor 660 (1:20; 50-3639-41; eBioscience; Thermo Fisher
Scientific, Inc.). Isotype control staining was performed using
mouse-IgG APC (1:80; 137214) and mouse-IgG PE (1:25; 400140)
(BioLegend, Inc.). Viability Dye 780 (1:10; 65-0865-14;
eBioscience; Thermo Fisher Scientific, Inc.) was used to detect
apoptotic and dead cells. Following blocking with 25 μg/ml
normal mouse immunoglobulin G (1:50, Nr. I5381, Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) for 15 min on ice, all cells
underwent cell surface staining on ice for 30 min, followed by
intracellular staining. For intracellular staining of Foxp3 and
CTLA-4, cells were treated with fixation buffer (eBioscience;
Thermo Fisher Scientific, Inc.) for 30 min at room temperature.
Permeabilisation buffer was subsequently applied (eBioscience;
Thermo Fisher Scientific, Inc.) followed by staining with
anti-Foxp3 and anti-CTLA-4 for 45 min at room temperature. All
antibodies were used according to the manufacturer's protocol. FACS
analysis was performed using an LSR II flow cytometer and the data
were analyzed using FlowJo software (FlowJo LLC, Ashland, OR,
USA).
Statistical analysis
Data are presented as mean ± standard deviation.
Statistical significance was analyzed by a two-tailed paired t-test
using GraphPad Prism Software 6.0c (GraphPad Software, Inc., La
Jolla, CA, USA). For non-parametric distribution the Wilcoxon test
was applied. P<0.05 was considered to indicate a statistically
significant difference.
Results
Patient characteristics
A total of 10 patients with CRSwNP were included in
the present study (8 male and 2 female). The mean age was 45±9.48
years. Eosinophilic polyposis was described in the histological
evaluation of most of the patients (7/10). Patient characteristics
are summarized in Table I.
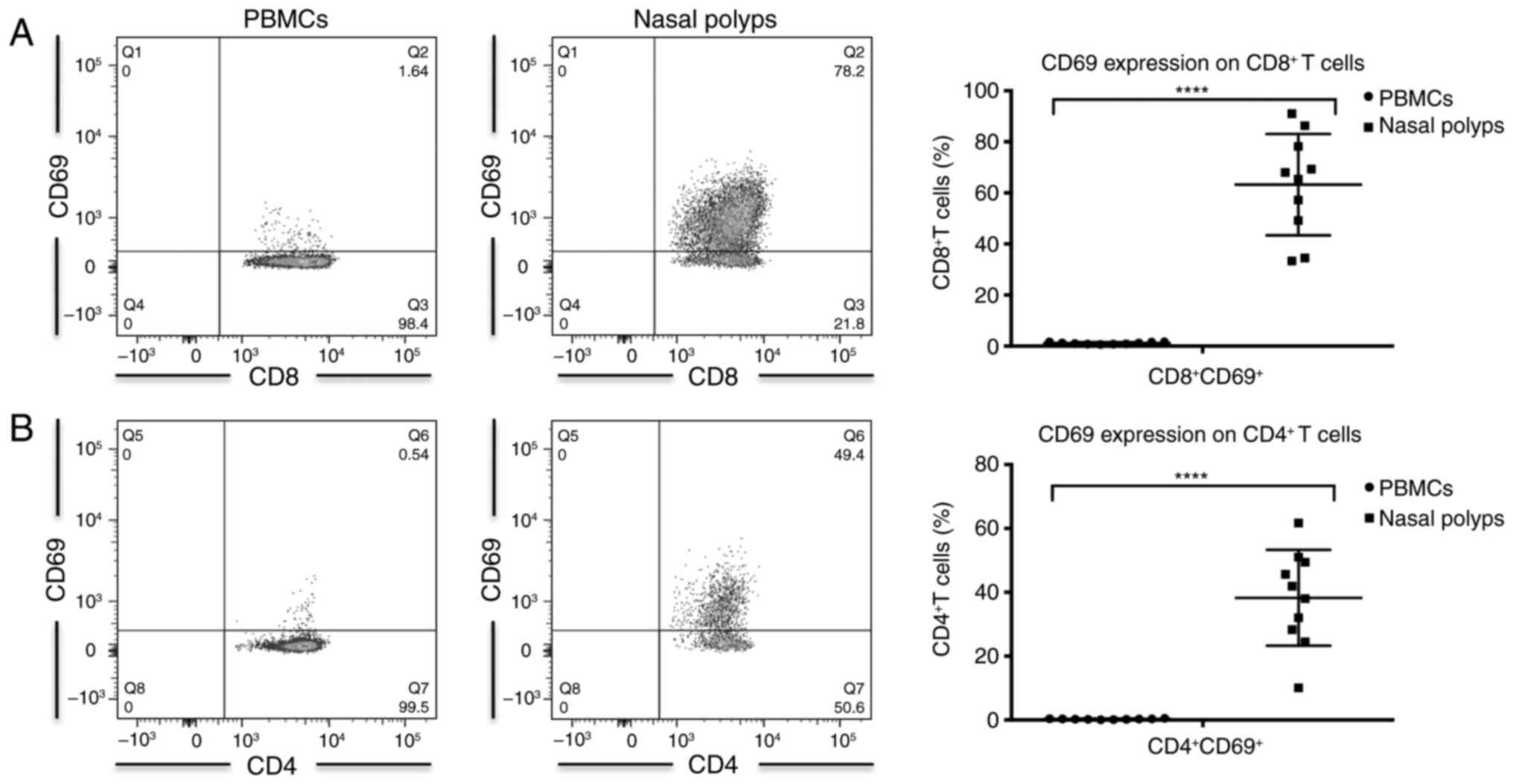
Higher frequency of CD69+
cells in CD4+ and CD8+ T cells in CRSwNP
The amount of CD3+ CD4+ T
cells among CD45+ leukocytes was significantly higher in
PBMCs compared with in nasal polyps from patients with CRSwNP
(Table II). In contrast,
CD3+ CD8+ T cells were significantly
increased in nasal polyps compared with PBMCs (Table II). A significant increase in the
frequency of CD69-expressing cells was observed among
CD3+ CD4+ and CD3+ CD8+
T cells in the nasal polyps compared with PBMCs (Table II; Fig. 1). However, these cells did not
constitute recently activated T cells, as T cells from nasal polyps
do not express elevated levels of human leukocyte antigen-antigen D
related (HLA-DR) compared with PBMCs (2). In PBMCs, a majority of cells were
CD69− (Table II;
Fig. 1). The percentage of
CD69+ cells was significantly higher among
CD8+ compared with CD4+ T cells in PBMCs and
polyps (Table II).
| Table IIComparison of CD4+ and
CD8+ Trm cells in patients with CRSwNP. |
Table II
Comparison of CD4+ and
CD8+ Trm cells in patients with CRSwNP.
| T cells | PBMCs | Nasal polyps | P-value |
|---|
| CD3+
CD4+ T cells | 37.47±10.18 | 20.67±8.71 | 0.0002 |
| CD4+
CD69+ Trm | 0.27±0.13 | 38.25±14.23 | <0.0001 |
| CD4+
CD69+ S1PR1− Trm | 0.28±0.13 | 23.61±15.26 | 0.0013 |
| CD4+
CCR7− Trm | 66.06±20.21 | 85.16±14.29 | 0.0093 |
| CD4+
S1PR1+ T cells | 4.56±6.46 | 33.82±27.98 | 0.0098 |
| CD3+
CD8+ T cells | 23.7±7.24 | 40.2±15.6 | 0.0089 |
| CD8+
CD69+ Trm | 1.14±0.36 | 63.24±18.83 | <0.0001 |
| CD8+
CD69+ S1PR1− Trm | 0.95±0.23 | 35.36±23.57 | 0.0017 |
| CD8+
CCR7− Trm | 79.04±13.43 | 97.18±5.48 | 0.0018 |
| CD8+
S1PR1+ T cells | 9.51±13.44 | 38.08±31.52 | 0.0488 |
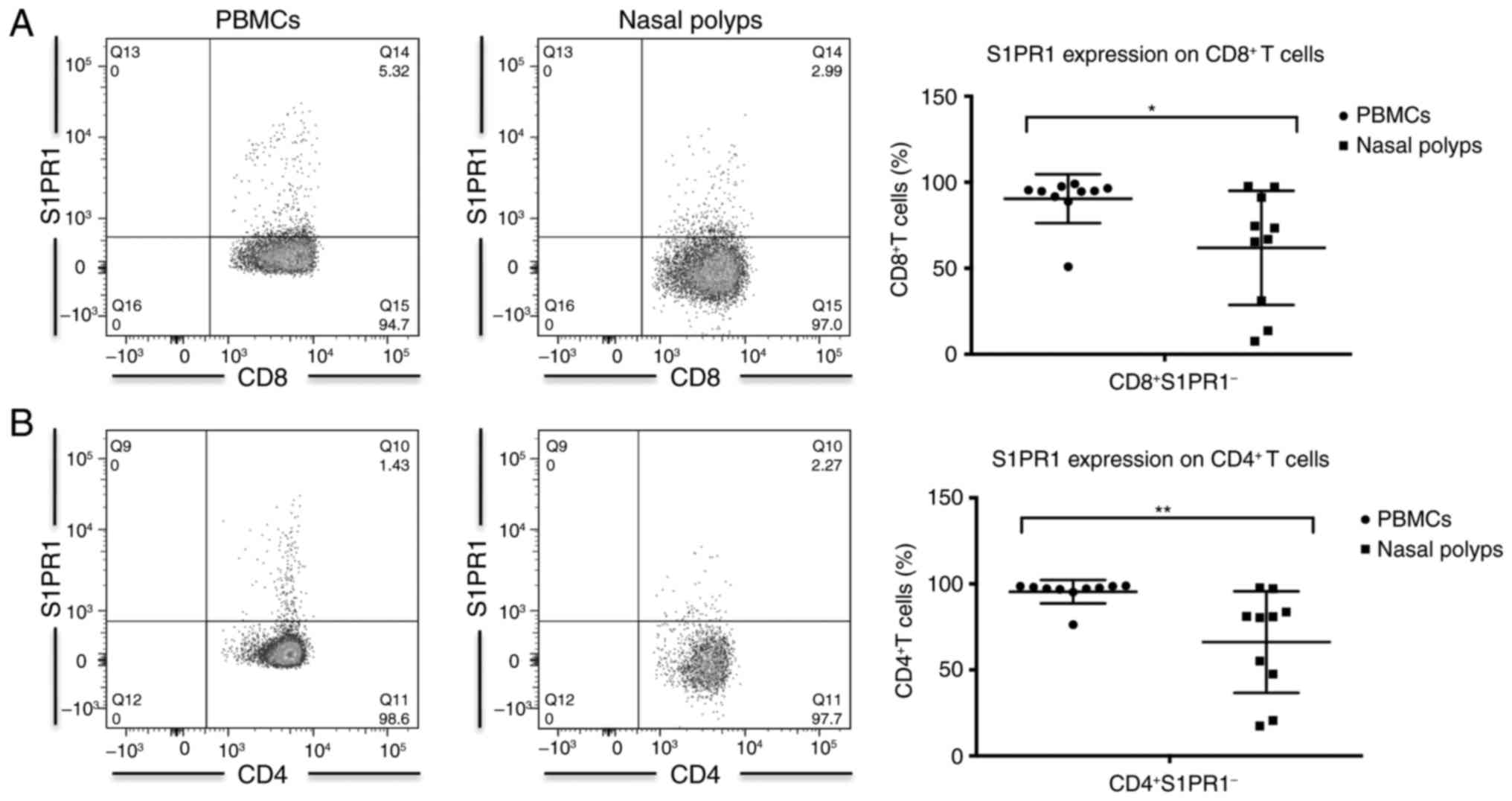
S1PR1 expression in CD4+ and
CD8+ T cells in CRSwNP
The percentage of S1PR1+ between
CD4+ and CD8+ T cells was significantly
higher in nasal polyps compared with PBMCs (Table II; Fig. 2). This is most likely attributable
to down modulation of S1PR1 expression by its ligand S1P, which is
abundantly present in PBMCs (18). In PBMCs the proportion of cells
expressing S1PR1 was significantly higher among CD8+ T
cells compared with CD4+ T cells, whereas no significant
difference in S1PR1 expression was observed between CD8+
and CD4+ T cells in nasal polyps (Table II).
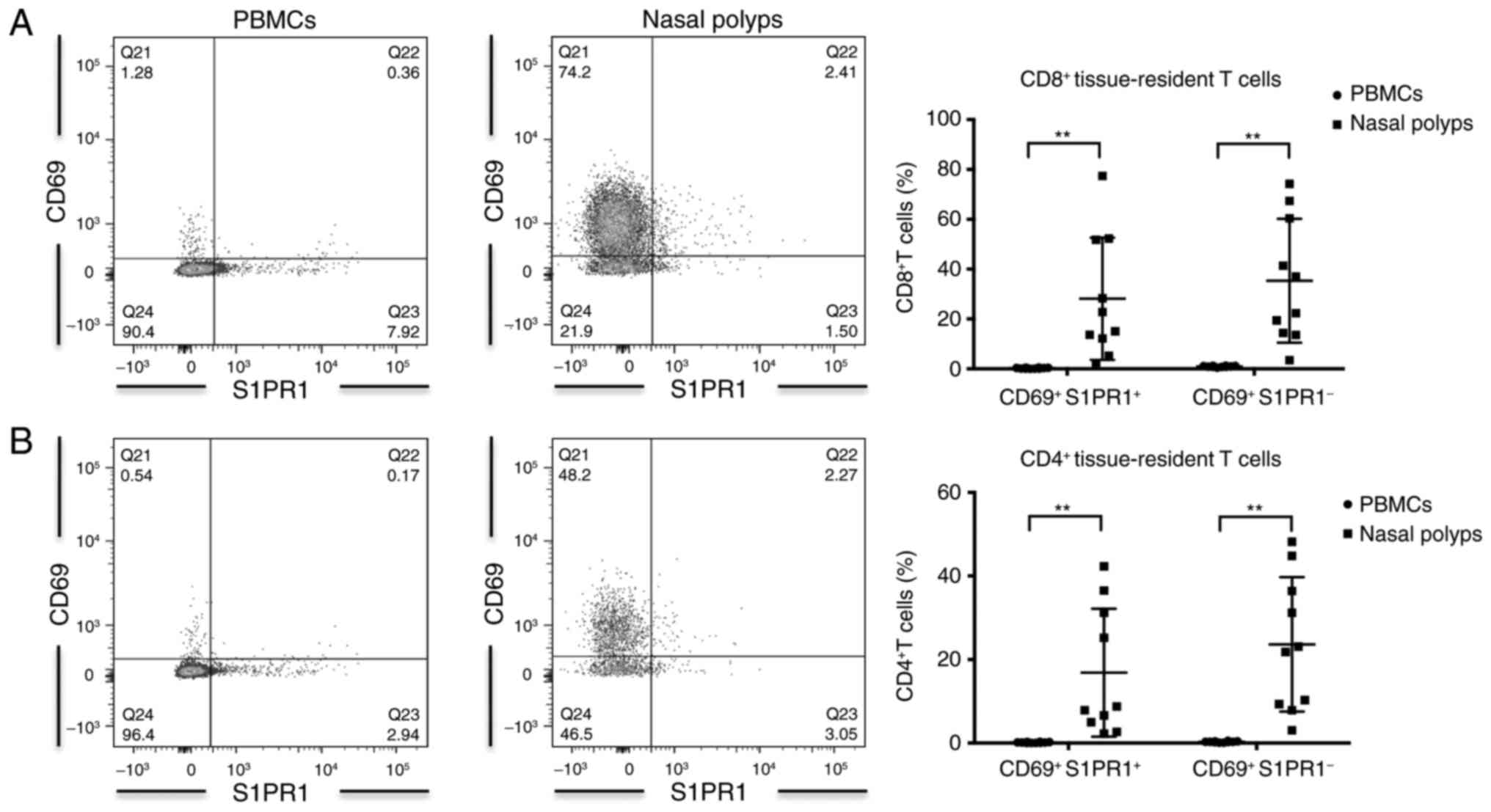
CD69+ S1PR1−
Trm was significantly increased in nasal polyps compared
with PBMCs
In patients with CRSwNP, the frequency of
CD69+ S1PR1− Trm in
CD4+ and CD8+ T cells was significantly
higher in nasal polyps compared with PBMCs (Table II; Fig. 3). CD8+ T cells
contained more CD69+ S1PR1− Trm
compared with CD4+ T cells, irrespective of the
anatomical compartment analyzed (Table II).
CD69 overexpression, activated
Treg (aTreg) and conventional memory T cells
(Tconv) in CRSwNP
Further analysis of CD4+ T cell subsets
with respect to CD45RA and FoxP3 expression revealed significantly
more CD3+ CD4+ CD45RA+
FoxP3+ naïve T cells in PBMCs compared with nasal polyps
in patients with CRSwNP (Table
III). However, CD69 expression was significantly higher in
phenotypically naïve CD4+ T cells in nasal polyps
compared with PBMCs (Table III;
Fig. 4). The number of
CD45RA− FoxP3− conventional Tconv
cells was significantly higher among CD4+ T cells in
nasal polyps with significantly higher expression of CD69 in these
cells compared with PBMCs (Table
III; Fig. 4). The proportion
of CD45RA− FoxP3low memory T cells with Th17
potential was significantly elevated among CD4+ T cells
in nasal polyps, while CD69 expression was also significantly
higher in these cells in nasal polyps compared with PBMCs (Table III; Fig. 4). Percentages of CD4+
CD45RA+ FoxP3low resting Treg
(rTreg) were not significantly different in PBMCs and
polyps (Table III). In
contrast, CD4+ T cells in nasal polyps from patients
with CRSwNP contained significantly more CD45RA−
FoxP3high aTreg compared with in PBMCs
(Table III). rTreg
and aTreg cells harbored significantly more
CD69+ cells in nasal polyps compared with in PBMCs
(Table III; Fig. 4). In nasal polyps,
aTreg contained significantly more CD69+
cells compared with rTreg (Fig. 4). Among the different
CD4+ T cell subsets in nasal polyps, aTreg
had the highest number of CD69-expressing cells followed by
Tconv (Table
III).
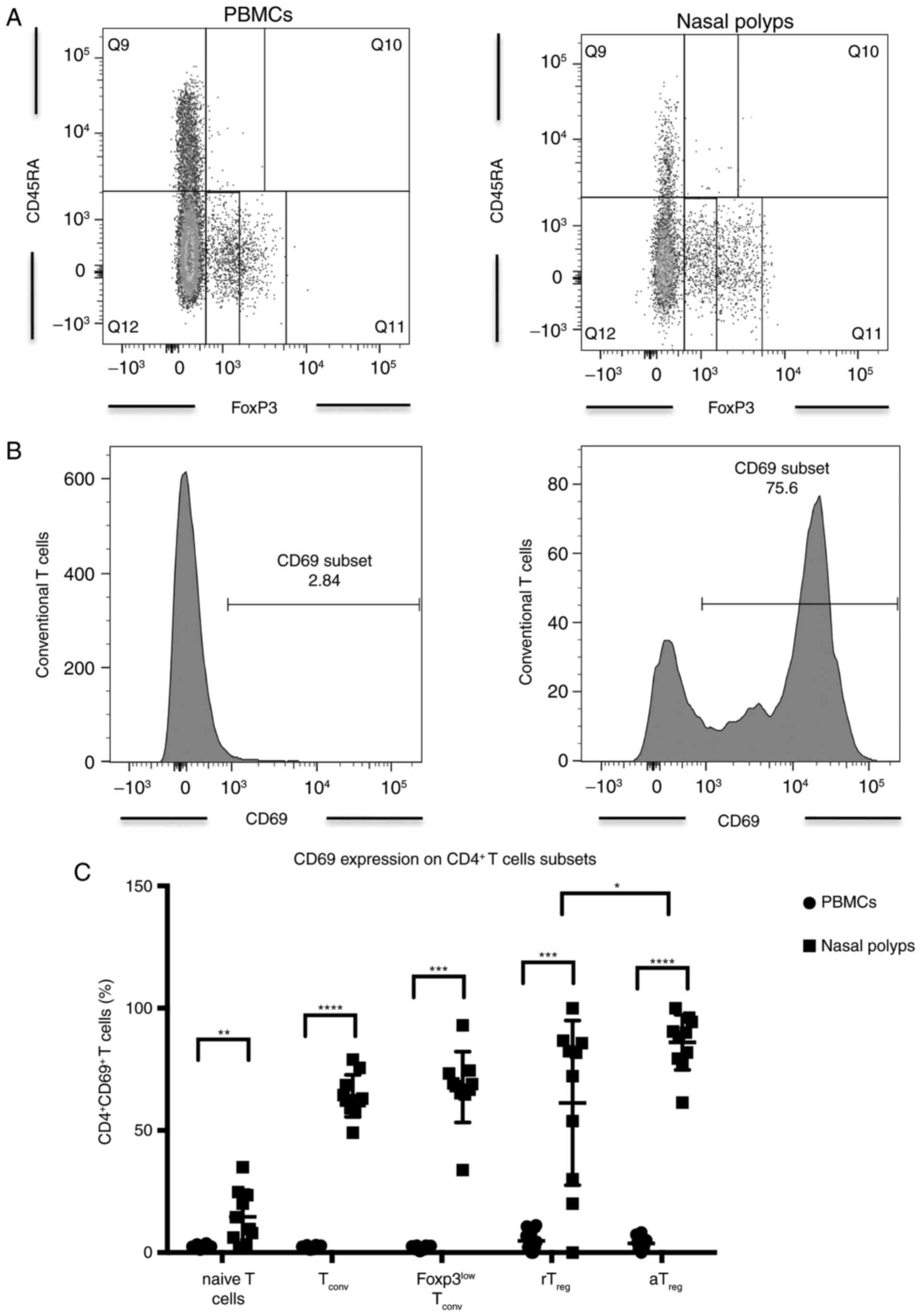 | Figure 4(A) Incidence of CD4+ naïve T cells,
Tconv, FoxP3low Tconv, rTreg and
aTreg in PBMCs and nasal polyps in patients with CRSwNP.
(B) Expression of CD69 in Tconv in PBMCs and nasal
polyps in patients with CRSwNP. (C) Expression of CD69 in CD4+ T
cell subsets in PBMCs and nasal polyps in patients with CRSwNP.
Data are presented as the mean ± standard deviation of 11 patients.
*P<0.05, **P<0.01,
***P<0.001 and ****P<0.0001. CD,
cluster of differentiation; Tconv, conventional memory T
cell; rTreg, resting regulatory T cell;
aTreg, activated regulatory T cell; CRSwNP, chronic
rhinosinusitis with nasal polyps. |
| Table IIICD4+ T cell subpopulations
and their CD69 expression. |
Table III
CD4+ T cell subpopulations
and their CD69 expression.
| CD3+
CD4+ T cells | PBMCs | Nasal polyps | P-value |
|---|
| CD45RA+
FoxP3low CTLA-4low resting
Treg | 0.52±0.30 | 1.24±1.41 | 0.275 |
|
CD69+ | 4.81±3.99 | 61.26±31.97 | <0.0001 |
| CD45RA−
FoxP3high CTLA-4high activated
Treg | 1.29±0.82 | 5.74±2.18 | 0.0004 |
|
CD69+ | 3.78±2.42 | 86.01±10.68 | <0.0001 |
| CD45RA−
Foxp3low memory T cells | 3.63±1.26 | 6.6±1.45 | 0.0004 |
|
CD69+ | 1.99±0.76 | 67.75±13.74 | 0.002 |
| CD45RA−
Foxp3− memory T cells | 56.87±16.27 | 74.73±8.88 | 0.014 |
|
CD69+ | 2.23±0.64 | 64.15±8.15 | <0.0001 |
| CD45RA+
Foxp3− naïve T cells | 37.83±16.64 | 11.01±9.90 | 0.002 |
|
CD69+ | 2.33±0.72 | 14.69±10.27 | 0.002 |
Homing receptor CCR7 on CD4+
and CD8+ T cells in CRSwNP
A significantly reduced proportion of
CCR7+ cells among CD3+ CD4+ and
CD3+ CD8+ T cells was apparent in nasal
polyps compared with PBMCs (Table
II). CD8+ T cells had a significantly lower
incidence of CCR7+ cells compared with CD4+ T
cells in nasal polyps and PBMCs (Table II).
Lack of CD4+ and
CD8+ T cells in CRSsNP
Evaluation of lymphocytes in the nasal mucosa of
patients with CRSsNP was not possible due to the low amounts of
these cells in the tissue harvested intraoperatively. Only cell
counts between 3 and 207 were found for CD4+ and
CD8+ T cells, thus a statistically appropriate analysis
was not possible. For this reason, a comparison of T cell subsets
in samples from patients with CRsSNP and CRSwNP was not possible in
the present study.
Discussion
In the present study, a detailed quantification of
Trm in PBMCs and nasal polyps from patients with CRSwNP
is presented. Percentages of CD69+ cells were
significantly increased in nasal polyps compared with PBMCs.
Furthermore, the incidence of CD69+ cells was
significantly higher among CD8+ T cells compared with
CD4+ T cells in polypoid tissue. Extending the analysis
to S1PR1 expression, the proportion of CD69+
S1PR1− Trm cells was significantly increased
among both CD4+ and CD8+ T cells in nasal
polyps compared with PBMCs in patients with CRSwNP. Furthermore,
the number of CD69+ S1PR1− Trm was
significantly higher among CD8+ compared with
CD4+ T cells. The frequency of S1PR1+ cells
was also significantly increased in edaphic CD4+ and
CD8+ T cells compared with PBMCs. Thus, the number of
Trm identified by double staining of CD69 and S1PR1 was
lower compared with CD69 alone. Nonetheless, the percentage of
CCR7− cells was significantly increased among
CD4+ and CD8+ T cells in edaphic lymphocytes
compared with PBMCs in patients with CRSwNP.
Effector memory T cells migrate from PBMCs into the
local tissue as a result of acute infection. Following further
differentiation into Trm, a high percentage of these
cells will remain in the local tissue (19). Ma et al (19) discussed transforming growth factor
(TGF)-β as one of the major signals for the differentiation of
kidney-resident T cells. However, TGF-β-independent differentiation
of Trm in the intestinal lamina propria has also been
reported (20). In CRSwNP, an
accumulation of effector CD4+ and CD8+ T
cells has been discussed (2).
TGF-β concentrations in CRSwNP differ from CRSsNP and vary between
patients from different countries (21). Therefore, future studies should
focus on the possible factors that drive Trm generation
in CRSwNP.
Different subsets of tissue-resident lymphocytes
have previously been described (22). Tissue residency was mainly
attributed to CD8+ T cells and they were observed in
many different organs (23,24). Memory T cells were subdivided into
Tcm and Tem by the homing receptor CCR7
(25). In the present study, high
numbers of CD8+ CCR7− Tem were
identified in nasal polyps compared with PBMCs from patients with
CRSwNP. The characterization of CD8+ Trm is
heterogeneous, often lacking CCR7 and highly expressing CD69
(15,26). In the present study, significantly
more CD8+ T cells were observed in nasal polyps compared
with in PBMCs. Almost 97% lacked the homing receptor CCR7 and ~63%
of these CD8+ T cells were CD69+. Whether
Trm depend (27) on
specific antigen presentation or not (24) remains controversial. However, a
low incidence of HLA-DR-expressing T cells in nasal polyps
(2) suggests that repeated
antigenic stimulation is not responsible for maintaining T cells
within the polyps. Rather, multiple triggers serve a role in this
chronic disease, including fungal (4–6)
infections or staphylococcal (7,8)
superantigens, which may generate a niche for Trm
development and maintenance independent of antigens.
Similar to CD8+ Trm,
CD4+ Trm are described as
CD69+ T cells and lack the homing receptor CCR7.
In the present study, ~38% of the CD4+ T cells were also
positive for CD69 and 85% lacked the homing receptor CCR7, which
suggests a high percentage of CD4+ Trm
in nasal polyps. Interestingly, CD8+
CD69+ Trm were significantly increased
compared with CD4+ CD69+ T
cells in nasal polyps. The reason for higher CD8+ than
CD4+ Trm numbers in polyps remains unclear
and should be the focus of future studies. Besides identical
expression of CD69, CCR7 and S1PR1 in CD4+ and
CD8+ T cells, the signals for tissue residency are
differentially described for CD4+ compared with
CD8+ Trm in the literature (28). The precise mechanisms for keeping
these T cells inside the tissue are still unclear.
Another subpopulation of CD4+ T cells
which are responsible for several autoimmune disorders are
Treg cells (29,30). Lynch et al (31) reported that Treg do not
recirculate in the blood. In contrast, Luo et al (32) demonstrated that Treg do
not persist in the local tissue for a long period of time. Like
CD4+ and CD8+ Trm, Treg
require the expression of CD69 as a signal to remain in the local
tissue (22). In the present
study, Treg were differentiated into CD3+
CD4+ CD45RA+ FoxP3low
rTreg and CD3+ CD4+
CD45RA− FoxP3high aTreg.
rTreg and aTreg exhibited a significantly
higher expression of CD69 in nasal polyps compared with PBMCs. In
nasal polyps, aTreg had a significantly higher CD69
expression compared with rTreg. These findings suggest
that Treg populations in the polyps primarily consist of
tissue-resident cells.
Skon et al (12) critically remarked that CCR7
downregulation alone is not a reliable marker for Trm.
For a more appropriate characterization of Trm, evidence
of S1PR1 downregulation is required (12,13). Following detection of its ligand,
S1P, S1PR1 is a necessary signal for naïve lymphocytes to exit the
local tissue and recirculate (11). CD69/S1PR1 double staining revealed
significantly more CD3+ CD4+ and
CD3+ CD8+ CD69+ S1PR1−
Trm in nasal polyps compared with PBMCs in patients with
CRSwNP. Furthermore, the frequency of CD69+
S1PR1− Trm was significantly higher among
CD8+ compared with CD4+ T cells, which
underscores the dominating role of CD8+ T cells in
CRSwNP. Interestingly, the expression of S1PR1 alone was
significantly higher in total CD3+ CD4+ and
CD3+ CD8+ T cells from nasal polyps compared
with PBMCs. This may be because S1PR1 expression is modulated by
the binding of S1P and there is a high concentration of this ligand
in PBMCs, with a downregulation on lymphocytes in PBMCs (18).
The pathophysiological function of Trm
has been described in the literature. They are regarded as a potent
sentinel mechanism against acute re-infections, thereby supporting
protective immunity (14). In
contrast, whether a high percentage of Trm in chronic
diseases, including CRSwNP, may act as a possible pathogenic
trigger of the disease itself or of early-onset recurrence
following therapy remains unclear. Schmidt et al discussed
allergen-specific CD8+ Trm as key mediators
for acute contact dermatitis (33). In addition, they may be
responsible for the development of novel sensitizations (33). Park et al (34) outlined the important role of
accumulating resident memory T cells in a various diseases of
barrier and non-barrier tissues. Furthermore, pathological
accumulation of hyperactive Trm as a response to an
extended inflammatory reaction may lead to further disease
(34).
One limitation of the present study is the lack of
a control group comprising the nasal mucosa of patients with
CRSsNP. It is therefore difficult to assess whether the
accumulation of Trm in nasal polyps is pathological or
the normal physiological condition. An analysis of lymphocytes from
the nasal mucosa of patients with CRSsNP was attempted, however the
number of cells was too small for a reliable evaluation. As very
few lymphocytes were able to be isolated from the nasal mucosa of
patients with CRSsNP, an accumulation of Trm seems
unlikely. Sathaliyawala et al (35) performed a unique analysis of human
T cells in healthy lymphoid and mucosal tissue obtained from
individual donors, thus describing a steady state of T cells.
Interestingly, the majority of Trm identified, even in
respiratory mucosae, were CD4+ memory T cells. This is
in contrast to the present study in which the majority of
Trm in polypoid tissue were CD8+ T cells.
This suggests a pathological increase in the percentage of
CD8+ Trm in compared with in healthy
respiratory mucosae from patients with CRSwNP.
Summarizing the findings of this study and the data
in the literature, there are two different T cell pools in nasal
polyps: A high percentage of CD8+ Trm and a
lower percentage of predominantly CD4+ Tem.
Interestingly, these T cells are HLA-DR− (2), therefore there are no recently
activated T cells in the polypoid tissue. Trm may be
important mediators of the chronic inflammatory process in CRSwNP.
Selective inhibition, or eliminating these cells by modifying their
ability to persistently reside in tissue, may be a possible
approach for the development of novel therapeutic strategies
(34). Hypothetically, targeting
and blocking CD69 could, for example, support the elimination of
pathogenic Trm in the tissue. The clinical impact of
Trm in recurrent CRSwNP should be further investigated
in the future.
To the best of our knowledge, this is the first
study to describe resident memory T cells in nasal polyps compared
with PBMCs from patients with CRSwNP. CD8+
Trm dominated CD4+ Trm within
nasal polyps. The role of Trm in nasal polyps as a
pathogenic trigger of the local inflammatory reaction must be
further investigated in future studies; however, the results of the
present study suggest local regulation of the immune response
within the nasal polyps. Thus, Trm can be may be a
potential trigger in the pathogenesis of nasal polyps.
Funding
Dr Niklas Beyersdorf was supported by the Deutsche
Forschungs gemeinschaft (DFG) (SFB/TR 124 FungiNet, Project
C6).
Availability of data and materials
All data generated or analyzed during this study
are included in this published article.
Authors' contributions
PI performed all experiments, analyzed the results
and was the main author of the manuscript. XD, NB and TK conceived
the study and analyzed the results. NK, RH and CG analyzed the data
and were major contributors to the manuscript. SH conceived the
study, analyzed the results and was a major contributor to the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Board of the
Medical Faculty, Julius-Maximilian-University Wuerzburg (vote no.
12/06). Ethics approval and written informed consent have been
obtained from every patient.
Consent for publication
Not applicable.
Competing interests
The authors declare that there are no competing
interests.
Abbreviations:
|
CRSwNP
|
chronic rhinosinusitis with nasal
polyps
|
|
Treg
|
regulatory T cells
|
|
aTreg
|
activated regulatory T cells
|
|
rTreg
|
resting regulatory T cells
|
|
Tconv
|
conventional T cells
|
|
Trm
|
tissue-resident memory T cells
|
|
Tem
|
effector memory T cells
|
|
Tcm
|
central memory T cells
|
Acknowledgments
Not applicable.
References
|
1
|
Bachert C and Akdis CA: Phenotypes and
emerging endotypes of chronic rhinosinusitis. J Allergy Clin
Immunol Pract. 4:621–628. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ickrath P, Kleinsasser N, Ding X, Ginzkey
C, Beyersdorf N, Hagen R, Kerkau T and Hackenberg S:
Characterization of T-cell subpopulations in patients with chronic
rhinosinusitis with nasal polyposis. Allergy Rhinol (Providence).
8:139–147. 2017. View Article : Google Scholar
|
|
3
|
Miyara M, Yoshioka Y, Kitoh A, Shima T,
Wing K, Niwa A, Parizot C, Taflin C, Heike T, Valeyre D, et al:
Functional delineation and differentiation dynamics of human
CD4+ T cells expressing the FoxP3 transcription factor.
Immunity. 30:899–911. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ponikau JU, Sherris DA, Kern EB, Homburger
HA, Frigas E, Gaffey TA and Roberts GD: The diagnosis and incidence
of allergic fungal sinusitis. Mayo Clin Proc. 74:877–884. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pant H, Hughes A, Miljkovic D, Schembri M,
Wormald P, Macardle P, Grose R, Zola H and Krumbiegel D:
Accumulation of effector memory CD8+ T cells in nasal
polyps. Am J Rhinol Allergy. 27:e117–e126. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pant H and Macardle P: CD8(+) T cells
implicated in the pathogenesis of allergic fungal rhinosinusitis.
Allergy Rhinol (Providence). 5:146–156. 2014. View Article : Google Scholar
|
|
7
|
Bachert C, Zhang N, Patou J, van Zele T
and Gevaert P: Role of staphylococcal superantigens in upper airway
disease. Curr Opin Allergy Clin Immunol. 8:34–38. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Van Zele T, Gevaert P, Holtappels G, van
Cauwenberge P and Bachert C: Local immunoglobulin production in
nasal polyposis is modulated by superantigens. Clin Exp Allergy.
37:1840–1847. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ickrath P, Kleinsasser N, Ding X, Ginzkey
C, Beyersdorf N, Kerkau T, Hagen R and Hackenberg S: Impact and
modulations of peripheral and edaphic B cell subpopulations in
chronic rhino-sinusitis with nasal polyposis. Clin Exp
Otorhinolaryngol. Feb 8–2018.Epub ahead of print. View Article : Google Scholar
|
|
10
|
Bromley SK, Thomas SY and Luster AD:
Chemokine receptor CCR7 guides T cell exit from peripheral tissues
and entry into afferent lymphatics. Nat Immunol. 6:895–901. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Matloubian M, Lo CG, Cinamon G, Lesneski
MJ, Xu Y, Brinkmann V, Allende ML, Proia RL and Cyster JG:
Lymphocyte egress from thymus and peripheral lymphoid organs is
dependent on S1P receptor 1. Nature. 427:355–360. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Skon CN, Lee JY, Anderson KG, Masopust D,
Hogquist KA and Jameson SC: Transcriptional downregulation of S1pr1
is required for the establishment of resident memory
CD8+ T cells. Nat Immunol. 14:1285–1293. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mackay LK, Braun A, Macleod BL, Collins N,
Tebartz C, Bedoui S, Carbone FR and Gebhardt T: Cutting edge: CD69
interference with sphingosine-1-phosphate receptor function
regulates peripheral T cell retention. J Immunol. 194:2059–2063.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schenkel JM and Masopust D:
Tissue-resident memory T cells. Immunity. 41:886–897. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Brown MN, Fintushel SR, Lee MH, Jennrich
S, Geherin SA, Hay JB, Butcher EC and Debes GF: Chemoattractant
receptors and lymphocyte egress from extralymphoid tissue: Changing
requirements during the course of inflammation. J Immunol.
185:4873–4882. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiang X, Clark RA, Liu L, Wagers AJ,
Fuhlbrigge RC and Kupper TS: Skin infection generates non-migratory
memory CD8+ T(RM) cells providing global skin immunity.
Nature. 483:227–231. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schrijvers R, Gilissen L, Chiriac AM and
Demoly P: Pathogenesis and diagnosis of delayed-type drug
hypersensitivity reactions, from bedside to bench and back. Clin
Transl Allergy. 5:312015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Aoki M, Aoki H, Ramanathan R, Hait NC and
Takabe K: Sphingosine-1-phosphate signaling in immune cells and
inflammation: Roles and therapeutic potential. Mediators Inflamm.
2016:86068782016.PubMed/NCBI
|
|
19
|
Ma C, Mishra S, Demel EL, Liu Y and Zhang
N: TGF-β controls the formation of kidney-resident T cells via
promoting effector T cell extravasation. J Immunol. 198:749–756.
2016. View Article : Google Scholar
|
|
20
|
Bergsbaken T and Bevan MJ: Proinflammatory
microenvironments within the intestine regulate the differentiation
of tissue-resident CD8(+) T cells responding to infection. Nat
Immunol. 16:406–414. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang X, Zhang N, Bo M, Holtappels G, Zheng
M, Lou H, Wang H, Zhang L and Bachert C: Diversity of TH cytokine
profiles in patients with chronic rhinosinusitis: A multicenter
study in Europe, Asia, and Oceania. J Allergy Clin Immunol.
138:1344–1353. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mackay LK and Kallies A: Transcriptional
regulation of tissue-resident lymphocytes. Trends Immunol.
38:94–103. 2017. View Article : Google Scholar
|
|
23
|
Anderson KG, Sung H, Skon CN, Lefrancois
L, Deisinger A, Vezys V and Masopust D: Cutting edge: Intravascular
staining redefines lung CD8 T cell responses. J Immunol.
189:2702–2706. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Casey KA, Fraser KA, Schenkel JM, Moran A,
Abt MC, Beura LK, Lucas PJ, Artis D, Wherry EJ, Hogquist K, et al:
Antigen-independent differentiation and maintenance of
effector-like resident memory T cells in tissues. J Immunol.
188:4866–4875. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sallusto F, Lenig D, Forster R, Lipp M and
Lanzavecchia A: Two subsets of memory T lymphocytes with distinct
homing potentials and effector functions. Nature. 401:708–712.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Steinert EM, Schenkel JM, Fraser KA, Beura
LK, Manlove LS, Igyártó BZ, Southern PJ and Masopust D: Quantifying
memory CD8 T cells reveals regionalization of immunosurveillance.
Cell. 161:737–749. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Steinbach K, Vincenti I, Kreutzfeldt M,
Page N, Muschaweckh A, Wagner I, Drexler I, Pinschewer D, Korn T
and Merkler D: Brain-resident memory T cells represent an
autonomous cytotoxic barrier to viral infection. J Exp Med.
213:1571–1587. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gebhardt T, Whitney PG, Zaid A, Mackay LK,
Brooks AG, Heath WR, Carbone FR and Mueller SN: Different patterns
of peripheral migration by memory CD4+ and
CD8+ T cells. Nature. 477:216–219. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sakaguchi S, Yamaguchi T, Nomura T and Ono
M: Regulatory T cells and immune tolerance. Cell. 133:775–787.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Robinson DS: The role of regulatory T
lymphocytes in asthma pathogenesis. Curr Allergy Asthma Rep.
5:136–141. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lynch L, Michelet X, Zhang S, Brennan PJ,
Moseman A, Lester C, Besra G, Vomhof-Dekrey EE, Tighe M, Koay HF,
et al: Regulatory iNKT cells lack expression of the transcription
factor PLZF and control the homeostasis of T(reg) cells and
macrophages in adipose tissue. Nat Immunol. 16:85–95. 2015.
View Article : Google Scholar
|
|
32
|
Luo CT, Liao W, Dadi S, Toure A and Li MO:
Graded Foxo1 activity in Treg cells differentiates tumour immunity
from spontaneous autoimmunity. Nature. 529:532–536. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schmidt JD, Ahlström MG, Johansen JD,
Dyring-Andersen B, Agerbeck C, Nielsen MM, Poulsen SS, Woetmann A,
Ødum N, Thomsen AR, et al: Rapid allergen-induced interleukin-17
and interferon-γ secretion by skin-resident memory CD8+
T cells. Contact Dermatitis. 76:218–227. 2016. View Article : Google Scholar
|
|
34
|
Park CO and Kupper TS: The emerging role
of resident memory T cells in protective immunity and inflammatory
disease. Nat Med. 21:688–697. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sathaliyawala T, Kubota M, Yudanin N,
Turner D, Camp P, Thome JJ, Bickham KL, Lerner H, Goldstein M,
Sykes M, et al: Distribution and compartmentalization of human
circulating and tissue-resident memory T cell subsets. Immunity.
38:187–197. 2013. View Article : Google Scholar :
|