Introduction
Atopic dermatitis (AD) is a common chronic skin
disorder that can have a significant impact on the human population
worldwide. AD precedes the development of allergic disorders,
including asthma, rhinitis and food allergy (1,2).
The pathogenesis of AD has been largely attributed to abnormalities
in the adaptive immune system. All AD conditions are characterized
by elevated peripheral eosinophilia counts and increased serum
immunoglobulin E (IgE) levels (3,4).
Proliferating T helper (Th) cells that develop into effector T
cells differentiate into two major subtypes of cells, known as Th1
and Th2 cells. Th1 cells are the host immune effectors against
intracellular bacteria and protozoa. Th1 cells are triggered by
interleukin (IL)-12, and their effector cytokines are interferon-γ
and IL-2. Th2 cells are the host immune effectors against
extracellular parasites, including helminths. They are triggered by
IL-4 and IL-2, and their effector cytokines are IL-4, IL-5, IL-9,
IL-10, IL-13 and IL-25. Peripheral eosinophilia is associated with
Th2 cytokines, including IL-4 and IL-13, which are involved in AD
and other allergic disorders. IL-4 and IL-13 are essential in the
initial phase of tissue inflammation and in the increased
expression of adhesion molecules on endothelial cells (5). These two cytokines are also
responsible for the differentiation of allergen-specific Th2 cells
and the class switching of activated B cells to IgE-producing
cells. The balance of the earliest determined CD4+ T
helper cell subsets, Th1 and Th2, is important in
allergic and autoimmune diseases.
Eosinophils are important in the pathogenesis of
allergic diseases, including asthma (6,7)
and AD (8,9). Their role in the occurrence of
tissue damage during the chronic phase of the disease is
particularly significant. IgE mediates mast cell activation, which
results in the release of preformed histamine and inflammatory
mediators, including chemokines and cytokines (10). In AD skin lesions, CD4+
T cells, eosinophils, mast and dendritic cells are markedly
increased in the dermis; these cytokines promote Th2-type T cell
responses (11) and are found in
skin lesions during the acute phase of AD (12). However, Th1 cells are also
involved. In addition, regulated immune responses are accompanied
by a combination of Th2 and Th1 responses during the chronic phase
of AD (12). AD triggers the
production of pro-inflammatory cytokines, including tumor necrosis
factor (TNF)-α and macrophage-derived cytokines (IL-6) (13). Therefore, the acute and chronic
phases of AD are characterized by the marked infiltration of cells,
particularly, CD4+ T cells, into the region of skin
involved (14).
BALB/c mice exhibit traits similar to those noted in
AD, including an increased level of IgE and chronic dryness. These
mice are generally adopted as animal experimental models for AD. In
addition, this is supported by the effects of
2,4-dinitrochlorobenzene (DNCB) sensitization on the production of
Th1 cytokines and Th2 cytokines (15). DNCB is an organic compound with
the formula
(O2N)2C6H3Cl. It is a
yellow solid that is soluble in organic solvents. DNCB induces a
type IV hypersensitivity reaction in almost all individuals exposed
to it, therefore, it is used medically to assess T cell activity in
patients. This diagnostic test is useful in immunocompromised
patients. DNCB can also be used to treat warts, however, DNCB can
cause contact dermatitis. Previous studies have indicated that
contact allergens can be discriminated from respiratory sensitizers
on the basis of cytokine production in the lymph nodes of BALB/c
mice (16,17). Cervus nippon mantchuricus
extract, which is a deer bone extract known as nok-gol (NGE) in
Korean, is one of the most famous Korean traditional medicines; NGE
has been widely used as a bone nutrient. In our previous study, it
was found that NGE alleviated neutropenia and activated macrophages
in vivo and in vitro (18). In previous studies, it has also
been found that the oral administration of NGE is useful for the
treatment of memory loss (19)
and bone resorption (20), as an
anti-aging treatment (21), for
the activation of macrophages (22), enhancement of immune system
activity (23), and the control
of various inflammatory diseases (24-26). It has been confirmed that NGE is
effective against a variety of inflammatory diseases, therefore, it
is expected to be effective for inflammatory skin diseases,
including AD.
The aim of the present study was to examine whether
NGE suppresses the progression of AD-like skin inflammation in
BALB/c mice. Skin thickness measurements and histological
evaluation, in addition to the estimation of leukocyte levels,
proinflammatory cytokine mRNA levels, and serum total IgE and
cytokine levels, were assessed in the study. The results showed
that NGE significantly inhibited DNCB-induced AD-like skin
inflammation in BALB/c mice. In addition, the anti-inflammatory
effects of NGE on human mast cells were investigated. For this
purpose, the cells were stimulated with phorbol 12-myristate
13-acetate (PMA) + ionomycin to produce proinflammatory cytokines.
PMA is a diester of phorbol and a potent tumor promoter often used
in biomedical investigations to activate the signal transduction
enzyme protein kinase C (PKC). The effects of PMA on PKC result
from its similarity to one of the natural activators of classic PKC
isoforms, diacylglycerol. PMA is a small molecule drug. Ionomycin
is an ionophore produced by the bacterium Streptomyces
conglobatus. It is used to increase the intracellular level of
calcium (Ca2+) and as a tool for understanding
Ca2+ transport across biological membranes. It is also
used to stimulate the intracellular production of the following
cytokines: Interferon, perforin, IL-2 and IL-4, usually in
conjunction with PMA (27). These
cytokines are important in the inflammatory response (27). The results of the present study
demonstrated that NGE inhibited the development of DNCB-induced
AD-like symptoms, suggesting that NGE may be a useful therapeutic
drug for the treatment of AD.
Materials and methods
NGE preparation
The NGE was prepared by Nongshim Corporation (Seoul,
Korea), and contained 88.8% crude protein, 1.9% crude fat, 2.2%
crude ash and 3.0% moisture. The total ganglioside content as
sialic acid in the extract was 0.09%. The total amino acid and free
amino acid contents in the extract were 922.5 and 8.21 mg/g,
respectively. NGE, as a water-extracted white powder, was dissolved
in water for the in vivo and in vitro experiments.
Further information on the NGE production process can be requested
from the company (http://nongshim.co.kr).
Cell culture
The HMC-1 human mast cells were purchased from the
Korea Cell Line Bank (Seoul, Korea). The cell line was grown in
Iscove's modified Dulbecco's medium (Welgene, Daegu, Korea)
supplemented with heat-inactivated 10% fetal bovine serum (FBS;
Welgene) and 1% Penicillin-Streptomycin solution (1X; Welgene) in a
5% CO2 incubator at 37°C.
Cell viability assay
The HMC-1 cells (1×104 cells/well) were
plated in 96-well culture plates and incubated for 24 h.
Subsequently, the HMC-1 cells were treated with 5 ng/ml PMA and 500
ng/ml ionomycin (Sigma, EMD Millipore, Billerica, MA, USA) in the
presence or absence of various concentrations of NGE (5, 25, 100,
250, 500, and 1,000 mg/ml). Following 24 h of incubation, 10 µl of
WST solution was added to each well of the plate, and the plate was
incubated in the dark at 37°C for an additional 1 h. The optical
density was measured at 450 nm using an enzyme-linked immunosorbent
assay (ELISA) plate reader (Versa Max; Molecular Devices LLC,
Sunnyvale, CA, USA).
Animals
Male, 6-week-old BALB/c mice (20±2 g) were purchased
from Orient Bio, Inc. (Sungnam, Korea). The mice were randomized
into three groups: Vehicle (saline), DNCB, and DNCB + NGE, each
comprising six mice. All mice were maintained in a pathogen-free
environment and allowed free access to food and water; they were
maintained on a 12-h light/dark cycle at a temperature of 24°C. All
procedures performed on the mice were approved by the Animal Care
Center of Kyung Hee University [approval no. KHUASP (SE)-16-095].
All methods were performed in accordance with the relevant
guidelines and regulations. At the end of the experiment, the mice
were sacrificed by CO2 inhalation, and cardiac blood was
collected.
Sensitization and treatment
The procedure for the induction of AD-like skin
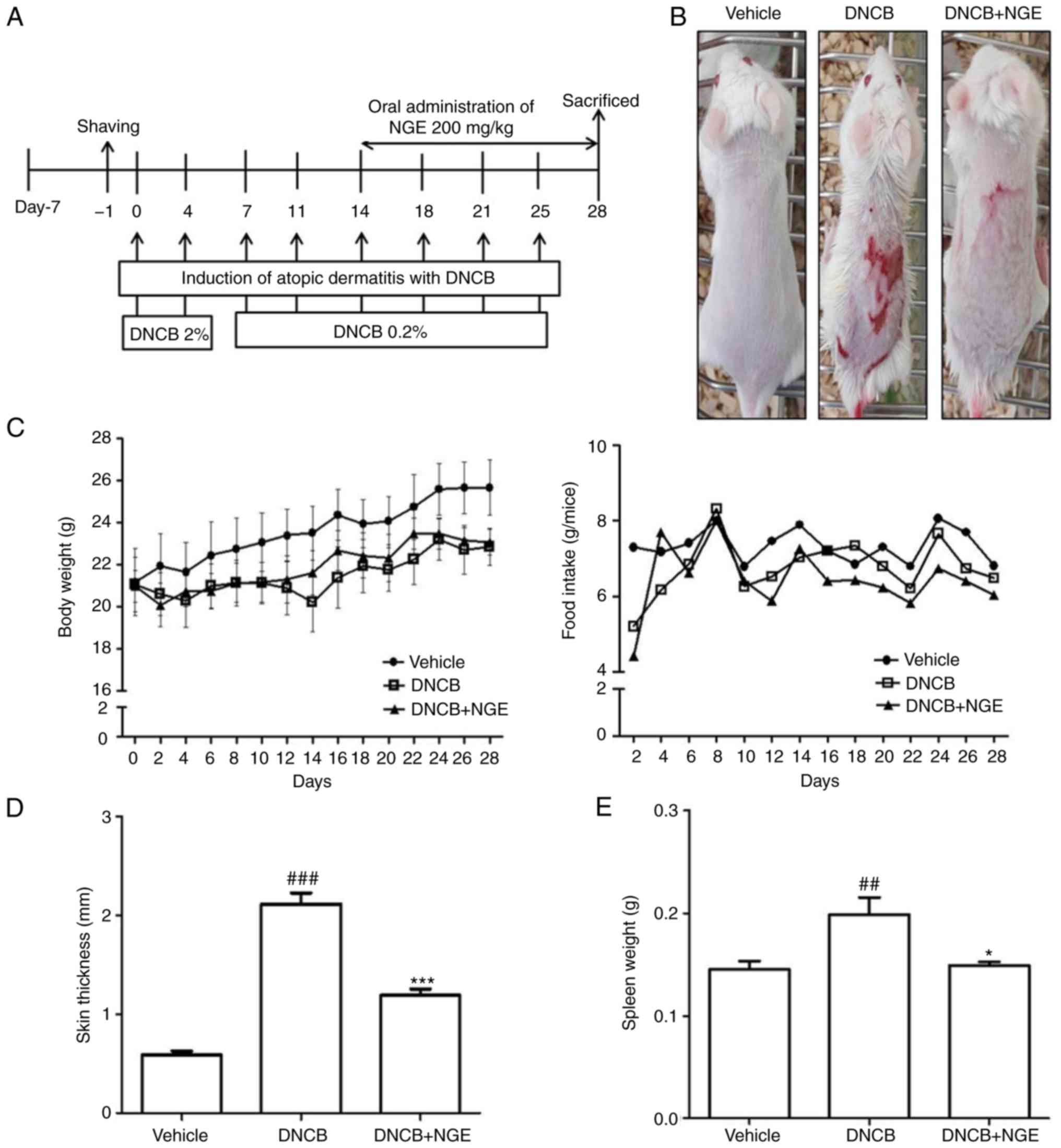
lesions is shown in Fig. 2A. For
this purpose, the dorsal skin of the mouse was shaved and coated
with 100 µl of 2% DNCB in 1×1 cm patches. At 1 week
post-sensitization, the dorsal skin was challenged with 100 µl of
0.2% DNCB solution twice per week (first and fourth day of each
week). The NGE was administered orally to the mice for 2 weeks
following sensitization with DNCB. Following the final application
of NGE, the mice were sacrificed, and immunological and
histological assessments were performed. The method was performed
as described previously (28).
Determination of skin thickness and
spleen weight
The thickness of the dorsal skin was measured with
digital calipers (Mitutoyo, Kawasaki, Japan) prior to sacrifice.
For each mouse, three different sites on the dorsal skin were
measured randomly, and the measurements were averaged. The entire
spleen was removed from the mice following sacrifice and was
weighed.
Histological observation
A section of the skin biopsies was fixed in 4%
paraformaldehyde and embedded in frozen section compound (Surgipath
FSC22 Clear Leica Biosystems GmbH, Wetzlar, Germany) on dry ice.
Skin sections measuring 20 µm in thickness were cut and stained
with hematoxylin and eosin (H&E) to visualize inflammatory
cells (neutrophils and mononuclear cells) or with toluidine blue to
visualize mast cells, and were examined under light microscopy
(Olympus Corporation, Tokyo, Japan). The numbers of mast cells and
inflammatory cells were counted in 10 regions of high-power fields
at ×400 magnification.
Immunohistochemistry
The expression of CD4+ lymphocytes was
detected by immunohistochemical analysis using a mouse monoclonal
anti-CD4+ antibody (cat. no. SC-7219; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). The skin sections were
hydrated. Following microwave treatment, the sections were treated
with 3% hydrogen peroxide in PBS for 15 min to inhibit the
endogenous peroxidase activity of the blood cells. The sections
were blocked with 5% bovine serum albumin (Sigma, EMD Millipore) in
PBS for 1 h at room temperature. The skin sections were incubated
with the mouse monoclonal CD4+ antibody (1:100 dilution)
overnight at 4°C and subsequently incubated with a biotinylated
anti-mouse IgG secondary antibody (1:50 dilution; Vectastain ABC
kit; cat. no. PK-6102; Vector Laboratories, Inc., Burlingame, CA,
USA) for 1 h at room temperature. The sections were treated with
avidin-biotin horseradish peroxidase (HRP) complex (Vectastain ABC
kit; cat. no. PK-4000; Vector Laboratories, Inc., Burlingame, CA,
USA) for 30 min at 4°C and finally stained with diaminobenzidine
tetrachloride (DAB) as a substrate. The slides were mounted with an
aqueous mounting solution (Permount; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) and cover-slipped. All the sections were
analyzed using a ZEISS Scope A1 light microscope (Carl Zeiss AG,
Oberkochen, Germany), and images were captured using a digital
video camera.
Blood analysis
Whole blood samples were collected by cardiac
puncture. The blood was placed in Vacutainer™ tubes containing EDTA
(BD Biosciences, San Jose, CA, USA) by centrifugation (200 × g, 20
min, 4°C). Anticoagulated blood was processed to determine the
hematological parameters of white blood cells (WBCs), lymphocytes,
monocytes, eosinophils, basophils and neutrophils in a HEMAVET 950
hematology system (Drew Scientific, Inc., Dallas, TX, USA) in
accordance with the manufacturer's recommendations.
ELISA
The total IgE levels in the serum of the mice were
determined by sandwich ELISA using the BD Pharmingen mouse IgE and
human ELISA sets (BD Biosciences, San Diego, CA, USA). Briefly, the
plates were coated with capture antibody in ELISA coating buffer
(Sigma, EMD Millipore) and incubated overnight at 4°C. The plates
were then washed with PBS-Tween-20 (0.05%) and subsequently blocked
(10% FBS in PBS) for 1 h at 20°C. Serial dilutions of standard
antigen or sample in dilution buffer (10% FBS in PBS) were added to
the plates, and the plates were incubated for 2 h at 20°C.
Following washing, biotin-conjugated anti-mouse IgE (1:500
dilution) and streptavidin-HRP conjugate (1:250 dilution) were
added to the plates, and the plates were incubated for 1 h at 20°C.
Finally, tetramethylbenzidine substrate solution was added to the
plates for 15 min incubation in the dark, and a 2N
H2SO4 solution was added to terminate the
reaction. The optical densities were measured at 450 nm on an
automated ELISA reader (VersaMax; Molecular Devices LLC). The
levels of IL-10 and IL-12 in the plasma and the levels of IL-4,
IL-13, TNF-α and IL-6 in the cell line were also measured by
sandwich ELISA using the BD Pharmingen mouse and human ELISA sets.
The subsequent procedure followed the same protocol described
above.
Reverse transcription-semi-quantitative
polymerase chain reaction analysis
The cells were harvested by centrifugation (500 × g,
20 min, 4°C) and the pellet was washed with ice-cold PBS. RNA was
isolated from the cells using the Easy-Blue RNA extraction kit
(iNtRON Biotech, Sungnam, Korea) according to the manufacturer's
protocol. The isolated RNA content was measured using a NanoDrop
ND-1000 spectrophotometer (NanoDrop Technologies, Inc., Wilmington,
DE, USA). Total cellular RNA (2 µg) from each sample was reverse
transcribed using a cDNA synthesis kit (Takara Bio, Inc., Otsu,
Japan). The sqPCR analysis was performed with a 20-µl reaction
mixture consisting of DNA template, each gene-specific primer at a
concentration of 10 pM, 10X Taq buffer, 2.5 mM dNTP mixture, and 1
unit of Taq DNA polymerase (Takara Bio, Inc.). PCR was performed
according to the instructions of the Takara Taq kit as follows:
Pre-DNA denaturation at 95°C for 3 min; and 30 cycles of DNA
denaturation at 95°C for 45 sec; annealing for 40 sec at 56°C;
elongation at 72°C for 50 sec. The PCR reaction was performed using
a SimpliAmp Thermal Cycler (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The PCR products were subsequently separated
using agarose gel electrophoresis [5% (w/v)] and were stained with
ethidium bromide at room temperature for 5 min. All experiments
were performed in triplicate. The relative optical density ratio
was calculated using ImageJ software (version 1.42q; National
Institutes of Health, Bethesda, MD, USA) with GAPDH as an internal
control. The primer sequences used for human and/or mouse IL-4,
IL-6, TNF-α and GAPDH are shown in Table I.
 | Table ISequences of primers for polymerase
chain reaction analysis. |
Table I
Sequences of primers for polymerase
chain reaction analysis.
| Primer type | Primer name | Primer
sequence |
|---|
| Mouse | IL-6 | F:
5′-CAAGAGACTTCCATCCAGTTGC-3′ |
| | R:
5′-TTGCCGAGTTCTCAAAGTGAC-3′ |
| IL-4 | F:
5′-TCGGCATTTTGAACGAGGTC-3′ |
| | R:
5′-GAAAAGCCCGAAAGAGTCTC-3′ |
| TNF-α | F:
5′-ATGAGCACAGAAAGCATGATC-3′ |
| | R:
5′-TACAGGCTTGTCACTGGAATT-3′ |
| GAPDH | F:
5′-GAGGGGCCATCCACAGTCTTC-3′ |
| | R:
5′-CATCACCATCTTCCAGGAGCG-3′ |
| Human | IL-6 | F:
5′-AACCTTCCAAAGATGGCTGAA-3′ |
| | R:
5′-CAGGAACTGGATCAGGACTTT-3′ |
| IL-4 | F:
5′-TGCCTCCAAGAACACAACTG-3′ |
| | R:
5′-CTCTGGTTGGCTTCCTTCAC-3′ |
| TNF-α | F:
5′-TGAGCACTGAAAGCATGATCC-3′ |
| | R:
5′-ATCACTCCAAAGTGCAGGAG-3′ |
| GAPDH | F:
5′-CGTCTTCACCACCATGGAGA-3′ |
| | R:
5′-CGGCCATCACGCCACAGTTT-3′ |
Statistical analysis
All experimental results were expressed as the mean
± standard deviation or the mean ± standard error of the mean of at
least three separate tests. P<0.05 was considered to indicate a
statistically significant difference and P<0.05, <0.01, and
<0.001 have been assigned respective symbols in figures.
Statistical significance was determined using a one-way analysis of
variance followed by Tukey-Kramer multiple comparisons post-tests
to analyse differences between groups. Statistical analyses were
performed using PRISM software (version 5.0; GraphPad Software
Inc., La Jolla, CA, USA.).
Results
NGE inhibits the agonist-induced
production of cytokines in HMC-1 cells
Mast cell mediators, including tryptase and
histamine, contribute to the induction of pruritus in AD (29). In the present study, the viability
of HMC-1 cells was examined following treatment with various
concentrations of NGE (Fig. 1A).
NGE did not induce any cytotoxicity in HMC-1 cells over a 24-h
period. The PMA + ionomycin-mediated stimulation of HMC-1 cells has
been shown to significantly increase the production of cytokines
and chemokines (27). The present
study examined the role of NGE in the suppression of cytokine
expression in HMC-1 cells. NGE cotreatment significantly reduced
the agonist-stimulated production of IL-4, IL-13, TNF-α and IL-6 in
the HMC-1 cells (Fig. 1B).
Further experiments were performed to determine whether NGE
regulates the expression of cytokines at the transcriptional level.
RT-qPCR analysis showed that the mRNA levels of IL-4, IL-6 and
TNF-α were induced by agonist treatment and were decreased by NGE
cotreatment in HMC-1 cells (Fig.
1C). In particular, treatment with NGE in a concentration of
250 µg/ml reduced the levels of IL-4 and IL-6 to the control
levels. These results suggested that NGE regulates proinflammatory
cyto-kine production in HMC-1 cells.
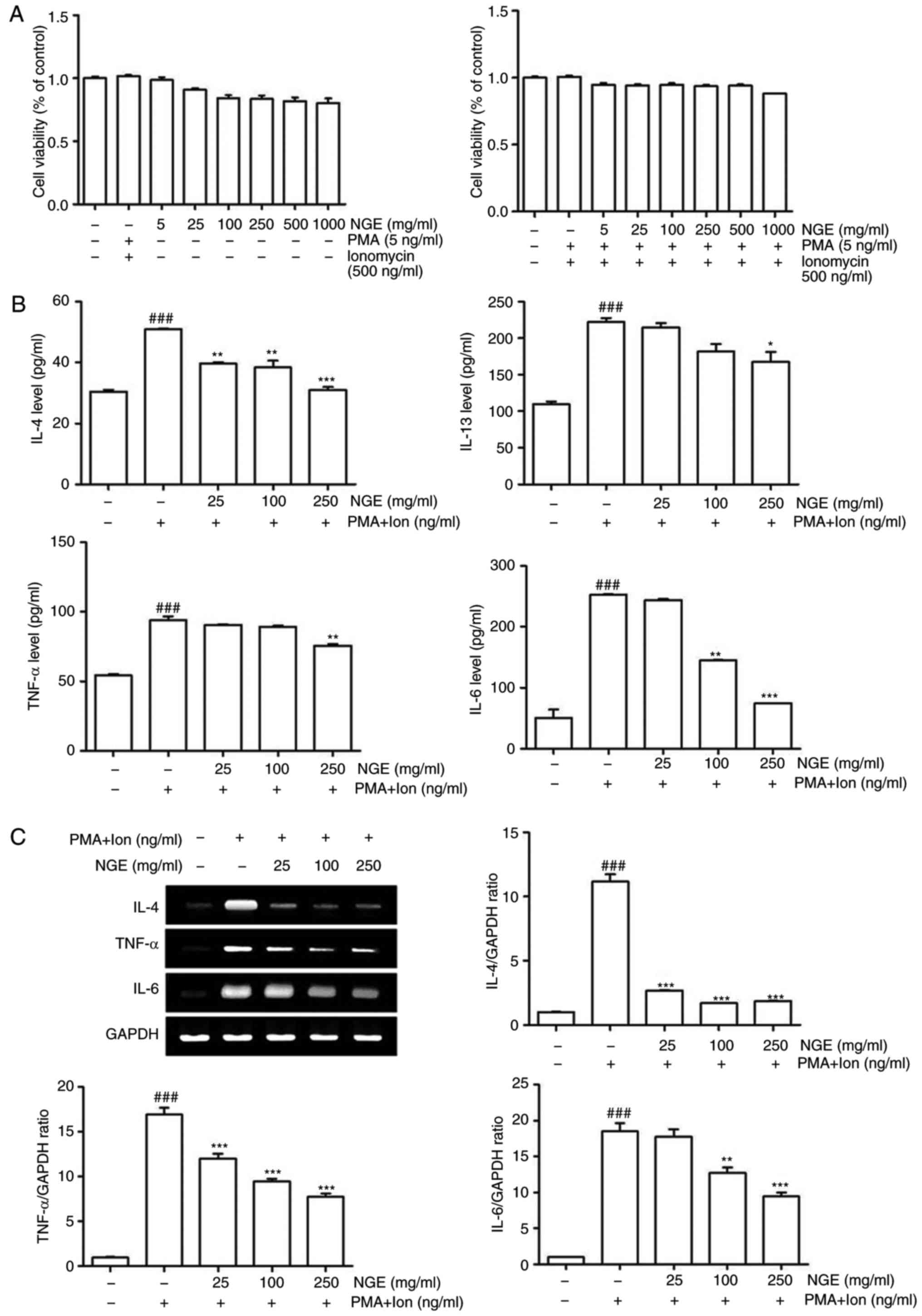 | Figure 1Effect of NGE on HMC-1 cells with PMA
+ ionomycin. (A) HMC-1 cells were treated with various
concentrations of NGE for 24 h following pretreatment with vehicle
(left panel) or with PMA + ionomycin (right panel) for 1 h.
Following treatment, cell viability was measured using a WST assay.
(B) Levels of IL-4, IL-13, TNF-α and IL-6 in cell culture
supernatants were measured via enzyme-linked immunosorbent assay.
HMC-1 cells were stimulated with 5 ng/ml PMA + 500 ng/ml ionomycin
for 1 h and treated with varying concentrations of NGE for 24 h.
(C) mRNA levels of IL-4, TNF-α and IL-6 were measured by RT-sqPCR
analysis in the treated HMC-1 cells. The bar graphs show
quantitation of the RT-sqPCR data; data are presented as the mean ±
standard error of the mean. ###P<0.001, compared with
the untreated group; **P<0.01 and
***P<0.001, compared with the PMA +
ionomycin-stimulated group. NGE, nok-gol; PMA, phorbol 12-myristate
13-acetate; Ion, ionomycin; RT-sqPCR, reverse
transcription-semi-quantitative polymerase chain reaction; IL,
interleukin; TNF-α, tumor necrosis factor-α. |
NGE suppresses the development of AD in
BALB/c mice
Following the application of DNCB to the skin of
mice to induce an acute allergic reaction, NGE was administered
orally for 2 weeks at a regular time every day. The level of
toxicity and the body weight was measured twice a week to determine
the degree of stress caused by the NGE treatment (Fig. 2A). Body weight and food intake
were also monitored throughout the study. As a result, it was found
that the mice in the vehicle group (n=6) were higher in weight
compared with those in the DNCB-treated (n=6) or NGE-treated (n=6)
groups. It was also found that the mice in the DNCB-treated group
did not exhibit any changes in food intake but exhibited a marginal
decrease (15%) in body weight compared with those in the
vehicle-treated group (Fig. 2C).
To investigate the effects of NGE on DNCB-induced AD-like symptoms
in BALB/c mice, skin lesions were examined and the skin thickness
and spleen weight were measured. It was found that DNCB induced AD
in BALB/c mice (Fig. 2B). In
addition, the skin thickness was gradually increased in the
DNCB-treated BALB/c mice compared with that in the non-DNCB-treated
mice, whereas NGE markedly attenuated the DNCB-induced increase in
skin thickness (Fig. 2D).
Similarly, the spleen weight of the DNCB-treated BALB/c mice was
heavier than that of the mice in the vehicle group, whereas NGE
significantly decreased the DNCB-induced increase in spleen weight
(Fig. 2E). These finding
suggested that NGE alleviated DNCB-induced AD.
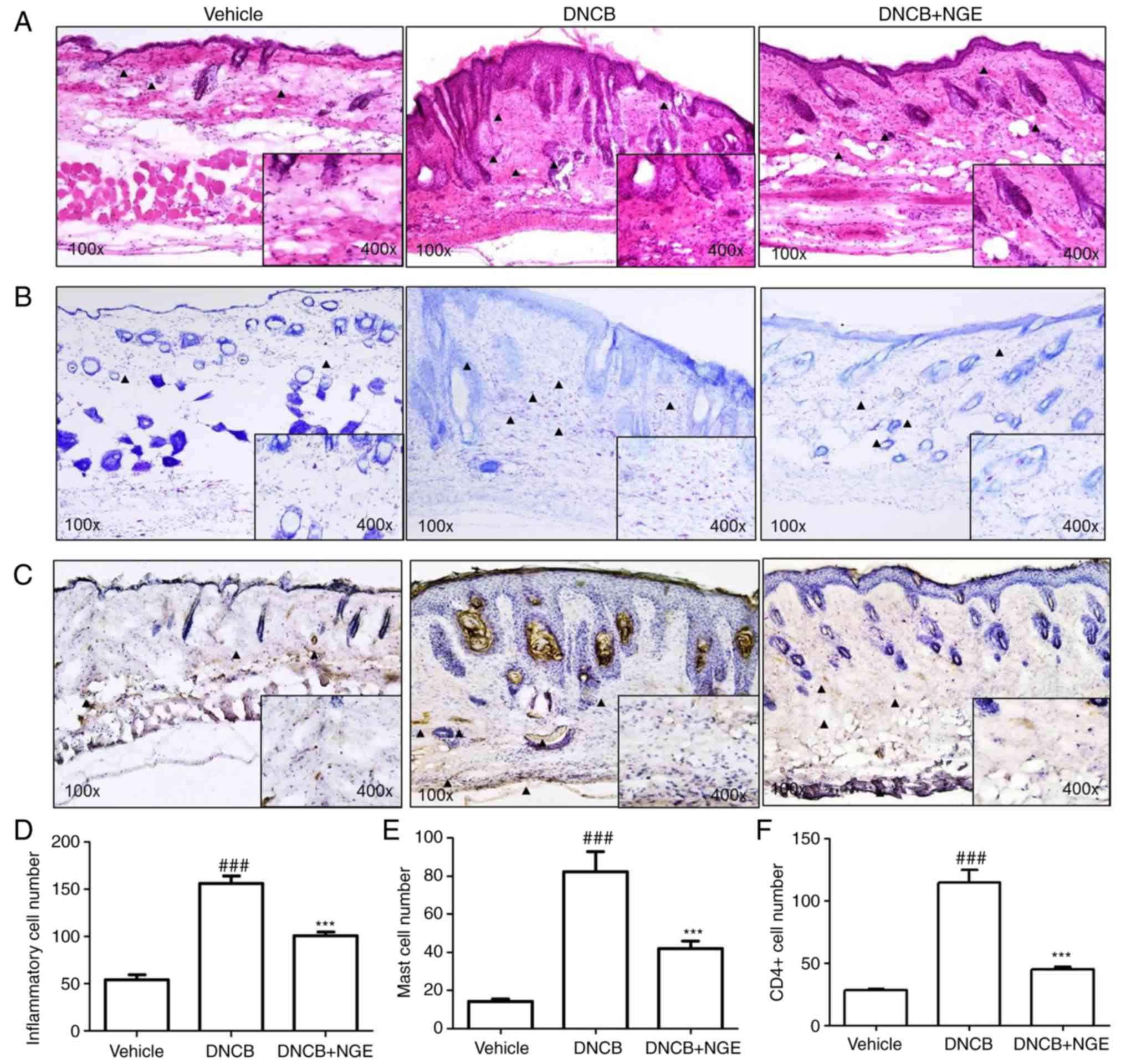
NGE decreases the infiltration of
inflammatory cells, mast cells and CD4+ T cells into AD
skin lesions
To determine whether NGE decreases the infiltration
of inflammatory cells (neutrophils and mononuclear cells) into AD
skin lesions, H&E staining of skin sections was performed
following topical administration of NGE. The infiltration of
inflammatory cells into the epidermis and dermis of mice in the
DNCB group was observed, whereas topical NGE decreased the
infiltration of inflammatory cells into the skin (Fig. 3A). Subsequently, toluidine blue
staining was performed for mast cell visualization. The repeated
cutaneous application of DNCB increased the dermal mast cell
number. However, this effect was significantly suppressed by NGE
(Fig. 3B). Immunocytochemistry
was performed to measure the intracellular level of CD4+
T cells. It was found that DNCB increased the number of
CD4+ T cells, whereas NGE decreased the number of
CD4+ T cells in the epidermis and dermis (Fig. 3C). These results indicated that
NGE treatment significantly decreased the number of inflammatory
cells, mast cells and CD4+ T cells, compared with DNCB
treatment (Fig. 3D-F).
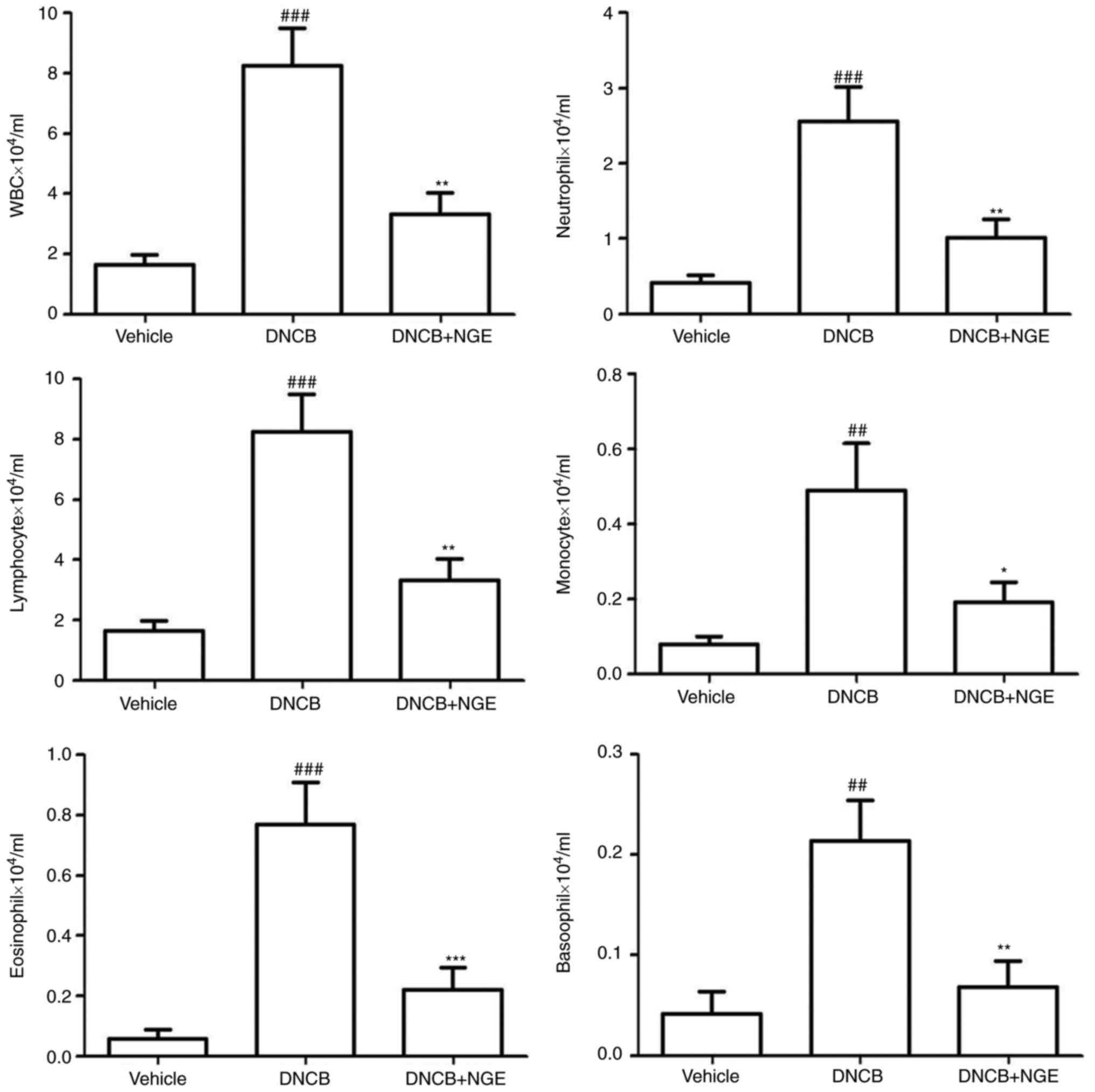
NGE decreases the levels of leukocytes
induced by DNCB
To investigate whether cutaneous NGE administration
can decrease the levels of inflammatory cells, leukocyte levels in
cardiovascular blood samples were measured using a HEMAVET 950
hematology system. It is known that WBCs, neutrophils, lymphocytes,
monocytes, eosinophils and basophils are activated in the blood of
patients with AD (17,30). In particular, eosinophils have
been shown to be present in the majority of patients with AD, and
they are correlated with disease activity (31). NGE significantly decreased the
numbers of WBCs, basophils, monocytes, neutrophils, eosinophils and
lymphocytes induced by DNCB (Fig.
4).
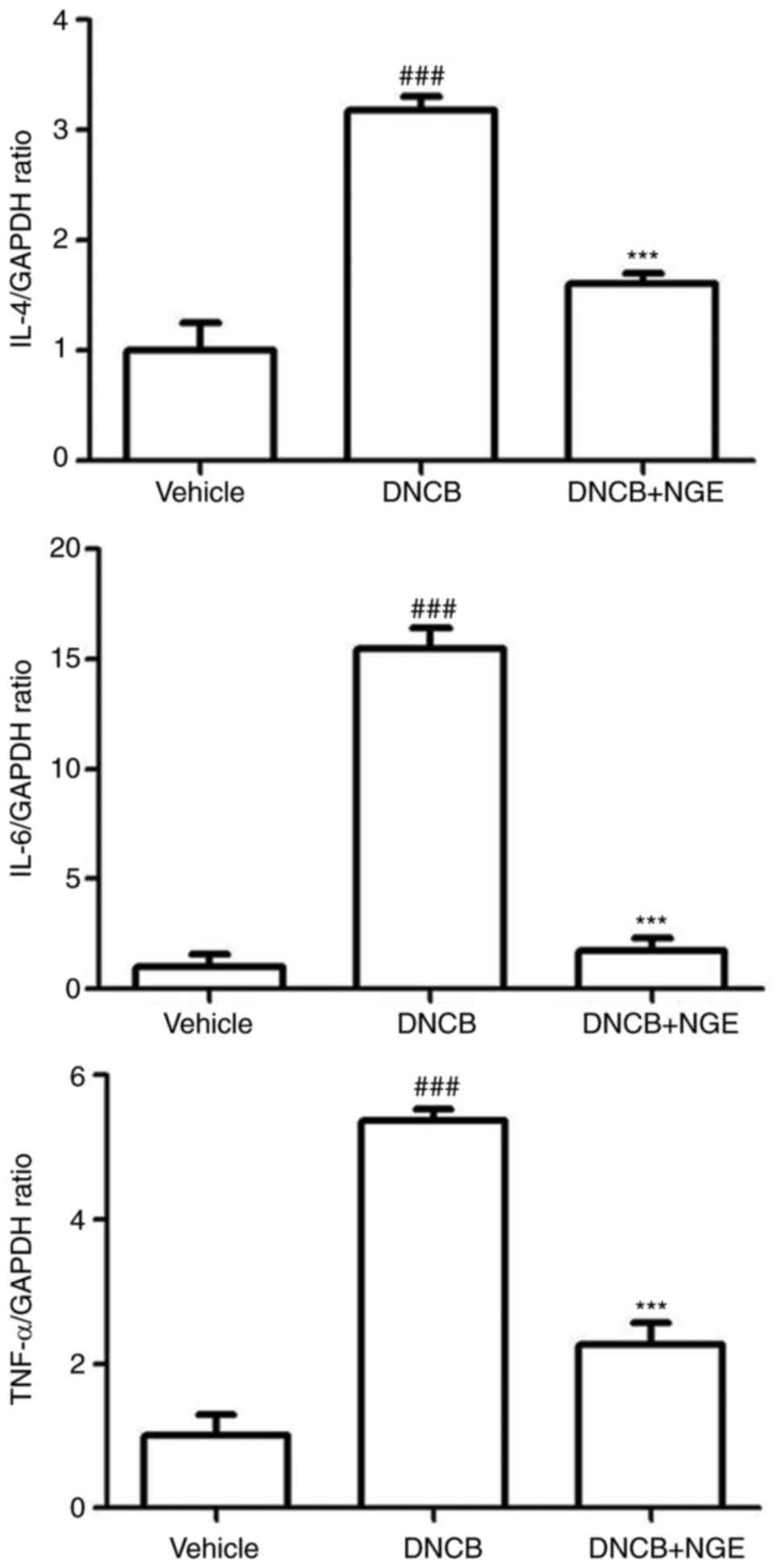
NGE administration downregulates the mRNA
expression of inflammatory cytokines induced by DNCB
In the histological analysis, the repeated cutaneous
application of DNCB increased dermal mast cell numbers and
CD4+ T cell numbers, and this effect was suppressed by
oral administration of NGE. Activated mast cells and
CD4+ T cells secrete various chemokines and cytokines,
including IL-4, IL-6 and TNF-α. To examine whether NGE decreases
the inflammatory response, RT-qPCR analysis was performed to
measure the mRNA expression of IL-4, IL-6 and TNF-α in the AD-like
skin lesions (Fig. 5). The oral
administration of NGE significantly suppressed the mRNA expression
of IL-4, IL-6 and TNF-α in the AD-like skin lesions. These results
indicated that NGE downregulated the DNCB-induced mRNA expression
of inflammatory cytokines.
NGE decreases the serum IgE concentration
and inflammatory cytokine levels (IL-10 and IL-12) in DNCB-treated
BALB/c mice
To obtain a better understanding of the mechanisms
underlying the anti-inflammatory activity of NGE, the IgE and
levels of inflammatory cytokines in serum were measured by ELISA.
The hyperproduction of IgE is a major characteristic of AD, and
patients with AD often exhibit elevated levels of total and
allergen-specific IgE antibodies in their serum. As shown in
Fig. 6, the total level of IgE
was markedly elevated in the DNCB-treated group compared with that
in the vehicle group. However, the increase in the serum level of
IgE induced by DNCB was significantly decreased by NGE treatment.
In addition, the repeated topical application of DNCB significantly
increased the levels of IL-10 and IL-12 in mouse serum, whereas NGE
significantly inhibited this increase. These results demonstrated
that NGE suppressed the DNCB-induced elevation of IgE and
inflammatory cytokine levels, thereby leading to the inhibition of
skin inflammation.
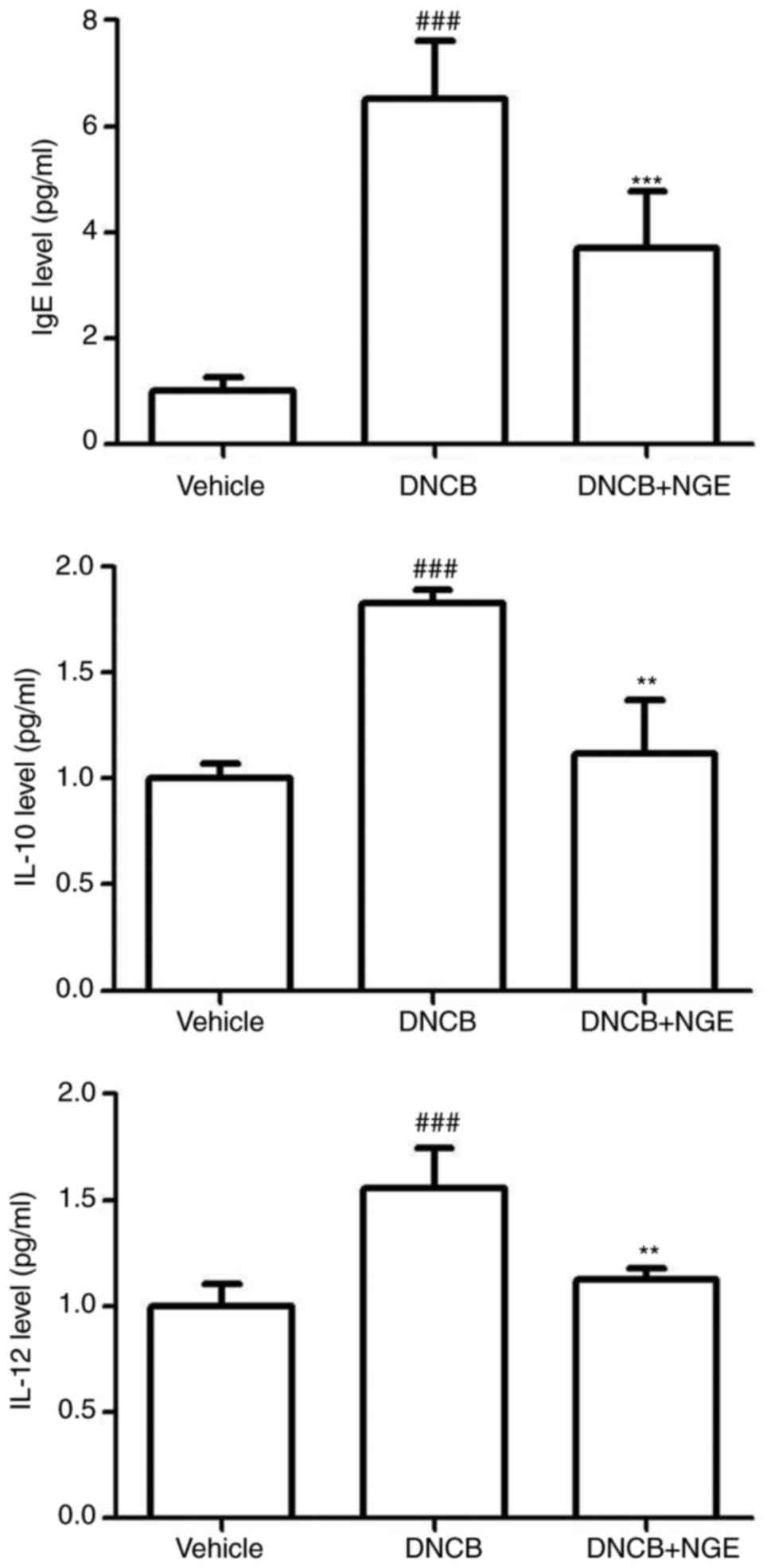 | Figure 6Effect of NGE on DNCB-induced plasma
IgE and cytokine levels. Blood samples were collected, and the
plasma was isolated. The plasma levels of IgE, IL-10, and IL-12 in
the indicated groups were measured using an enzyme-linked
immunosorbent assay. The data are presented as the mean ± standard
error of the mean. ###P<0.001, compared with the
vehicle group; **P<0.01 and ***P<0.001,
compared with the DNCB-stimulated group. NGE, nok-gol; DNCB,
2,4-dinitrochlorobenzene; IgE, immunoglobulin E; IL,
interleukin. |
Discussion
In the present study, the effect of NGE on
DNCB-induced AD in BALB/c mice was investigated. AD was induced by
the repeated alternative application of DNCB. DNCB is the most
important allergen associated with AD, and BALB/c mice offer a
useful model, exhibiting AD-like skin lesions and an elevated level
of blood IgE in response to DNCB (15-17). AD is regarded as a Th2-associated
inflammatory disease of the skin characterized by eosinophil
recruitment, inflammatory mediator release, and IgE production. IgE
molecules bind to their surface receptors on mast cells, causing
the release of mediators that recruit inflammatory cells and
stimulate the secretion of cytokines, including IL-4 and IL-13,
which are considered to drive disease pathology in patients with AD
(32). The oral administration of
NGE suppressed the development of AD in BALB/c mice, alleviating
the AD-like symptoms. NGE markedly attenuated the DNCB-induced
increase in skin thickness and spleen weight. Histopathological
analysis revealed that NGE decreased the infiltration of
inflammatory cells, mast cells and CD4+ cells into AD
skin lesions.
Mast cells are critical for the induction of
allergic diseases, including AD (29,33). Mast cells are localized in several
tissues, and the activation of mast cells induces the release of
histamine, tryptase and proinflammatory cytokines in tissues,
resulting in allergic responses (34,35). Mast cells are activated by the
binding of allergen-bound IgE to the FcεRI receptor (36). In the present study, NGE inhibited
IgE-mediated mast cell activation, resulting in suppression of the
expression of proinflammatory cytokines in mice with DNCB-induced
AD and mast cell activation. Notably, although PMA treatment was an
effective inducer of chemokine transcripts in HMC-1 cells,
treatment with PMA + ionomycin has been shown to be ineffective for
inducing chemokine expression in T cells (37,38). The present study found that NGE
markedly downregulated the mRNA expression of IL-4, IL-6 and TNF-α
in HMC-1 cells exposed to PMA + ionomycin, indicating that NGE
alleviates several AD symptoms by controlling the transcriptional
expression of Th2 cytokines. In addition, analysis via ELISA in
HMC-1 cells revealed that NGE inhibited the production of IL-4,
IL-13, TNF-α and IL-16 induced by PMA + ionomycin. These results
suggested that NGE has an anti-allergy effect. Of note,
CD4+ T cell subsets are involved in the pathogenesis of
several diseases (39).
CD4+ T cells generate cytokines that can stimulate other
T cell effectors and enhance antibody production by B cells
(40). CD4+ T cells
are also key factors in allergic inflammatory diseases. Th2 cells
produce IL-4, which induces B cell activation and antibody class
switching to IgE; the increased production of IL-4 or induction of
the IL-4 signaling pathway causes allergic diseases (41).
NGE was shown to decrease the levels of leukocytes
induced by DNCB. Eosinophils are involved in the pathology of
allergic diseases, particularly AD (9,42).
The elevated eosinophil count is associated with inflammation and a
change in eosinophil cell number may be a result of disease. In
addition, NGE downregulated the mRNA expression of inflammatory
cytokines induced by DNCB. NGE also decreased the serum IgE
concentration and inflammatory cytokine levels in the DNCB-treated
BALB/c mice.
The most common side effect associated with ointment
treatment is a sensation of mild to moderate skin burning at the
site of ointment application. It has been reported that the
long-term use of corticosteroids results in treatment resistance
(43,44). The currently used medications
include antihistamines, steroids and immunosuppressants, however,
these medications show various limitations in treatment efficacy
(6). Therefore, the
identification of novel compounds which effectively treat AD is
urgently required.
With the aim of developing an appropriate treatment
agent for AD using a natural substance, the present study performed
an in vivo experiment using BALB/c mice to assess the
anti-AD effect of NGE. The results showed that NGE suppressed
DNCB-induced AD in the BALB/c mice. As a herbal medicine, NGE may
not have substantial side effects when used to treat AD.
In conclusion, the present study revealed for the
first time, to the best of our knowledge, that NGE, which is
derived from an anti-inflammatory traditional medicine, alleviates
AD-like inflammatory symptoms in mice by suppressing the production
of CD4+ T cells, mast cells, eosinophils and
proinflammatory cytokines. These results suggest that NGE may be a
useful drug for the treatment of AD.
Funding
This study was supported by grants from the Korean
Medicine R&D Project of the Ministry of Health and Welfare
(grant nos. HI12C1889 and HI13C0530) and Nongshim Corporation
(grant no. 20121086).
Availability of data and materials
All data and materials are described within the
article. The corresponding author will provide data and materials
upon request.
Authors' contributions
SHH, JMK, and S-GK conceived and designed the
experiments; SHH, HIK, SJL and YSL performed the experiments; SHH
and YCS analyzed the data; SHH and HSS contributed to the writing
of the paper. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All procedures performed on mice were approved by
the Animal Care Center of Kyung Hee University [approval no. KHUASP
(SE)-16-095]. All methods were performed in accordance with the
relevant guidelines and regulations.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Leung DY and Bieber T: Atopic dermatitis.
Lancet. 361:151–160. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Spergel JM and Paller AS: Atopic
dermatitis and the atopic march. J Allergy Clin Immunol.
112:S118–S127. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Simon D, Braathen LR and Simon HU:
Eosinophils and atopic dermatitis. Allergy. 59:561–570. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rousset F, Robert J, Andary M, Bonnin JP,
Souillet G, Chrétien I, Brière F, Pène J and de Vries JE: Shifts in
interleukin-4 and interferon-gamma production by T cells of
patients with elevated serum IgE levels and the modulatory effects
of these lymphokines on spontaneous IgE synthesis. J Allergy Clin
Immunol. 87:58–69. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bieber T: Atopic dermatitis. Ann Dermatol.
22:125–137. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gleich GJ: Mechanisms of
eosinophil-associated inflammation. J Allergy Clin Immunol.
105:651–663. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Holt PG, Macaubas C, Stumbles PA and Sly
PD: The role of allergy in the development of asthma. Nature.
402:B12–B17. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Leiferman KM: Eosinophils in atopic
dermatitis. Allergy. 9:20–26. 1989. View Article : Google Scholar
|
|
9
|
Ogawa K, Hashida R, Miyagawa M, Kagaya S,
Sugita Y, Matsumoto K, Katsunuma T, Akasawa A, Tsujimoto G and
Saito H: Analysis of gene expression in peripheral blood
eosinophils from patients with atopic dermatitis and in vitro
cytokine-stimulated blood eosinophils. Clin Exp Immunol.
131:436–445. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hanifin JM, Cooper KD, Ho VC, Kang S,
Krafchik BR, Margolis DJ, Schachner LA, Sidbury R, Whitmore SE,
Sieck CK, et al: Guidelines of care for atopic dermatitis,
developed in accordance with the American Academy of Dermatology
(AAD)/American Academy of Dermatology Association 'Administrative
Regulations for Evidence-Based Clinical Practice Guidelines'. J Am
Acad Dermatol. 50:391–404. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Novak N, Valenta R, Bohle B, Laffer S,
Haberstok J, Kraft S and Bieber T: FcepsilonRI engagement of
Langerhans cell-like dendritic cells and inflammatory dendritic
epidermal cell-like dendritic cells induces chemotactic signals and
different T-cell phenotypes in vitro. J Allergy Clin Immunol.
113:949–957. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ong PY and Leung DY: Immune dysregulation
in atopic dermatitis. Curr Allergy Asthma Rep. 6:384–389. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Homey B, Steinhoff M, Ruzicka T and Leung
DY: Cytokines and chemokines orchestrate atopic skin inflammation.
J Allergy Clin Immunol. 118:178–189. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Leung DY, Boguniewicz M, Howell MD, Nomura
I and Hamid QA: New insights into atopic dermatitis. J Clin Invest.
113:651–657. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dearman RJ, Basketter DA and Kimber I:
Differential cytokine production following chronic exposure of mice
to chemical respiratory and contact allergens. Immunology.
86:545–550. 1995.PubMed/NCBI
|
|
16
|
Dearman RJ, Basketter DA and Kimber I:
Characterization of chemical allergens as a function of divergent
cytokine secretion profiles induced in mice. Toxicol Appl
Pharmacol. 138:308–316. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Warbrick EV, Dearman RJ, Basketter DA and
Kimber I: Analysis of interleukin 12 protein production and mRNA
expression in mice exposed topically to chemical allergens.
Toxicology. 132:57–66. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Choi HS, Kim SR, Hong SH, Ku JM, Kim MK,
Seo HS, Cho SG, Shin S, Shin YC and Ko SG: Water extract of deer
bones activates macrophages and alleviates neutropenia. Evid Based
Complement Alternat Med. 2013:6173022013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Du CN, Min AY, Kim HJ, Shin SK, Yu HN,
Sohn EJ, Ahn CW, Jung SU, Park SH and Kim MR: Deer bone extract
prevents against scopolamine-induced memory impairment in mice. J
Med Food. 18:157–165. 2015. View Article : Google Scholar :
|
|
20
|
Li YJ, Kim TH, Kwak HB, Lee ZH, Lee SY and
Jhon GJ: Chloroform extract of deer antler inhibits osteoclast
differentiation and bone resorption. J Ethnopharmacol. 113:191–198.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kuo CY, Wang T, Dai TY, Wang CH, Chen KN,
Chen YP and Chen MJ: Effect of the velvet antler of formosan sambar
deer (Cervus unicolor swinhoei) on the prevention of an allergic
airway response in mice. Evid Based Complement Alternat Med.
2012:4813182012. View Article : Google Scholar
|
|
22
|
Dai TY, Wang CH, Chen KN, Huang IN, Hong
WS, Wang SY, Chen YP, Kuo CY and Chen MJ: The antiinfective effects
of velvet antler of formosan sambar deer (Cervus unicolor swinhoei)
on staphylococcus aureus-infected mice. Evid Based Complement
Alternat Med. 2011:5340692011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hong SH, Ku JM, In Kim H, Ahn CW, Park SH,
Seo HS, Shin YC and Ko SG: The immune-enhancing activity of Cervus
nippon mantchuricus extract (NGE) in RAW264.7 macrophage cells and
immunosuppressed mice. Food Res Int. 99:623–629. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kang SK, Kim KS, Kim SI, Chung KH, Lee IS
and Kim CH: Immunosuppressive activity of deer antler extracts of
Cervus korean TEMMINCK varmantchuricus Swinhoe, on type II
collagen-induced arthritis In Vitro. Cell Dev Biol Anim.
42:100–107. 2006. View Article : Google Scholar
|
|
25
|
Kim KS, Choi YH, Kim KH, Lee YC, Kim CH,
Moon SH, Kang SG and Park YG: Protective and anti-arthritic effects
of deer antler aqua-acupuncture (DAA), inhibiting dihydroorotate
dehydrogenase, on phosphate ions-mediated chondrocyte apoptosis and
rat collagen-induced arthritis. Int Immunopharmacol. 4:963–973.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee H, Choi HS, Park Y, Ahn CW, Jung SU,
Park SH and Suh HJ: Effects of deer bone extract on the expression
of pro-inflammatory cytokine and cartilage-related genes in
monosodium iodoacetate-induced osteoarthritic rats. Biosci
Biotechnol Biochem. 78:1703–1709. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Galli SJ, Tsai M and Piliponsky AM: The
development of allergic inflammation. Nature. 454:445–454. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ku JM, Hong SH, Kim HI, Seo HS, Shin YC
and Ko SG: Effects of Angelicae dahuricae radix on
2,4-dinitrochloro-benzene-induced atopic dermatitis-like skin
lesions in mice model. BMC Complement Altern Med. 17:982017.
View Article : Google Scholar
|
|
29
|
Liu FT, Goodarzi H and Chen HY: IgE, mast
cells, and eosinophils in atopic dermatitis. Clin Rev Allergy
Immunol. 41:298–310. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lubach D, Bensmann A and Bornemann U:
Steroid-induced dermal atrophy. Investigations on discontinuous
application. Dermatologica. 179:67–72. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Reitamo S, Rissanen J, Remitz A, Granlund
H, Erkko P, Elg P, Autio P and Lauerma AI: Tacrolimus ointment does
not affect collagen synthesis: Results of a single-center
randomized trial. J Invest Dermatol. 111:396–398. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Werfel T, Allam JP, Biedermann T, Eyerich
K, Gilles S, Guttman-Yassky E, Hoetzenecker W, Knol E, Simon HU,
Wollenberg A, et al: Cellular and molecular immunologic mechanisms
in patients with atopic dermatitis. J Allergy Clin Immunol.
138:336–349. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kawakami T, Ando T, Kimura M, Wilson BS
and Kawakami Y: Mast cells in atopic dermatitis. Curr Opin Immunol.
21:666–678. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Metz M, Grimbaldeston MA, Nakae S,
Piliponsky AM, Tsai M and Galli SJ: Mast cells in the promotion and
limitation of chronic inflammation. Immunol Rev. 217:304–328. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Theoharides TC, Kempuraj D, Tagen M, Conti
P and Kalogeromitros D: Differential release of mast cell mediators
and the pathogenesis of inflammation. Immunol Rev. 217:65–78. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kalesnikoff J and Galli SJ: New
developments in mast cell biology. Nat Immunol. 9:1215–1223. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Miller MD, Hata S, De Waal Malefyt R and
Krangel MS: A novel polypeptide secreted by activated human T
lymphocytes. J Immunol. 143:2907–2916. 1989.PubMed/NCBI
|
|
38
|
Zipfel PF, Balke J, Irving SG, Kelly K and
Siebenlist U: Mitogenic activation of human T cells induces two
closely related genes which share structural similarities with a
new family of secreted factors. J Immunol. 142:1582–1590.
1989.PubMed/NCBI
|
|
39
|
Hamid Q, Boguniewicz M and Leung DY:
Differential in situ cytokine gene expression in acute versus
chronic atopic dermatitis. J Clin Invest. 94:870–876. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ahmadzadeh M, Hussain SF and Farber DL:
Heterogeneity of the memory CD4 T cell response: Persisting
effectors and resting memory T cells. J Immunol. 166:926–935. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hershey GK, Friedrich MF, Esswein LA,
Thomas ML and Chatila TA: The association of atopy with a
gain-of-function mutation in the alpha subunit of the interleukin-4
receptor. N Engl J Med. 337:1720–1725. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Matsumoto K and Saito H: The role of
eosinophils in asthma: Sarastro or the Queen of the Night? Int Arch
Allergy Immunol. 125:290–296. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gottlieb AB: Therapeutic options in the
treatment of psoriasis and atopic dermatitis. J Am Acad Dermatol.
53:S3–S16. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Del Rosso J and Friedlander SF:
Corticosteroids: Options in the era of steroid-sparing therapy. J
Am Acad Dermatol. 53:S50–S58. 2005. View Article : Google Scholar : PubMed/NCBI
|