Introduction
Colorectal cancer (CRC) is considered as one of the
most common malignancies with a high incidence and mortality
worldwide. It is estimated that 1.4 million individuals were newly
diagnosed with CRC in 2012, which resulted in 693,900 mortalities
(1). Surgery, chemotherapy and
radiation treatment are the three main standard therapies for CRC.
Chemotherapy is an important treatment for cancer, particularly in
tumors with a propensity to invade adjacent tissues and metastasize
to other organs. The introduction of advanced surgical and systemic
therapeutic options has improved the prognosis for CRC over the
past few years. However, challenges remain that require the
continued search for novel effective and less toxic
chemotherapeutic agents for the treatment of colon cancer (2).
Artesunate (ART), a natural sesquiterpene extracted
from a Chinese medicinal herb, is recognized as a safe compound for
treating malaria (3). It is
considered to be suitable for drug development due to its aqueous
solubility. In 1996, Efferth et al (4) discovered for the first time that ART
was able to induce tumor cell apoptosis. Currently, the potential
anticancer effects of ART have been reported in numerous tumors,
including myelodysplastic syndrome (5,6),
multiple myeloma (7), chronic
myeloid leukemia (8), Burkitt’s
lymphoma (9), bladder cancer
(10), renal carcinoma (11), cervical (12), breast (13), prostate (14), and head and neck (15) cancer, hepatocellular carcinoma
(16), esophageal cancer
(17) and CRC (18). However, the specific mechanism of
its antitumor activity remain unclear.
Presently, the ART-induced tumor cell death has been
reported to mainly involve apoptosis (5,16,18,19), autophagy (13), oncosis (11) and ferroptosis (15). Cell death signaling is complex,
and the different processes function in concert. Apoptosis and
autophagy are two distinct processes that coordinately regulate
cell survival and death, and occur simultaneously in numerous types
of cancer (20). Furthermore,
these two processes have been identified in CRC (21).
It has been reported that ART induces apoptosis and
autophagic excitation of Raji cells in Burkitt’s lymphoma (9) and suppresses HCT116 cell
proliferation through apoptosis pathways (18). Therefore, in the present study,
the effects of ART on apoptosis and autophagy of HCT116 cells were
investigated, and the association between apoptosis and autophagy
was explored in vitro. In addition, the inhibitory effect of
ART on subcutaneous xenografts was assessed in nude mice.
Materials and methods
Cells and animals
The human HCT116 CRC cell line was routinely
cultured at the Centre Laboratory of the Affiliated Hospital of
Nanjing University of TCM. The cells were cultured in RPMI 1640
medium (HyClone; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal calf serum (Evergreen; Zhejiang
Tianhang Biotechnology Co., Ltd, Luoshe, China). The cells were
incubated at 37°C in a humidified atmosphere of 5%
CO2.
The experimental animal protocols were reviewed and
approved by the Animal Ethics Committee of the Affiliated Hospital
of Nanjing University of TCM (2017-DW-11). A total of 20 6-week-old
BALB/c nude mice weighing 18–22 g (no. 201716339) were purchased
from Changzhou Cavens Laboratory Animal Co., Ltd (Changzhou,
China). They were maintained under specific pathogen-free
conditions at room temperature (23±2°C) and relative humidity
(40–60%), with a12 h light/dark cycle, and their cages, litter,
food and water were strictly sterilized.
Materials
ART was purchased from Guilin Pharmaceutical Co.,
Ltd. (Guilin, China). According to the manufacturer’s protocol, ART
was dissolved to a concentration of 20 mg/ml and then stored at
−20°C. Prior to the experiments, the ART stock was diluted to the
final desired concentrations with complete RPMI 1640. 5-Fu was
purchased from Tianjin King York Pharmaceutical Co., Ltd. (Tianjin,
China).
Antibodies against caspase 3 (cat. no. 9665),
caspase 9 (cat. no. 9502), poly-ADP ribose polymerase (PARP; cat.
no. 9532), B-cell lymphoma-2 (Bcl-2; cat. no. 2876),
Bcl-extra-large (Bcl-xL; cat. no. 2764), Bcl-2-associated X protein
(Bax; cat. no. 2772), light chain 3 (LC3)-I/II (cat. no. 12741),
beclin-1 (cat. no. 3495), autophagy protein 5 (Atg5; cat. no.
12994), Atg12 (cat. no. 4180), β-actin (cat. no. 4970) and
anti-rabbit IgG HRP-linked Antibody (cat. no. 7074P2) were
purchased from Cell Signaling Technology, Inc., (Danvers, MA, USA).
Primary antibody was purchased from Wuhan Servicebio Technology
Co., Ltd. (Wuhan, China; cat. no. G2025), and the dilution ratio
used for each antibody was 1:1,000. Hydroxychloroquine sulfate
(HCQ) was purchased from Selleck Chemicals (Houston, TX, USA).
3-(4,5-Dimethylthiazol-2-yl)2,5-diphenyl
tetrazolium bromide (MTT) assay
Cell viability was evaluated using an MTT assay.
Briefly, the cells were seeded in 96-well plates at a cell density
of 4,000 cells/well. Following treatment of the cells with ART at
various concentrations (2.5, 10, 40 and 80 µg/ml) or the
control for the indicated time (24, 48 and 72 h), the MTT reagent
was added to each well at a final concentration of 5 mg/ml,
followed by incubation at 37°C for 4 h. Next, the medium was
removed, and 150 µl dimethyl sulfoxide was added to each
well. The absorbance was measured at 490 nm using a microplate
reader (BioTek Instruments, Inc., Winooski, VT, USA). The relative
cell viability was calculated as follows: Relative cell viability
(%)=(Mean absorbance of the test wells/mean absorbance of the
control wells) ×100. The half maximal inhibitory concentration
(IC50) was calculated using SPSS Software based on the
results of MTT. Ranjan et al (22) identified the IC50 of
penfluridol was 6.5 µM in Panc-1 cells following 24 h
treatment. Thus, 5 µM and 10 µM around 6.5 µM
were selected for the study. Wang et al (23) selected 20 µM and 40
µM emodin for the experiment in accordance with the results
of IC50. In the present experiment, the IC50
of ART on HCT116 was 2.18 µg/ml following 48 h. When the
concentration of ART was <1 µg/ml, the inhibition was
<30%, so 2 µg/ml was selected as the low dosage group,
indicating 30–50% inhibition. Twice the low dosage was used for the
high dosage group (4 µg/ml ART), which resulted in >50%
inhibition.
Cell morphological changes examined using
4′,6-diamidino- 2-phenylindole (DAPI) staining
HCT116 cells were treated with ART (2 and 4
µg/ml) and 5-Fu (0.3 µg/ml) for 48 h respectively,
fixed with 4% paraformaldehyde for 10 min at room temperature and
then stained with DAPI (Beyotime Institute of Biotechnology,
Shanghai, China) for 15 min. Subsequently, the cells were observed
under an LSM710 microscope (Carl Zeiss, Oberkochen, Germany).
Detection of apoptosis using flow
cytometry
The apoptosis of treated cells was examined using an
Annexin V-FITC/PI apoptosis kit (MultiSciences Biotech Co., Ltd.,
Hangzhou, China) according to the manufacturer’s protocol. Briefly,
the cells were collected and washed twice in phosphate-buffered
saline (PBS). Cells were then labeled with 500 µl 1X binding
buffer containing 5 µl Annexin V-fluorescein isothiocyanate
(FITC) and 10 µl propidium iodide (PI) in the dark for 5
min. Subsequently, apoptosis was detected using a flow cytometer
(BD Falcon; BD Biosciences, Franklin Lakes, NJ, USA), and the
Annexin V and PI values were set as the horizontal and vertical
axes, respectively, for the plot construction. Mechanically
damaged, late apoptotic, dual negative/normal, and early apoptotic
cells were located in the upper left, upper right, lower left and
lower right quadrants of the flow cytometric dot plot,
respectively.
Transmission electron microscopy (TEM)
analysis
HCT116 cells were postfixed in 2.5% glutaraldehyde
for 4 h at 4°C (Wuhan Goodbio Technology Co., Ltd, Wuhan, China),
rinsed with PBS and then fixed in 1% osmium tetroxide
(OsO4; Sinopharm Chemical Reagents Co., Ltd., Shanghai,
China) in 0.1 M PBS for 1 h at room temperature. Following
dehydration with ethanol (Sinopharm Chemical Reagent Co., Ltd.,
Shanghai, China), the cells were embedded in Epon resin (Sales
Performance International Greater China Co., Beijing, China), and
ultrathin sections of the selected areas were cut using the LKB
NOVA ultramicrotome (Leica Biosystems, Solms, Germany) using a
diamond knife (Daito Me Holdings Co., Ltd, Tokyo, Japan). Cells
were then stained with a saturated solution of uranyl acetate in
methanol (50:50) (Sinopharm Chemical Reagent Co., Ltd.) for 12 min
at 45°C, followed by incubation in an aqueous solution of
concentrated bismuth subnitrate (Sinopharm Chemical Reagent Co.,
Ltd.) for 10 min at 25°C. Subsequently, all the sections were
examined under a Hitachi TEM system (Hitachi, Ltd., Tokyo,
Japan).
Animal studies
HCT116 cells in the logarithmic growth phase were
resuspended and digested to a density of 1×107 cells/ml.
Each mouse was inoculated with 0.2 ml of the cell suspension into
the armpit. Approximately 2 weeks after inoculation, 15 mice that
exhibited induration with a diameter of 8–10 mm, which suggested
that a successful tumor model was established, were randomly
divided into three groups. Mice in the three groups were
administered physiological saline [NS; 0.2 ml/day,
intraperitoneally (i.p.), every other day (qod)], 5-fluorouracil
(5-Fu; 12.5 mg/kg, i.p., qod) or ART (100 mg/kg, i.p., daily),
respectively, for 2 weeks.
Observation of antitumor effects
The daily diet, defecation, urination, activity
status and tumor growth of the mice were observed. Subsequent to
treatment, the short- and long-axis diameters of the tumors were
measured every other day, and the tumor volumes were calculated as
follows using the Steel formula (17): Tumor volume (V)
(cm3)=(long diameter x short diameter2)/2.
The volume inhibition rate (%) was also determined as follows:
(1-Vexperimental group/Vcontrol group) ×100%.
The mice were euthanized on day 14 after drug administration. The
primary tumors were excised, and the tumor volumes were measured as
V=4/3πr3, where r is the mean radius of three
measurements. Half of the tumor tissue was fixed in formalin for 24
h at room temperature and embedded in paraffin for TdT-mediated
dUTP nick end labelling (TUNEL) and immunohistochemical assays,
while the other part of the tumor was snap frozen in liquid
nitrogen and stored at −80°C for western blotting analysis.
Immunohistochemistry
Paraffin-embedded tumor tissue samples were cut into
3-µm sections, deparaffinized and subjected to antigen
recovery with citric acid buffer (Wuhan Servicebio Technology Co.,
Ltd.) under high pressure for 2 min. Following blocking with goat
serum (OriGene Technologies, Inc., Beijing, China) at room
temperature for 10 min, then the samples were stained with rabbit
anti-cleaved caspase 3 (1:300), anti-Bax (1:200) and anti-Bcl-2
(1:200) antibodies at 4°C overnight. 3,3′-Diaminobenzidine (Wuhan
Servicebio Technology Co., Ltd.) was used to detect the
immunocomplexes, whereas hematoxylin was used for nuclear
counterstaining. An immunoglobulin-negative control was used to
eliminate nonspecific binding. The sections were examined by light
microscopy (Leica Biosystems, Solms, Germany) at ×200
magnification. Images were analyzed using Image Pro Plus (version
6.0; Media Cybernetics, Inc., Rockville, MD, USA). The Integrated
optical density from three fields of each slice was calculated
using Intel IPP software (version 6.0; Intel, Mountain View, CA,
USA). Three fields were selected from the top to the bottom of each
slice randomly.
TUNEL assay
Sections (3 µm thick) cut from the
paraffin-embedded tissue were dewaxed with xylene twice for 15 min,
hydrated using an ethanol gradient (twice with 100% for 5 min, then
85% for 5 min and 75% for 5 min), fixed in 4% formaldehyde solution
for 20 min at room temperature and then incubated with proteinase K
at 37°C for 30 min. The TUNEL assay kit (Roche Diagnostics, Basel,
Switzerland) containing TdT was prepared immediately before use
according to the manufacturer’s protocol. Subsequent to washing
with PBS, the sections were counterstained with DAPI. Apoptotic
cells in the sections were detected using a microscope (Nikon
Corp., Tokyo, Japan).
Western blot analysis
The cells and tumor tissues were lysed with ice-cold
radio immunoprecipitation assay buffer containing a protease
inhibitor and phosphatase inhibitors (KeyGen Biotech Co., Ltd.,
Nanjing, China). Protein concentrations were determined using a
modified bovine serum albumin assay kit (Thermo Fisher Scientific,
Inc.). Equal amounts of protein (30 µg) from each sample
were separated using 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis, followed by transfer onto polyvinylidene
difluoride membranes (EMD Millipore, Billerica, MA, USA) and then
incubation with 5% BSA for 1 h at room temperature for blocking.
The membranes were subsequently probed with different primary
antibodies (Cell Signaling Technology, Inc.) at 4°C overnight.
Next, the membranes were washed with Tris-buffered saline with
Tween-20 (Sangon Biotech Co., Ltd. Shanghai, China) and incubated
with anti-rabbit IgG HRP-conjugated antibody. An enhanced
chemiluminescence kit (EMD Millipore) was subsequently used to
detect the immuno-reactive signals. Image J software (version 2.1;
National Institutes of Health, Bethesda, MD, USA) was used analyze
the results and calculate the expression of the protein.
Statistical analyses
The results are expressed as the mean ± standard
error of the mean. The data were analyzed using the t-test and
one-way analysis of variance. Differences among the groups were
analyzed using Duncan’s multiple range test with the SPSS software
(version 22.0; IBM Corp., Armonk, NY, USA). A value of P<0.05
was considered to denote a statistically significant
difference.
Results
ART inhibits the viability and growth of
HCT116 cells
The structure of ART (Fig. 1A) was derived from the
instructions of pharmaceutical company. Treatment of HCT116 cells
with different concentrations of ART for various times inhibited
the cell viability in a time- and concentration-dependent manner as
determined by the MTT assay results (Table I). Upon treatment with different
concentrations, the inhibition rates at 48 and 72 h were
significantly different as compared with that at 24 h (P<0.01).
Furthermore, at 48 and 72 h, incubation with 40 and 80 µg/ml
ART resulted in significantly inhibited cell viability when
compared with that observed for the 2.5 and 10 µg/ml
concentrations (P<0.05), whereas no significant difference was
observed among the various concentrations at 24 h. (Fig. 1B).
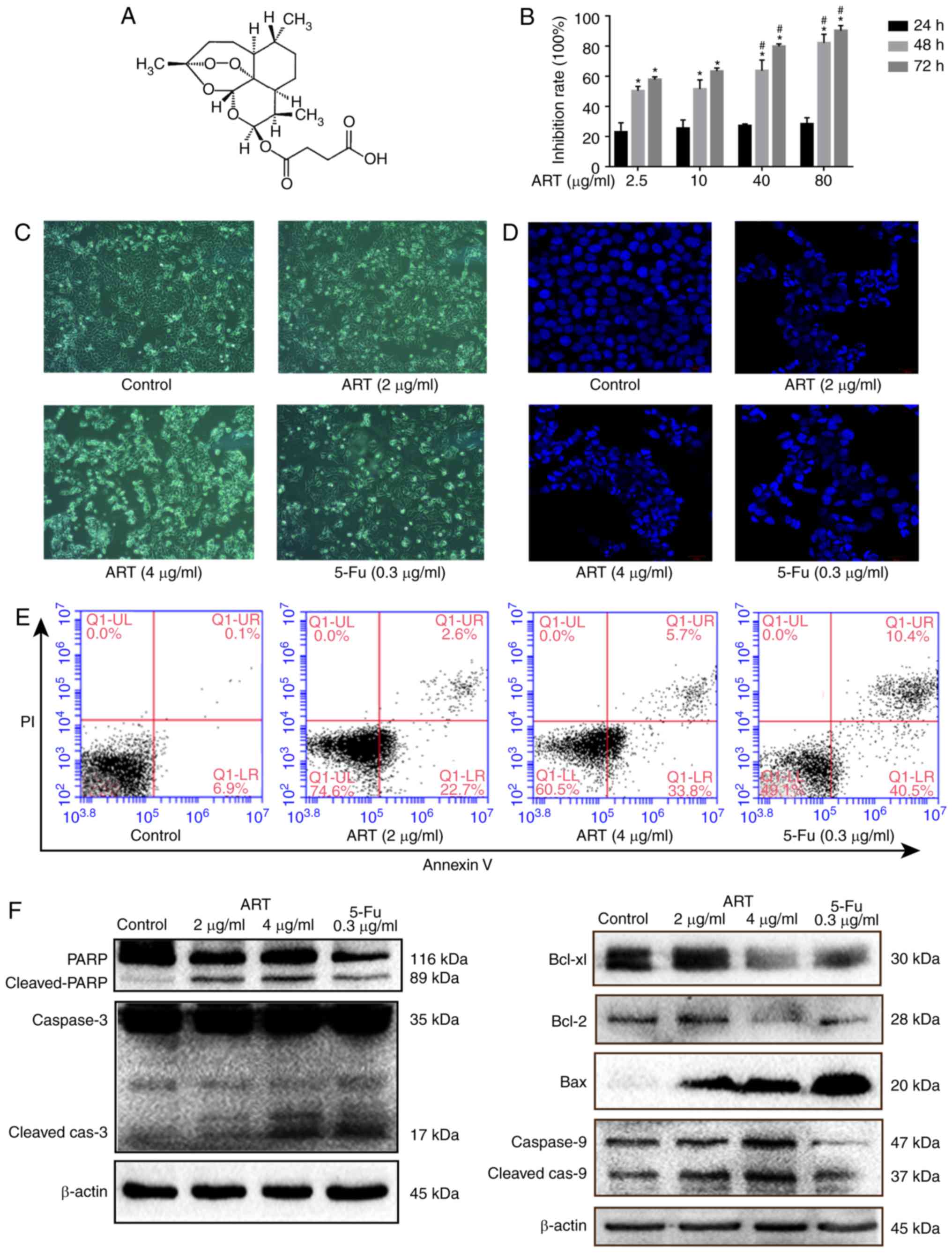 | Figure 1ART inhibits the growth of HCT116
cells and induces apoptosis. (A) Structure of ART. (B) Cell
viability of HCT116 cells treated with ART (2.5, 10, 40 and 80
µg/ml) for 24, 48 and 72 h, determined using an MTT assay.
In HCT116 cells treated with 2 and 4 µg/ml ART for 48 h, (C)
morphological changes were examined using phase-contrast microscope
(magnification, ×200), (D) DAPI staining and confocal microscopy
were used to assess changes in the nucleus, (E) apoptosis was
analyzed using flow cytometry, and (F) apoptosis-associated
proteins (including caspase 3, PARP, Bax, Bcl-2, Bcl-xL and caspase
9) were examined using western blotting. *P<0.01 vs.
corresponding inhibition rate at 24 h; #P<0.05 vs.
inhibition rates of 2.5 and 10 µg/ml ART at the same time
point. ART, artesunate; PARP, poly-ADP ribose polymerase; Bcl-2,
B-cell lymphoma-2; Bax, Bcl-2 associated X protein; Bcl-xL,
Bcl-extra-large protein; 5-Fu, 5-fluorouracil. |
 | Table IGrowth inhibition rate of HCT116
cells at different ART concentrations and incubation times. |
Table I
Growth inhibition rate of HCT116
cells at different ART concentrations and incubation times.
| ART concentration
(µg/ml) | Cell growth
inhibition rate (%)
|
|---|
| 24 h | 48 h | 72 h |
|---|
| 2.5 | 22.95±6.13 | 50.40±2.87a | 57.81±1.79a |
| 10 | 25.38±5.59 | 51.56±6.03a | 63.38±2.07a |
| 40 | 27.07±1.34 | 63.60±7.19a,b | 79.91±1.60a,b |
| 80 | 28.30±4.17 | 82.24±5.68a,b | 90.29±3.30a,b |
ART promotes apoptosis in HCT116 cells
via the mitochondrial pathway
Treatment of HCT116 cells with 2, 4 µg/ml ART
and 0.3 µg/ml 5-Fu for 48 h evidently reduced the number of
viable cells. The cell morphology was markedly changed, with cell
elongation and membrane foaming observed (Fig. 1C). DAPI staining also revealed the
typical apoptotic morphological changes of nuclear condensation,
fragmentation, and chromatin shrinkage (Fig. 1D).
To further confirm the apoptosis-inducing activity
of ART, HCT116 cells were subjected to Annexin V-FITC/PI staining
with flow cytometry, while 5-Fu was used as the positive
proapoptotic drug. The analysis of the results revealed that
apoptotic cells accounted for 22.7 and 33.8% of the cells in early
apoptosis (lower right quadrant) following a 48-h treatment with 2
and 4 µg/ml ART, while that accounted for 40.5% treated with
5-Fu, respectively (Fig. 1E).
To explore the pattern of ART-induced apoptosis of
HCT116 cells, the levels of key proteins involved in
mitochondria-associated apoptosis were measured using western
blotting. As shown in Fig. 1F,
procaspase 3 was cleaved to active caspase 3 following ART
treatment. In addition, ART increased the proteolytic cleavage of
116 kDa PARP to its 89 kDa fragment, which is a hallmark of
apoptosis. ART also increased the protein expression of Bax and
decreased the levels of Bcl-2 and Bcl-xL, which are
mitochondria-associated proteins. The expression of relative
proteins was also increased in the 5-Fu group. These findings
suggest that ART activates mitochondria-mediated apoptosis in
HCT116 cells.
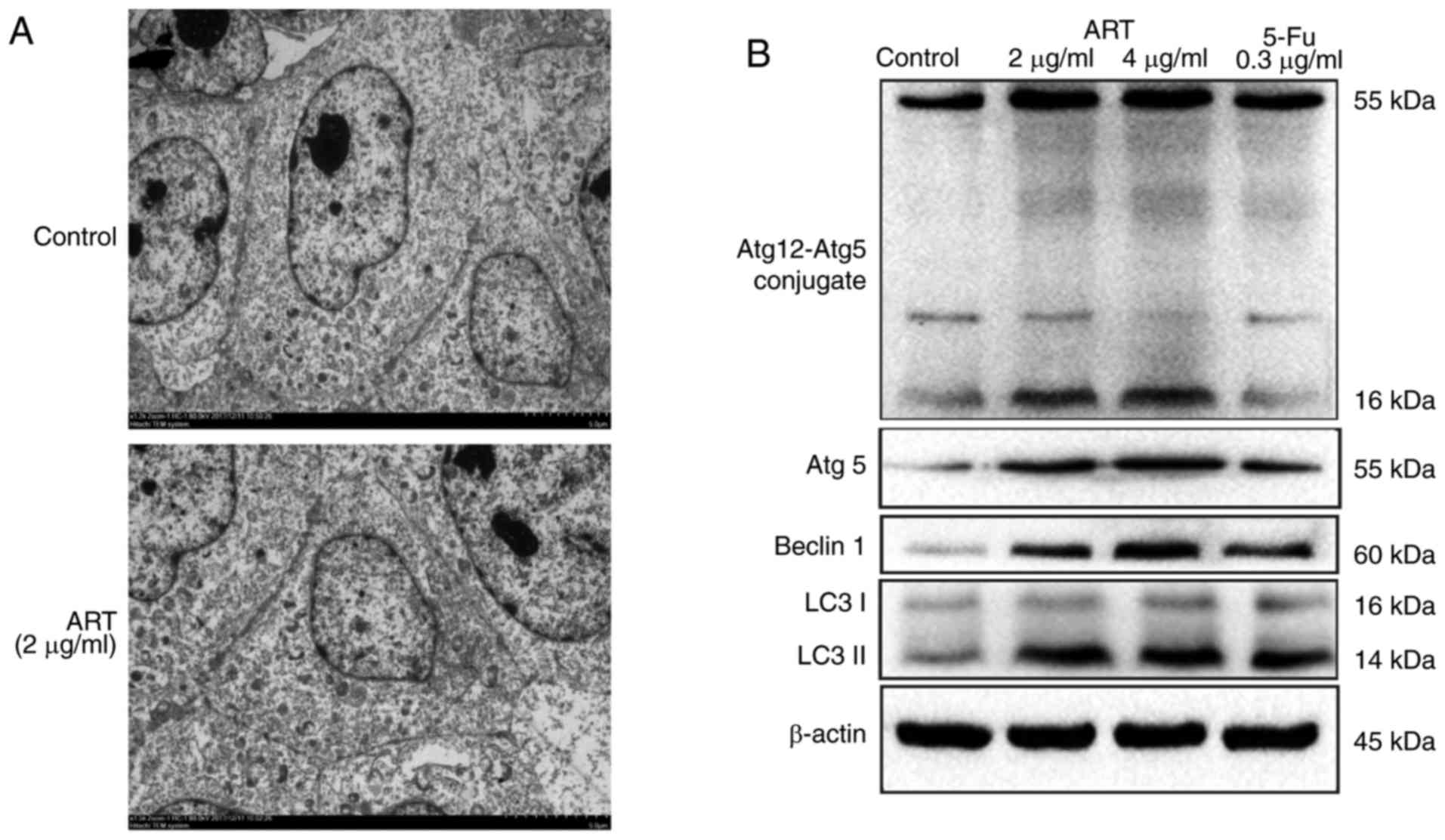
ART induces autophagy in HCT116
cells
Treatment of HCT116 cells with 2 and 4 µg/ml
ART for 48 h induced the appearance of autophagosomes in the cells
(Fig. 2A). The protein expression
levels of the autophagy-specific proteins beclin-1 and LC3-I/II,
the level of Atg5 and the complexation of Atg12-Atg5 increased
following ART exposure, all the protein in 5-Fu group were not
markedly increased (Fig. 2B).
These findings suggest that ART induced autophagy in HCT116
cells.
Autophagy inhibition promotes ART-induced
apoptosis in HCT116 cells
HCT116 cells were treated with 4 µg/ml ART, 4
µg/ml ART + 10 µM HCQ (which is a classic autophagy
inhibitor) (22), or 10 µM
HCQ. As shown in Fig. 3A, the
inhibition of autophagy enhanced the ART-induced inhibition of cell
proliferation (P<0.05). Furthermore, the examination of HCT116
cells in each group using Annexin V-FITC/PI staining with flow
cytometry revealed that the early apoptotic cells accounted for
13.5% in ART group, but increased to 21.9% in the ART + HCQ group
(Fig. 3B). Examination of the
expression levels of autophagy- and apoptosis-associated proteins
revealed that cleaved caspase 3 and pro-PARP levels increased,
while those of LC3-I/II and beclin-1 decreased in the ART+HCQ group
(Fig. 3C). These results
indicated that the number of apoptotic cells following ART
treatment increased when autophagy was inhibited by HCQ.
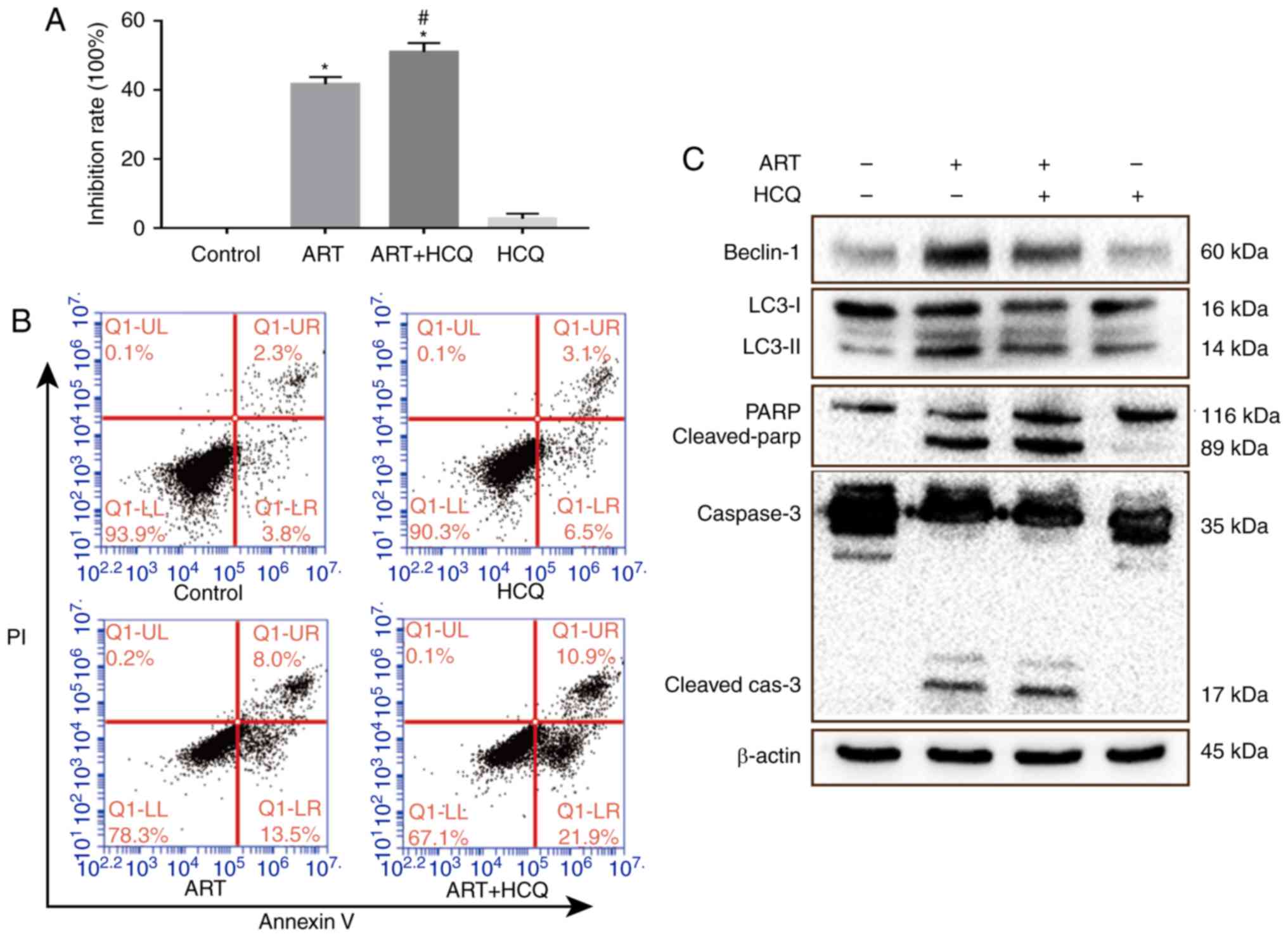 | Figure 3Autophagy inhibition enhances the
ART-induced apoptosis of HCT116 cells. (A) MTT assay of cell
viability of HCT116 cells pretreated with an autophagy inhibitor
(10 µM HCQ; 1 h) followed by ART (4 µg/ml; 48 h). (B)
Apoptosis of cells treated with ART (4 µg/ml), ART (4
µg/ml) +HCQ (10 µM), and HCQ (10 µM) alone for
48 h was analyzed by flow cytometry. (C) Cells were incubated with
ART, ART+HCQ and HCQ for 48 h, and then caspase 3, PARP, LC3 and
beclin-1 protein expression levels were determined using western
blotting. ART, artesunate; HCQ, hydroxychloroquine sulfate; PARP,
poly-ADP ribose polymerase; LC3, light chain 3. |
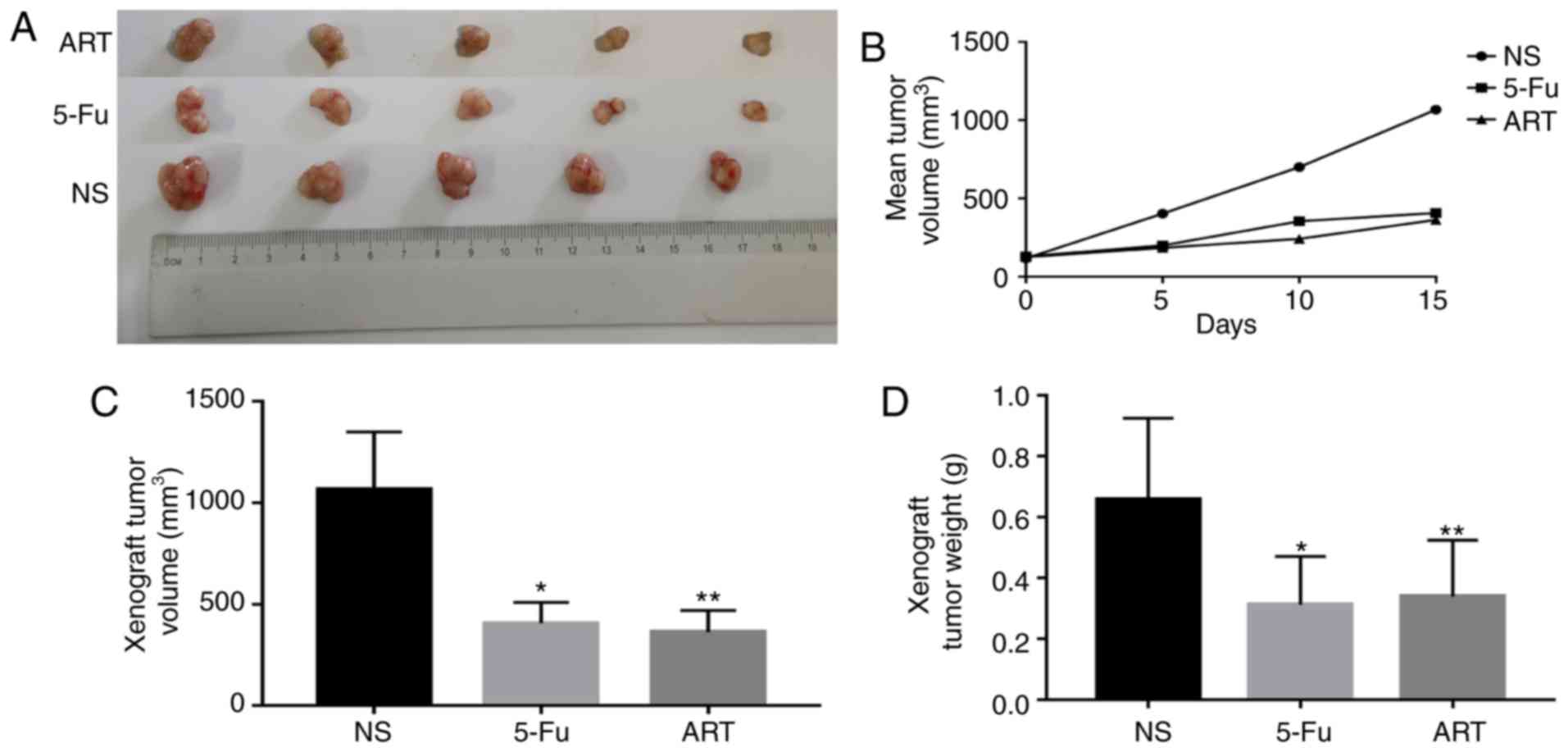
ART inhibits transplanted tumor growth in
mice
The mental state of the animals, intake of food and
water, urination, defecation and other activities were not
significantly different between the NS, 5-Fu and ART groups.
Compared with the control group, the weight of mice in the two
treatment groups was not significantly different (P>0.05; data
not shown). After a 14-day treatment with the test substances, the
mean tumor volumes of the mice in the ART and 5-Fu groups were
smaller compared with those in the control group (Fig. 4A and B). At the end of the
experiment, the ART and 5-Fu groups exhibited more significant
inhibition of tumor growth in comparison with that observed in the
control group (P<0.01; Fig. 4A, C
and D)
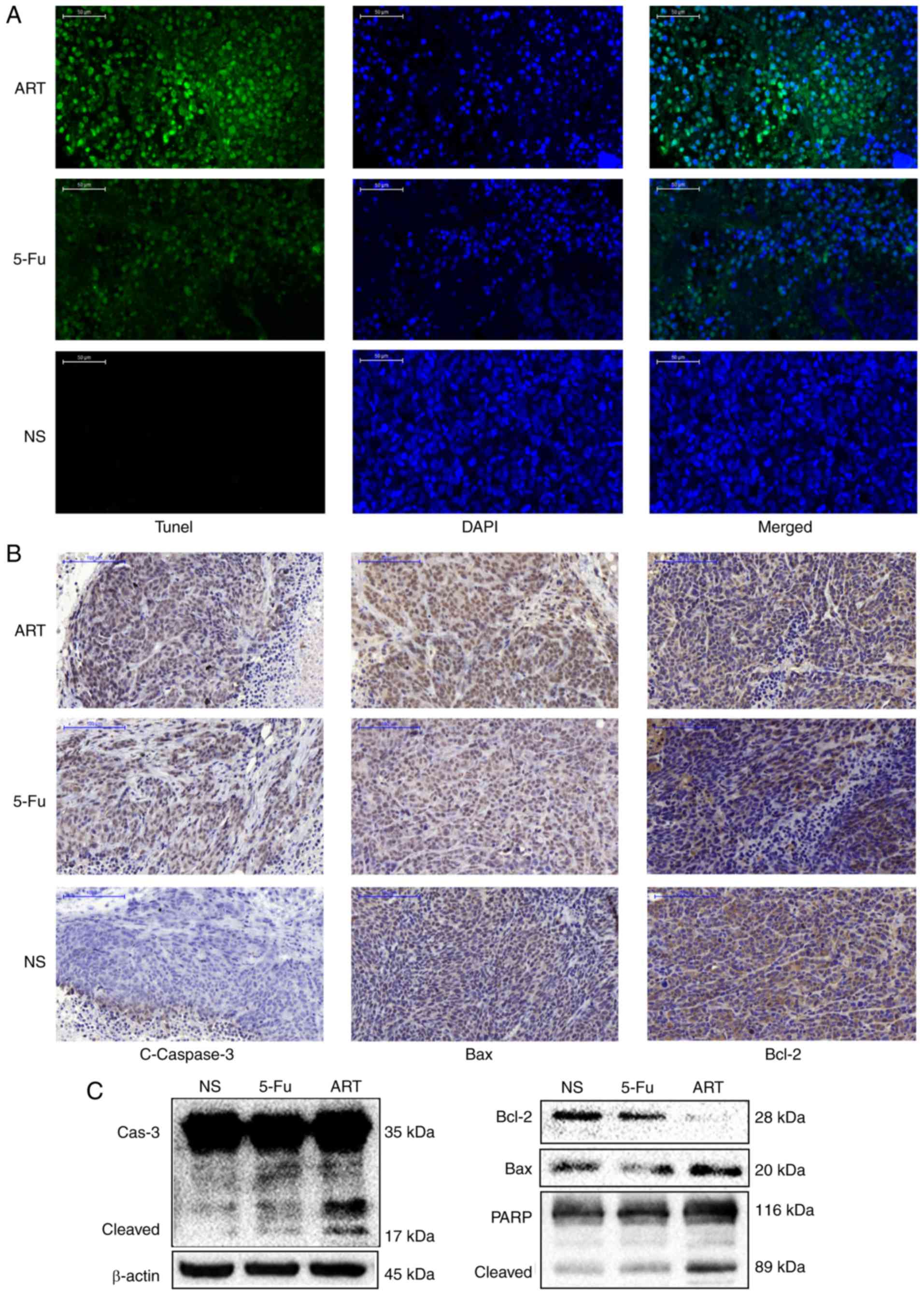
ART promotes the apoptosis of HCT116
cells in transplanted mouse tumors
To elucidate the mechanism of the ART-induced tumor
size and weight suppression, a TUNEL assay was performed to examine
apoptosis and associated protein expression levels in tumor
tissues. ART and 5-Fu promoted the apoptosis of the tumor cells of
transplanted mice as evidenced by the detection of TUNEL-positive
cells with increased DNA fragmentation in the ART and 5-Fu treated
group compared with that in the NS group (Fig. 5A). The immunohistochemical and
western blot analyses also revealed that the protein expression
levels of cleaved caspase 3 and Bax increased, while the level of
Bcl-2 decreased in the ART and 5-Fu group (Fig. 5B and C). The results further
confirmed that the antitumor activity of ART in transplanted tumors
was associated with increased apoptosis.
Discussion
The US Food and Drug Administration-approved ART is
currently the most potent antimalarial agent available (24). The selective killing of cancer but
not normal cells may also be a major advantage of ART, which is
already known to be a generally safe and well-tolerated drug, even
in the fetus during the first trimester of pregnancy (25). To fully understand the mechanisms
of the antitumor actions of ART, the present study investigated for
the first time the association between apoptosis and autophagy in
HCT-116 cells treated with ART.
In the present study, it was observed that ART
significantly inhibited the proliferation of HCT116 cells after 48
h of treatment, whereas the effect was not significant after 24 h.
This result was not consistent with the observations of a previous
study, indicating ART significantly inhibited the cell viability of
HCT116 after 24 h of treatment (18), and a possible explanation for this
apparent discrepancy may be the different drug sources. In the
current study, a commercially available injectable formulation was
used that may contain a number of water-soluble ingredients, while
ART as a monomer was used in the previous investigation that was
purchased from Sigma-Aldrich (Merck, Darmstadt, Germany) (18).
Apoptosis is an active process of programmed cell
death that occurs in multicellular organisms, while the
mitochondrial pathway is an important form of apoptosis. Bax and
Bcl-2 proteins are members of the Bcl-2 family, which consists of
pro- and anti-apoptotic proteins that exert opposing effects on the
mitochondria (26). Upregulation
of Bax and caspase 3, and downregulation of Bcl-2 may result in
cell apoptosis (27). Bax
permeabilizes the mitochondrial outer membrane, releasing
pro-apoptotic factors that activate caspases (28), which can then active the nuclear
protein PARP (29). In the
present study, HCT116 cell apoptosis following intervention with
ART was assessed by DAPI staining and flow cytometry in
vitro. The western blot analysis revealed that the protein
expression levels of Bax, cleaved caspase 3 and cleaved PARP were
increased, while those of Bcl-2 and Bcl-xL were decreased, which
are all mitochondria-associated proteins. In vivo, ART
significantly inhibited tumor size and weight, and evidently
induced the apoptosis of transplanted tumor cells by increasing the
levels of proteins associated with the mitochondrial pathway.
Therefore, the present study suggested that ART induces the
apoptosis of HCT116 cells through the mitochondrial pathway, which
was also observed in other cells, including HepG2 (19), Eca109 and Ec9706 cells (17).
Autophagy is important in normal development and
cellular response to environmental stimuli. It is an intracellular
degradation system that delivers cytoplasmic constituents to the
lysosome and involves the formation of a double-membrane vesicle,
also termed autophagosome. Autophagosomes encapsulate the cytoplasm
and organelles, and fuse with lysosomes, thereby degrading the
vesicle contents (30). This
physiological process is mediated by the interaction of various
molecules, such as beclin1 and LC3, as well as Atg5 and Atg12
(21), which form a conjugate
that is essential for autophagosome formation (31). In CRC cells with high
microsatellite instability (such as HCT116 cells), Atg5 and Atg12
mutations may contribute to the tumor progression by deregulating
the autophagy process (32). In
the present study, intracellular autophagosomes were identified
using TEM, indicating that autophagy occurred in the ART-treated
HCT116 cells. In addition, western blotting revealed that the
levels of LC3-II, beclin-1 and Atg5, as well as the Atg5-Atg12
conjugate, increased after HCT116 cells were treated with ART.
These results indicate that ART induced autophagy in HCT116 cells,
which was also previously reported in breast cancer cells (13).
Autophagy and apoptosis are well-regulated
biological processes that have important roles in tumor development
and progression. Whether autophagy is pro-tumorigenic or
antitumorigenic in cancer development and therapy remains unclear.
Several studies have demonstrated that autophagy has an
anti-apoptotic role in numerous types of cancer (33–36), whereas other studies reported its
essential pro-apoptotic role in certain cancer tissues (22–23,37,38). In the current study, it was
observed that the inhibition of autophagy enhanced the ART-induced
inhibition of cell proliferation. In addition, the percentage of
early apoptotic cells increased after co-treatment with ART and HCQ
compared with that of cells treated with ART alone. Similarly,
autophagy and apoptosis are closely linked in the expression of
specific proteins. The inhibition of beclin-1 by RNAi suppressed
autophagic activity and proliferation, but promoted apoptosis in
the CRC HCT116 and SW620 cell lines (39). The present study revealed that the
protein expression of cleaved caspase 3 and PARP increased, while
that of beclin1 and LC3 decreased in the ART + HCQ group compared
with cells treated with ART alone. Therefore, the results are
consistent with those of other published studies, demonstrating
that autophagy inhibition promotes the apoptosis of certain cancer
cells (33–36).
In conclusion, ART induced apoptosis both in
vitro and in vivo, as well as autophagy in HCT116 cells.
However, autophagy protected HCT116 cells from the apoptosis
induced by ART. These findings provide a basis for further
investigation and development of ART as a potential drug for CRC.
Furthermore, co-administration of ART with an autophagy inhibitor
may improve the efficacy of ART; however, further research would be
required to verify the antitumor activity of ART.
Acknowledgments
Not applicable.
Abbreviations:
|
ART
|
artesunate
|
|
Bcl-2
|
B-cell lymphoma-2
|
|
Bax
|
Bcl-2 associated X protein
|
|
Bcl-xL
|
Bcl-extra-large protein
|
|
CRC
|
colorectal cancer
|
|
DAPI
|
4′,6-diamidino-2-phenylindole
|
|
HCQ
|
hydroxychloroquine sulfate
|
|
TUNEL
|
TdT-mediated dUTP nick end
labelling
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)2,5-diphenyl tetrazolium bromide
|
|
PARP
|
poly-ADP ribose polymerase
|
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81373645, 81573978). This
study was also supported by the Priority Academic Program
Development of Jiangsu Higher Education Institutions and Jiangsu
Province Special Program of Medical Science (grant no. BL2014100)
and by the Peak Academic Talents plan (grant no. BRA2017536) of the
Jiangsu Province Hospital of TCM (Jiangsu, China).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors’ contributions
FJ and YC were responsible for conception and design
of the experiments. FJ and JZ performed the experiments. FJ and DZ
analyzed the data. DZ and ML supplied reagents, materials and
analysis tools. FJ and YC wrote this manuscript. FJ and ML edited
all the pictures. JZ and DZ revised the manuscript critically. All
authors have agreed to the publication of this manuscript.
Ethics approval and consent to
participate
The experimental animal protocols were reviewed and
approved by the Animal Ethics Committee of the Affiliated Hospital
of Nanjing University of TCM (Jiangsu, China; 2017-DW-11).
Patient consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interests
regarding the publication of this manuscript.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Coghlin C and Murray GI: Biomarkers of
colorectal cancer: Recent advances and future challenges.
Proteomics Clin Appl. 9:64–71. 2015. View Article : Google Scholar
|
|
3
|
Efferth T and Kaina B: Toxicity of the
antimalarial artemisinin and its dervatives. Crit Rev Toxicol.
40:405–421. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Efferth T, Rücker G, Falkenberg M, Manns
D, Olbrich A, Fabry U and Osieka R: Detection of apoptosis in KG-1a
leukemic cells treated with investigational drugs.
Arzneimittelforschung. 46:196–200. 1996.PubMed/NCBI
|
|
5
|
Xu N, Zhou X, Wang S, Xu LL, Zhou HS and
Liu XL: Artesunate induces SKM-1 cells apoptosis by inhibiting
hyperactive β-catenin signaling pathway. Int J Med Sci. 12:524–529.
2015. View Article : Google Scholar :
|
|
6
|
Wang Y, Yang J, Chen L, Wang J, Wang Y,
Luo J, Pan L and Zhang X: Artesunate induces apoptosis through
caspase-dependent and -independent mitochondrial pathways in human
myelodysplastic syndrome SKM-1 cells. Chem Biol Interact.
219:28–36. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Papanikolaou X, Johnson S, Garg T, Tian E,
Tytarenko R, Zhang Q, Stein C, Barlogie B, Epstein J and Heuck C:
Artesunate overcomes drug resistance in multiple myeloma by
inducing mitochondrial stress and non-caspase apoptosis.
Oncotarget. 5:4118–4128. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim C, Lee JH, Kim SH, Sethi G and Ahn KS:
Artesunate suppresses tumor growth and induces apoptosis through
the modulation of multiple oncogenic cascades in a chronic myeloid
leukemia xenograft mouse model. Oncotarget. 6:4020–4035. 2014.
|
|
9
|
Wang ZC, Liu Y, Wang H, Han QK and Lu C:
Research on the relationship between artesunate and Raji cell
autophagy and apoptosis of Burkitt’s lymphoma and its mechanism.
Eur Rev Med Pharmacol Sci. 21:2238–2243. 2017.PubMed/NCBI
|
|
10
|
Zuo W, Wang Z-Z and Xue J: Artesunate
induces apoptosis of bladder cancer cells by miR-16 regulation of
COX-2 expression. Int J Mol Sci. 15:14298–14312. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jeong DE, Song HJ, Lim S, Lee SJ, Lim JE,
Nam DH, Joo KM, Jeong BC, Jeon SS, Choi HY and Lee HW: Repurposing
the anti-malarial drug artesunate as a novel therapeutic agent for
metastatic renal cell carcinoma due to its attenuation of tumor
growth, metastasis, and angiogenesis. Oncotarget. 6:33046–33064.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang LX, Liu ZN, Ye J, Sha M, Qian H, Bu
XH, Luan ZY, Xu XL, Huang AH, Yuan DL, et al: Artesunate exerts an
anti-immunosuppressive effect on cervical cancer by inhibiting PGE2
production and Foxp3 expression. Cell Biol Int. 38:639–646. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen K, Shou L-M, Lin F, Duan WM, Wu MY,
Xie X, Xie YF, Li W and Tao M: Artesunate induces G2/M cell cycle
arrest through autophagy induction in breast cancer cells.
Anticancer Drugs. 25:652–662. 2014.PubMed/NCBI
|
|
14
|
Wang Z, Wang C, Wu Z, Xue J, Shen B, Zuo
W, Wang Z and Wang SL: Artesunate suppresses the growth of
prostatic cancer cells through inhibiting androgen receptor. Biol
Pharm Bull. 40:479–485. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Roh JL, Kim EH, Jang H and Shin D: Nrf2
inhibition reverses the resistance of cisplatin-resistant head and
neck cancer cells to artesunate-induced ferroptosis. Redox Biol.
11:254–262. 2017. View Article : Google Scholar :
|
|
16
|
Pang Y, Qin G, Wu L, Wang X and Chen T:
Artesunate induces ROS-dependent apoptosis via a Bax-mediated
intrinsic pathway in Huh-7 and Hep3B cells. Exp Cell Res.
347:251–260. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu L, Zuo LF, Zuo J and Wang J:
Artesunate induces apoptosis and inhibits growth of Eca109 and
Ec9706 human esophageal cancer cell lines in vitro and in vivo. Mol
Med Rep. 12:1465–1472. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen X, Wong YK, Lim TK, Lim WH, Lin Q,
Wang J and Hua Z: Artesunate activates the intrinsic apoptosis of
HCT116 cells through the suppression of fatty acid synthesis and
the NF-kappaB pathway. Molecules. 22:E12722017. View Article : Google Scholar
|
|
19
|
Qin G, Wu L, Liu H, Pang Y, Zhao C, Wu S,
Wang X and Chen T: Artesunate induces apoptosis via a
ROS-independent and Bax-mediated intrinsic pathway in HepG2 cells.
Exp Cell Res. 336:308–317. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
O’Donovan TR, Rajendran S, O’Reilly S,
O’Sullivan GC and McKenna SL: Lithium modulates autophagy in
esophageal and colorectal cancer cells and enhances the efficacy of
therapeutic agents in vitro and in vivo. PLoS One. 10:e01346762015.
View Article : Google Scholar
|
|
21
|
Coker-Gürkan A, Arisan ED, Obakan P,
Akalın K, Özbey U and Palavan-Unsal N: Purvalanol induces
endoplasmic reticulum stress-mediated apoptosis and autophagy in a
time-dependent manner in HCT116 colon cancer cells. Oncol Rep.
33:2761–2770. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ranjan A and Srivastava SK: Penfluridol
suppresses pancreatic tumor growth by autophagy-mediated apoptosis.
Sci Rep. 6:261652016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Luo Q, He X, Wei H, Wang T, Shao J
and Jiang X: Emodin induces apoptosis of colon cancer cells via
induction of autophagy in a ROS-dependent manner. Oncol Res. 2017
Jul;25 View Article : Google Scholar
|
|
24
|
Rosenthal PJ: Artesunate for the treatment
of severe falciparum malaria. N Engl J Med. 358:1829–1836. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Clark RL: Embryotoxicity of the
artemisinin antimalarials and potential consequences for use in
women in the first trimester. Reprod Toxicol. 28:285–296. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Oh KJ, Barbuto S, Pitter K, Morash J,
Walensky LD and Korsmeyer SJ: A membrane-targeted BID BCL-2
homology 3 peptide is sufficient for high potency activation of BAX
in vitro. J Biol Chem. 281:36999–37008. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Siddiqui WA, Ahad A and Ahsan H: The
mystery of BCL2 family: Bcl-2 proteins and apoptosis: An update.
Arch Toxicol. 89:289–317. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Martinou JC and Green DR: Breaking the
mitochondrial barrier. Nat Rev Mol Cell Biol. 2:63–67. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chaitanya GV, Steven AJ and Babu PP:
PARP-1 cleavage fragments: Signatures of cell-death proteases in
neurodegeneration. Cell Commun Signal. 8:312010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mizushima N: Autophagy: Process and
function. Genes Dev. 21:2861–2873. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hanada T and Ohsumi Y: Structure-function
relationship of Atg12, a ubiquitin-like modifier essential for
autophagy. Autophagy. 1:110–118. 2005. View Article : Google Scholar
|
|
32
|
Kang MR, Kim MS, Oh JE, Kim YR, Song SY,
Kim SS, Ahn CH, Yoo NJ and Lee SH: Frameshift mutations of
autophagy-related genes ATG2B, ATG5, ATG9B and ATG12 in gastric and
colorectal cancers with microsatellite instability. J Pathol.
217:702–706. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang Y, Chen P, Hong H, Wang L, Zhou Y
and Lang Y: JNK pathway mediates curcumin-induced apoptosis and
autophagy in osteosarcoma MG63 cells. Exp Ther Med. 14:593–599.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jung HJ, Kang JH, Choi S, Son YK, Lee KR,
Seong JK, Kim SY and Oh SH: Pharbitis Nil (PN) induces apoptosis
and autophagy in lung cancer cells and autophagy inhibition
enhances PN-induced apoptosis. J Ethnopharmacol. 208:253–263. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hseu YC, Tsai TJ, Korivi M, Liu JY, Chen
HJ, Lin CM, Shen YC and Yang HL: Antitumor properties of Coenzyme
Q0 against human ovarian carcinoma cells via induction of
ROS-mediated apoptosis and cytoprotective autophagy. Sci Rep.
7:80622017. View Article : Google Scholar :
|
|
36
|
Cave DD, Desiderio V, Mosca L, Ilisso CP,
Mele L, Caraglia M, Cacciapuoti G and Porcelli M:
S-Adenosylmethionine-mediated apoptosis is potentiated by autophagy
inhibition induced by chloroquine in human breast cancer cells. J
Cell Physiol. 233:1370–1383. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Huang CC, Lee CC, Lin HH, Chen MC, Lin CC
and Chang JY: Autophagy-regulated ROS from xanthine oxidase acts as
an early effector for triggering late mitochondria-dependent
apoptosis in cathepsin S-targeted tumor cells. PLoS One.
10:e01280452015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Won SJ, Yen CH, Liu HS, Wu SY, Lan SH,
Jiang-Shieh YF, Lin CN and Su CL: Justicidin A-induced autophagy
flux enhances apoptosis of human colorectal cancer cells via class
III PI3K and Atg5 pathway. J Cell Physiol. 230:930–946. 2015.
View Article : Google Scholar
|
|
39
|
Liu L, Zhao WM, Yang XH, Sun ZQ, Jin HZ,
Lei C, Jin B and Wang HJ: Effect of inhibiting Beclin-1 expression
on autophagy, proliferation and apoptosis in colorectal cancer.
Oncol Lett. 14:4319–4324. 2017. View Article : Google Scholar : PubMed/NCBI
|