Introduction
It has been reported that the incidence of breast
cancer is increasing, and it is predicted to increase further in
the next 20 years worldwide (1).
Substantial evidence has suggested that risk factors, including
genetic mutations, family inheritance, chemicals, radiation and
lifestyle factors, such as being overweight, excessive alcohol
consumption and insufficient physical exercise, contribute to the
high incidence of breast cancer (2). Breast cancer is a solid tumor, and
the surrounding tumor microenvironment is particularly important
for its development (3).
Furthermore, expression of the estrogen receptor, progesterone
receptor or the human epidermal growth factor receptor 2 oncogene
is a significant indicator for breast cancer classification
(4). Recently, breast cancer
treatment has markedly improved due to advances in chemotherapy,
including anthracyclines, alkylating agents and cytoskeletal
disruptors; radiation therapy; hormonal blockade, including
tamoxifen and aromatase inhibitors; and targeted biological
therapies, including trastuzumab and lapatinib. However, the side
effects of various therapies, including the risk of cardiovascular
diseases and breast cancer recurrence remain complex issues that
urgently need to be addressed (5).
MicroRNAs (miRNAs/miRs) are a class of non-coding
RNA molecules, 18-24 nucleotides long, which regulate gene
expression at the post-transcriptional level by specifically
binding to their targets (6).
Various miRNAs, including miR-206, miR-221/222, miR-22, let-7
family, miR-34 and miR-194 serve regulatory roles in breast cancer
progression (7). Furthermore,
miR-194 is a vertebrate-specific miRNA, which has pivotal roles in
energy production, inflammatory inhibition and malignancies, such
as breast cancer, non-small cell lung cancer (8), renal cell carcinoma (9), acute myeloid leukemia (10) and colorectal cancer (11). Furthermore, upregulation of
miR-194 is associated with the recurrence of breast cancer
(12). However, the exact roles
of miR-194 in breast cancer remain to be elucidated.
The Wnt/β-catenin pathway is a complex signaling
cascade, which serves significant biological roles in embryonic
development, cellular polarity, cell proliferation, differentiation
and apoptosis (13). Wnt ligands
are able to interact with the Frizzled family of seven-pass
transmembrane receptors, and/or co-receptors, such as lipoprotein
receptor-related protein 5/6, receptor tyrosine kinase like orphan
receptor 2 and receptor-like tyrosine kinase. Subsequently,
activation of the Wnt pathway leads to the accumulation of
β-catenin in the nucleus. The binding reaction of β-catenin to T
cell factor/lymphoid enhancer factor (LEF) gives rise to gene
expression (14). The
Wnt/β-catenin signaling pathway is associated with regulation of a
wide range of biological effects, due to its ability to up- or
downregulate downstream gene expression, thus resulting in the
alteration of other signaling pathways (15). SRY-box 17 (SOX17) belongs to the
high-mobility group-box transcription factor family. It has been
reported that SOX17 gene methylation in breast cancer may promote
the degradation of β-catenin in the Wnt singling pathway (16). However, the roles of SOX17 in
breast cancer remain to be determined.
The present study aimed to investigate the
expression of miR-194-5p and its roles in breast cancer. To verify
that SOX17 is a direct target gene of miR-194-5p, bioinformatics
analysis using TargetScan (http://www.targetscan.org/vert_71/) indicated that
SOX17 is a direct target of miR-194-5p. Therefore, the present
study investigated the correlation between miR-194-5p and SOX17,
and the biological effects of miR-194-5p knockdown on the
Wnt/β-catenin signaling pathway in breast cancer.
Materials and methods
Tissues and cells
The human studies were approved by the ethics
committee of Yinzhou People's Hospital of Ningbo City (Ningbo,
China). Written informed consent was obtained from all patients. A
total of 30 pairs of breast cancer and paracarcinoma tissues (≥5 cm
away from the tumor edge) were obtained from patients with breast
cancer that had not undergone radiotherapy and chemotherapy (age
range, 25-55 years old). The patients were admitted to the Yinzhou
People's Hospital of Ningbo City between January 2015 and March
2017. After tissue collection, the specimens were fixed at 4°C
overnight in 10% formalin for storage. The normal breast epithelial
cell line MCF-10A, and the breast cancer cell lines, MCF-7, T47D
and MDA-MB-231, were obtained from the American Type Culture
Collection (Manassas, VA, USA). All cells were cultured in Roswell
Park Memorial Institute-1640 medium supplemented with 10% fetal
bovine serum (FBS) (both from Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR was performed to investigate the correlation
between miR-194-5p and SOX17 in breast cancer. Once cells reached
60% confluence, TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) was applied for RNA extraction. The same
method was used for RNA extraction from tissues. RNA concentration
was determined using a NanoDrop 2000 instrument (NanoDrop
Technologies; Thermo Fisher Scientific, Inc., Wilmington, DE, USA).
M-MLV RT (cat no. 28025-013; Invitrogen; Thermo Fisher Scientific,
Inc.) and GoTaq® DNA Polymerase (cat no. M3005; Promega
Corporation, Madison, WI, USA) were used for cDNA synthesis and
PCR, respectively. cDNA synthesis was conducted at 95°C for 12 min,
and the PCR thermocycling conditions were as follows: 97°C for 5
min, followed by 35 cycles at 95°C for 30 sec, 65°C for 30 sec and
73°C for 1 min, and a final step at 73°C for 10 min, after which
samples were maintained at 4°C until use. The miR-194-5p and SOX17
PCR primer sequences were as follows: miR-194-5p, forward, 5'-ACA
CTC CAG CTG GGT GTA ACA GCA ACT CC-3' and reverse, 5'-TGG TGT CGT
GGA GTC G-3'; and SOX17, forward, 5'-GTG GAC CGC ACG GAA TTT G-3'
and reverse, 5'-GGA GAT TCA CAC CGG AGT CA-3'. The expression
levels were normalized to U6 or GAPDH, the primer sequences were as
follows: U6, forward, 5'-AGTA AGC CCT TGC TGT CAG TG-3' and
reverse, 5'-CCT GGG TCT GAT AAT GCT GGG-3'; and GAPDH, forward,
5'-ACA ACT TTG GTA TCG TGG AAG G-3' and reverse, 5'-GCC ATC ACG CCA
CAG TTT C-3'. The 2−ΔΔCq method was used for relative
quantification (17).
Cell transfection
MCF-7 cells (1×105) were seeded into
24-well plates, and the miR-194-5p inhibitor (miR20000460-1-5) and
miR-194-5p overexpression plasmids (miR10000460-1-5) were obtained
from Guangzhou RiboBio Biotechnology Co., Ltd. (Guangzhou, China).
The overexpression plasmid was used for luciferase reporter assays.
Briefly, 500 ng plasmids were transfected into cells using 1
µl Lipo6000™ Transfection Reagent (Beyotime Institute of
Biotechnology, Shanghai, China); the cells were incubated at 37°C
in an atmosphere containing 5% CO2 for 6 h.
Subsequently, the transfected cells were incubated for a further 72
h. The empty vector was transfected into cells in the
overexpression negative control (NC) groups. The following
sequences were transfected for miR-194-5p overexpression: Forward,
5'-CAG GAG TTG TAA ATC CGA GCC G-3' and reverse, 5'-TTC ATA GGT CAG
AGC CCT GTG CA-3'.
MTT assay
After 72 h incubation, the MTT assay was performed
for cell proliferation analysis. Briefly, MCF-7 cells
(5×105 cells/well) were placed into 96-well plates and
the cell proliferation rate was determined at various time points
(24, 48 and 72 h). Briefly, 20 µl MTT reagent
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was added to cells
and incubated for 4 h at 37°C. Subsequently, formazan crystals were
dissolved in dimethyl sulfoxide and the optical density (OD) value
was detected at 490 nm using a microplate reader.
Colony formation assay
The colony formation assay was performed after
transfection and 72 h incubation. Cells (1,500 cells/well) were
plated into 12-well plates and cultured for 10 days to form
colonies. The cells were then fixed with methanol for 15 min at 4°C
and stained with Giemsa dye solution for 30 min at room temperature
to visualize cell colonies. Images were captured under a light
microscope.
Scratch and Transwell assays
Post-transfection, the migration and invasion of
MCF-7 cells was evaluated by scratch and Transwell assays. For the
scratch assay, MCF-7 cells in log phase were seeded into 96-well
plates to obtain a monolayer cell culture. Across the center of the
well, the cell monolayer was scratched using a fresh 1-ml pipette
tip; the width of the scratch was equal to the outer diameter of
the tip. Subsequently, the cells were incubated at 37°C in an
atmosphere containing 5% CO2 for 72 h. Images of the
migrated cells were captured after 24 h under an inverted
microscope (CKX41; Olympus Corporation, Tokyo, Japan) and were
analyzed using ImageJ v1.8.0 (National Institutes of Health,
Bethesda, MD, USA). For the Transwell assay, Transwell culture
inserts (pore size, 8 mm; Falcon; BD Biosciences, Franklin Lakes,
NJ, USA) were placed into the wells of 96-well plates, thus leading
to separated upper and lower chambers. The upper side of the
membrane was precoated with Matrigel (BD Biosciences) and incubated
at 37°C for 1 h for gel formation. FBS was used to hydrate the
membrane 2 h prior to use. Subsequently, Dulbecco's modified
Eagle's medium (Gibco; Thermo Fisher Scientific, Inc.; 600
µl) containing 10% FBS was added to the lower chamber,
whereas 1×105 cells/well were added to the upper
chamber. After 72 h at 37°C, the number of invading cells was
counted using a counting chamber under an inverted microscope
(CKX41; Olympus Corporation).
Western blotting
Western blot analysis was conducted after 72 h
incubation. Proteins were isolated from transfected MCF-7 cells
using radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology) and protein concentration was measured
using a bicinchoninic acid kit (Beyotime Institute of
Biotechnology). Cytosolic protein was extracted using the Cytosol
Protein Extraction kit (#P0033; Beyotime Institute of
Biotechnology). Nuclear protein was extracted using the
Nucleoprotein Extraction kit (#C500009-0050; Sangon Biotech Co.,
Ltd., Shanghai, China). Subsequently, the protein samples (20
µg/per lane) were separated by 15% SDS-PAGE. Polyvinylidene
fluoride (PVDF) membranes (EMD Millipore, Billerica, MA, USA) were
used for protein transfer. Subsequently, the PVDF membranes were
blocked with 5% skimmed milk for 2 h at room temperature, and were
incubated with the following primary antibodies: Anti-SOX17
(#81778, 1:2,000), anti-Wnt (#2915, 1:2,000), anti-β-catenin
(#8480, 1:500), anti-phosphorylated (p)-β-catenin (#9567, 1:500)
and anti-GAPDH (#5174, 1:2,000) (Cell Signaling Technology, Inc.,
Danvers, MA, USA) for 1 h at room temperature, followed by
incubation with secondary antibodies (sc-2004, 1:1,000; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) for 45 min at room
temperature. An enhanced chemiluminescence kit (#32209; Pierce;
Thermo Fisher Scientific, Inc.) was applied for visualization and
the images were analyzed using ImageJ v1.8.0 (National Institutes
of Health).
Bioinformatics analysis and luciferase
reporter assays
Bioinformatics analysis with TargetScan, and
luciferase reporter assays were conducted to verify that SOX17 was
a direct target gene of miR-194-5p. For luciferase reporter assays,
MCF-7 cells were seeded into 96-well plates at a density of
2×105 cells/well and grown to 70% confluence. Following
the implementation of site-directed mutagenesis using the
QuikChange Lightning Site-Directed Mutagenesis kit (Guangzhou
RiboBio Biotechnology Co., Ltd.), cells were co-transfected with
miR-194-5p mimic or miR-194-5p NC for 48 h at 37°C (50 ng;
Guangzhou RiboBio Biotechnology Co., Ltd.), and SOX17-3'
untranslated region (UTR)-wild-type (WT) (50 ng) or
SOX17-3'UTR-mutant (MUT) (50 ng) plasmids (Guangzhou RiboBio
Biotechnology Co., Ltd.) using Lipo6000™ Transfection Reagent (0.2
µl). Furthermore, transfection efficiency was normalized to
a Renilla luciferase vector (pRL-CMV; Promega Corporation).
The luciferase assay kit (BioLux® Gaussia; New
England Biolabs, Inc., Ipswich, MA, USA) was used to evaluate
lucif-erase activity according to the manufacturer's protocol.
Animals
The animal studies were approved by the laboratory
animal management and welfare ethical review committee of Yinzhou
People's Hospital of Ningbo City. A total of 18 female BALB/c-nu/nu
nude mice (age, 4-5 weeks; weight, 18-25 g) were obtained from
Zhejiang Experimental Animal Center. The mice were housed in a
specific pathogen-free laboratory and were maintained under the
following conditions: Constant temperature, 22-26°C; humidity,
40-70%; 12-h light/dark cycle; free access to food and water).
Briefly, the mice were randomly divided into groups A and B; mice
in group A were used for tumor weight and volume analysis, whereas
mice in group B were used for immunohistochemical analysis.
Briefly, 500 ng miR-194-5p inhibitor and negative control (NC) were
transfected into MCF-7 cells with 1 µl Lipo6000™
Transfection Reagent at 37°C with 5% CO2 for 6 h.
Subsequently, 1×107/0.3 ml MCF-7 cells in the log phase
were inoculated into the left side of the breast to generate a
murine model of breast cancer. After 20 days ad libitum
feeding, the tumor-burdened nude mice exhibited tumors ~0.5 cm in
diameter. Mice were divided into the control, NC and miR-194-5p
inhibitor groups.
Tumor weight and size
Tumor weight was measured using an electric scale
and tumor volume was recorded using a vernier caliper 20 days after
MCF-7 cell inoculation.
Immunohistochemistry (IHC)
After 20 days ad libitum feeding, the mice
were anesthetized and sacrificed. Subsequently, the tumor tissues
were obtained and fixed in 10% formalin at 4°C overnight. Sections
were incubated with anti-SOX17 primary antibodies (#81778S,
1:1,000; Cell Signaling Technology, Inc.) for 1 h at room
temperature, after which they were incubated with secondary
antibodies (#7074, 1:2,000; Cell Signaling Technology, Inc.) for 10
min at room temperature. Images of staining were captured under an
inverted microscope (CKX41; Olympus Corporation) and were analyzed
using Image-Pro Plus 6.0 (Media Cybernetics, Inc., Rockville, MD,
USA).
Statistical analysis
Each experiment was repeated in triplicate and all
data are presented as the means ± standard deviation. Additionally,
Student's t-test was used for two group comparisons, whereas
one-way analysis of variance followed by the Dunnett's post hoc
test was used for multiple group comparisons, and the Pearson's
correlation coefficient was employed for correlation analysis. Data
were analyzed using SPSS 14.0 (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Expression levels of miR-194-5p and SOX17
in breast cancer tissues, and correlation analysis
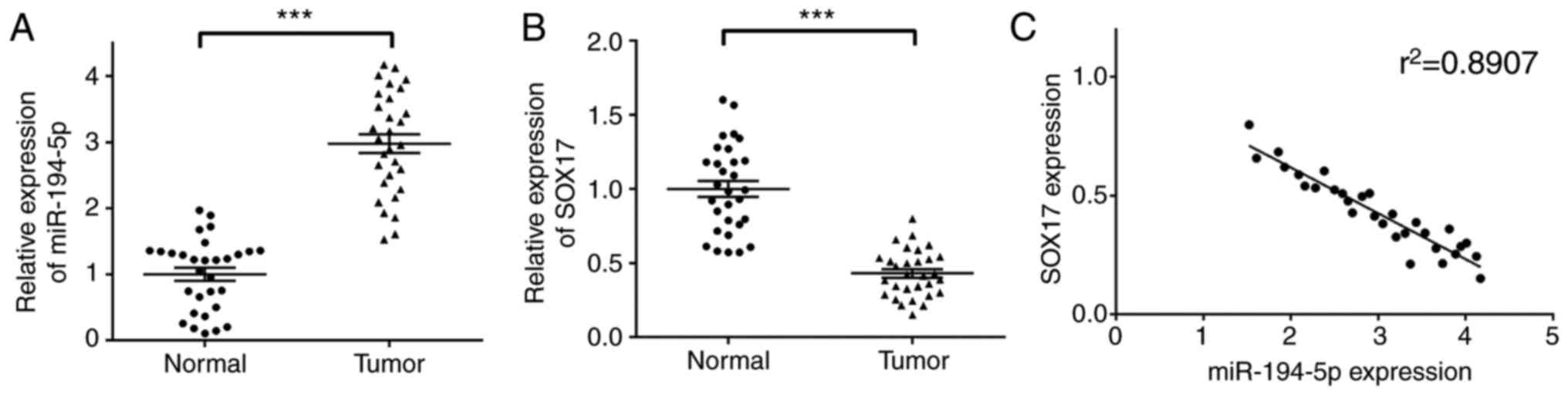
In order to investigate the correlation between
miR-194-5p and SOX17 in breast cancer, the expression levels of
miR-194-5p and SOX17 were determined by RT-qPCR. As shown in
Fig. 1A and B, miR-194-5p was
significantly upregulated, whereas SOX17 was markedly downregulated
in breast cancer tissues compared with in normal tissues. In
addition, a negative correlation was detected between miR-194-5p
and SOX17 (r2=0.8907; Fig.
1C), thus indicating that the expression of miR-194-5p was
negatively correlated with SOX17 in breast cancer. It is of great
significance to further elucidate the correlation between
miR-194-5p and SOX17, and the regulatory relationship of the Wnt
signaling pathway.
Upregulation of miR-194-5p in various
breast cancer cell lines
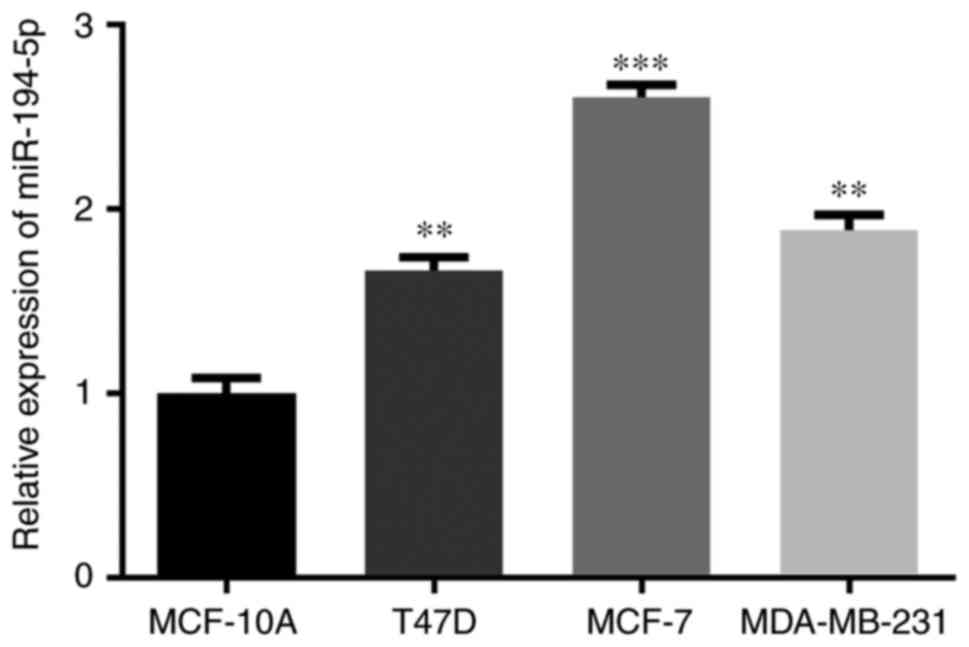
The expression levels of miR-194-5p were also
detected in numerous breast cancer cell lines, in order to select
an appropriate cell line for subsequent experiments. As shown in
Fig. 2, miR-194-5p was
significantly increased in MCF-7, T47D and MDA-MB-231 cells
compared with in MCF-10A cells. Furthermore, miR-194-5p was most
upregulated in MCF-7 cells; therefore, MCF-7 was chosen as the
target cell line in the present study.
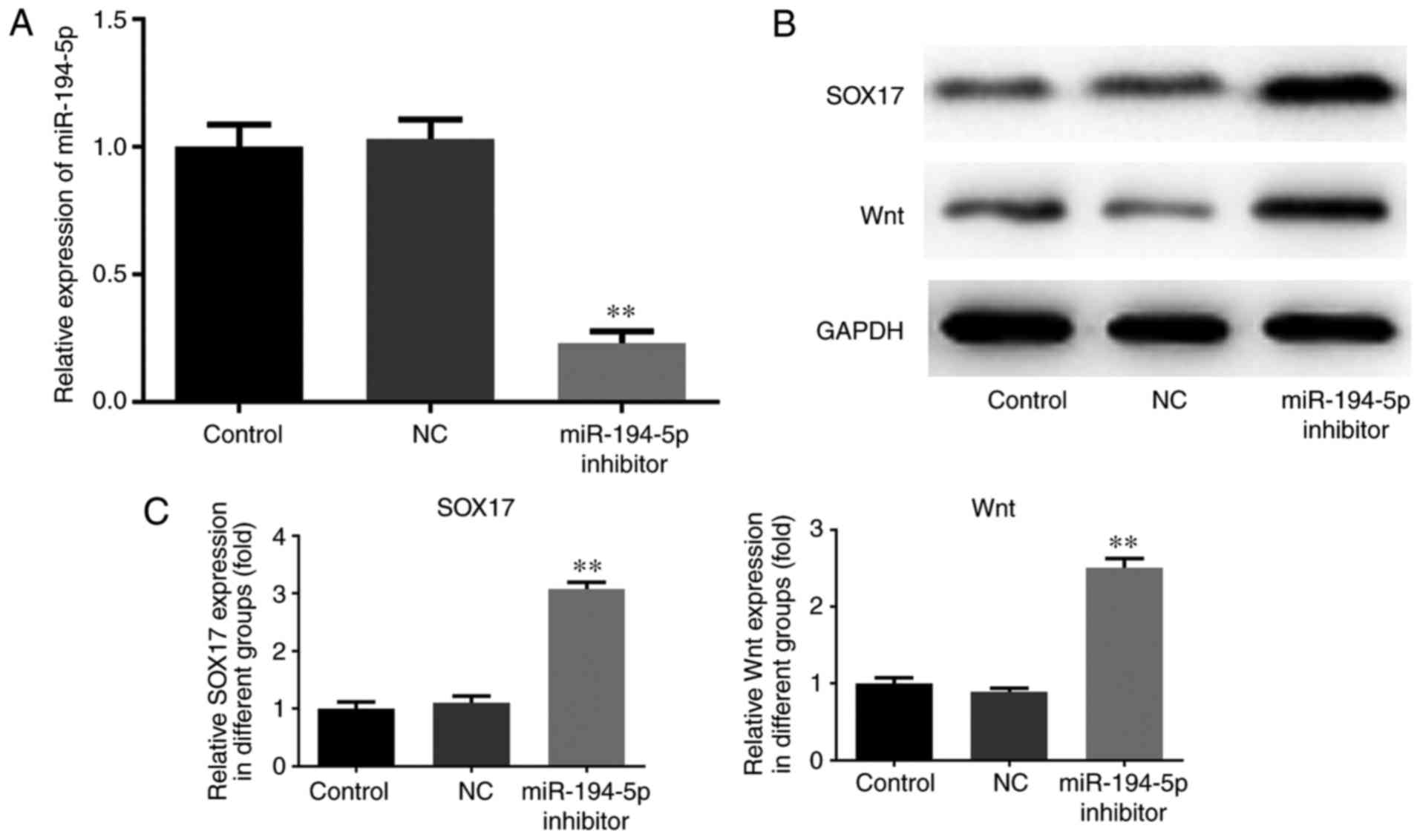
Knockdown of miR-194-5p promotes SOX17
and Wnt expression in MCF-7 cells
The expression levels of miR-194-5p were measured by
RT-qPCR, and the protein expression levels of SOX17 and Wnt in
transfected MCF-7 cells were determined by western blotting. As
shown in Fig. 3A, the expression
levels of miR-194-5p were markedly decreased in the miR-194-5p
inhibitor group compared with in the control groups. Furthermore,
the expression levels of SOX17 and Wnt were upregulated in the
miR-194-5p inhibitor-transfected group compared with in the control
groups (Fig. 3B and C).
Therefore, knockdown of miR-194-5p in MCF-7 cells may promote the
expression of SOX17.
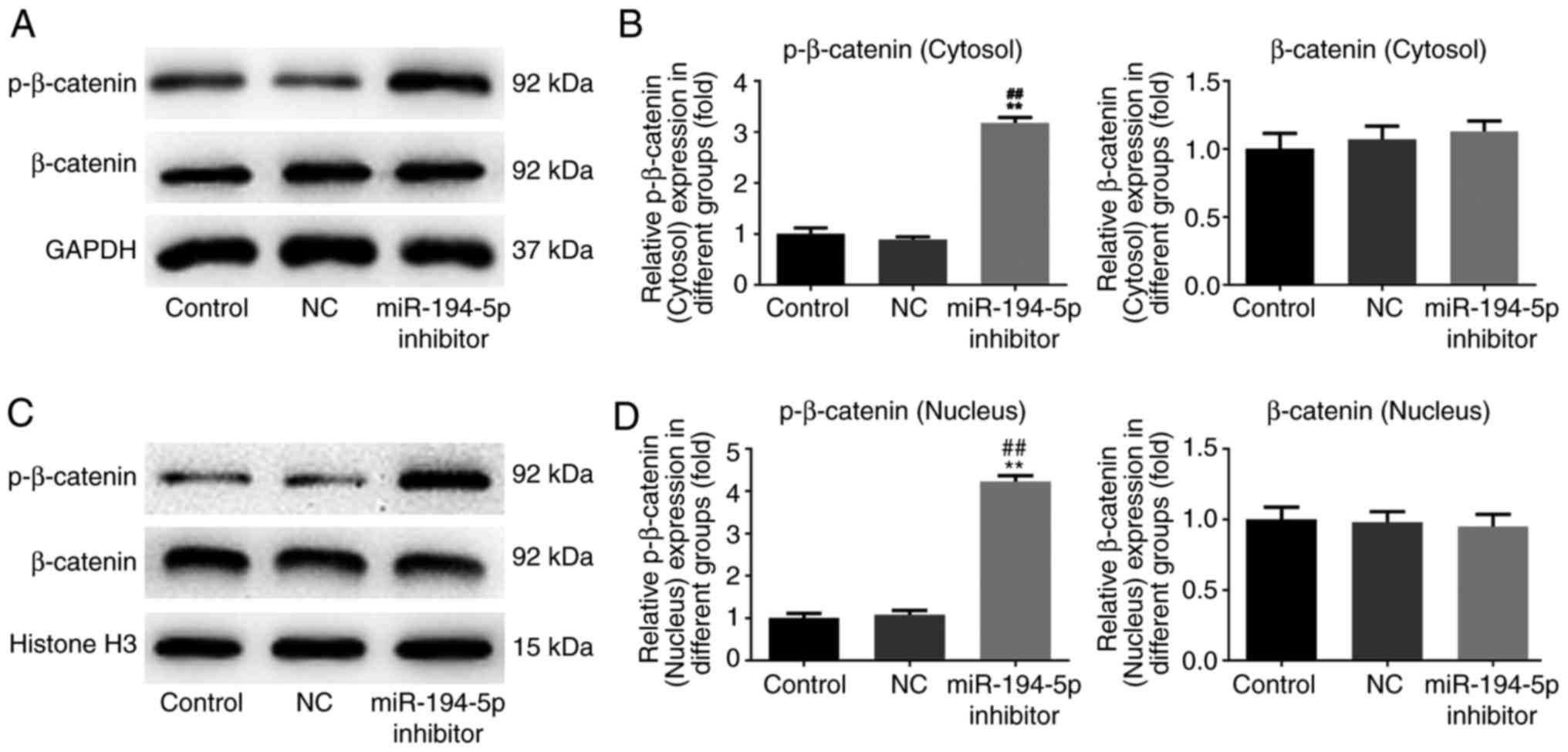
Knockdown of miR-194-5p upregulates the
expression of p-β-catenin in the cytosol and nucleus of MCF-7
cells
The cytosolic and nuclear protein expression levels
of p-β-catenin and β-catenin were also evaluated by western
blotting. The results demonstrated that p-β-catenin was
significantly upregulated in the cytosol and nucleus in the
miR-194-5p inhibitor group compared with in the control groups.
However, there was no difference in the expression levels of
β-catenin in the cytosol and nucleus between the miR-194-5p
inhibitor group and the control groups (Fig. 4). Therefore, knockdown of
miR-194-5p in MCF-7 cells might facilitate the expression of
p-β-catenin in the cytosol and nucleus, with no effects on
β-catenin expression.
Knockdown of miR-194-5p suppresses the
proliferation of MCF-7 cells
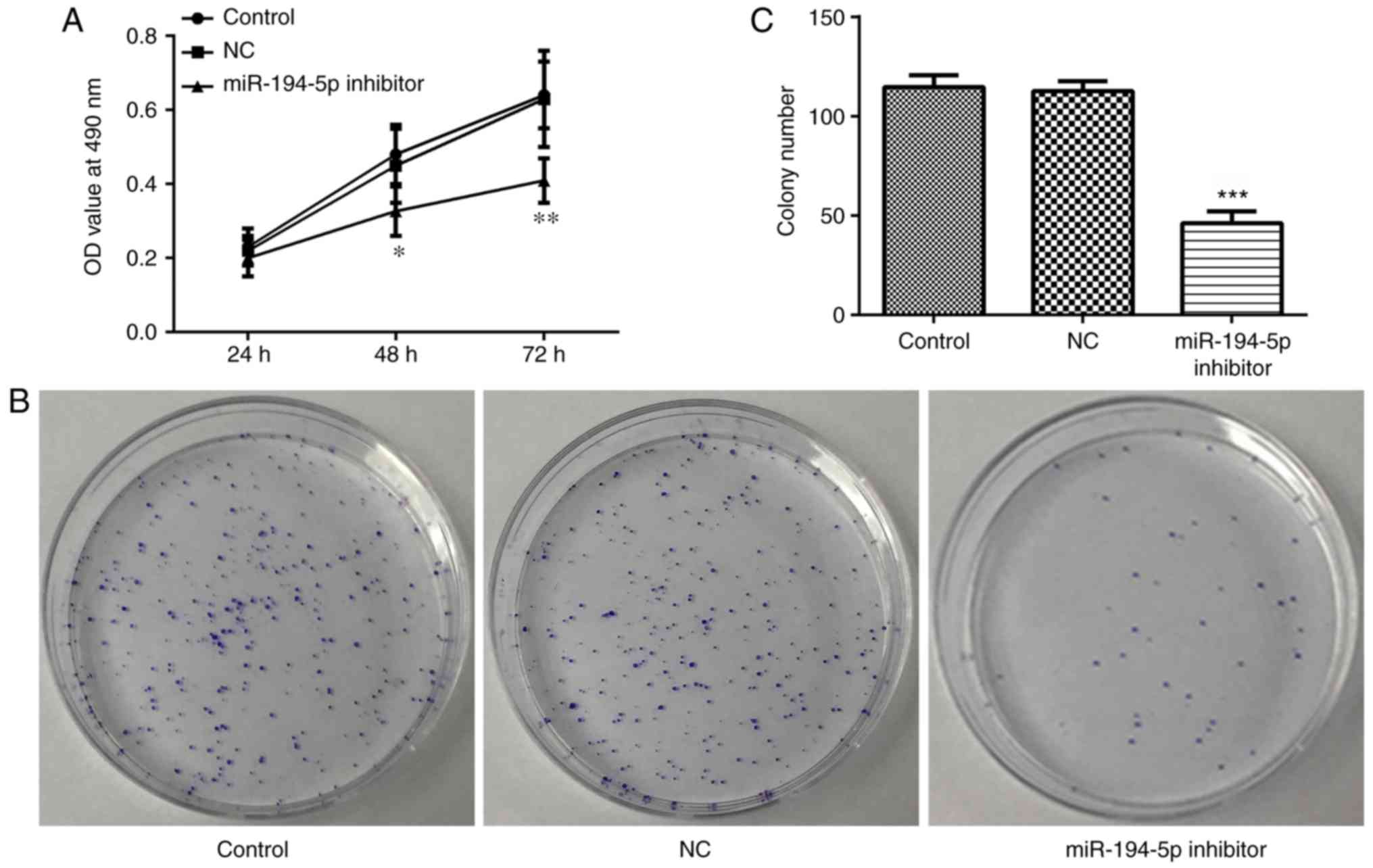
The MTT assay was performed to assess proliferation
of miR-194-5p inhibitor-transfected MCF-7 cells. The proliferation
rate was determined by OD values. As shown in Fig. 5A, proliferation was significantly
decreased in the miR-194-5p inhibitor group compared with in the
control groups. As presented in Fig.
5B and C, the proliferation rate was further determined by
colony formation assay. The results indicated that proliferation
was significantly decreased in the miR-194-5p inhibitor group.
These results suggested that knockdown of miR-194-5p may suppress
the proliferation of MCF-7 cells.
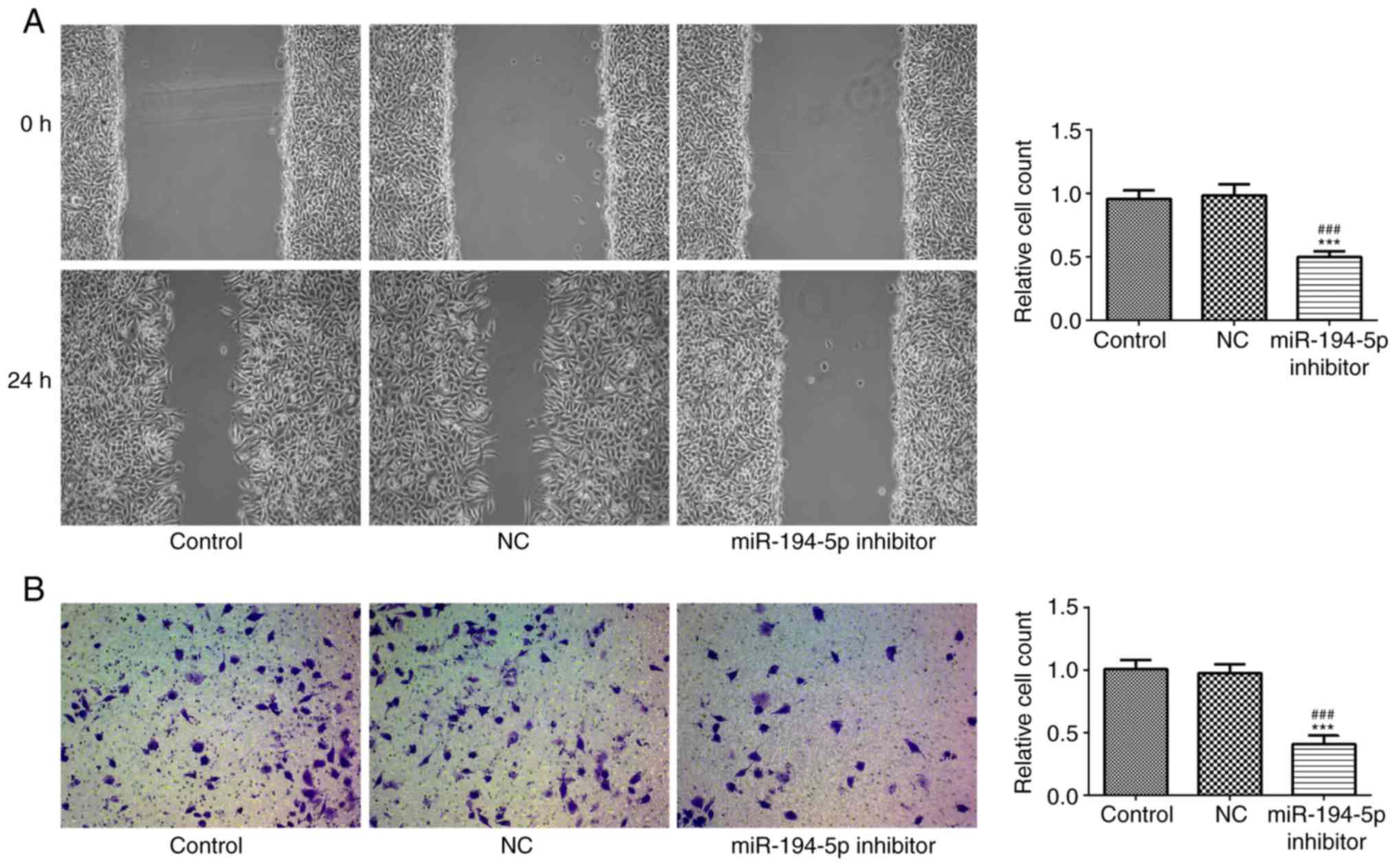
Knockdown of miR-194-5p inhibits the
migration and invasion of MCF-7 cells
The migration and invasion of transfected MCF-7
cells were measured by scratch and Transwell assays, respectively.
As shown in Fig. 6A and B, cell
migration and invasion were significantly inhibited in the
miR-194-5p inhibitor group compared with in the control groups.
Therefore, knockdown of miR-194-5p in MCF-7 cells might inhibit
cell migration and invasion.
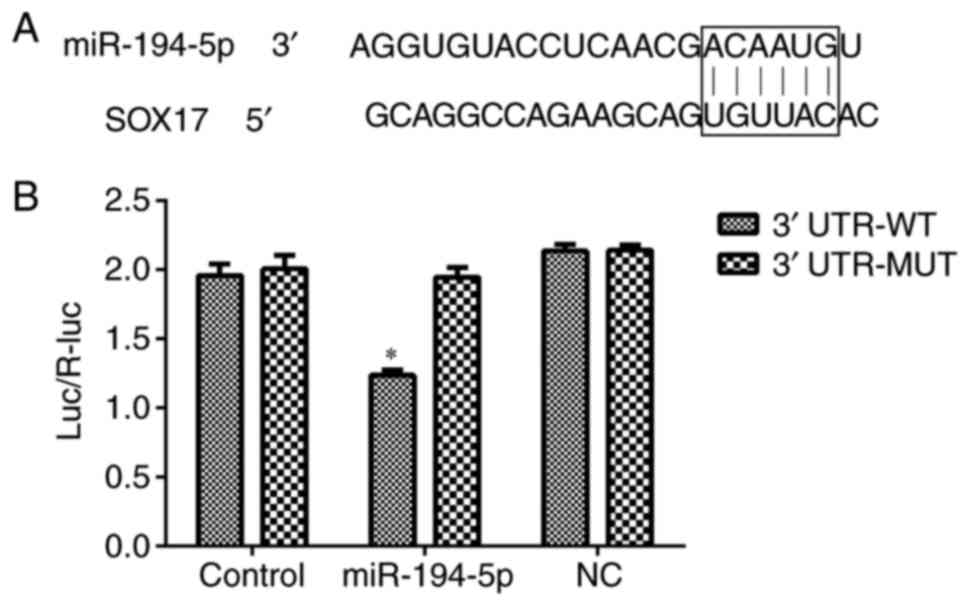
Luciferase reporter assay for target
verification
The results of a TargetScan analysis suggested that
SOX17 was a target gene of miR-194-5p, due to a complementary
binding region in its 3'UTR. A luciferase reporter assay was
performed to verify this prediction. As shown in Fig. 7, luciferase activity was decreased
in the SOX17 3'UTR WT group transfected with miR-194-5p mimics,
thus suggesting that SOX17 may be a direct target gene of
miR-194-5p.
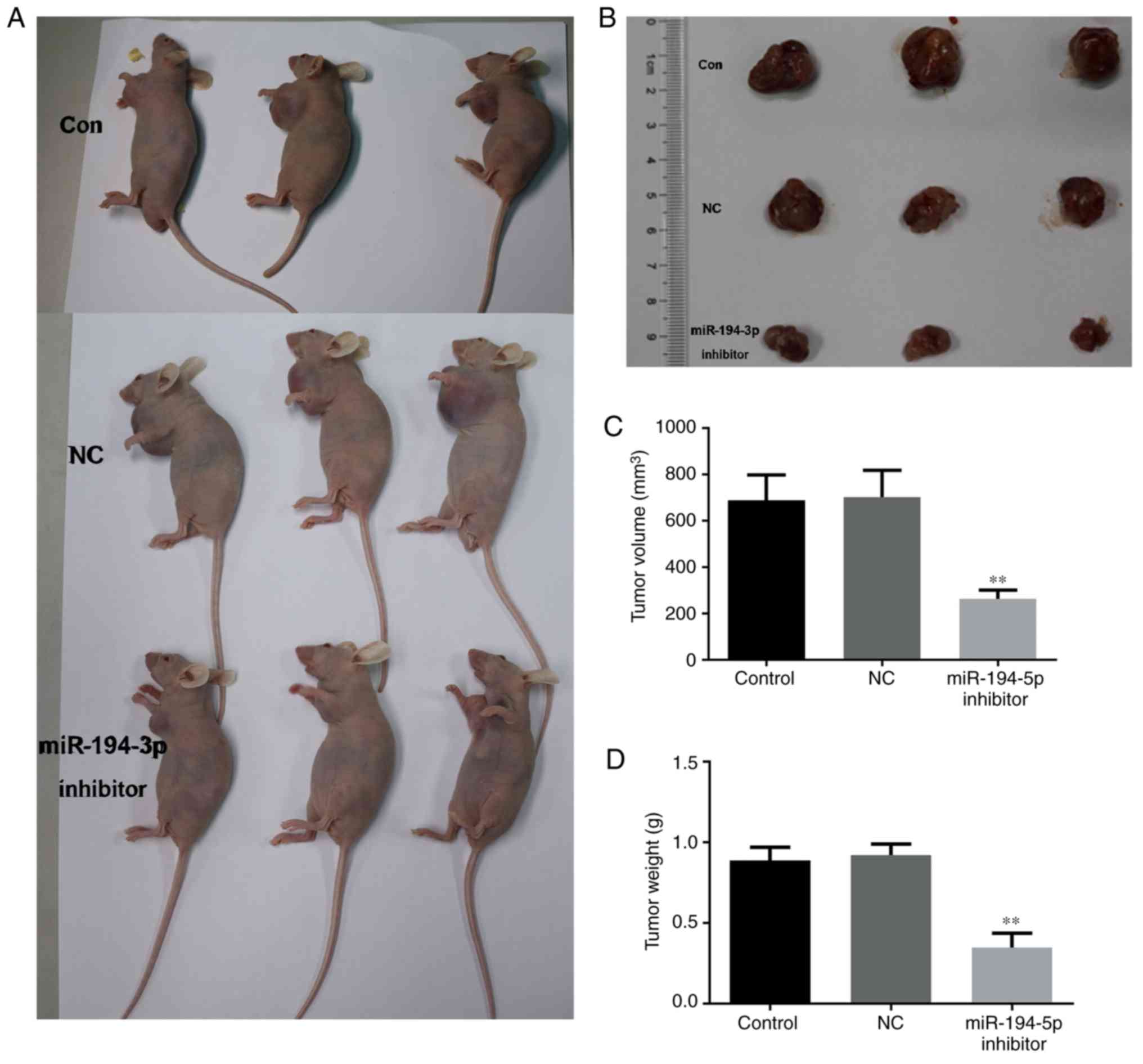
Knockdown of miR-194-5p inhibits tumor
growth in nude mice
A total of 20 days after injection with miR-194-5p
inhibitor-transfected cells, tumor growth was measured according to
tumor volume and weight. As shown in Fig. 8, tumor volume and weight were
significantly decreased in the miR-194-5p inhibitor group compared
within the control groups. These findings indicated that knockdown
of miR-194-5p may contribute to tumor growth suppression in breast
cancer.
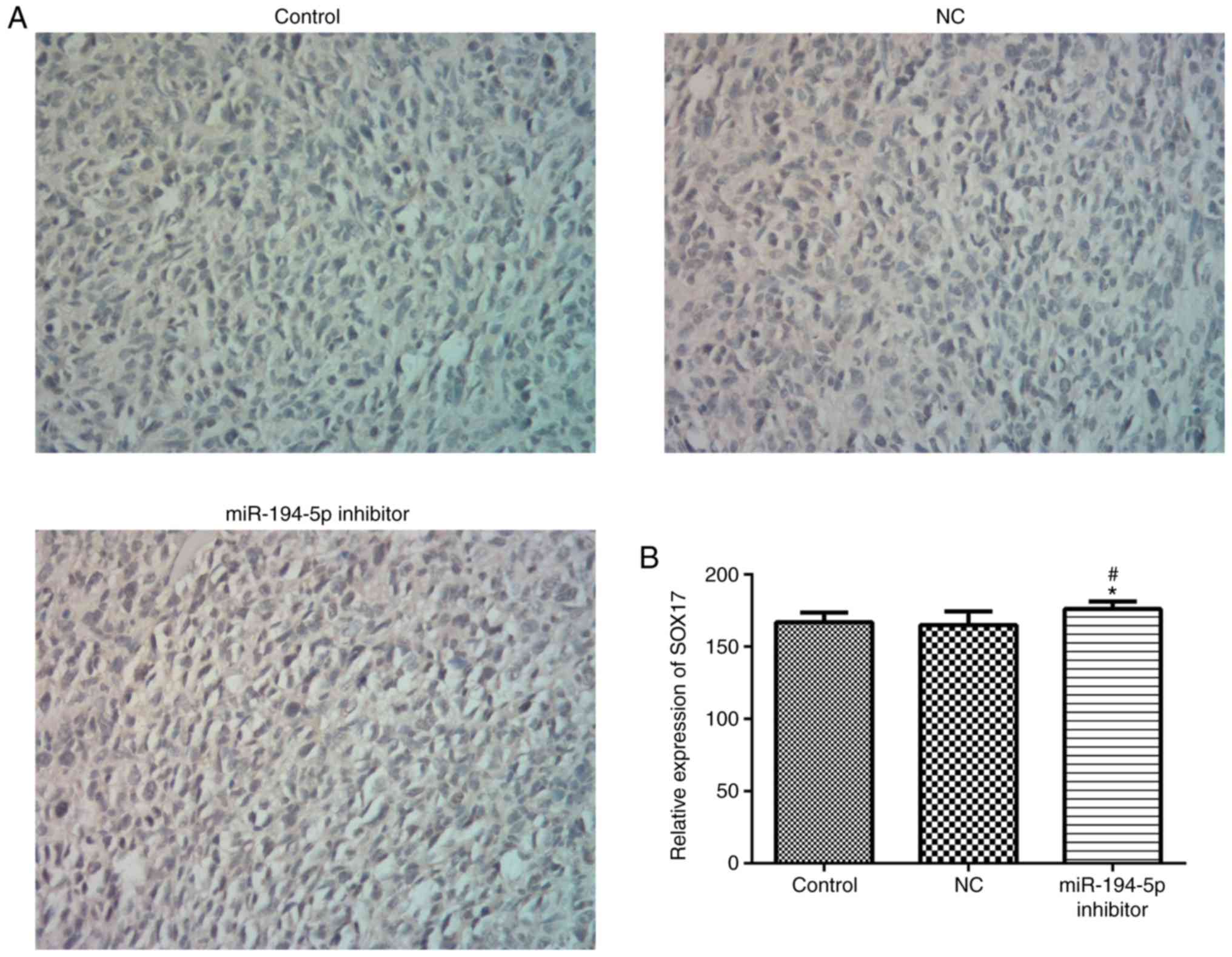
Knockdown of miR-194-5p promotes SOX17
expression
SOX17 expression was measured by IHC after injection
with miR-194-5p inhibitor-transfected cells for 20 days. The
results indicated that the expression levels of SOX17 were markedly
increased in the miR-194-5p inhibitor group compared with in the
control groups (Fig. 9).
Therefore, knockdown of miR-194-5p may give rise to the
upregulation of SOX17 in breast cancer tissues.
Discussion
The present study investigated the effects of
miR-194-5p knockdown on SOX17 and the Wnt/β-catenin signaling
pathway in breast cancer. The results demonstrated that miR-194-5p
was upregulated in breast cancer, which is consistent with the
previous findings by Huo et al (18). In a previous study, the SOX family
was confirmed as a target of miR-194, and it was revealed that the
expression of SOX is regulated by miR-194 (19). In the present study, SOX17 was
identified as a target gene of miR-194-5p. Subsequently, knockdown
of miR-194-5p resulted in the upregulation of SOX17. The present
study demonstrated that miR-194-5p knockdown upregulated the
expression levels of Wnt and p-β-catenin in MCF7 cells. An increase
in p-β-catenin by miR-194-5p knockdown may result in reduced
activation of the Wnt/β-catenin pathway. Furthermore, miR-194-5p
knockdown increased the expression of SOX17, which may have led to
inhibition of the Wnt/β-catenin signaling pathway; however,
increased Wnt expression indicated that this inhibition may involve
regulation of other factors. Therefore, knockdown of miR-194-5p may
have multiple effects on the Wnt/β-catenin pathway.
Numerous studies have reported that SOX17 is an
important tumor suppressor in various types of cancer, which can
modulate the Wnt/β-catenin signaling pathway by binding to LEF
(20-23). In addition, the interaction
between SOX17 and LEF leads to the reduced combination of SOX17 to
β-catenin, which further results in suppression of the
Wnt/β-catenin signaling pathway. The Wnt/β-catenin signaling
pathway is closely associated with numerous types of cancer due to
overactivated Wnt/β-catenin (24,25). Furthermore, it has been reported
that blockade of the Wnt/β-catenin signaling pathway could
effectively suppress breast cancer metastasis, invasion and cell
proliferation (26-29). The present study revealed that
knockdown of miR-194-5p inhibited cell proliferation, migration and
invasion of MCF-7 cells, which is in concordance with these
previous findings. Although the direct regulatory mechanism of
miR-194-5p knockdown on the Wnt/β-catenin pathway has not been
confirmed, it does seem to affect the activity of the signaling
pathway and therefore may have an effect on the Wnt/β-catenin
pathway. In addition, reduced tumor weight and volume in the
miR-194-5p inhibitor group of mice with breast cancer indicated
that miR-194-5p knockdown might suppress tumor growth. The results
of IHC demonstrated that knockdown of miR-194-5p was able to
promote the expression of SOX17 in mice with breast cancer. The
results of the present mouse studies were in line with the results
of experiments in cells, thus suggesting that miR-194-5p knockdown
may be an effective potential therapeutic strategy for breast
cancer.
However, the expression of SOX17 and the
Wnt/β-catenin signaling pathway is complex; therefore, we cannot
completely eliminate other forms of the interference, including
methylation of SOX17 and the effects of other miRNAs. It has been
reported that SOX17 promoter methylation leads to poor survival in
breast cancer (30).
Additionally, other miRNAs, such as the miR-200 family, miR-141 and
miR-26 (31-34), are associated with SOX17
expression and the Wnt/β-catenin signaling pathway. Therefore,
further experiments should be performed to deeply investigate the
association among miR-194-5p, SOX17 and the Wnt/β-catenin signaling
pathway.
In conclusion, the present results indicated that
miR-194-5p inhibitor-induced regulation of SOX17 expression and the
Wnt/β-catenin signaling pathway suppressed cell proliferation,
migration and invasion in breast cancer. Although the exact roles
of miR-194-5p and SOX17 in the Wnt/β-catenin signaling pathway
remain to be elucidated, they may be considered targets for the
inhibition and treatment of breast cancer progression.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FY designed the research. ZX collected and bred the
mice, and conducted animal studies. ZX and SZ performed the
experiments. FY and ZX analyzed the data and wrote the manuscript.
All authors reviewed the results and approved the final version of
the manuscript.
Ethics approval and consent to
participate
The human studies were approved by the ethics
committee of Yinzhou People's Hospital of Ningbo City (Ningbo,
China). Written informed consent was obtained from all patients.
The animal studies were approved by the laboratory animal
management and welfare ethical review committee of Yinzhou People's
Hospital of Ningbo City.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Howell A, Anderson AS, Clarke RB, Duffy
SW, Evans DG, Garcia-Closas M, Gescher AJ, Key TJ, Saxton JM and
Harvie MN: Risk determination and prevention of breast cancer.
Breast Cancer Res. 16:4462014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gray JM, Rasanayagam S, Engel C and Rizzo
J: State of the evidence 2017: An update on the connection between
breast cancer and the environment. Environ Health. 16:942017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ward R, Sims AH, Lee A, Lo C, Wynne L,
Yusuf H, Gregson H, Lisanti MP, Sotgia F, Landberg G and Lamb R:
Monocytes and macrophages, implications for breast cancer migration
and stem cell-like activity and treatment. Oncotarget.
6:14687–14699. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Need EF, Selth LA, Trotta AP, Leach DA,
Giorgio L, O'Loughlin MA, Smith E, Gill PG, Ingman WV, Graham JD
and Buchanan G: The unique transcriptional response produced by
concurrent estrogen and progesterone treatment in breast cancer
cells results in upregulation of growth factor pathways and
switching from a luminal a to a basal-like subtype. BMC Cancer.
15:7912015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bodai BI and Tuso P: Breast cancer
survivorship: A comprehensive review of long-term medical issues
and lifestyle recommendations. Perm J. 19:48–79. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lages E, Ipas H, Guttin A, Nesr H, Berger
F and Issartel JP: MicroRNAs: Molecular features and role in
cancer. Front Biosci (Landmark Ed). 17:2508–2540. 2012. View Article : Google Scholar
|
|
7
|
Greene SB, Herschkowitz JI and Rosen JM:
Small players with big roles: MicroRNAs as targets to inhibit
breast cancer progression. Curr Drug Targets. 11:1059–1073. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhu X, Li D, Yu F, Jia C, Xie J, Ma Y, Fan
S, Cai H, Luo Q, Lv Z and Fan L: miR-194 inhibits the
proliferation, invasion, migration, and enhances the
chemosensitivity of non-small cell lung cancer cells by targeting
forkhead box A1 protein. Oncotarget. 7:13139–13152. 2016.PubMed/NCBI
|
|
9
|
Okato A, Arai T, Yamada Y, Sugawara S,
Koshizuka K, Fujimura L, Kurozumi A, Kato M, Kojima S, Naya Y, et
al: Dual strands of pre-miR-149 inhibit cancer cell migration and
invasion through targeting FOXM1 in renal cell carcinoma. Int J Mol
Sci. 18:E19692017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dell'Aversana C, Giorgio C, D'Amato L,
Lania G, Matarese F, Saeed S, Di Costanzo A, Belsito Petrizzi V,
Ingenito C, Martens JHA, et al: miR-1945p/BCLAF1 deregulation in
AML tumorigenesis. Leukemia. 31:2315–2325. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chang HY, Ye SP, Pan SL, Lania G, Matarese
F, Saeed S, Di Costanzo A, Belsito Petrizzi V, Ingenito C, Martens
JHA, et al: Overexpression of miR-194 reverses HMGA2-driven
signatures in colorectal cancer. Theranostics. 7:3889–3900. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hironaka-Mitsuhashi A, Matsuzaki J,
Takahashi RU, Yoshida M, Nezu Y, Yamamoto Y, Shiino S, Kinoshita T,
Ushijima T, Hiraoka N, et al: A tissue microRNA signature that
predicts the prognosis of breast cancer in young women. PLoS One.
12:e01876382017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ring A, Kim YM and Kahn M: Wnt/catenin
signaling in adult stem cell physiology and disease. Stem Cell Rev.
10:512–525. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lien WH and Fuchs E: Wnt some lose some:
Transcriptional governance of stem cells by Wnt/β-catenin
signaling. Genes Dev. 28:1517–1532. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Aminuddin A and Ng PY: Promising druggable
target in head and neck squamous cell carcinoma: Wnt signaling.
Front Pharmacol. 7:2442016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yin D, Jia Y, Yu Y, Brock MV, Herman JG,
Han C, Su X, Liu Y and Guo M: SOX17 methylation inhibits its
antagonism of Wnt signaling pathway in lung cancer. Discov Med.
14:33–40. 2012.PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
18
|
Huo D, Clayton WM, Yoshimatsu TF, Chen J
and Olopade OI: Identification of a circulating microRNA signature
to distinguish recurrence in breast cancer patients. Oncotarget.
7:55231–55248. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jung KH, McCarthy RL, Zhou C, Uprety N,
Barton MC and Beretta L: MicroRNA regulates hepatocytic
differentiation of progenitor cells by targeting YAP1. Stem Cells.
34:1284–1296. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu X, Luo M, Xie W, Wells JM, Goodheart
MJ and Engelhardt JF: Sox17 modulates Wnt3A/beta-catenin-mediated
transcriptional activation of the Lef-1 promoter. Am J Physiol Lung
Cell Mol Physiol. 299:L694–L710. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang H, Lee S, Lee S, Kim K, Yang Y, Kim
JH, Adams RH, Wells JM, Morrison SJ, Koh GY and Kim I: Sox17
promotes tumor angiogenesis and destabilizes tumor vessels in mice.
J Clin Invest. 123:418–431. 2013. View
Article : Google Scholar :
|
|
22
|
Lv Li, He ZY, Wang G, Zhang J, Lu X, Ren
G, Wang X, Zhu F, Ding XY, et al: The SOX17/miR-3715p/SOX2 axis
inhibits EMT, stem cell properties and metastasis in colorectal
cancer. Oncotarget. 6:9099–9112. 2015.
|
|
23
|
Guimarães-Young A, Neff T, Dupuy AJ and
Goodheart MJ: Conditional deletion of Sox17 reveals complex effects
on uterine adenogenesis and function. Dev Biol. 414:219–227. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gao L, Chen B, Li J, Yang F, Cen X, Liao Z
and Long X: Wnt/β-catenin signaling pathway inhibits the
proliferation and apoptosis of U87 glioma cells via different
mechanisms. PLoS One. 12:e01813462017. View Article : Google Scholar
|
|
25
|
Lisanti MP, Tsirigos A, Pavlides S, Reeves
KJ, Peiris-Pagès M, Chadwick AL, Sanchez-Alvarez R, Lamb R, Howell
A, Martinez-Outschoorn UE and Sotgia F: JNK1 stress signaling is
hyper-activated in high breast density and the tumor stroma:
Connecting fibrosis, inflammation, and stemness for cancer
prevention. Cell Cycle. 13:580–599. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jang GB, Kim JY, Cho SD, Park KS, Jung JY,
Lee HY, Hong IS and Nam JS: Blockade of Wnt/β-catenin signaling
suppresses breast cancer metastasis by inhibiting CSC-like
phenotype. Sci Rep. 5:124652015. View Article : Google Scholar
|
|
27
|
Dey N, Barwick BG, Moreno CS,
Ordanic-Kodani M, Chen Z, Oprea-Ilies G, Tang W, Catzavelos C,
Kerstann KF, Sledge GW Jr, et al: Wnt signaling in triple negative
breast cancer is associated with metastasis. BMC Cancer.
13:5372013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu T, Hu K, Zhao Z, Chen G, Ou X, Zhang
H, Zhang X, Wei X, Wang D, Cui M and Liu C: MicroRNA-1
down-regulates proliferation and migration of breast cancer stem
cells by inhibiting the Wnt/β-catenin pathway. Oncotarget.
6:41638–41649. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yin X, Xiang T, Li L, Su X, Shu X, Luo X,
Huang J, Yuan Y, Peng W, Oberst M, et al: DACT1, an antagonist to
Wnt/β-catenin signaling, suppresses tumor cell growth and is
frequently silenced in breast cancer. Breast Cancer Res.
15:R232013. View
Article : Google Scholar
|
|
30
|
Fu D, Ren C, Tan H, Wei J, Zhu Y, He C,
Shao W and Zhang J: Sox17 promoter methylation in plasma DNA is
associated with poor survival and can be used as a prognostic
factor in breast cancer. Medicine (Baltimore). 94:e6372015.
View Article : Google Scholar
|
|
31
|
Manavalan TT, Teng Y, Litchfield LM,
Muluhngwi P, Al-Rayyan N and Klinge CM: Reduced expression of
miR-200 family members contributes to antiestrogen resistance in
LY2 human breast cancer cells. PLoS One. 8:e623342013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang G, Zhang W, Li B, Stringer-Reasor E,
Chu C, Sun L, Bae S, Chen D, Wei S, Jiao K, et al: MicroRNA-200c
and microRNA- 141 are regulated by a FOXP3-KAT2B axis and
associated with tumor metastasis in breast cancer. Breast Cancer
Res. 19:732017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Peng Y, Zhang X, Feng X, Fan X and Jin Z:
The crosstalk between microRNAs and the Wnt/β-catenin signaling
pathway in cancer. Oncotarget. 8:14089–14106. 2017.
|
|
34
|
Jia Y, Yang Y, Zhan Q, Brock MV, Zheng X,
Yu Y, Herman JG and Guo M: Inhibition of SOX17 by microRNA 141 and
methylation activates the WNT signaling pathway in esophageal
cancer. J Mol Diagn. 14:577–585. 2012. View Article : Google Scholar : PubMed/NCBI
|