Introduction
The accumulation of amyloid β (Aβ) is a
histopathological hallmark of Alzheimer's disease (AD) (1). Substantial evidence suggests that
astroglial-mediated interstitial fluid (ISF) bulk flow, known as
the paravascular pathway, may contribute to a large portion of Aβ
clearance (2,3). In the paravascular pathway,
subarachnoid cerebrospinal fluid (CSF) driven by vasomotion rapidly
recirculates through the brain along paravascular spaces
surrounding cerebral arteries. ISF and interstitial solutes are
cleared through the paravascular spaces surrounding cerebral veins
(2,4,5).
The astroglial water channel protein aquaporin-4 (AQP4) is key in
the paravascular pathway (2).
AQP4 deficiency or dysfunction significantly impairs the function
of the paravascular pathway. In the aging brain, the function of
AQP4 decreases due to the increasing reactivity of astrocytes,
thereby leading to a 40% reduction in Aβ clearance by the
paravascular pathway (3).
The secreted glycoprotein slit guidance ligand 2
(Slit2) was first identified as an axonal repellent in the
development of the central nervous system (CNS) through interaction
with four cognate roundabout (Robo) receptors, Robo1-4 (6). The interactions between Slit2 and
its receptors is context dependent, creating a multifunctional
platform for cell-cell or cell-matrix interactions, impacting cell
migration, polarity and adhesion (7). Slit2 has been reported to have
beneficial and detrimental effects in diseases of the brain. For
example, in the ischemic brain, Slit2 is secreted by astrocytes as
an autocrine or paracrine molecule interacting with Robo, which
reduces immune cell recruitment to ischemic tissue and mediates
neuroprotection (8). The function
of Slit2 in neuro-inflammation is closely associated with reactive
astrocytes (9). By contrast, the
overexpression of Slit2 increases the permeability of the blood
brain barrier (BBB), which is associated with AD-like alterations
in animals (10,11). As disruption of the BBB and
inflammation are closely linked to aging-related neurodegenerative
disease (12,13), it is necessary to examine the role
of Slit2 in the pathogenesis of neurodegenerative diseases.
In the present study, using Slit2 overexpression
transgenic mice (Slit2-Tg mice), the role of Slit2 in maintaining
the function of the paravascular pathway in the aging mouse brain
was evaluated, and the effects of Slit2 on reducing the risk of
neurodegenerative diseases were examined.
Materials and methods
Animals
All animal experiments in the present study were
approved by the Institutional Animal Care and Use Committee of
Guangdong Laboratory Animals Monitoring Institute (Guangzhou,
China; IACUC no. 2015023). All procedures were performed in
accordance with the AAALAC guidelines (14).
The Slit2-Tg mice overexpressing human Slit2 were
from Guangdong Pharmaceutical University (Guangzhou, China), as
previously described (15). The
heterozygous transgenic mice were crossed with C57BL/6 mice (Stock
no. 000664; Jackson Laboratory, Ben Harbor, ME, USA) to generate
Slit2-Tg mice and wild-type littermates (WT mice). Unless otherwise
noted, the animals used in the present study defined as aging were
15-month-old adult male mice. All mice were provided with water and
a standard chow diet ad libitum. The mice were housed in a
specific pathogen-free facility with a 12 h light/dark cycle at
23±2°C and 50±10% humidity.
The transgenic offspring were identified by
polymerase chain reaction (PCR) using the following primer
sequences: Slit2 forward 5′-CCC TCC GGA TCC TTT ACC TGT CAA GGT
CCT-3′ and Slit2 reverse 5′-TGG AGA GAG CTC ACA GAA CAA GCC ACT
GTA-3′ (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA); the product size was 645 bp. In all experiments, the animals
were anesthetized with chloral hydrate (4.2%, 0.01 ml/g).
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
Following CO2 euthanasia, mouse brains
were removed and total RNA extraction using TRIzol (Invitrogen;
Thermo Fisher Scientific, Inc.) and RT was performed using the
PrimeScript™ RT reagent kit (Takara Bio, Inc., Otsu, Japan) at 37°C
for 30 min and 85°C for 1 min, according to the manufacturer's
protocol. The primers used for Slit2 were provided by Invitrogen;
Thermo Fisher Scientific, Inc. and were as follows: Forward, 5′-AGC
CGA GGT TCA AAA ACG AGA-3′ and reverse, 5′-GGC AGT GCA AAA CAC TAC
AAG A-3′ (Invitrogen; Thermo Fisher Scientific, Inc.). The qPCR
analysis was performed as follows: 95°C for 30 sec followed by 40
cycles of 95°C for 5 sec and 60°C for 34 sec, using an ABI-7500
Fast Real-Time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.) in a total volume of 25 µl containing 2
µM primers, 80 µg cDNA and 12.5 µl of the
SYBR® Premix Ex Taq™ (Takara Bio, Inc.). The results
were quantified using the 2−ΔΔCq method (16).
In vivo 2-photon fluorescence
imaging
The fluorescent CSF tracer penetrating into the
intact living brain was visualized through a closed cranial window
using a custom-built 2-photon laser scanning microscope (Leica
Microsystems GmbH, Wetzlar, Germany), as described previously
(17,18). Briefly, the animals were
anesthetized and securely placed on a stereotaxic device with the
skull level between the bregma and lambda. The stereotaxic
coordinates of the right parietal cortex were 2 mm caudal from the
bregma, and 1.7 mm lateral from the midline (19). A thin cranial window over the
parietal area was prepared for 2-photon in vivo imaging. As
described by Ren et al (20), FITC-conjugated dextran (40 kDa;
D-1841; Thermo Fisher Scientific, Inc.) was first reconstituted
with artificial CSF into a concentration of 0.5%. This was injected
it into the subarachnoid CSF via cisterna magna puncture at a rate
of 2 µl/min for a period of 5 min with a microsyringe pump
controller, following which 200 µl of dextran rhodamine B
(70 kDa; D-1841; Thermo Fisher Scientific, Inc.) was injected
intravenously immediately prior to imaging to visualize the
vasculature. The mice were immediately transferred to the Leica
DM6000 CFS stage (Leica Microsystems GmbH) and their head
immobilized.
Imaging was performed using a Leica SP5 2-photon
imaging system (Leica Microsystems GmbH) equipped with a
Ti:Sapphire laser (Coherent Chameleon Ultra II; Coherent, Inc.,
Santa Clara, CA, USA) and 25X/0.95 NA water-immersion objective and
controlled with Leica LAS X 3.3.0 software (Leica Microsystems
GmbH). Images of the cerebral vasculature were first captured with
512×512 pixel frames from the surface to a depth of 300 µm
with 2-µm z-steps, to avoid the influence of laser
irradiation. Quantification of the mean pixel intensity of the
tracer was processed from 40 µm (xyz) of the 3-dimensional
image stacks obtained 5, 15, 30, 45, and 60 min following
intra-cisternal injection. To examine the dissipation of tracer in
the paravascular space of penetrating arterioles, the 2-photon
in vivo image of 100-µm below the surface was
captured. Donut-shaped region of interests (ROIs) were defined and
the mean pixel intensity of the tracer within these ROIs in the
paravascular space was quantified. The para-arteriolar ROIs at each
time point in each animal were separately averaged to generate
values for a single parameter.
Immunofluorescence
For histological evaluation, the mice were
transcardially perfused with 50 ml of ice-cold saline followed by
200 ml of 4% (w/v) formaldehyde in PBS. The brains were removed,
immersed in 20–30% sucrose solution overnight and embedded in
Tissue-Tek Optimal Cutting Temperature compound (Sakura
Finetechnical, Co., Ltd., Tokyo, Japan) at −20°C. Brain tissue
blocks were consecutively cryosectioned into 10-µm thick
cross sections with a cryostat microtome (Leica Microsystems
GmbH).
For immunofluorescence staining, the frozen sections
were treated with 0.3% triton and 10% anti-donkey serum (Abcam,
Cambridge, UK) for 1 h at room temperature. Subsequently, the
sections were incubated overnight at 4°C in the dark with the
following primary antibodies: Mouse anti-glial fibrillary acidic
protein (GFAP; cat. no. MAB360), rabbit anti-AQP4 (cat. no.
AB3068), mouse anti-Aβ1-40 (cat. no. MABN11), rabbit anti-Aβ1-42
(cat. no. AB5078P; all 1:500; EMD Millipore, Billerica, MA, USA).
Alexa Fluor® 488-conjugated immunoglobulin G (heavy and
light chain), F(ab)2 Fragment antibodies were used as
secondary antibodies and incubated with the membrane at 37°C for 1
h in the dark. These secondary antibodies were anti-mouse (cat. no.
4409) for GFAP, anti-rabbit (cat. no. 4412) for AQP4 and Aβ1-40,
and anti-rabbit (cat. no. 4413; all 1:300; Cell Signaling
Technology, Inc., Danvers, MA, USA) for Aβ1-42. All sections were
mounted with DAPI as a nuclear stain. A Leica TCS SP5 Spectral
one-photon microscope (Leica Microsystems GmbH) was used to acquire
immunofluorescent staining data. The excitation powers were 5 mW
for IgG Alexa Fluor 488 and 0.1 mW for IgG Alexa Fluor 555. The
photomultiplier tube value was 800 V without offset. All
immunofluorescence staining was repeated three times. All images
were captured at the same exposure time.
Quantitative analysis for polarization of
AQP4 water channel
The polarization of astrocytic AQP4 was evaluated in
accordance with a previous study (21). The color channels in the
histological sections labeled for GFAP and AQP4 were separated, and
each image was uniformly captured at two levels (high and a low
stringency thresholds). The low-stringency threshold defined the
overall area of AQP4-immunoreactivity, whereas the high-stringency
threshold defined the area of intense AQP4-immunoreactivty that was
localized to perivascular endfeet. The ratio of the low stringency
area:high stringency area was defined as 'AQP4 polarity'. A higher
AQP4 polarity represented a greater proportion of immunoreactivity
restricted to perivascular regions, whereas a lower proportion
indicated that the distributed immunoreactivity was between the
perivascular endfeet and the soma.
Morris water maze
The Morris water maze experiment was performed
according to the protocols in a previous report by our group
(17). The investigators were
blinded during the experiment. The maze consisted of a circular tub
(120 cm in diameter, 50 cm in height) and a white circular platform
(10 cm). The tub was surrounded by a curtain, which was located ~1
m from the tub wall and painted with distinct geometric cues, the
water (24±1°C) was rendered opaque with white tempera paint to
conceal the platform. Over 4 consecutive days, the platform was
submerged 1 cm under the surface of the water in the center of one
of the pool quadrants. The mice were subjected to four trials (up
to 60 sec) per day from each of the four start locations. Animals
that failed to locate the platform within the allotted 60 sec were
gently guided to the platform. All mice remained on the platform
for 10 sec at the end of each trial. On day 5, the platform was
removed and a single 60 sec probe trial was performed. The swim
paths were recorded using an overhead video camera and tracked by
ANY-maze 6.0 (San Diego Instruments, San Diego, CA, USA). The
velocity during the probe trial, the number of times the target
area (former platform) was crossed and the time spent in each
quadrant during the probe trial were recorded.
Statistical analysis
All data are presented as the mean ± standard
deviation or standard error of the mean. An independent sample
t-test or Wilcoxon rank sum test was used for comparison between
two groups. One-way analysis of variance (ANOVA) or Kruskal-Wallis
test and LSD t-test or Bonferroni test were used for comparison of
mean pixel intensity with the PVS and the latency to the platforms
during the water maze training. SPSS 20.0 (IBM SPSS, Armonk, NY,
USA) software was used for the statistical analysis. Images and
sections were analyzed by an investigator, who was blinded to the
experimental conditions. ImageJ 1.50i (National Institutes of
Health, Bethesda, MD, USA) software was applied for analysis of the
immunohistochemical results. The histology data were analyzed
according to a previous study (22). Briefly, four locations per sample
(three fields per section; six sections per mouse) were used for
analysis. Differences in fluorescent CSF tracer, perivascular GFAP
and polarization of AQP4, Aβ1-40 and Aβ1-42 immunofluorescence
between the Slit2-Tg mice and WT mice were compared using an
unpaired t-test. Differences in the Morris water maze results were
evaluated by one-way ANOVA followed by Tukey's post hoc test for
multiple comparisons. P<0.05 was considered to indicate a
statistically significant difference.
Results
Overexpression of Slit2 restores the
function of the paravascular pathway in the aging brain
Impairment of paravascular pathway function in the
aging brain has an adverse effect on glymphatic CSF recirculation
(3). To investigate the effect of
Slit2 on paravascular pathway function in the aging brain, the
present study verified whether Slit2 was expressed in the mouse
brain using RT-qPCR analysis, the results of which showed the
overexpression of Slit2 in the brain of the Slit2-Tg mice, compared
with the WT mice (Fig. 1A).
Following this, the dynamics of the paravascular CSF-ISF exchange
in vivo were evaluated by 2-photon microscopy and the
intra-cisternal injection of fluorescent CSF tracer
(FITC-conjugated dextran, MW 40 kDa). The cerebral vasculature was
visualized through a thinned-skull window over the parietal area
following caudal vein injection of Rhodamine B. As shown in
Fig. 1B, the intra-cisternal
injection of FITC tracer was followed by a distinct paravascular
influx, which moved rapidly into the cortex along penetrating
arterioles and entered the interstitium of the parenchyma. One-way
ANOVA indicated that the quantification of mean pixel intensity of
the 3D image stacks (Fig. 1C) was
significantly different at different time points in the WT group
(F=9.927, P<0.001). The LSD-t test showed that interstitial
accumulation of the tracer appeared in the parenchyma within 5 min
(29.22±12.53) and increased at 15 min (31.34±3.65), although there
was no significant difference from that at 5 min (P>0.05). The
mean pixel intensity of the CSF tracer peaked at ~30 min
(58.50±5.66, P<0.001) following injection in the aging WT mice,
and gradually reduced at 45 min (45.84±8.85, P<0.05) and at 60
min (41.16±4.41, P>0.05). In the Slit2-Tg mice, interstitial
accumulation of the CSF tracer was also observed within 5 min
(41.11±12.66), and peaked at ~15 min (60.75±6.90). Subsequently,
the mean pixel intensity was significantly decreased at 30 min
(39.73±7.77), 45 min (32.60±4.98) and 60 min (19.61±5.22). However,
one-way ANOVA indicated that the mean pixel intensities were not
significantly different from each other (F=1.385, P>0.05). The
independent sample t-test indicated no significant difference in
the pixel intensity at 5 min post-CSF tracer injection (t=1.492,
P>0.05) or between the peak pixel intensity of the CSF tracer
between the WT mice and Slit2-Tg mice (t=0.563, P>0.05).
However, there was significant attenuation of the pixel intensity
of CSF tracer accumulation in the parenchyma of the Slit2-Tg mice
compared with that in the WT mice at 45 min (t=2.917, P<0.05)
and 60 min (t=7.051, P<0.001).
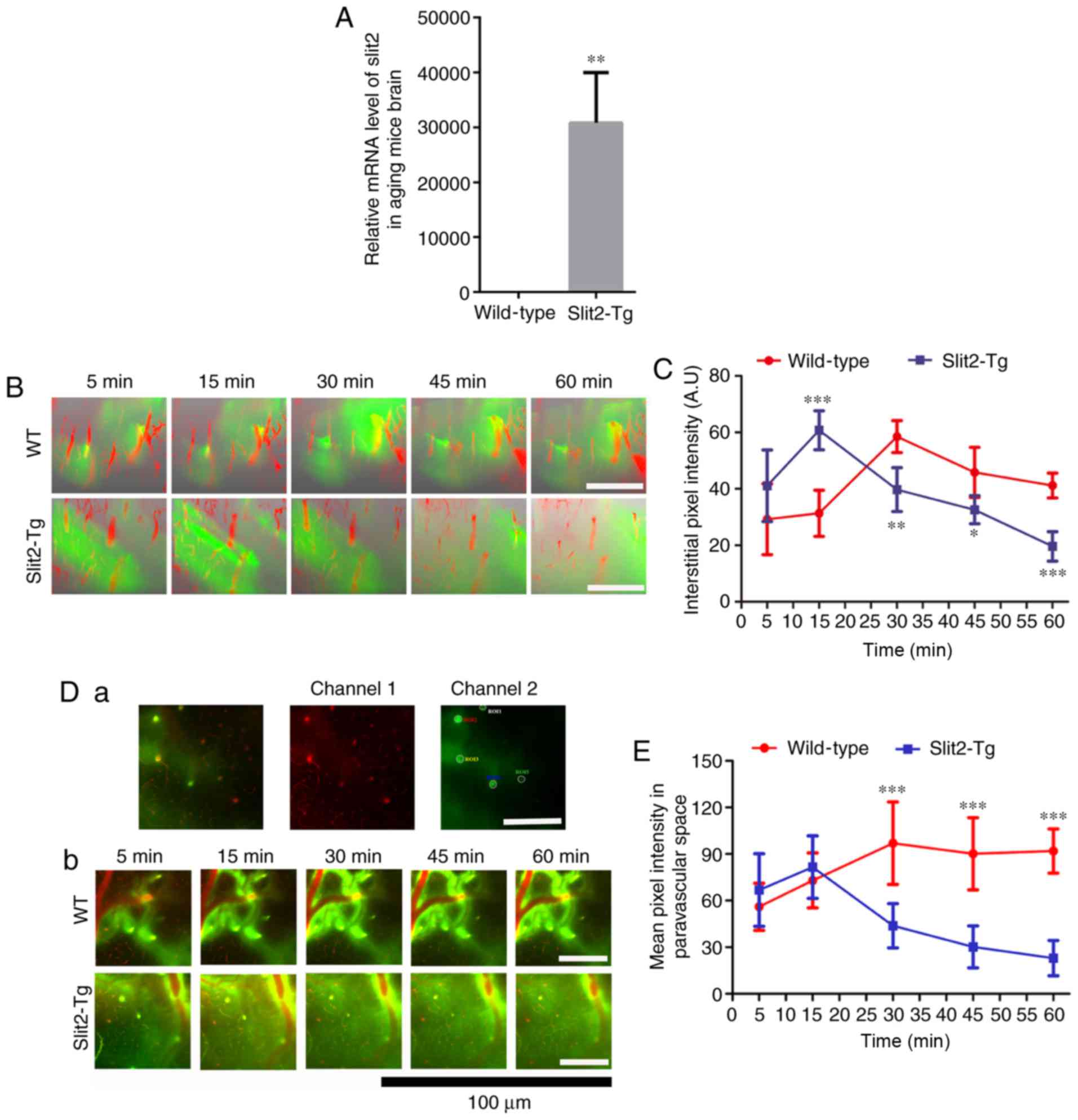 | Figure 1In vivo 2-photon imaging
revealing Slit2 ameliorates paravascular glymphatic CSF
recirculation in aging mice. (A) Relative mRNA level of Slit2 in
the brain of Slit-Tg and WT mice. (B) 3D image stacks of CSF tracer
penetration into the mouse cortex revealed by in vivo
2-photon microscopy following intra-cisternal injection of
FITC-conjugated dextran (green, 40 kDa). Cerebral vasculature was
visualized by intravenous injection of dextran rhodamine B (red, 70
kDa). Magnification, ×250; scale bar=250 µm. (C)
Quantitative analysis of the mean pixel intensity of the tracer in
the 3D image stacks. (D) Accumulation of CSF tracer along
perivascular spaces penetrating into the brain parenchyma,
evaluated by in vivo 2-photon microscopy (a) region of
interest used for analysis (magnification, ×250; scale bar=250
µm); (b) dynamic change of CSF tracer around perivascular
spaces in WT and Slit2-Tg mice (magnification, ×750; scale bar=100
µm). (E) quantitative analysis of the fluorescence intensity
of the CSF tracer. Each value is expressed as the mean ± standard
deviation (*P<0.05, **P<0.01 and
***P<0.001, vs. Slit-Tg group; n=6 per group.).
Slit2, slit guidance ligand 2; CSF cerebrospinal fluid; Tg,
transgenic; WT, wild-type. |
The CSF tracer was analyzed in the perivascular
space of penetrating arteries 100 µm below the cortical
surface (Fig. 1D-a). In the aging
brain of the WT mice, one-way ANOVA indicated that the accumulation
of CSF tracer along perivascular spaces was significantly different
at different time points (F=8.643, P<0.001). The LSD-test showed
that the CSF tracer penetrating into the brain parenchyma was
observed within 5 min (56.03±15.18), increased at 15 min
(72.98±17.68, P<0.05) and peaked at 30 min (96.98±26.53)
(Fig. 1D-b and E, P<0.01). No
significant decrease in the fluorescence intensity of the CSF
tracer was observed at 45 min (90.20±23.20; t=0.667, P>0.05) or
60 min (91.67±14.27). By contrast, the Kruskal-Wallis test
indicated that the accumulation of CSF tracer along perivascular
spaces was significantly different at different time points in the
Slit2-Tg mice (P<0.001). It was present at 5 min (66.83±23.36),
but decreased at 15 min (49.89±20.43) (Fig. 1D-b and E). The fluorescence
intensity of CSF tracer in the paravascular space gradually
decreased at 30 min (34.60±15.29), 45 min (30.21±13.48) and 60 min
(22.96±11.36). Notably, the peak intensity of CSF tracer in the WT
mice was significantly higher than that in the Slit2-Tg mice
(t=0.243, P<0.001). An independent t-test showed that the
fluorescence intensity of the CSF tracer was significantly
increased at 60 min, compared with that at 5 min (t=0.276,
P<0.001) in the aging WT mice, whereas the Wilcoxon rank sum
test on the fluorescence intensity was significantly decreased at
60 min, compared with that at 5 min (P<0.001) in the aging
Slit2-Tg mice. These results indicated that the overexpression of
Slit2 accelerated paravascular CSF-ISF exchange in the aging
brain.
Overexpression of Slit2 inhibits the
reactivity of astrocytes and improves AQP4 polarity
The depolarization of AQP4 in reactive astrocytes is
closely associated with impairment of the paravascular pathway in
the aging brain (3). To
understand why the overexpression of Slit2 restores the function of
the paravascular pathway, the activation of astrocytes in the brain
parenchyma and the polarization of AQP4 were evaluated. As shown in
Fig. 2A, the GFAP-positive
astrocytes were widespread in the cortex and hippocampus of the
aging brain in WT and Slit2-Tg mice. An independent sample t-test
indicated that the mean fluorescence intensity of GFAP-positive
cells was significantly decreased in the Slit2-Tg mice, compared
with that in the WT mice in the cortex (43.21±8.16, vs. 54.21±8.58;
t=0.814, P<0.05; Fig. 2B-a)
and hippocampus (40.02±4.28, vs. 59.08±9.89; t=0.069, P 0.01;
Fig. 2B-b).
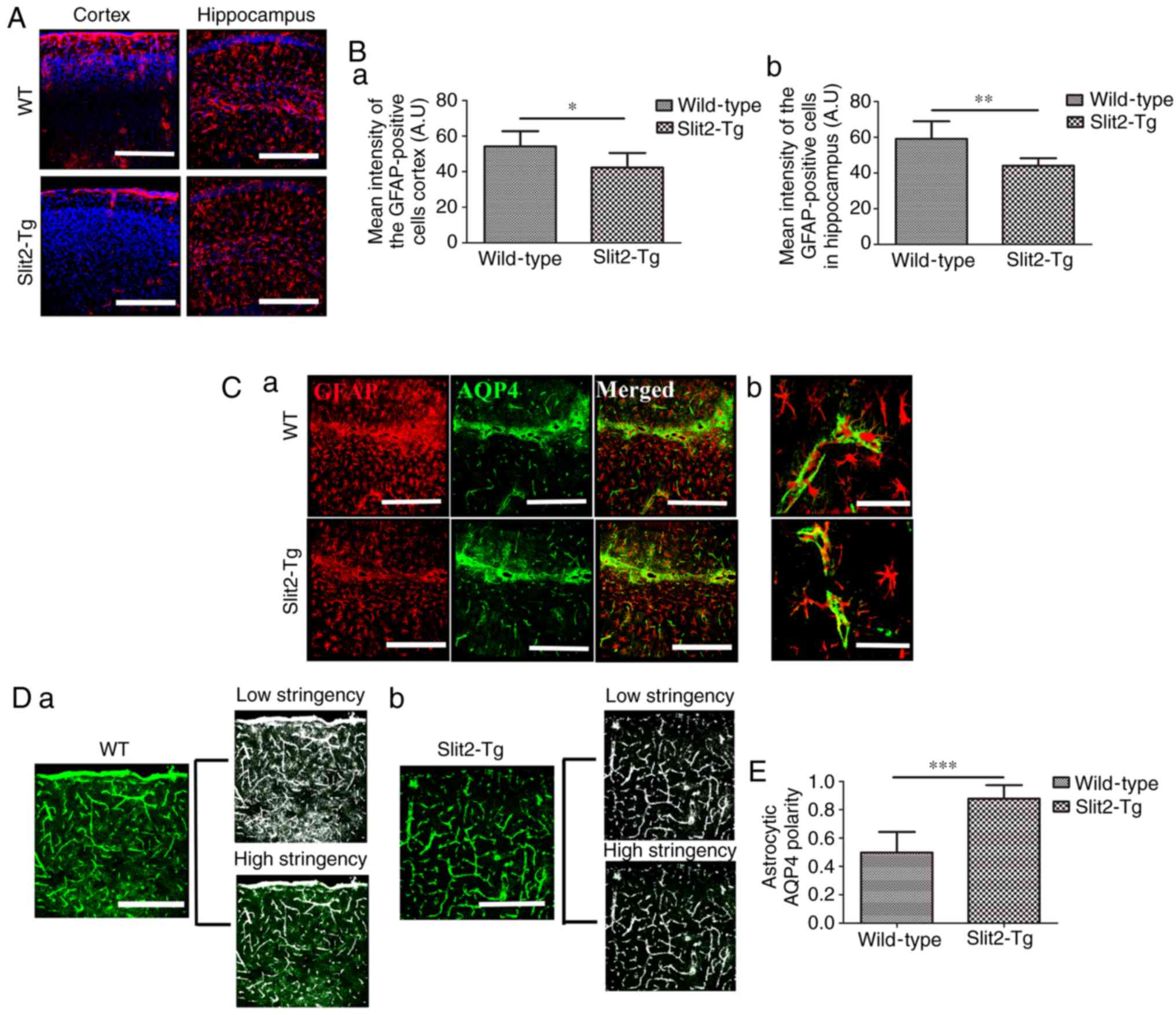 | Figure 2Slit2 inhibits reactivity of
astrocytes and ameliorates AQP4 polarization in the aging mouse
brain. The polarity of AQP4 and reactivity of astrocytes
(GFAP-positive cells) was evaluated by immunofluorescence staining.
(A) GFAP-positive cells were widespread in the cortex and
hippocampus of the aging brains of Slit2-Tg and WT mice
(magnification, ×250; scale bar=250 µm). (B) Quantitative
analysis of the mean pixel intensity of GFAP in the (a) cortex and
(b) hippocampus. (C) Immunofluorescence (a) double-labeling of GFAP
and AQP4 (magnification, ×250; scale bar=250 µm) showed (b)
expression of AQP4 distributed around the astrocytic endfeet, with
less in the astrocytic soma in Slit2-Tg mice, whereas the opposite
was observed in the WT mice (magnification, ×750; scale bar=75
µM). (D) Low stringency images show all AQP4-immunoreactive
pixels in the image, high stringency images captured all pixels
around perivascular endfeet in (a) WT mice and (b) Slit2 mice
(magnification, ×250; scale bar=250 µm). (E) AQP4 polarity
was derived as the ratio of low stringency:high stringency. Each
value is expressed as the mean ± standard deviation.
*P<0.05, **P<0.01 and
***P<0.001; n=6 per group). Slit2, slit guidance
ligand 2; Tg, transgenic; WT, wild-type; GFAP, glial fibrillary
acidic protein; AQP4, aquaporin-4. |
As a main component of water channel proteins
expressed by astrocytes, AQP4 is polarized in the perivascular
astrocytic endfeet in the healthy young brain, but not in the aging
brain. AQP4 delocalization from the endfeet to the soma of
astrocytes is, in part, associated with the failure of the
paravascular pathway (3).
Therefore, the present study investigated the polarization of AQP4
in the aging brain of WT and Slit2-Tg mice (Fig. 2C-a). In the Slit2-Tg mice, the
expression of AQP4 was well distributed around the perivascular
region, where AQP4 sheathed the astrocytic endfeet, and was less
widespread in the astrocytic soma (Fig. 2C-b), whereas AQP4 was mis-located
in the soma of astrocytes in the WT mice (Fig. 2C-b). According to a previous
report (21), the value of AQP4
polarity was analyzed, which was defined as the low stringency area
(overall area of AQP4-immunoreactivity in the image): High
stringency area (area of intense AQP4-immoreactivty localized to
the perivascular endfeet in the image) in the WT mice (Fig. 2D-a) and Slit2-Tg mice (Fig. 2D-b). An independent sample t-test
indicated that astrocytic AQP4 polarity was significantly increased
in the aging Slit2-Tg mice (0.88±0.10), compared with that in the
WT mice (0.50±0.15; t=0.368, P<0.001, Fig. 2E). This result suggested that the
improved paravascular pathway function in the aging brain induced
by the overexpression of Slit2 was accomplished by the enhancement
of astroglial water transport.
Overexpression of Slit2 maintains the
integrity of the BBB in the aging brain
The disruption of the BBB caused by aging results in
loss of vasomotion and decreases the efficiency of paravascular
pathway clearance of Aβ (3,23),
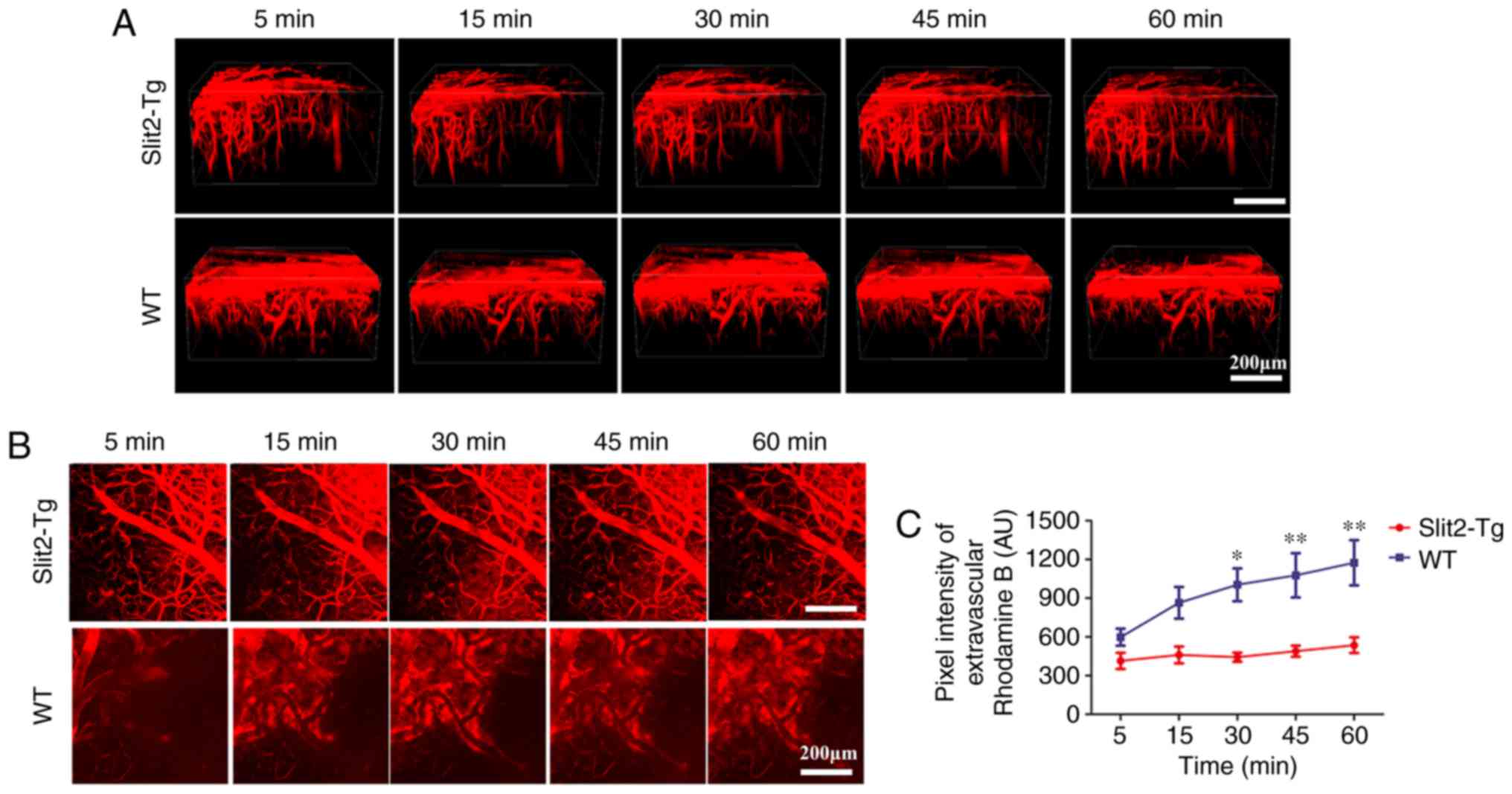
In the present study, the dynamic change of BBB function was
evaluated by in vivo 2-photon microscopy and intravenous
injection of dextran rhodamine B (MW 40 kDa). The 3D image stacks
(Fig. 3A) showed that intravenous
injection of dextran rhodamine B rapidly leaked from blood vessels
into the brain parenchyma of WT mice. However, rhodamine B was
restricted inside the blood vessels of the brain and minimal
leakage was observed in the brain parenchyma of the Slit2-Tg
mice.
To quantify the leakage of rhodamine B from the BBB,
the total fluorescence intensity in the extravascular compartment
was analyzed (24) (Fig. 3B). Two-way repeated ANOVA
indicated no significant interaction between group and time factors
(P>0.05). The main effect of the group and time factors were
significant (F=4.152, P<0.05 and F=41.52, P<0.001,
respectively). Bonfferoni's post hoc test was used to analyze the
fluorescence intensity to examine the BBB permeability. No
significant difference between the WT and Slit 2-Tg mice was
observed at 5 min (598.50±162.11, vs. 414.41±153.84 AU, P>0.05)
or 15 min (864.48±299.30, vs. 460.78±159.32 AU, P>0.05). The
fluorescence intensity in the extravascular compartment was
significantly decreased in the Slit-Tg mice, compared with that in
the WT mice at 30 min (443.08±85.49, vs. 1,004.13±310.60 AU,
P<0.05), 45 min (1,077.08±420.20, vs. 489.39±104.72 AU,
P<0.01) and 60 min (1,174.16±427.65, vs. 536.12±148.46 AU,
P<0.01) (Fig. 3C). These
results indicated that the overexpression of Slit2 maintained the
integrity of the BBB in the aging brain.
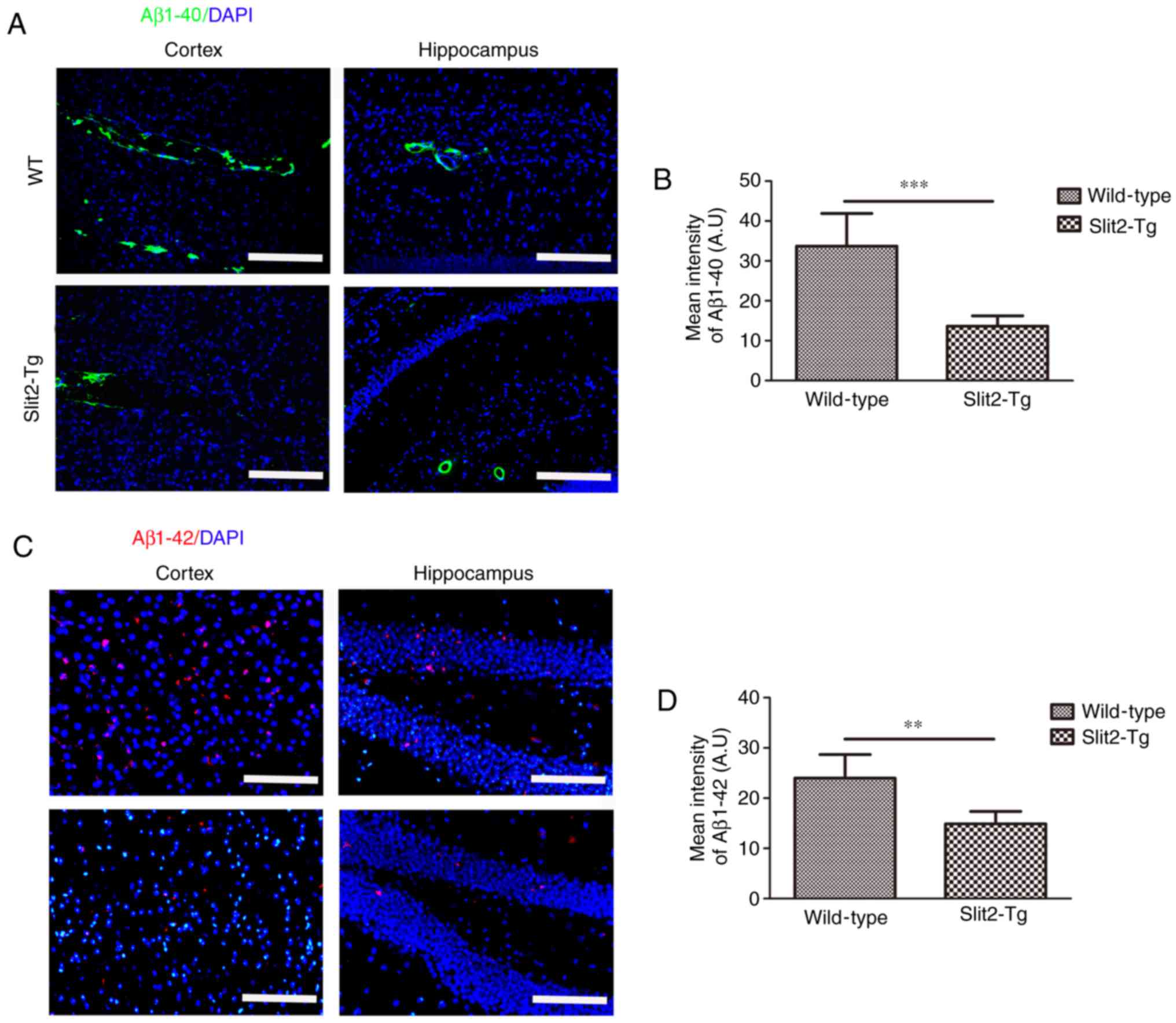
Overexpression of Slit2 reduces the
accumulation of Aβ in the aging brain
The paravascular pathway and interstitial waste
removal are suppressed with aging, which may contribute to the
accumulation of Aβ leading to the pathogenesis of neurodegenerative
diseases, including AD (3). To
evaluate the effect of Slit2 on the accumulation of Aβ,
immunofluorescent staining was performed to analyze the deposition
of Aβ1-42 and Aβ1-40 in the brain parenchyma of aging mice. It was
found that increased Aβ1-40 moved out from the blood vessels of the
WT mice than that in the Slit2-Tg mice in the cortex and
hippocampus (Fig. 4A). An
independent sample t-test indicated that the overall fluorescence
intensity was significantly decreased in the cortex of the Slit2-Tg
mice (13.65±2.57), compared with that of the WT mice (33.70±8.18;
t=5.726, P<0.001) (Fig. 4B).
The accumulation of intra-neuronal Aβ1-42 was further analyzed in
the two regions (Fig. 4C), and
the mean fluorescence intensity of Aβ1-42 was also significantly
decreased in the cortex of the Slit2-Tg mice (14.88±2.47), compared
with that of the WT mice (23.98±4.70; t=4.194, P<0.01) (Fig. 4D). These results indicated that
the over expression of Slit2 restored the function of the
paravascular pathway and reduced Aβ deposition in the aging
brain.
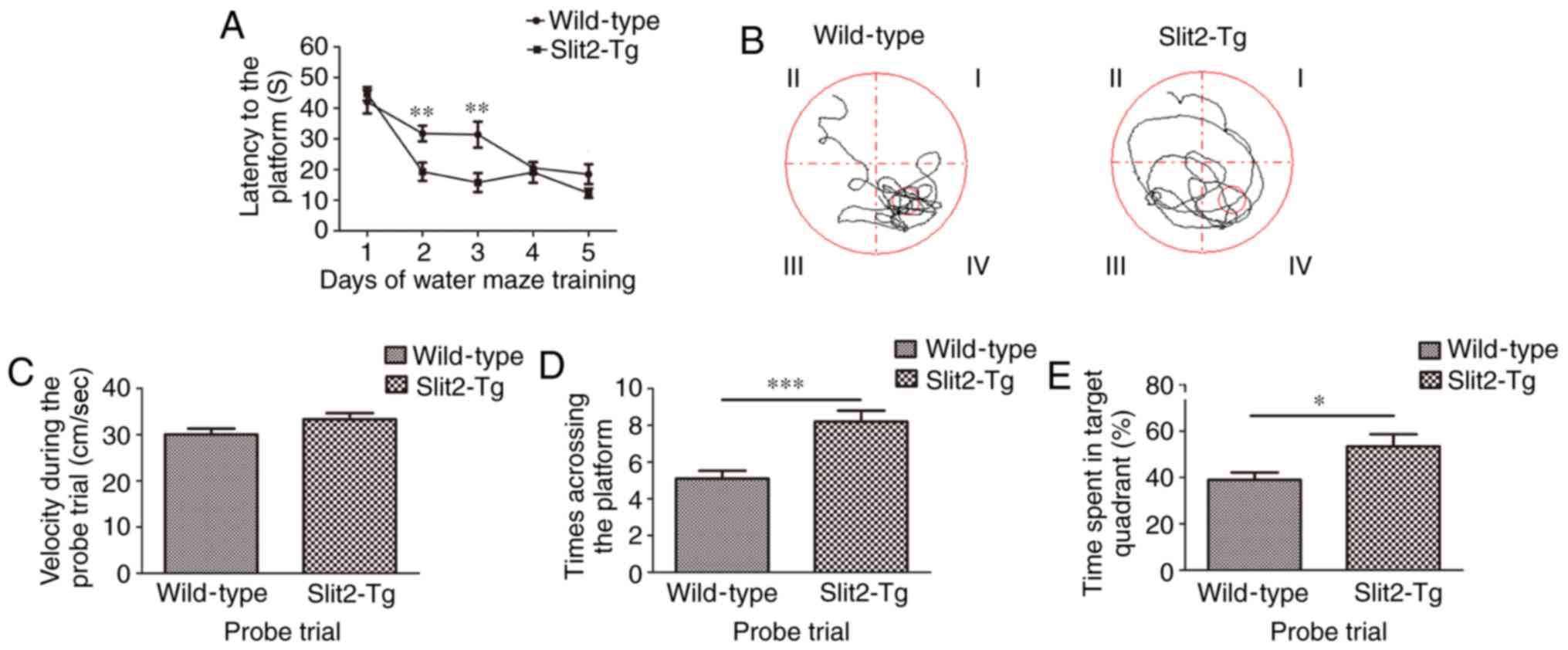
Overexpression of Slit2 improves the
spatial memory cognition of aging mice
To investigate the effect of the overexpression of
Slit2 on the spatial memory cognition of aging mice, Morris water
maze tests were performed. As shown in Fig. 5A, the Wilcoxon rank sum test
revealed no significant difference in the latency to reach
platforms between the WT mice and Slit2-Tg mice (42.097±3.842, vs.
44.739±2.194; P>0.05) on day 1. The Kruskal-Wallis test showed
that the latency on different training days was significantly
different (P<0.001) in the WT group. The Bonferroni test showed
that the latency on day 4 (20.593±1.800) and day 5 (18.499±3.238)
was significantly decreased, compared with that on day 1
(P<0.001; P<0.001); the latency on day 5 was also
significantly decreased, compared with that on day 2 (31.788±2.554)
(P<0.05). However, the latency on day 3 (31.402±4.214) did not
differ significantly from that on the other days (all P>0.05).
In the Slit2-Tg mice, the Kruskal-Wallis test showed that the
latencies on different training days were significantly different
(P<0.001). The Bonferroni test was used to compare with the
latency on day 1. The latency to reach the platform was
significantly lower on day 2 (19.326±2.995; P<0.001), day 3
(15.780±3.102; P<0.001), day 4 (19.108±3.436; P<0.001) and
day 5 (12.327±1.479, P<0.001), however, no significantly
difference was observed among these consecutive 3 days. The latency
was significantly decreased in the Slit2-Tg mice, compared with
that in the WT mice on days 2 and 3 (t=3.166 and 2.985, P<0.01);
whereas no significant differences were observed on days 4 and 5
between the two groups (t=0.383 and 1.734, P>0.05). In the probe
trial (Fig. 5B), the independent
sample t-test indicated no significant difference in velocities
between the WT mice (30.03±1.30 cm/s) and Slit2-Tg mice
(33.308±1.34 cm/s; t=1.753, P>0.05; Fig. 5C), whereas the time to the target
area (previous platform) was significantly increased in the
Slit2-Tg mice (8.20±0.59), compared with that in the WT mice
(5.10±0.433; t=4.223, P<0.001; Fig. 5D). Finally, the time spent in the
target quadrant (%) was analyzed (Fig. 5E), independent sample t-test
indicated that the time spent in the target quadrant (%) was
significantly increased in Slit2-Tg mice (53.417±5.287), compared
with that in WT mice (38.982±3.215; t=2.333; P<0.05). These data
collectively suggested that the overexpression of Slit2 restored
the function of the paravascular pathway, which assisted in
improving spatial memory cognition in the aging mice.
Discussion
The paravascular pathway has a 'glymphatic' role,
responsible for water and waste exchange between the CSF and ISF,
and the clearance of interstitial solutes in the brain (2,5,25).
Dysfunction of the paravascular pathway has been linked to the
accumulation of Aβ (26).
Reactive astrogliosis and neuro-inflammation are prominent features
of aging and the injured brain (3,18,27). Reactive astrocytes directly lead
to a loss of paravascular astroglial AQP4 polarization from the
endfeet to the soma, which is important in maintaining paravascular
pathway function (3,28). Slit2 is widely expressed in
various tissues, including the brain (29). During inflammation, Slit2 inhibits
the secretion of certain inflammatory cytokines/chemokines, which
is mediated by its Robo receptors (30,31). In neuroinflammation, cytokines
have been shown to induce astrocyte activation (32); cytokines and chemokines produced
by activated astrocytes further amplify inflammatory responses in
the brain (33). Although, the
way in which Slit2 reduces aging-related reactive gliosis remains
to be fully elucidated, an early study indicated that Slit2 was
expressed at a high level in GFAP-positive reactive astrocytes
surrounding the necrotic tissue of the injured brain (34). Another study indicated that the
administration of recombinant Slit2 reduces the neuroinflammation
caused by brain injury (35).
Therefore, the effect of Slit2 in improving paravascular pathway
function in the aging brain may be associated with the inhibition
of astrocyte activation by its anti-inflammatory property.
Substantial evidence had shown that Slit2 is
important in promoting vascular stability by inhibiting endothelial
hyper-permeability (31,36,37). Aging induces disruption of the BBB
by increasing endothelial permeability. Disruption of the BBB
results in loss of cerebrovascular contractile function through
interacting with smooth muscle cells (38), and the impairment of vasomotion
decreases the efficiency of paravascular pathway clearance of Aβ
(23). In the present study,
using transgenic mice overexpressing Slit2 in the brain, it was
observed that the integrity of the BBB was maintained and the
accumulation of Aβ was reduced in the brains of aging mice; these
results were in accordance with the improved paravascular pathway
found in the same transgenic aging mice. However, this result was
unexpected, as homozygous Slit2-Tg mice with an intumescent head
have been reported to show increased BBB permeability and Aβ
deposition (10,15). This inconsistency may be due to
differences in the methodology of previous studies, including the
use of homozygous mice, and the use of Evans blue staining for
assessment of the BBB and thioflavin staining for Aβ deposition.
The abnormal phenotype in terms of development of the head was not
observed in heterozygous transgenic mice in the present study,
therefore, using heterozygous mice may avoid the negative effect of
the transgene vector inserting randomly into the genome changing
the expression of other genes. In addition, there are problems in
using Evans blue as a marker for BBB leakage assessment, including
residual dye in brain capillaries, binding of dye to plasma
proteins and spectral shifts (39). In the present study,
fluorescence-labeled dextrans were used for examination of the
dynamic leakage of BBB under in vivo 2-photon microscopy, as
labeled dextrans are considered more suitable for quantification in
tissue (39).
In addition, Aβ deposition was detected using
thioflavin staining in the previous study (15). Thioflavin staining is an easy and
sensitive assay for amyloid. However, its lack of specificity for
amyloid is a major drawback as it may react with several other
proteins. In addition, the autofluorescence of granules, including
elastin fibers and lipofuchsin, may increase the difficulty of data
interpretation. Notably, in the previous study (15), thioflavin staining revealed marked
Aβ deposition in the 9-month-old Tg2576 mice. Tg2576 mice are one
of the widely used mouse models of AD, which overexpresses a mutant
form of APP (isoform 695) with the Swedish mutation (KM670/671NL).
This result is contradictory with general findings that increased
Aβ levels and amyloid plaques in Tg2576 mice are evident at 11–13
months of age (40). By contrast,
using specific antibodies for Aβ1-40 and Aβ1-42, the present study
found that the accumulation of Aβ1-40 and Aβ1-42 was significantly
decreased in the aging brains of Slit2-Tg mice. Aβ, a major
component of senile plaques, has distinctive toxic effects on
neurons and astrocytes. Aβ can induce reactive morphological
changes and the upregulation of GFAP in astrocytes in vitro
(41). In addition, cerebral
amyloid angiopathy results in loss of the perivascular localization
of AQP4 in mouse models and humans with AD (42). Therefore, in the present study,
the decrease of Aβ deposition in the brains of Slit2-Tg mice was
consistent with the inhibition of astroglial reactivity and
maintenance of AQP4 polarity.
In conclusion, the overexpression of Slit2 in the
aging brain improved the function of the paravascular pathway,
maintained the integrity of the BBB, and decreased Aβ accumulation
and age-related impaired spatial memory cognition. Further
understanding of the mechanism underlying the function of Slit2 in
the paravascular pathway of the aging brain may provide a novel
strategy to reduce excess protein waste deposition and delay, or
prevent the onset of neurodegenerative diseases.
Acknowledgments
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China [grant nos. 31472061, 31702074,
81371441, 81371255, 81572230 and 81671102] and the Guangdong
Provincial Science & Technology Project [gran t nos.
2013B060300025, 2013B051000036, 2013B051000018, 2014B020212001,
2014A030304018 and 2014B040404053]. Funding sources had no
involvement in the experimental design or interpretation of
results.
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GL, XH, ZP, RH, YLa and YZ designed the experiments.
XH, HL, YW, YG, SL, YLi and HJ performed the experiments. GL, XH,
ZP, RH, LW and YZ analyzed the data. GL and XH and LW wrote the
paper. RH and LW edited and revised the manuscript. YZ and YLa
reviewed the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal experiments in the present study were
approved by the Institutional Animal Care and Use Committee of
Guangdong Laboratory Animals Monitoring Institute (IACUC no.
2015023). All procedures were performed in accordance with the
AAALAC guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ross CA and Poirier MA: Protein
aggregation and neurodegenerative disease. Nat Med. 10(Suppl):
S10–S17. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Iliff JJ, Wang M, Liao Y, Plogg BA, Peng
W, Gundersen GA, Benveniste H, Vates GE, Deane R, Goldman SA, et
al: A paravascular pathway facilitates CSF flow through the brain
parenchyma and the clearance of interstitial solutes, including
amyloid beta. Sci Transl Med. 4:147ra1112012. View Article : Google Scholar
|
|
3
|
Kress BT, Iliff JJ, Xia M, Wang M, Wei HS,
Zeppenfeld D, Xie L, Kang H, Xu Q, Liew JA, et al: Impairment of
paravascular clearance pathways in the aging brain. Ann Neurol.
76:845–861. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Iliff JJ, Wang M, Zeppenfeld DM,
Venkataraman A, Plog BA, Liao Y, Deane R and Nedergaard M: Cerebral
arterial pulsation drives paravascular CSF-interstitial fluid
exchange in the murine brain. J Neurosci. 33:18190–18199. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Iliff JJ, Lee H, Yu M, Feng T, Logan J,
Nedergaard M and Benveniste H: Brain-wide pathway for waste
clearance captured by contrast-enhanced MRI. J Clin Invest.
123:1299–1309. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Brose K, Bland KS, Wang KH, Arnott D,
Henzel W, Goodman CS, Tessier-Lavigne M and Kidd T: Slit proteins
bind Robo receptors and have an evolutionarily conserved role in
repulsive axon guidance. Cell. 96:795–806. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ypsilanti AR, Zagar Y and Chedotal A:
Moving away from the midline: New developments for Slit and Robo.
Development. 137:1939–1952. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Altay T, McLaughlin B, Wu JY, Park TS and
Gidday JM: Slit modulates cerebrovascular inflammation and mediates
neuroprotection against global cerebral ischemia. Exp Neurol.
207:186–194. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Park JH, Pak HJ, Riew TR, Shin YJ and Lee
MY: Increased expression of Slit2 and its receptors Robo1 and Robo4
in reactive astrocytes of the rat hippocampus after transient
forebrain ischemia. Brain Res. 1634:45–56. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Han HX and Geng JG: Over-expression of
Slit2 induces vessel formation and changes blood vessel
permeability in mouse brain. Acta Pharmacol Sin. 32:1327–1336.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yuen DA and Robinson LA: Slit2-Robo
signaling: A novel regulator of vascular injury. Curr Opin Nephrol
Hypertens. 22:445–451. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Elahy M, Jackaman C, Mamo JC, Lam V,
Dhaliwal SS, Giles C, Nelson D and Takechi R: Blood-brain barrier
dysfunction developed during normal aging is associated with
inflammation and loss of tight junctions but not with leukocyte
recruitment. Immun Ageing. 12:22015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gorlé N, Van Cauwenberghe C, Libert C and
Vandenbroucke RE: The effect of aging on brain barriers and the
consequences for Alzheimer's disease development. Mamm Genome.
27:407–420. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
National Research Council (U.S.),
Committee for the Update of the Guide for the Care and Use of
Laboratory Animals, Institute for Laboratory Animal Research (U.S.)
and National Academies Press (U.S.): Guide for the Care and Use of
Laboratory Animals. National Academies Press; Washington, D.C.:
2011
|
|
15
|
Li JC, Han L, Wen YX, Yang YX, Li S, Li
XS, Zhao CJ, Wang TY, Chen H, Liu Y, et al: Increased permeability
of the blood-brain barrier and Alzheimer's disease-like alterations
in slit-2 transgenic mice. J Alzheimers Dis. 43:535–548. 2015.
View Article : Google Scholar
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
17
|
He XF, Lan Y, Zhang Q, Liu DX, Wang Q,
Liang FY, Zeng JS, Xu GQ and Pei Z: Deferoxamine inhibits
microglial activation, attenuates blood-brain barrier disruption,
rescues dendritic damage and improves spatial memory in a mouse
model of microhemorrhages. J Neurochem. 138:436–447. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Luo C, Yao X, Li J, He B, Liu Q, Ren H,
Liang F, Li M, Lin H, Peng J, et al: Paravascular pathways
contribute to vasculitis and neuroinflammation after subarachnoid
hemorrhage independently of glymphatic control. Cell Death Dis.
7:e21602016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Harvey CD, Coen P and Tank DW:
Choice-specific sequences in parietal cortex during a
virtual-navigation decision task. Nature. 484:62–68. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ren Z, Iliff JJ, Yang L, Yang J, Chen X,
Chen MJ, Giese RN, Wang B, Shi X and Nedergaard M: 'Hit & Run'
model of closed-skull traumatic brain injury (TBI) reveals complex
patterns of post-traumatic AQP4 dysregulation. J Cereb Blood Flow
Metab. 33:834–845. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang M, Iliff JJ, Liao Y, Chen MJ,
Shinseki MS, Venkataraman A, Cheung J, Wang W and Nedergaard M:
Cognitive deficits and delayed neuronal loss in a mouse model of
multiple microinfarcts. J Neurosci. 32:17948–17960. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu H, Wu T, Li M and Wang J: Efficacy of
the lipid-soluble iron chelator 2,2′-dipyridyl against hemorrhagic
brain injury. Neurobiol Dis. 45:388–394. 2012. View Article : Google Scholar
|
|
23
|
Di Marco LY, Farkas E, Martin C, Venneri A
and Frangi AF: Is vasomotion in cerebral arteries impaired in
Alzheimer's disease? J Alzheimers Dis. 46:35–53. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nhan T, Burgess A, Cho EE, Stefanovic B,
Lilge L and Hynynen K: Drug delivery to the brain by focused
ultrasound induced blood-brain barrier disruption: Quantitative
evaluation of enhanced permeability of cerebral vasculature using
two-photon microscopy. J Control Release. 172:274–280. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hladky SB and Barrand MA: Mechanisms of
fluid movement into, through and out of the brain: Evaluation of
the evidence. Fluids Barriers CNS. 11:262014. View Article : Google Scholar
|
|
26
|
Bakker EN, Bacskai BJ, Arbel-Ornath M,
Aldea R, Bedussi B, Morris AW, Weller RO and Carare RO: Lymphatic
Clearance of the Brain: Perivascular, paravascular and significance
for neurodegenerative diseases. Cell Mol Neurobiol. 36:181–194.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Iliff JJ, Chen MJ, Plog BA, Zeppenfeld DM,
Soltero M, Yang L, Singh I, Deane R and Nedergaard M: Impairment of
glymphatic pathway function promotes tau pathology after traumatic
brain injury. J Neurosci. 34:16180–16193. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Verkman AS, Anderson MO and Papadopoulos
MC: Aquaporins: Important but elusive drug targets. Nat Rev Drug
Discov. 13:259–277. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Blockus H and Chedotal A: Slit-Robo
signaling. Development. 143:3037–3044. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
London NR and Li DY: Robo4-dependent Slit
signaling stabilizes the vasculature during pathologic angiogenesis
and cytokine storm. Curr Opin Hematol. 18:186–190. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao H, Anand AR and Ganju RK: Slit2-Robo4
pathway modulates lipopolysaccharide-induced endothelial
inflammation and its expression is dysregulated during endotoxemia.
J Immunol. 192:385–393. 2014. View Article : Google Scholar
|
|
32
|
Pekny M, Pekna M, Messing A, Steinhäuser
C, Lee JM, Parpura V, Hol EM, Sofroniew MV and Verkhratsky A:
Astrocytes: A central element in neurological diseases. Acta
Neuropathol. 131:323–345. 2016. View Article : Google Scholar
|
|
33
|
Farina C, Aloisi F and Meinl E: Astrocytes
are active players in cerebral innate immunity. Trends Immunol.
28:138–145. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hagino S, Iseki K, Mori T, Zhang Y, Hikake
T, Yokoya S, Takeuchi M, Hasimoto H, Kikuchi S and Wanaka A: Slit
and glypican-1 mRNAs are coexpressed in the reactive astrocytes of
the injured adult brain. Glia. 42:130–138. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sherchan P, Huang L, Wang Y, Akyol O, Tang
J and Zhang JH: Recombinant Slit2 attenuates neuroinflammation
after surgical brain injury by inhibiting peripheral immune cell
infiltration via Robo1-srGAP1 pathway in a rat model. Neurobiol
Dis. 85:164–173. 2016. View Article : Google Scholar
|
|
36
|
Jones CA, London NR, Chen H, Park KW,
Sauvaget D, Stockton RA, Wythe JD, Suh W, Larrieu-Lahargue F,
Mukouyama YS, et al: Robo4 stabilizes the vascular network by
inhibiting pathologic angiogenesis and endothelial
hyperpermeability. Nat Med. 14:448–453. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jones CA, Nishiya N, London NR, Zhu W,
Sorensen LK, Chan AC, Lim CJ, Chen H, Zhang Q, Schultz PG, et al:
Slit2-Robo4 signalling promotes vascular stability by blocking Arf6
activity. Nat Cell Biol. 11:1325–1331. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Moody DM, Brown WR, Challa VR, Ghazi-Birry
HS and Reboussin DM: Cerebral microvascular alterations in aging,
leukoaraiosis, and Alzheimer's disease. Ann NY Acad Sci.
826:103–116. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Saunders NR, Dziegielewska KM, Møllgård K
and Habgood MD: Markers for blood-brain barrier integrity: How
appropriate is Evans blue in the twenty-first century and what are
the alternatives? Front Neurosci. 9:3852015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Deacon RMJ: APP-based transgenic models:
The Tg2576 model. Animal Models of Dementia. De Deyn PP and Van Dam
D: Humana Press; Totowa, NJ: pp. 387–398. 2011, View Article : Google Scholar
|
|
41
|
Yang W, Wu Q, Yuan C, Gao J, Xiao M, Gu M,
Ding J and Hu G: Aquaporin-4 mediates astrocyte response to
beta-amyloid. Mol Cell Neurosci. 49:406–414. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wilcock DM, Vitek MP and Colton CA:
Vascular amyloid alters astrocytic water and potassium channels in
mouse models and humans with Alzheimer's disease. Neuroscience.
159:1055–1069. 2009. View Article : Google Scholar : PubMed/NCBI
|