Introduction
Randomized trials have reported that radiotherapy
can substantially reduce cancer recurrence and moderately reduce
cancer mortality; however, cardiac radiation exposure increases the
risk of heart failure, thus limiting the use of radiotherapy
(1,2). Cardiac radiation exposure in a
rodent model resulted in cardiomyocyte hypertrophy, left
ventricular diastolic dysfunction, oxidative stress and myocardial
fibrosis (3). However, the
mechanisms responsible for these effects of radiotherapy remain
unclear. A previous study suggested that exposure to radiation
affected cardiac physiology-associated gene networks and molecular
signaling in cardiomyocytes, with changes in cellular proliferation
and cell cycle-associated genes and signaling during the early
phase of radiation exposure (4).
Furthermore, cardiomyocyte senescence has been demonstrated to be
involved in cardiac pathological processes (5). Therefore, the present study aimed to
determine whether radiation induced senescence in
cardiomyocytes.
MicroRNAs (miRs) are a class of noncoding RNAs that
have been reported to serve a significant role in gene regulation
by targeting a variety of transcripts. miRs have also recently been
identified as important regulators of cellular senescence and aging
(6). miR-34a was demonstrated to
be involved in the senescence process in cardiomyocytes (7), and exerted a pro-senescence effect
by regulating the sirtuin 1 (SIRT1) signaling pathway (8), which serves a crucial role in the
maintenance of cardiac homeostasis and energy balance, and is
disturbed by radiotherapy (9).
Therefore, the current study examined the role of the miR-34a/SIRT1
signaling pathway in radiation-induced cellular senescence.
Macrophage migration inhibitory factor (MIF) is a
multifunctional cytokine secreted by various tissues, including the
myocardium (10). Recent evidence
has suggested that MIF may have a cardioprotective role in
pathological conditions, including ischemia injury, hypertension
and cellular senescence (11-13). Several mechanisms have been
suggested to account for the MIF-induced biological responses,
including modulation of the energy balance, which is known to be
disrupted in radiation-induced ischemic heart disease (14). Thus, the role of MIF in the
regulation of radiation-induced cellular senescence was also
investigated in the present study.
SIRT1, a member of the SIRT family of class III
histone deacetylases, is one of the targets of miR-34a, known to be
widely involved in the regulation of cellular senescence (15,16). Furthermore, SIRT1 has been
reported to be involved in the development and progression of heart
failure through the regulation of cell senescence-associated
signaling in chemotherapy-induced cardiac damage (17). Consequently, activation of SIRT1
may have a great potential in the treatment of radiation-associated
cardiotoxicity.
Oxidative stress is an important inducer of
cardiomyocyte senescence (18),
with radiation being a major cause of reactive oxygen species (ROS)
production (19). Upregulation of
antioxidant proteins, such as endothelial nitric oxide synthase
(eNOS) and catalase, has been demonstrated to protect against
oxidative stress-mediated senescence (20). Growing evidence has also indicated
that miR-34a functions asa senescence promoter by inhibiting SIRT1
and inducing oxidative stress (21). Therefore, the present study
explored whether radiation-induced oxidative stress exerted a
pro-senescence effect in cardiomyocytes.
In the current study, the aim was to examine whether
radiation induces cardiomyocyte senescence, and whether MIF reduces
cellular susceptibility to radiation-induced senescence by
inhibiting miR-34a expression and oxidative stress. The results
suggested that MIF may represent a promising therapeutic strategy
for radiation-associated cardiac damage.
Materials and methods
Cell culture and treatment
Human cardiomyocytes (HCMs; PromoCell GmbH,
Heidelberg, Germany) were cultured in Dulbecco's modified Eagle's
medium (HyClone; GE Healthcare Life Sciences, Logan, UT, USA)
supplemented with 10% fetal calf serum (HyClone; GE Healthcare Life
Sciences) and 1% penicillin-streptomycin (Beyotime Institute of
Biotechnology, Jiangsu, China) at 37°C with 5% CO2. In
order to induce radiation exposure, 1×105 HCMs growing
on 6-well plates were irradiated in a 15×15 cm2 square
field with 5 Gy (6-MeV electron rays; current, 6 A; dose rate, 2
Gy/min), as described previously (22). HCM was treated with 100 ng/ml
human recombinant MIF (cat. no. 289-MF; R&D Systems, Inc.,
Minneapolis, MN, USA) for 1 h at 37°C.
Cell transfection
HCMs were replated into 6-well plates at a density
of 1×105 cells per well and then transfected with
miR-34a mimic or miR-negative control (NC) mimic (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) using
X-tremeGENE™ HP DNA transfection reagent (Roche Applied Science,
Penzberg, Germany) according to the manufacturer's protocol. The
miRNA sequences are listed in Table
I. Cells were irradiated with 5 Gy, treated with 100 ng/ml MIF
and harvested 48 h after transfection for further analysis. The
transfection efficiency was analyzed by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR).
 | Table ITransfected sequences. |
Table I
Transfected sequences.
| Name | Sequence |
|---|
| miR-34a mimic |
5′-UGGCAGUGUCUUAGCUGGUUGUAACCAGCUAAGACACUGCCAUU-3′ |
| miR-negative
control |
5′-UGUCAGCUUUGGAGCUGGUUGUAACCUAAGAUGCCACCAGCAUU-3′ |
| siRNA-Sirtuin
1 | F:
5′-AAGTACAATCCACTCCGGAATGA-3′ |
| siRNA-non
targeting | F:
5′-CTCTATGAACACCTAGCT-3′ |
Small interfering RNA (siRNA) gene
knockdown
HMCs (1×105) were seeded in 6-well plates
at 30-40% confluence 1 day prior to transfection and reached 70-80%
confluence the following day. Cells were then transfected with 100
nM SIRT1 siRNA and siRNA-non targeting (siRNA-NT; both Invitrogen;
Thermo Fisher Scientific, Inc.) using X-tremeGENE HP DNA
transfection reagent according to the manufacturer's protocol. The
siRNA sequences are listed in Table
I. siRNA-NT was used as the control. After a 6-h incubation at
37°C, the transfection reagent-siRNA mixture was replaced with
fresh growth medium, and the cells were harvested for further
experiments at 48 h after transfection.
Cell proliferation assay
Cell proliferation was assessed using a Cell
Counting Kit-8 assay (CCK-8) according to the manufacturer's
protocol (Beyotime Institute of Biotechnology). Briefly, cells were
seeded at a density of 2×103 cells/well in 96-well
plates and incubated at 37°C for 24, 48 and 72 h, respectively.
CCK-8 solution (10 µl) was added to each well, and the
plates were incubated for 1 h at 37°C. The absorbance of cells at
450 nm was then measured with a microplate reader.
RT-qPCR
Isolated cells were collected for RT-qPCR analysis.
Total RNA was extracted from HCMs using TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the protocol
described by the manufacturer. RNA was then reverse transcribed
into cDNA using Transcriptor First Strand cDNA Synthesis kit (Roche
Applied Science), followed by analysis by qPCR. The primers used in
qPCR for miR-34a, cyclin-dependent kinase inhibitor 1a (Cdkn1a),
Cdkn2c, SIRT1, eNOS, catalase and glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) are shown in Table II. qPCR was conducted using the
FastStart Universal SYBR Green Master (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) and Applied Biosystems Step One Plus Real-Time
PCR system (Thermo Fisher Scientific, Inc.). The thermocycling
conditions were as follows: 40 cycles consisting of 95°C for 10
sec, 60°C for 10 sec and 72°C for 10 sec. Quantification was
performed relative to the levels of the housekeeping gene GAPDH.
Data were analyzed using the 2−ΔΔCq method (23).
| Table IIPrimer sequences used in quantitative
polymerase chain reaction. |
Table II
Primer sequences used in quantitative
polymerase chain reaction.
| Gene | Sequence |
|---|
| microRNA-34a | F:
5′-CAGAGCATCACACGCAAGC-3′ |
| R:
5′-CAGGAAACAGAAACCCCAGC-3′ |
| Cdkn1a | F:
5′-TCACTGTCTTGTACCCTTGTGC-3′ |
| R:
5′-GGCGTTTGGAGTGGTAGAAA-3′ |
| Cdkn2c | F:
5′-CGGGAGGTTCTTGTTCTG-3′ |
| R:
5′-TTTGTTGGCTTGCTTGAC-3′ |
| Sirtuin 1 | F:
5′-CAGAGCATCACACGCAAGC-3′ |
| R:
5′-CAGGAAACAGAAACCCCAGC-3′ |
| Telomere
length | F:
5′-CGGTTTGTTTGGGTTTGGGTTTGGGTTTGGGTTTGGGTT-3′ |
| R:
5′-GGCTTGCCTTACCCTTACCCTTACCCTTACCCTTACCCT-3′ |
| Endothelial nitric
oxide synthase | F:
5′-GCAACCACATCAAGTATGCCACCAA-3′ |
| R:
5′-GCTGTTCCAGATTCGGAAGTCTCCT-3′ |
| Catalase | F:
5′-CAAGCTGGTTAATGCGAATGG-3′ |
| R:
5′-TTGAAAAGATCTCGGAGGCC-3′ |
| GAPDH | F:
5′-CTGCACCACCAACTGCTTAG-3′ |
| R:
5′-AGGTCCACCACTGACACGTT-3′ |
Western blotting
Cells were lysed in ice-cold lysis buffer to obtain
the total protein (Beyotime Institute of Biotechnology), and
protein concentrations were measured using the BCA Protein Assay
kit (Beyotime Institute of Biotechnology). Samples were then
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and blotted onto polyvinylidene fluoride membranes.
The membranes were incubated with SIRT1 (cat. no. 9475) and β-actin
(cat. no. 4970) primary antibodies (both 1:1,000; Cell Signaling
Technology, Inc., Danvers, MA, USA) overnight, followed by
incubation with horseradish peroxidase-conjugated secondary
antibodies (cat. no. 7074; 1:2,000; Cell Signaling Technology,
Inc.). Subsequently, the membranes were visualized using the
BeyoECL Plus (Beyotime Institute of Biotechnology). The stained
protein bands were visualized using a Bio-RadChemiDoc XRS system
and analyzed using Quantity One software (Bio-Rad Laboratories,
Inc., Hercules, CA, USA).
Relative telomere length
The relative telomere length in HCMs was measured
using a qPCR approach, as described previously (24), with GAPDH serving as the
normalizing gene. Table II lists
the primers used for the detection of the telomere length.
Relative telomerase activity
A TeloTAGGG Telomerase PCR ELISA PLUS Assay kit
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was used to analyze
the relative telomerase activity, according to the protocol
provided by the manufacturer.
Senescence-associated β-galactosidase
(SA-β-gal) assay
Cellular senescence was measured by SA-β-gal assay
(Cell Signaling Technology, Inc., Danvers, MA, USA). Briefly, cells
were seeded in a 6-well plate at a density of 2×104
cells/well were washed with phosphate-buffered saline, fixed with
2% paraformaldehyde for 30 min at room temperature and then
incubated with fresh SA-β-gal staining solution, as described
previously (25). SA-β-gal
activity was measured using a microplate reader at a wavelength of
50 µm.
ROS measurement
Intracellular ROS levels were measured using
2,7-dichlorodihydrofluorescein diacetate (Beyotime Institute of
Biotechnology), according to the protocol described by the
manufacturer. A fluorescence spectrophotometer was used to
determine the fluorescence intensity of the cells, at an excitation
and emission wavelength of 488 and 525 nm, respectively.
Lipid peroxidation assays
Malondialdehyde (MDA) levels in HMCs were measured
using the Lipid Peroxidation (MDA) Assay kit (Abcam, Cambridge,
UK). Briefly, 2 ml HMCs at a density of 1×106 cells/ml
were homogenized on ice in 300 µl MDA lysis buffer, followed
by centrifugation at 12,000 × g and 4°C for 1 min to remove
insoluble material. Thiobarbituric acid was then added to the
supernatant and incubated at 95°C for 60 min. The samples were
allowed to cool at room temperature for 10 min, and the absorbance
at 532 nm was measured with a spectrophotometer.
Statistical analysis
Data are expressed as the mean ± standard deviation.
Differences among groups were tested by one-way analysis of
variance, and comparisons between two groups were evaluated by
Student's t-tests using SPSS version 19.0 software (IBM
Corporation, Armonk, NY, USA). A value of P<0.05 was considered
to denote a difference that was statistically significant.
Results
Radiation induces miR-34a expression and
senescence in HCMs
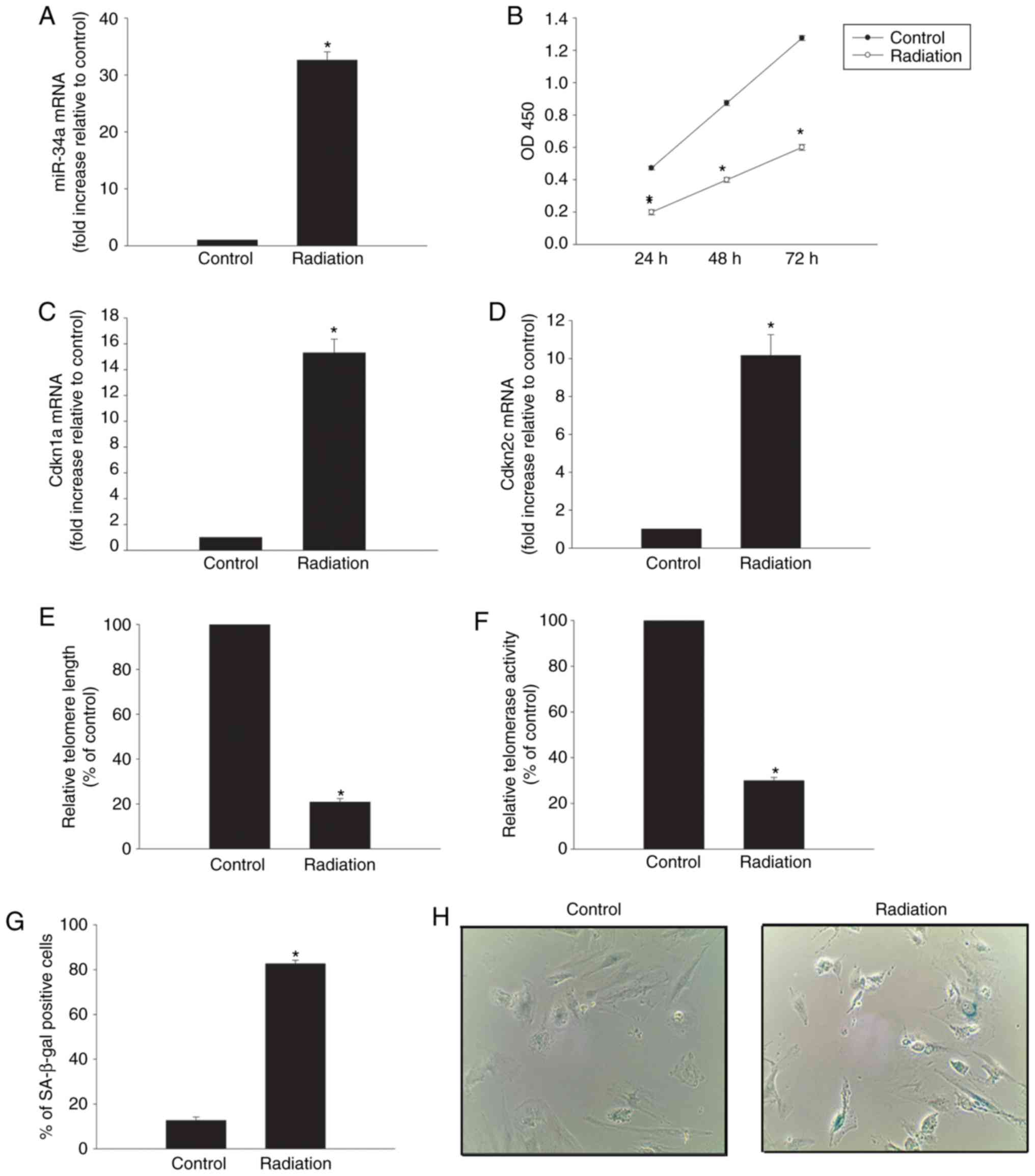
miR-34a expression levels in HCMs were significantly
increased following exposure to radiation (Fig. 1A). Radiation markedly inhibited
cellular proliferation (Fig. 1B)
and increased the expression levels of the cellular
senescence-associated genes Cdkn1a and Cdkn2c (Fig. 1C and D). Radiation exposure also
significantly shortened the relative telomere length (Fig. 1E), impaired the relative
telomerase activity (Fig. 1F) and
increased the percentage of SA-β-gal-positive cells (Fig. 1G and H).
Exogenous MIF alleviates
radiation-induced senescence via inhibition of miR-34a
The anti-senescence effect of MIF was explored by
adding 100 ng/ml MIF to HCMs prior to exposure to radiation.
Exogenous MIF significantly decreased the expression of miR-34a
induced by radiation (Fig. 2A).
Subsequently, the study further examined whether MIF exerted its
anti-senescence effect via inhibition of miR-34a by transfecting
HCMs with a miR-34a mimic to induce the overexpression of miR-34a
(Fig. 2B). MIF treatment in cells
exposed to radiation increased cellular proliferation (Fig. 2C), decreased the expression levels
of the senescence-associated genes Cdkn1a and Cdkn2c (Fig. 2D and E), recovered the impaired
relative telomere length and activity (Fig. 2F and G), and reduced the
percentage of SA-β-gal-positive cells (Fig. 2H and I). However, the effects of
MIF treatment were reversed by miR-34a overexpression in cells
exposed to radiation.
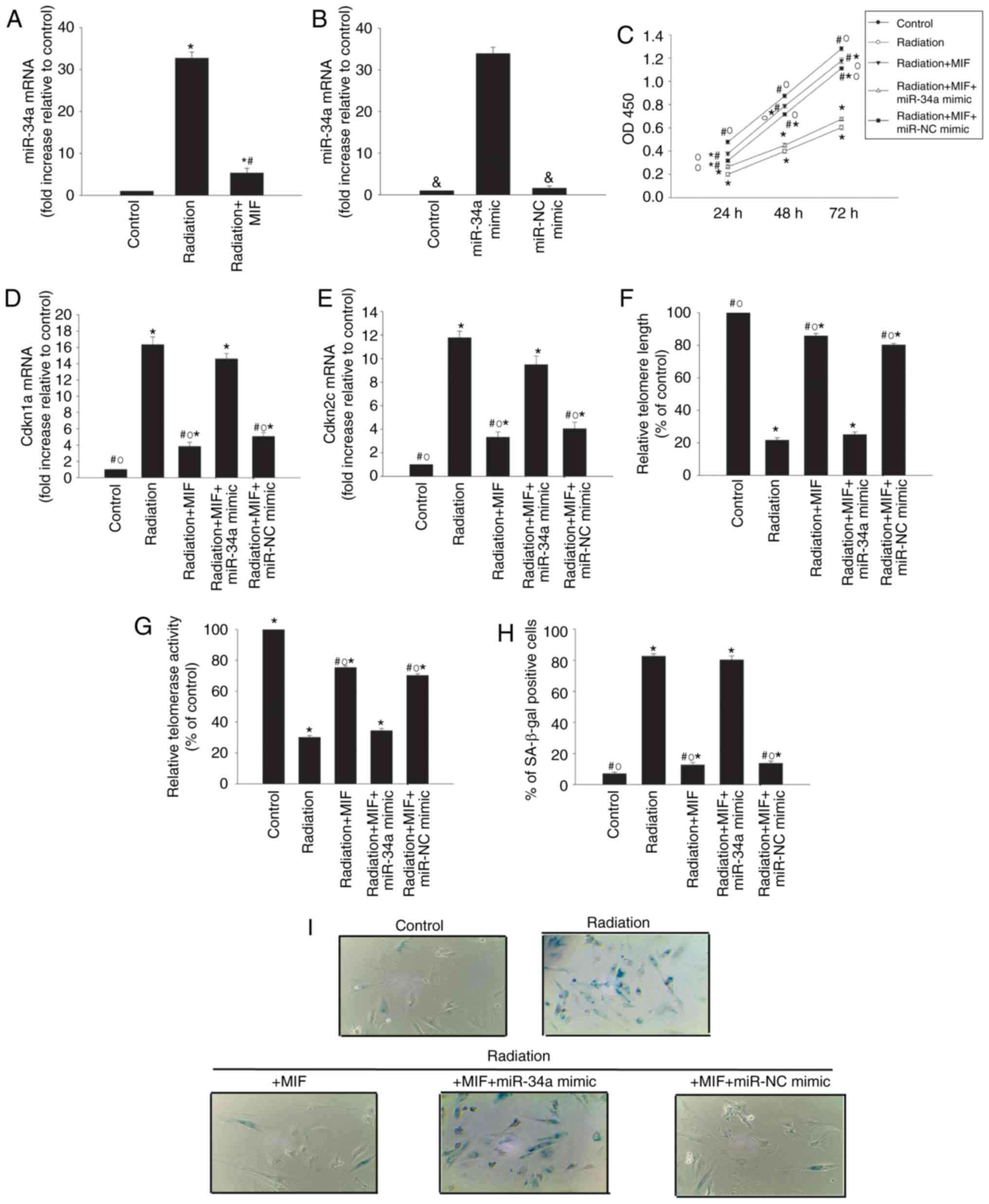 | Figure 2Exogenous MIF alleviates
radiation-induced senescence via inhibition of miR-34a. (A) miR-34a
expression levels in control and radiation-exposed cardiomyocytes
with or without MIF treatment (100 ng/ml), as determined by
RT-qPCR. (B) miR-34a expression levels in untransfected
cardiomyocytes, and cardiomyocytes transfected with miR-34a mimic
or miR-NC mimic, as determined by RT-qPCR. (C) Cell growth curves
determined by Cell Counting Kit-8 assay in cardiomyocytes
transfected with miR-34a mimic or miR-NC mimic, pretreated with MIF
and exposed to radiation. Untreated cardiomyocytes serve as the
control. (D) Cdkn1a, (E) Cdkn2c and (F) relative telomere length
mRNA levels were analyzed by RT-qPCR. (G) Relative telomerase
activity. (H) Percentage of SA-β-gal-positive cells, and (I)
representative images of SA-β-gal staining. Data represent the mean
± standard deviation from three independent experiments.
*P<0.05 vs. control group; #P<0.05 vs.
radiation group; &P<0.05 vs. miR-NC group;
○P<0.05 vs. radiation+MIF+miR-34a mimicgroup. MIF,
macrophage migration inhibitory factor; miR, microRNA; NC, negative
control; RT-qPCR, reverse transcription-quantitative polymerase
chain reaction; Cdkn, cyclin-dependent kinase inhibitor; SA-β-gal,
senescence-associated β-galactosidase. |
Rebalancing of the miR-34a/SIRT1
signaling pathway by MIF is essential for preventing
radiation-associated senescence
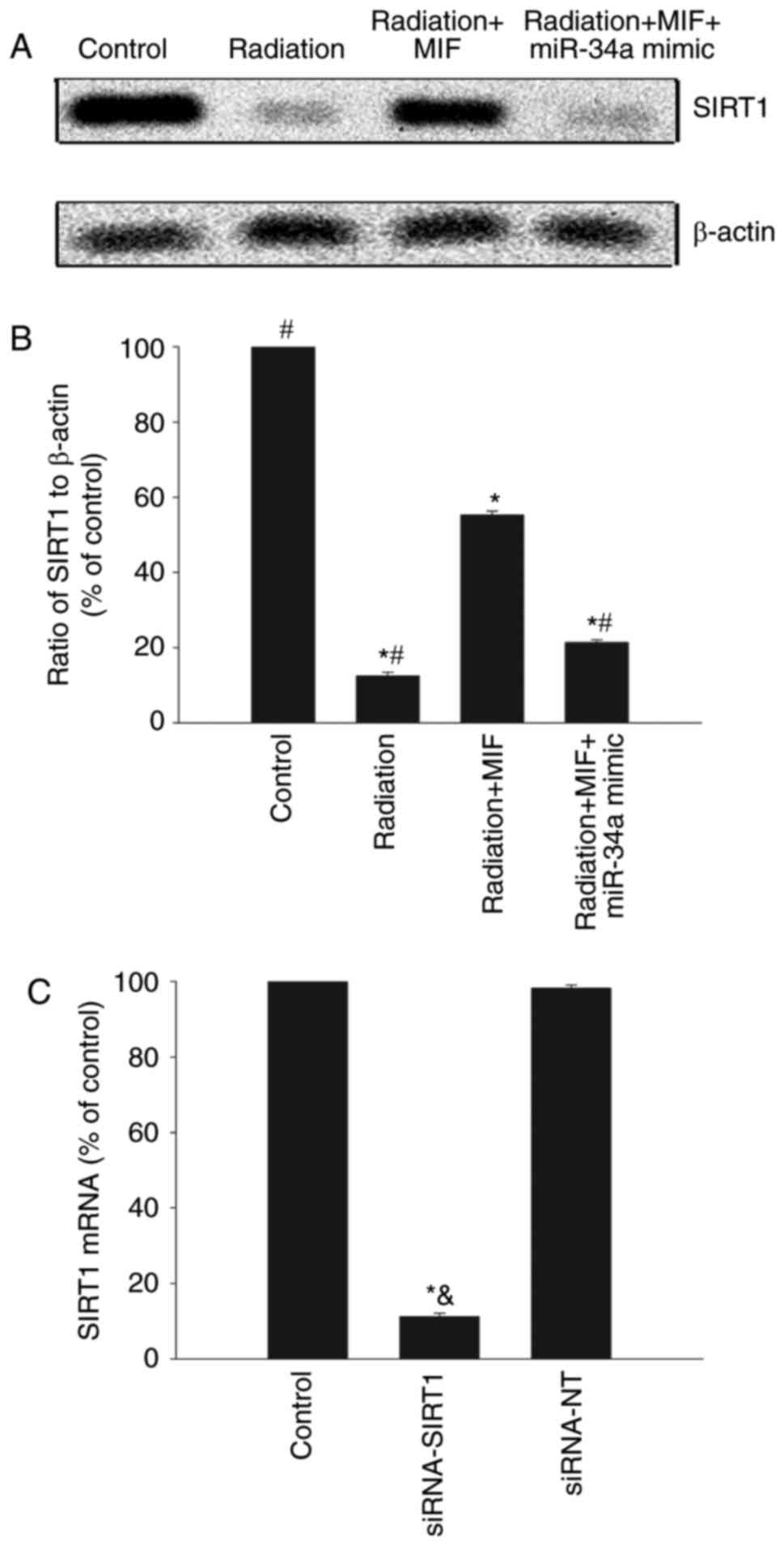
SIRT1 is as a well-known target of miR-34a and is
closely associated with cellular senescence (8). The present study analyzed SIRT1
expression to investigate the miR-34a target and clarify the
mechanism of MIF-mediated suppression of radiation-associated
cellular senescence in HCMs. SIRT1 protein expression was
significantly inhibited by radiation and then recovered by MIF;
however, the MIF-mediated recovery was reversed by miR-34a
overexpression (Fig. 3A and B).
Furthermore, the mechanism responsible for the anti-senescence
effect of MIF in relation to the miR-34a/SIRT1 signaling pathway
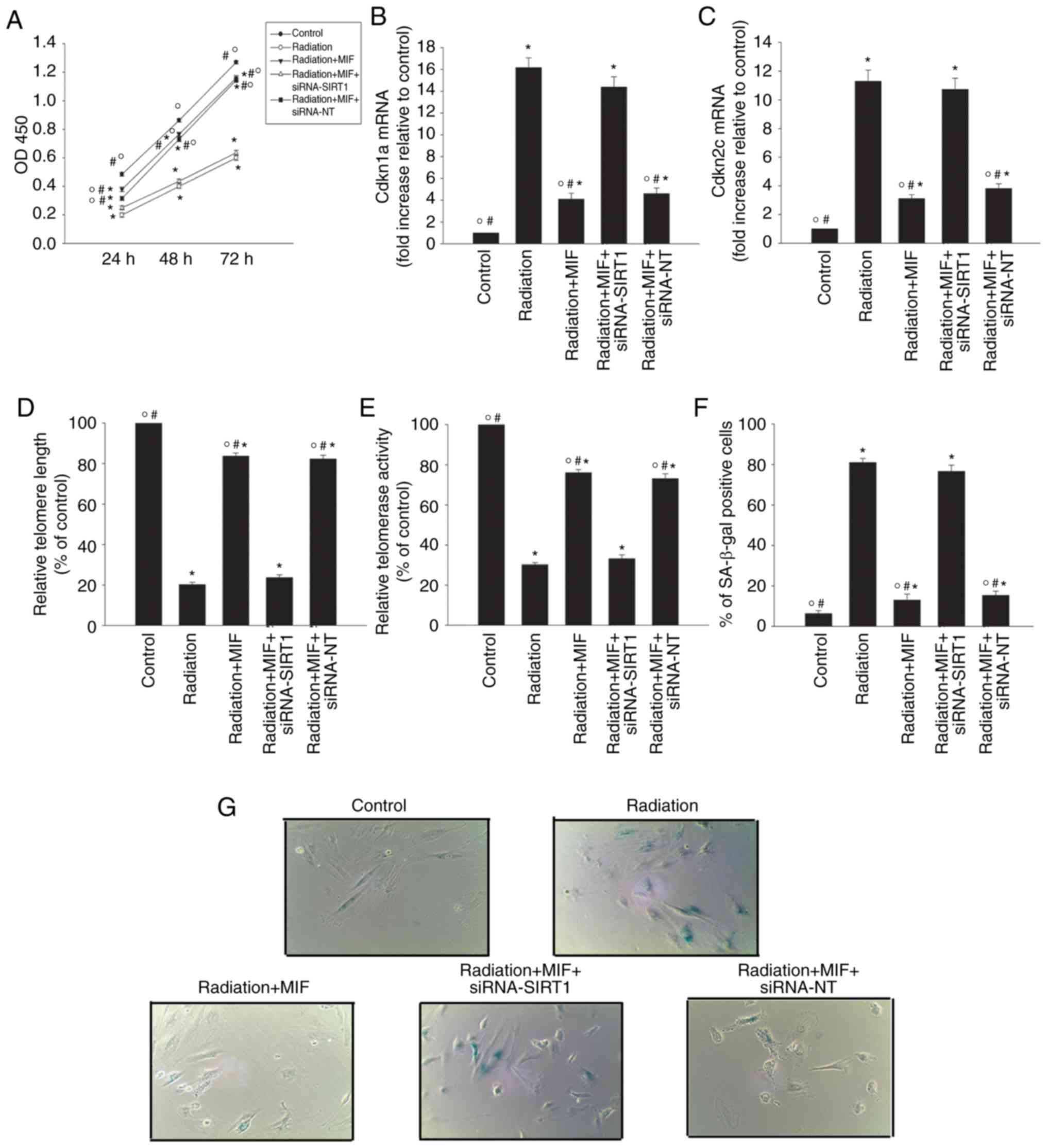
was further explored by silencing SIRT1 using siRNA (Fig. 3C). Downregulation of SIRT1in
MIF-treated radiation-exposed cells decreased the recovery of cell
proliferation induced by MIF (Fig.
4A), induced the expression of the senescence-associated genes
Cdkn1a and Cdkn2c (Fig. 4B and
C), reduced the telomere length and activity (Fig. 4D and E), and increased the
percentage of SA-β-gal-positive cells (Fig. 4F and G), as compared with the
cells treated with only MIF and radiation. By contrast, none of
these effects were observed in HCMs treated with the control,
siRNA-NT.
Modulation of oxidative stress by MIF
inhibits the radiation-induced senescence
The effects of MIF on oxidants and antioxidant gene
expression levels were assessed to determine whether MIF exerted
its anti-senescence effect via modulation of oxidative stress in
cardiomyocytes. Quantitative analysis demonstrated that MIF
inhibited the radiation-induced accumulation of cellular ROS and
generation of MDA in HCMs (Fig. 5A
and B), and reversed the radiation-mediated inhibition of eNOS
and catalase gene expression levels (Fig 5C and D). These antioxidant effects
of MIF were abolished by overexpression of miR-34a or silencing of
SIRT1 (Fig. 5).
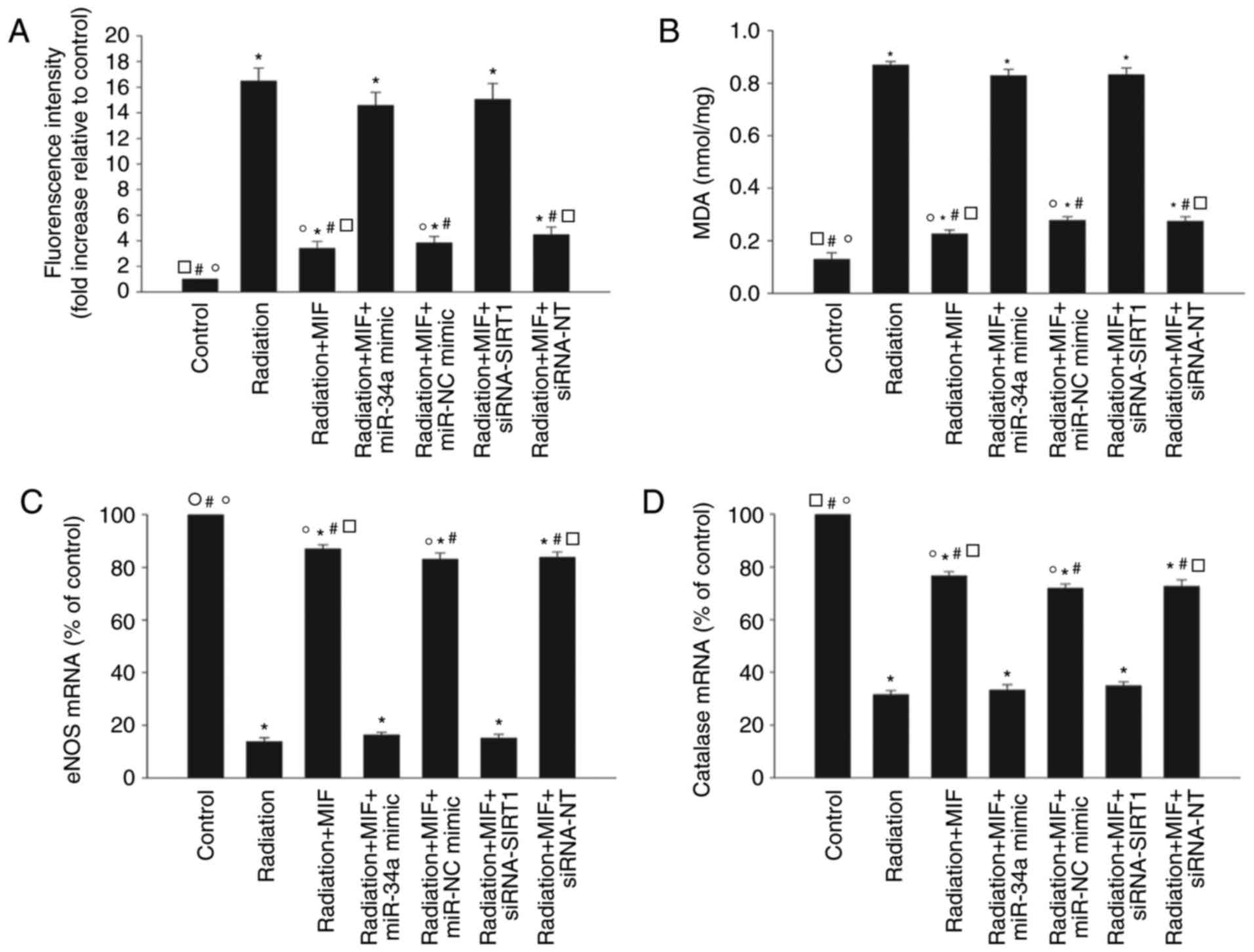 | Figure 5Modulation of oxidant stress by MIF
inhibits radiation-induced senescence. Cardiomyocytes were
transfected with miR-34a mimic, miR-NC mimic, siRNA-SIRT1 or
siRNA-NT, then pretreated with MIF and exposed to radiation.
Untreated cardiomyocytes were used as the control. (A)
Intracellular ROS production analyzed by fluorescence
spectrophotometry. (B) Lipid peroxidation evaluated by MDA
formation. (C) eNOS and (D) catalase mRNA levels measured by
reverse transcription-quantitative polymerase chain reaction. Each
column represents the mean ± standard deviation from three
independent experiments. *P<0.05 vs. control group;
#P<0.05 vs. radiation group; ○P<0.05
vs. radiation+MIF+miR-34a mimic group; □P<0.05 vs.
radiation+MIF+siRNA-SIRT1 group. MIF, macrophage migration
inhibitory factor; miR, microRNA; NC, negative control; siRNA,
small interfering RNA; SIRT1, sirtuin 1; ROS, reactive oxygen
species; MDA, malondialdehyde; eNOS, endothelial nitric oxide
synthase. |
Discussion
Recent progress in radiotherapy over the past two
decades has led to substantial improvements in life expectancy for
cancer patients, however, at the cost of increased risks of
unintended side effects (26).
Radiotherapy has been closely associated with cardiotoxicity and an
increased risk of symptomatic cardiac dysfunction (1,27).
Given the increasing number of long-term survivors following
treatment with radiotherapy, cardiotoxicity has become recognized
as a major concern in oncology (28). However, the mechanisms responsible
for radiotherapy-associated cardiac damage remain unclear. Heart
aging is associated with functional decline and increased
vulnerability to cardiovascular damage, and is the leading cause of
mortality worldwide (29).
Age-associated changes include decreased cardiac output, elevated
susceptibility to arrhythmia and impaired relaxation with increased
myocardial stiffness, all of which are included in the phenotype of
radiotherapy-associated cardiac damage (1,30).
The results of the current study revealed that radiation induced
senescence in HCMs.
miRs have emerged as a new class of modulators of
gene expression. Among these, miR-34a has been implicated in
cardiac-associated damage, particularly cardiac senescence
(31). Researchers recently
demonstrated that miR-34a was involved in modulating
radiation-induced senescence, and that radiation-induced senescence
was correlated with the upregulation of miR-34a expression
(32). Accordingly, the present
study found that radiation exposure induced miR-34a expression in
HCMs, accompanied by cellular senescence.
MIF is known as a prototype cytokine that regulates
macrophage function in inflammation and serves an important role in
cardiac metabolism (33).
Previously, it was confirmed that MIF was highly expressed in
cardiomyocytes and regulated cellular senescence (10). As a promoter of miR-34a, p53 is
associated with accelerated cellular senescence and is inhibited by
MIF (34,35). The results of the present study
revealed that MIF alleviated radiation-associated cellular
senescence through inhibiting miR-34a expression, while
overexpression of miR-34a abolished the anti-senescence effect of
MIF. miR-34a has been reported to trigger senescence partly through
genetic inhibition of SIRT1 (36); therefore, this observation
prompted us to explore the potential role of MIF in modulating
miR-34a-dependent SIRT1 expression in cardiomyocytes. It was
observed that MIF significantly suppressed the radiation-induced
expression of miR-34a, while silencing SIRT1 blocked the
anti-senescence effects of MIF. These results revealed that MIF
reduced cardiomyocyte senescence by interacting with miR-34a and
upregulating SIRT1.
Cellular senescence is defined as the stable cell
cycle arrest elicited in response to a variety of stressors
(37). The accumulation of
oxidative stress in cardiomyocytes has been reported to cause
senescence and reduce cardiac function (18). Radiation-associated oxidative
stress and telomere loss have all been linked to the induction of
the senescent phenotype through replication stress-induced DNA
damage (38,39). The current study demonstrated that
cardiomyocyte senescence was accompanied by radiation-induced
oxidative stress and telomere loss. MIF has been reported to
function as an antioxidant factor (40). Accordingly, the present study
found that MIF significantly inhibited ROS and MDA generation, and
stimulated eNOS and catalase gene expression in cardiomyocytes.
Furthermore, overexpression of miR-34a by transfection of amiR-34a
mimic or silencing SIRT1 abolished the MIF-induced antioxidant gene
expression.
In conclusion, the results of the present study
demonstrated that MIF had a protective role in radiation-induced
cardiac senescence. This cardioprotective effect of MIF may mainly
be attributed to the suppression of oxidative stress by preventing
miR-34a-mediated inhibition of antioxidant gene expression. This
increased understanding of the complex signaling networks involved
in radiation-associated cardiac injuries will help to establish
methods to alleviate cardiac damage induced by radiation.
Funding
The current study was supported by the National
Natural Science Foundation of China (grant nos. 81500261 and
81600278 to MH and WX, respectively) and the Medical Science and
Technology Project of Zhejiang Province (grant no. 2018KY517 to
MH).
Availability of data and materials
All data generated or analyzed during the present
study are included in the published article.
Authors' contributions
YH and WX made substantial contributions to the
acquisition, analysis and interpretation of data. MH was involved
in conception and design of the study, and in drafting the
manuscript. All authors read and approved the final manuscript
Ethics approval and consent to
participate
All animal procedures were approved by the
Institutional Animal Care and Use Committee of Wenzhou Medical
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Taylor C, Correa C, Duane FK, Aznar MC,
Anderson SJ, Bergh J, Dodwell D, Ewertz M, Gray R, Jagsi R, et al:
Estimating the risks of breast cancer radiotherapy: Evidence from
modern radiation doses to the lungs and heart and from previous
randomized trials. J Clin Oncol. 35:1641–1649. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
EBCTCG (Early Breast Cancer Trialists'
Collaborative Group); McGale P, Taylor C, Correa C, Cutter D, Duane
F, Ewertz M, Gray R, Mannu G, Peto R, et al: Effect of radiotherapy
after mastectomy and axillary surgery on 10-year recurrence and
20-year breast cancer mortality: Meta-analysis of individual
patient data for 8135 women in 22 randomised trials. Lancet.
383:2127–2135. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Saiki H, Moulay G, Guenzel AJ, Liu W,
Decklever TD, Classic KL, Pham L, Chen HH, Burnett JC, Russell SJ
and Redfield MM: Experimental cardiac radiation exposure induces
ventricular diastolic dysfunction with preserved ejection fraction.
Am J Physiol Heart Circ Physiol. 313:H392–H407. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Coleman MA, Sasi PS, Onufrak J, Natarajan
M, Manickam K, Schwab J, Muralidharan S, Peterson LE, Alekseyev YO,
Yan X and Goukassian DA: Low dose radiation affects cardiac
physiology: Gene networks and molecular signaling in
cardiomyocytes. Am J Physiol Heart Circ Physiol. 309:H1947–H1963.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Du WW, Yang W, Chen Y, Wu ZK, Foster FS,
Yang Z, Li X and Yang BB: Foxo3 circular RNA promotes cardiac
senescence by modulating multiple factors associated with stress
and senescence responses. Eur Heart J. 38:1402–1412. 2017.
|
|
6
|
Smith-Vikos T and Slack FJ: MicroRNAs and
their roles in aging. J Cell Sci. 125:7–17. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang Y, Cheng HW, Qiu Y, Dupee D, Noonan
M, Lin YD, Fisch S, Unno K, Sereti KI and Liao R: MicroRNA-34a
plays a key role in cardiac repair and regeneration following
myocardial infarction. Circ Res. 7:450–459. 2015. View Article : Google Scholar
|
|
8
|
Fomisonnurse I, Saw EEL, Gandhi S,
Munasinghe PE, Van Hout I, Williams MJA, Galvin I, Bunton R, Davis
P, Cameron V and Katare R: Diabetes induces the activation of
pro-ageing miR-34a in the heart, but has differential effects on
cardiomyocytes and cardiac progenitor cells. Cell Death Differ.
25:1336–1349. 2018. View Article : Google Scholar
|
|
9
|
Hong EH, Lee SJ, Kim JS, Lee KH, Um HD,
Kim JH, Kim SJ, Kim JI and Hwang SG: Ionizing radiation induces
cellular senescence of articular chondrocytes via negative
regulation of SIRT1 by p38 kinase. J Biol Chem. 285:1283–1295.
2010. View Article : Google Scholar :
|
|
10
|
Miller EJ, Li J, Leng L, McDonald C,
Atsumi T, Bucala R and Young LH: Macrophage migration inhibitory
factor stimulates AMP-activated protein kinase in the ischaemic
heart. Nature. 451:578–582. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ma H, Wang J, Thomas DP, Tong C, Leng L,
Wang W, Merk M, Zierow S, Bernhagen J, Ren J, et al: Impaired
macrophage migration inhibitory factor-AMP-activated protein kinase
activation and ischemic recovery in the senescent heart.
Circulation. 122:282–292. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu X, Hua Y, Nair S, Bucala R and Ren J:
Macrophage migration inhibitory factor deletion exacerbates
pressure overload-induced cardiac hypertrophy through mitigating
autophagy. Hypertension. 63:490–499. 2014. View Article : Google Scholar :
|
|
13
|
Xu X, Pacheco BD, Leng L, Bucala R and Ren
J: Macrophage migration inhibitory factor plays a permissive role
in the maintenance of cardiac contractile function under starvation
through regulation of autophagy. Cardiovasc Res. 99:412–421. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Azimzadeh O, Azizova T, Merlpham J,
Subramanian V, Bakshi MV, Moseeva M, Zubkova O, Hauck SM, Anastasov
N, Atkinson MJ and Tapio S: A dose-dependent perturbation in
cardiac energy metabolism is linked to radiation-induced ischemic
heart disease in Mayak nuclear workers. Oncotarget. 8:9067–9078.
2017. View Article : Google Scholar :
|
|
15
|
Haigis MC and Sinclair DA: Mammalian
sirtuins: Biological insights and disease relevance. Annu Rev
Pathol. 5:253–295. 2009. View Article : Google Scholar
|
|
16
|
Morris BJ: Seven sirtuins for seven deadly
diseases of aging. Free Radic Biol Med. 56:133–171. 2013.
View Article : Google Scholar
|
|
17
|
De Angelis A, Piegari E, Cappetta D, Russo
R, Esposito G, Ciuffreda LP, Ferraiolo FA, Frati C, Fagnoni F,
Berrino L, et al: SIRT1 activation rescues doxorubicin-induced loss
of functional competence of human cardiac progenitor cells. Int J
Cardiol. 189:30–44. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang C, Wang J, Ma X, Wang W, Zhao B,
Chen Y, Chen C and Bihl JC: ACE2-EPC-EXs protect ageing ECs against
hypoxia/reoxygenation-induced injury through the miR-18a/Nox2/ROS
pathway. J Cell Mol Med. 22:1873–1882. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Iglesias-Bartolome R, Patel V, Cotrim A,
Leelahavanichkul K, Molinolo AA, Mitchell JB and Gutkind JS: mTOR
inhibition prevents epithelial stem cell senescence and protects
from radiation-induced mucositis. Cell Stem Cell. 11:401–414. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guo Y, Li P, Gao L, Zhang J, Yang Z,
Bledsoe G, Chang E, Chao L and Chao J: Kallistatin reduces vascular
senescence and aging by regulating microRNA-34a-SIRT1 pathway.
Aging Cell. 16:837–846. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lewinska A, Adamczykgrochala J,
Kwasniewicz E, Deregowska A, Semik E, Zabek T and Wnuk M: Reduced
levels of methyltransferase DNMT2 sensitize human fibroblasts to
oxidative stress and DNA damage that is accompanied by changes in
proliferation-related miRNA expression. Redox Biol. 14:20–34. 2018.
View Article : Google Scholar
|
|
22
|
Gu A, Jie Y, Sun L, Zhao S, E M and You Q:
RhNRG-1β protects the myocardium against irradiation-induced damage
via the ErbB2-ERK-SIRT1 signaling pathway. Plos One.
10:e01373372015. View Article : Google Scholar
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
24
|
Crepin T, Carron C, Roubiou C, Gaugler B,
Gaiffe E, Simula-Faivre D, Ferrand C, Tiberghien P, Chalopin JM,
Moulin B, et al: ATG-induced accelerated immune senescence:
Clinical implications in renal transplant recipients. Am J
Transplant. 15:1028–1038. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Filippi-Chiela EC, Bueno e Silva MM, Thomé
MP and Lenz G: Single-cell analysis challenges the connection
between autophagy and senescence induced by DNA damage. Autophagy.
11:1099–1113. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ewer MS and Ewer SM: Cardiotoxicity of
anticancer treatments. Nat Rev Cardiol. 12:547–558. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cohen S, Liu A, Gurvitz M, Guo L, Therrien
J, Laprise C, Kaufman JS, Abrahamowicz M and Marelli AJ: Exposure
to low-dose ionizing radiation from cardiac procedures and
malignancy risk in adults with congenital heart disease.
Circulation. 137:1334–1345. 2018. View Article : Google Scholar
|
|
28
|
Dess RT, Sun Y, Matuszak MM, Sun G, Soni
PD, Bazzi L, Murthy VL, Hearn JWD, Kong FM, Kalemkerian GP, et al:
Cardiac events after radiation therapy: Combined analysis of
prospective multicenter trials for locally advanced non-small-cell
lung cancer. J Clin Oncol. 35:1395–1402. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kaushik G, Spenlehauer A, Sessions AO,
Trujillo AS, Fuhrmann A, Fu Z, Venkatraman V, Pohl D, Tuler J, Wang
M, et al: Vinculin network-mediated cytoskeletal remodeling
regulates contractile function in the aging heart. Sci Transl Med.
7:292ra2992015. View Article : Google Scholar
|
|
30
|
Blice-Baum AC, Zambon AC, Kaushik G,
Viswanathan MC, Engler AJ, Bodmer R and Cammarato A: Modest
overexpression of FOXO maintains cardiac proteostasis and
ameliorates age-associated functional decline. Aging Cell.
16:93–103. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dorn GW II: miR-34a and the cardiomyopathy
of senescence: SALT PNUTS, SALT PNUTS! Cell Metab. 17:629–630.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
He X, Yang A, Mcdonald DG, Riemer EC,
Vanek KN, Schulte BA and Wang GY: MiR-34a modulates ionizing
radiation-induced senescence in lung cancer cells. Oncotarget.
8:69797–69807. 2017.PubMed/NCBI
|
|
33
|
Merk M, Mitchell RA, Endres S and Bucala
R: D-dopachrome tautomerase 1 (D-DT or MIF-2): Doubling the MIF
cytokine family. Cytokine. 59:10–17. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yoshihisa Y, Rehman MU, Kondo T and
Shimizu T: Role of macrophage migration inhibitory factor in
heat-induced apoptosis in keratinocytes. FASEB J. 30:3870–3877.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
von Muhlinen N, Horikawa I, Alam F,
Isogaya K, Lissa D, Vojtesek B, Lane DP and Harris CC: p53 isoforms
regulate premature aging in human cells. Oncogene. 37:2379–2393.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhao T, Li J and Chen AF: MicroRNA-34a
induces endothelial progenitor cell senescence and impedes its
angiogenesis via suppressing silent information regulator 1. Am J
Physiol Endocrinol Metab. 299:E110–E116. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Venkatachalam G, Surana U and Clément MV:
Replication stress-induced endogenous DNA damage drives cellular
senescence induced by a sub-lethal oxidative stress. Nucleic Acids
Res. 45:10564–10582. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mallette FA, Gaumont-Leclerc MF and
Ferbeyre G: The DNA damage signaling pathway is a critical mediator
of oncogene-induced senescence. Genes Dev. 21:43–48. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Purvis JE, Karhohs KW, Mock C, Batchelor
E, Loewer A and Lahav G: p53 dynamics control cell fate. Science.
336:1440–1444. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lin KC, Yip HK, Shao PL, Wu SC, Chen KH,
Chen YT, Yang CC, Sun CK, Kao GS, Chen SY, et al: Combination of
adipose-derived mesenchymal stem cells (ADMSC) and ADMSC-derived
exosomes for protecting kidney from acute ischemia-reperfusion
injury. Int J Cardiol. 216:173–185. 2016. View Article : Google Scholar : PubMed/NCBI
|