Introduction
In Asia, diabetes is a common and disease with an
increasing prevalence rate estimated (1). Diabetes has become increasingly
widespread and is now recognized as an epidemic disease in China
which is specifically analyzed (2-4).
Diabetic cardiomyopathy (DCM) is recognized as a primary
complication and the main cause of morbidity and mortality in
diabetic patients, who have a 2-5-fold increased risk of developing
heart failure, reduced forced vital capacity and forced expiratory
volume/sec when compared with non-diabetic individuals (5,6).
Myocardial fibrosis, known as a pathological entity of
extracellular matrix remodeling, often induces myocardial stiffness
and exaggerates cardiac dysfunction (7). Myocardial fibrosis and cardiac
function are vital indicators of diabetes, and have been analyzed
and investigated previously (8-10).
Treatment against fibrosis with the early utilization of
angiotensin-converting enzyme inhibitors may be a potential
diabetic strategy (11).
Dimethylarginine dimethylaminohydrolase (DDAH) dysregulation has
been found to exert notable effect during the progression of
diabetic nephropathy through elevating asymmetric dimethylarginine
levels (12). Therefore, the
present study investigated the role of DDAH2 under experimental
diabetic conditions to test the hypothesis that DDAH2 dysregulation
is involved in the progression of fibrosis in DCM.
DDAH is an enzyme which can metabolize competitive
endogenous inhibitors of nitric oxide (NO) synthase (NOS),
including asymmetric NG,
NGdimethyl-L-arginine (ADMA) and
NG-monomethyl-L-arginine (13). DDAH consists of two isoforms,
namely DDAH1 and DDAH2 in mammals, which are expressed in the
cardiovascular system (14). Of
note, the expression of DDAH2 was shown to be noticeably
downregulated in the adipose tissue of diabetic rats (15). Diabetic endothelial dysfunction
may arise from the uncoupling of endothelial nitric oxide synthase
(eNOS) by forming superoxide anion [O(2)(−)] (16). ADMA, a naturally existing
L-arginine analog endogenously synthesized during the methylation
of protein arginine residues, is an inhibitor of NOS by competing
with L-arginine to bind to the active site of NOS, thus suppressing
NO synthesis and causing NOS uncoupling (17). The majority of ADMA can be
intracellularly degraded into citrulline and dimethylamine by DDAH
(18). A previous study revealed
that the level of exogenous ADMA increased, whereas the
overexpression of DDAH2 suppressed the senescence of high
glucose-induced endothelial progenitor cells (19). In addition, the activity of DDAH
is involved in endothelial dysfunction in diabetes, indicating that
preservation of the activity of DDAH in vessels and the reduction
of endogenous ADMA may underlie the protective mechanism
alleviating the impairment of endothelium-dependent vasodilatation
(20). In this regard, the
expression of DDAH2 was associated with endothelial dysfunction in
diabetic rats or cell damage in a high glucose environment
(19,20). Considering the role of DDAH2 in
diabetes, the present study hypothesized that DDAH2 may be
implicated in DCM through involvement of the DDAH/ADMA/NOS/NO
pathway. The present study was performed to examine whether DDAH2
is a novel target for improving the clinical outcome of DCM by
ameliorating cardiac function and myocardial fibrosis via the
DDAH/ADMA/NOS/NO pathway.
Materials and methods
Ethical statement
The animal experiments were approved by The First
People’s Hospital of Yunnan Province, Affiliated Hospital of
Kunming Science and Technology University (Kunming, China). Efforts
were made to minimize the suffering of animals. All experimental
procedures and protocols were in line with the recommendations of
the Guide for the Care and Use of Laboratory Animals of the
American National Institutes of Health.
Animal treatment
A total of 100 Wistar male rats (weighing 190-200 g;
aged 6 weeks) were purchased from the Laboratory Animal Center,
Kunming Medical University (Kunming, China) for the following in
vivo experiments. Streptozotocin (STZ) (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) injection was used for diabetic rat model
establishment. STZ was dissolved in 0.1 mmol/l citric acid buffer
(pH 4.5) to prepare 1% STZ solution. The rats were
intraperitoneally injected with STZ solution (30 mg/kg). After 72
h, venous blood from the rat tails was collected, with blood
glucose (BG) ≥16.7 mmol/l considered successful model
establishment. The rate of successful modeling was 83% (83/100). Of
the 100 rats, 10 died and seven rats were unsuccessfully modeled
with BG <16.7 mmol/l. The rats successfully modeled were
randomly grouped into a model group, DDAH2 group, DDAH2-NC group,
short hairpin RNA (sh)DDAH2 group, shDDAH2-NC group, L-NNA group
and L-NNA + DDAH2 group, with 10 animals in each group.
Subsequently, 2.5×1010 PFU/ml DDAH2 lentivirus (2
µl), DDAH2-NC lentivirus, shDDAH2 lentivirus (2 µl),
shDDAH2-NC lentivirus (2 µl) and shDDAH2-NC lentivirus (2
µl) were injected in the tail vein of the rats in the DDAH2
group, the DDAH2-NC group, the shDDAH2 group, the shDDAH2-NC group,
and the L-NNA+DDAH2 group respectively. All lentivirus vectors were
produced by Beijing Nuosai Biotechnology Co., Ltd. (Beijing,
China). After 1 week, the injection was repeated again. Meanwhile,
the NOS inhibitor L-NNA (5 mg/kg) was intraperitoneally injected
into the rats of the L-NNA and L-NNA + DDAH2 groups following
establishment of the rat models. The same dose of NOS inhibitor
L-NNA was injected again after 20 h. The rats were housed at room
temperature with a relative humidity of 50-60%, free access to food
and water, and a 12-h light/dark cycle.
Cell grouping and culture
The H9C2 primary rat myocardial cell line (Institute
of Biochemistry and Cell Biology, Shanghai Institutes for
Biological Sciences, Chinese Academy of Sciences, Shanghai, China)
was used for in vitro experiments. The cells were assigned
into the model group (H9C2 cells treated with high glucose), DDAH2
group (H9C2 cells treated with high glucose and transfected with
DDAH2 plasmid), DDAH2-NC group (H9C2 cells treated with high
glucose and transfected with DDAH2-NC plasmid), shDDAH2 group (H9C2
cells treated with high glucose and transfected with shDDAH2
plasmid), shDDAH2-NC group (H9C2 cells treated with high glucose
and transfected with shDDAH2-NC plasmid), L-NNA group (H9C2 cells
treated with high glucose and incubated with 50 µmol/l L-NNA
for 48 h) and L-NNA + DDAH2 group (H9C2 cells treated with high
glucose, incubated with 50 µmol/l NOS inhibitor L-NNA for 48
h and transfected with DDAH2 plasmid). The cells were cultured in
Dulbecco’s modified Eagle’s medium containing 10% fetal bovine
serum (FBS) (all purchased from Sigma-Aldrich; Merck KGaA)
1×105 U/l penicillin, and 100 mg/l streptomycin at 37°C
with 5% CO2. The medium was changed every 2 days. The
cells were subcultured when cell confluence reached 80-90%. The
cells prepared for high glucose treatment were cultured in culture
medium with 30 ml/l glucose, and cells at the exponential growth
phase were used for subsequent experiments.
Construction and transfection of
plasmid
According to the known DDAH2 sequence in NCBI
(https://www.ncbi.nlm.nih.gov/nuccore/NM_212532.2),
the DDAH2 plasmid, DDAH2-NC plasmid, shDDAH2 plasmid, and
shDDAH2-NC plasmid were constructed by Sangon Biotech Co., Ltd.,
(Shanghai, China). At the same time, three different plasmids were
screened to avoid off-target effects.
The third generation cells were detached by trypsin
and inoculated into a 24-well plate for the cells to grow into
monolayers with medium discarded. Transfection was performed using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) according to the manufacturer’s protocol. The
procedure was as follows: H9C2 cells were seeded into a 6-well
plate at a density of 2×105 cells/well and, when the
cells had adhered to the wall, the medium was replaced with culture
medium without penicillin/streptomycin 12 h prior to transfection.
Subsequently, 100 µl liposome was diluted in 250 µl
OPTI-MEM medium, triturated with a pipette and mixed prior to
incubation at room temperature for 5 min. The plasmid vectors (4
µg) were diluted in 250 µl OPTI-MEM, triturated with
a pipette and mixed prior to incubation at room temperature for 20
min, and then added to a 6-well plate containing 1.5 ml OPTI-MEM
and mixed. The cells were incubated in a 5% CO2
incubator at 37°C, and the medium was replaced with complete medium
at 6 h. To screen out the stably transfected cells at 48 h
post-transfection, the cells were seeded into G418 (1,000-2,000
µg/ml) medium for 4 weeks and the medium was replaced every
3-5 days.
Assessment of cardiac function in
rats
Following lentivirus injection for 12 weeks, five
rats in each group were heparinized by injection of 0.3% heparin (2
mg/kg) into the tail vein, intraperitoneally injected with 3%
sodium pentobarbital (40 mg/kg) and anesthetized. Following
fixation, the right common carotid artery of the rats was bluntly
separated via a cervical median incision with the arteries ligated
at the distal end of the heart. Subsequently, the proximal end of
the heart was clamped and a small opening was cut in the artery
wall to insert a catheter connected to the pressure transducer
(Miller, Houston, TX, USA). The catheter was then placed into the
left ventricle for several minutes with the ligature released. The
catheter was connected to the Power Lab biometrics experiment
system channel (AD Instruments, Oxford, UK) via a pressure
transducer to measure the left ventricular end-diastolic pressure
(LVEDP), left ventricular systolic pressure (LVSP), and the maximum
rate of left ventricular pressure rise/fall (LV ± dp/dt).
Blood biochemical assessment
The BG was measured in five rats selected from each
group using the glucose oxidase method at 0, 5, 15 and 20 weeks.
The steps of the glucose oxidase method were as follows: 10
µl plasma was added to 1.5 ml enzyme-buffer working
solution, mixed and placed in a 37°C water bath for 10 min. A
microplate reader (MK3, Thermo Fisher Scientific, Inc.) was used
for assessment of the optical density (OD) values at 500 nm. The
glucose levels were calculated according to formulas. The levels of
total cholesterol (TC), triglyceride (TG) and fasting blood glucose
(FBG) in the rats were analyzed using the Bayer 1650 blood
chemistry analyzer (Bayer AG, Leverkusen, Germany). An
enzyme-linked immunosorbent assay was used to detect the fasting
insulin (FINS) levels of the rats in each group.
Masson staining
Following the analysis of blood biochemistry, the
five rats in each group were sacrificed, limbs were fixed, the
chest was opened in a sterile environment and the heart was
removed. The heart was gently washed in phosphate-buffered saline
(PBS) and then the residual blood in heart was extruded. The heart
was trimmed and myocardial tissue samples of each group were
obtained. The myocardial tissues were used for Masson staining and
hematoxylin and eosin (H&E) staining, DDAH activity detection,
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) analysis and western blot analysis.
The myocardial tissues were fixed with 4%
paraformaldehyde, embedded with paraffin, and cut into sections
with a thickness of 4-µm. Following dewaxing, the tissue
sections were stained with hematoxylin for 5 min, differentiated
with hydrochloric acid ethanol, and then stained with Masson
ponceau acid fuchsin solution for 5 min. Following rapid washing
under water, the sample sections were stained with 1%
phosphomolybdic acid for 3 min and stained with toluidine blue for
5 min prior to drying. Following this, the sections were mounted
with gum and observed under a microscope (Nikon Ecliose 80i, Nikon
Corporation, Tokyo, Japan).
H&E staining
The paraffin sections in each group were dewaxed and
stained with hematoxylin for 5 min, followed by differentiation
with ethanolic hydrochloric acid. The paraffin sections were washed
and stained with eosin for 2 min. The sample sections were then
quickly washed with water, dried, mounted with gum and observed
under the microscope (Nikon Eclipse 80i, Nikon Corporation).
Transwell assay
Following transfection for 24 h, the cells in each
group were detached with trypsin and resuspended in serum-free
medium containing bovine serum albumin (Sigma-Aldrich; Merck KGaA)
for preparation of the cell suspension. Subsequently, 200 µl
cell suspension was added to an 8-mm Transwell chamber (Costar,
High Wycombe, UK) and placed in a 24-well plate. A total of 500
µl medium containing 20% FBS was added to the basolateral
chamber for incubation for 24 h at 37°C with 5% CO2.
Cells in the apical chamber were wiped off with a cotton swab,
washed with PBS 3-5 times, fixed with methanol for 20 min, and
stained with 0.1% crystal violet for 20 min. Under an inverted
microscope (Olympus, Corporation, Tokyo, Japan), five visual fields
were randomly selected and images were captured. The experiment was
repeated three times with the mean value obtained.
Measurement of ADMA concentration,
activity of DDAH in myocardial tissue, activity of NOS and NO
content in serum
The ADMA concentration was determined by high
performance liquid chromatography. Five rats were selected from
each group, and 1 ml serum from the rat was added to
5-sulfosalicylic acid precipitation protein at 4°C and centrifuged
at 2,000 × g for 5 min at 4°C. Following that, 10 µl serum
or standard samples were added to 100 µl derivatization
reagent (mixture of phthalaldehyde, borate buffer and
D-mercaptoethanol), triturated with pipette fully, mixed and placed
at room temperature for 3 min. The samples were then eluted using a
linear gradient (mobile phase: 4% acetonitrile and 0.4%
trifluoroacetic acid; flow rate: 0.2 ml/min). ADMA was then
separated on columns in samples and internal standard samples,
prior to being determined using a mass spectrometer (MS-2010;
ADMAM/Z: 203, N2 flow rate: 4.0 l/min). Subsequently, the cells in
each group were detached with 0.25% trypsin and centrifuged at
1,000 × g for 5 min at 4°C with the supernatant discarded. The
cells were resuspended with 1 ml ice phosphate buffer (pH 6.5, 0.1
mol/l), split by sonication, and then centrifuged at 3,500 × g for
30 min at 4°C. Subsequently, the ADMA concentration in 50 µl
of supernatant was measured as described above.
The activity of DDAH in myocardial tissues was
determined as follows: 0.1 mmol/l L-proline (500 µl) were
taken from a standard tube, and 50 µl myocardial homogenate
and 50 µl cell lysate were taken from a testing tube and a
control tube respectively. Subsequently, 100 µl ADMA
standard solution (1 mmol/l) was added into the testing tube and an
equal volume of PBS was added into the control tube and placed at
room temperature for 2 h. To each tube, 10% trichloroacetic acid
(0.5 ml) was added and centrifuged at 1,000 × g for 10 min at 4°C.
Following the addition of 0.5 ml of supernatant into a clean glass
test tube, 0.5 ml ddH2O, 1 ml colorimetric solution
containing 0.8% diacetyl monohydrazine and 0.5% antipyrine at the
ratio of 2:1 were added, prior to sealing with preservative film.
The tubes were placed in a water bath at 60°C for 100 min and
cooled in ice water for several seconds without exposure to light,
following which the OD value at 466 nm of each tube was determined
with a full-wavelength UV spectrophotometer. The enzymatic activity
of DDAH was the quantity of DDAH required to form 1 µmol/l
L-proline per min at 37°C (expressed as U/g protein).
A NOS activity assay kit (Nanjing Jiancheng
Biotechnology Co., Ltd., Nanjing, China) was used to measure NOS
activity. The procedure was as follows: 50 µl supernatant or
cell lysate of myocardial tissues following homogenization and
centrifugation was added into a testing tube, and 50 µl
ddH2O was added into a control tube. In each tube, 200
µl substrate buffer, 10 µl accelerant and 100
µl color reagent were added, mixed and incubated for 15 min
at 37°C. Following the addition of 100 µl clear reagent and
2 ml stop solution, the tissues were mixed and placed under plate
reader (MK3, Thermo Fisher Scientific, Inc.) for assessment of the
OD value at 530 nm. The NOS activity in each group was calculated
according to following formulae: NOS activity = (OD testing
tube-OD control tube)/molar extinction coefficient
× (the total volume of reaction liquid/the volume of sample) ×
(1/the time of reaction/the diameter of colorimetric ware)/the
content of protein.
A NO content assay kit (Nanjing Jiancheng
Biotechnology Co., Ltd., Nanjing, China) was used to determine the
NO content. The procedures were as follows: 0.5 ml supernatant or
cell lysate of myocardial tissues following homogeni-zation and
centrifugation was added into a testing tube, 0.5 ml
ddH2O into the control tube and 0.4 ml ddH2O
and 0.1 ml 10 µmol/l standard liquid into the standard tube.
Subsequently, 0.4 ml Reagent One and 0.4 ml Reagent Two were added
into each tube and mixed fully with vortex and placed in a water
bath at 37°C for 60 min. Subsequently, 0.2 ml Reagent Three and 0.1
ml Reagent Four were mixed fully for 30 sec, and allowed to stand
for 40 min at room temperature. Following centrifugation at 3,500 ×
g for 10 min at 4°C, 0.8 ml supernatant and 0.6 ml color reagent
were added, mixed thoroughly with a vortex and allowed to stand for
10 min. The OD values of the culture medium at 550 nm were measured
using a microplate reader (MK3, Thermo Fisher Scientific, Inc.).
The NO content in each group was calculated according to formulas:
NO content (µmol/gprot) = (OD
testing tube-OD control tube)/(OD
standard tube-OD control tube) ×
concentration standard liquid (20
µmol/l)/protein concentration
testing sample (gprot/l).
RT-qPCR analysis
Total RNA was extracted from the tissues and cells
in each group using TRIzol Reagent (Takara Biotechnology, Co.,
Ltd., Dalian, China) with the RNA purity and concentration measured
using a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies,
Inc. Rockland, DE, USA). Subsequently, the PrimeScript RT reagent
kit (Takara Biotechnology Co., Ltd.) was used for RT of the RNA
into cDNA. The reaction system was as follows: 0.5 µl
PrimeScript RT Enzyme mix, 4 µl 5X PrimeScript buffer, 1
µl RT primer, total RNA of 500 ng, with RNase Free
ddH2O added to a total PCR volume of 20 µl. The
reaction conditions were as follows: At 37°C for 15 min and at 85°C
for 5 sec. A UV spectrophotometer and 1% agarose gel were used to
measure DNA concentration and purity (A260/A280 >1.8).
The prepared cDNA was subjected to the following
experiments using the SYBR Premix Ex Taq II kit (Takara Bio, Inc.,
Tokyo, Japan) with glyceraldehyde-3-phosphate dehydrogenase (GAPDH)
used as an internal reference. The reaction conditions were as
follows: Pre-denaturation at 94°C for 10 min, 25 cycles of
denaturation at 94°C for 1 min, annealing at 60°C for 45 sec and
extension at 72°C for 2 min. The reaction system was as follows: 1
µl template cDNA, 0.4 µl forward primers and 0.4
µl reverse primers, 10 µl SYBR Premix Ex Taq, and 0.4
µl of ROX (50X), with ddH2O added to a total PCR
volume of 20 µl. The primers used are shown in Table I. All RT-qPCR experiments were
performed using an ABI7500 quantitative PCR instrument (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The dissolution curves
at 60-90°C were drawn to ensure that single products were amplified
and the Cq value was used. The 2−ΔΔCq method (21) was applied to detect the relative
expression of each gene.
 | Table IPrimer sequences for reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I
Primer sequences for reverse
transcription-quantitative polymerase chain reaction analysis.
| Gene | Sequence
(5′-3′) |
|---|
| PRMT1 | F:
AACCCTCACGTACCGCAACTCC |
| R:
CAGCCACTTGTCACGAGCGT |
| DDAH2 | F:
GCAACGACTAGGTCTGCAGCTTC |
| R:
GGTACCGTAGAGACAGCGAAGTC |
| DDAH1 | F:
AGCCGCAGGAAGGAGGTTGACATGAT |
| R:
GGTACTCTTCTGGGGTTGGGTGCA |
| Collagen I | F:
TTCACCTACTGCACGCTTGT |
| R:
TTGGGATGGAGGGAGTTTAC |
| MMP2 | F:
GATACCCTCAAGAAGATGCAGAAGT |
| R:
ATCTTGGCTTCCGCATGGT |
| TIMP2 | F:
GGCAAGATGCACATTACC |
| R:
AACTTGGCATTGTGGAAGG |
| GAPDH | F:
GTCTTCACTACCATGGAGAAG |
| R:
TCATGGATGACCTTGGCCAG |
Western blot analysis
A total of 20 mg myocardial tissues from the rats in
each group were supplemented with pre-cooled 2% sodium dodecyl
sulfate at ratio of 1:20 (m:V), homogenized on ice for 10 sec in an
electric homogenizer, and boiled at 100°C for 5 min. Following
cooling, the tissues were centrifuged at 12,000 × g for 4 min at
4°, and the supernatant was collected as tissue protein samples. In
each group, cells at the exponential growth phase were harvested
following 72 h of culture, and total protein was extracted using
the NucBusterä Protein Extraction kit (Novagen, Madison, WI,
USA).
The concentration of protein samples was determined
using a bicinchoninic acid kit (Pierce; Thermo Fisher Scientific,
Inc.) with 30 µg protein sample in each protein lane
adjusted by deionized water. Subsequently, the 10% SDS separation
gel and spacer gel were prepared, and the samples were mixed with
sample buffer prior to being boiled at 100°C for 5 min. After
cooling to room temperature, each sample was subjected to
electrophoresis by adding micro sampler into lanes. Subsequently,
the protein on the gel was transferred onto a polyvinylidene
fluoride membrane, which was then blocked with 5% skim milk
overnight at 4°C. The membrane was incubated with the following
diluted primary antibodies at 4°C overnight: Rabbit anti-rat
protein arginine N-methyltransferase 1 (PRMT1; 1:1,000; cat. no.
ab73246), rabbit anti-rat DDAH2 (1:500; cat. no. ab1383), rabbit
anti-rat DDAH1 (1:500; cat. no ab2231), rabbit anti-rat collagen I
(1:1,000; cat. no. ab34710), rabbit anti-rat matrix
metalloproteinase 2 (MMP2; 1:2,000, cat no. ab37150) and rabbit
anti-rat tissue inhibitor of metalloproteinase 2 (TIMP2; 1:500;
cat. no. ab180630). This was followed by three PBS washes (5 min
each time) at room temperature. All the above antibodies were
purchased form Abcam (Cambridge, MA, USA). Subsequently, the
membrane was incubated with secondary antibody, horseradish
peroxidase-labeled goat anti-rabbit IgG (1:3,000; cat. no. ab6721,
Abcam) at 37°C for 1 h, followed by three PBS washes (5 min each
time). The membrane was then immersed in electroluminescence
reaction liquid (Pierce; Thermo Fisher Scientific, Inc.) for 1 min
at room temperature and the liquid aspirated, followed by covering
with preservative film and observing under a chemiluminometer
(Shanghai Tianneng Co., Ltd., Shanghai, China) with GAPDH (1:
2,500; cat. no. ab9485’ Abcam) as an internal reference. Finally,
the ratio of the gray value of the target protein band/GAPDH
protein band was considered the relative protein expression.
Statistical analysis
SPSS 21.0 (IBM Corp., Armonk, NY, USA) was used for
statistical analysis. All experimental results were examined for
Gaussian distribution and homogeneity of variance. Continuous
variables are expressed as the mean ± standard deviation in the
case of Gaussian distribution and as median (interquartile range)
in the case of variables with non-normal distribution. Comparison
of measurement data among multiple groups was performed using
one-way analysis of variance, and post-hoc testing was conducted by
Tukey’s test. In addition, repeated-measures analysis of variance
was used to compare data at different time points. P<0.05 was
considered to indicate a statistically significant difference.
Results
DDAH2 improves cardiac function in
diabetic rats
The cardiac function was determined to examine the
effect of DDAH2 on cardiac function in diabetic rats with
myocardial fibrosis (Fig. 1A-D).
Compared with the diabetic rats of the model group, a significant
reduction in LVEDP was found in the diabetic rats of the DDAH2
group, whereas the levels of LVSP and LV ± dp/dt were increased
(all P<0.05). By contrast, the level of LVEDP increased
significantly and notable decreases in the levels of LVSP and LV ±
dp/dt were observed in the diabetic rats of the shDDAH2 and L-NNA
groups (all P<0.05). No notable differences in LVEDP, LVSP or LV
± dp/dt were observed in the diabetic rats of the model, DDAH2-NC,
shDDAH2-NC and L-NNA + DDAH2 groups (P>0.05). Taken together,
DDAH2 exhibited a protective effect on the cardiac function of
diabetic rats with myocardial fibrosis.
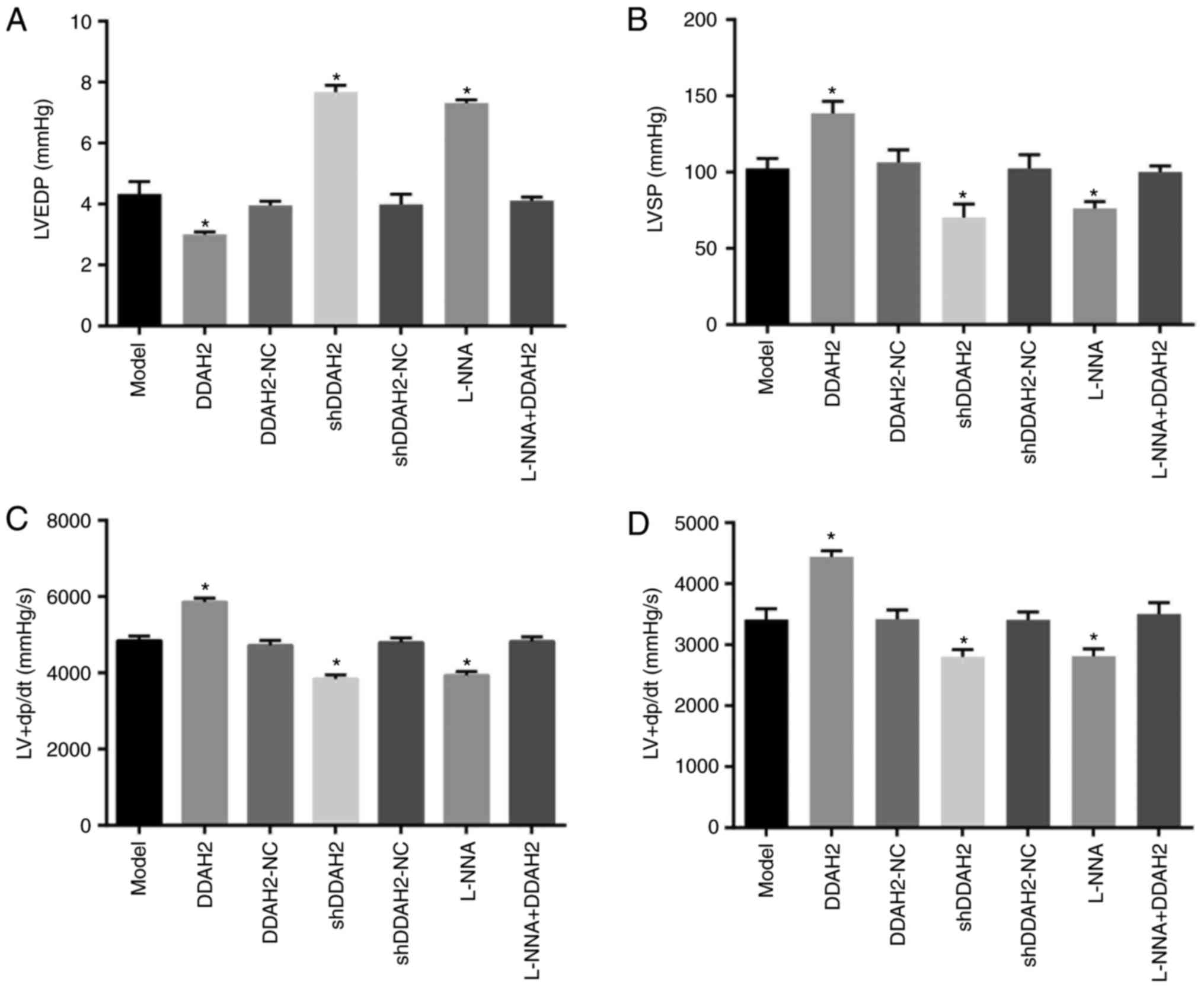 | Figure 1DDAH2 exerts a protective effect on
cardiac function in diabetic rats with myocardial fibrosis. (A)
LVEDP level in diabetic rats following 12 weeks of treatment with
lentivirus expressing DDAH2 or shDDAH2, and/or L-NNA. (B) LVSP
level in diabetic rats following 12 weeks of treatment with
lentivirus expressing DDAH2 or shDDAH2, and/or L-NNA. (C) LV +
dp/dt level in diabetic rats following 12 weeks of treatment with
lentivirus expressing DDAH2 or shDDAH2, and/or L-NNA. (D) LV-dp/dt
level in diabetic rats following 12 weeks of treatment with
lentivirus expressing DDAH2 or shDDAH2, and/or L-NNA. Data are
presented as the mean ± standard deviation and analyzed by the
one-way analysis of variance. All data are representative of three
independent experiments. n=5. *P<0.05, vs. model
group. DDAH2, dimethylarginine dimethylaminohydrolase 2; NC,
negative control; sh, short hairpin RNA; LVEDP, left ventricular
end-diastolic pressure, LVSP, left ventricular systolic pressure,
LV ± dp/dt, maximum rate of left ventricular pressure
rise/fall. |
DDAH2 decreases levels of biochemical
indicators in diabetic rats
The levels of biochemical indicators were detected
for determination of the role of DDAH2 in diabetic rats with
myocardial fibrosis (Fig. 2A-E).
From week 5 following treatment, the levels of BG, TC, TG, FBG and
FINS in the rats of the DDAH2 group were lower than those in the
diabetic rats of the model group (P<0.05), whereas the levels of
BG, TC, TG, FBG, and FINS were increased in the diabetic rats of
the shDDAH2 and L-NNA groups (P<0.05). No significant
differences in BG, TC, TG, FBG or FINS were found in the diabetic
rats of the model, DDAH2-NC, shDDAH2-NC and L-NNA + DDAH2 groups
(P>0.05). Taking these results into consideration, the positive
role of DDAH2 was identified as it alleviated myocardial fibrosis
in diabetic rats.
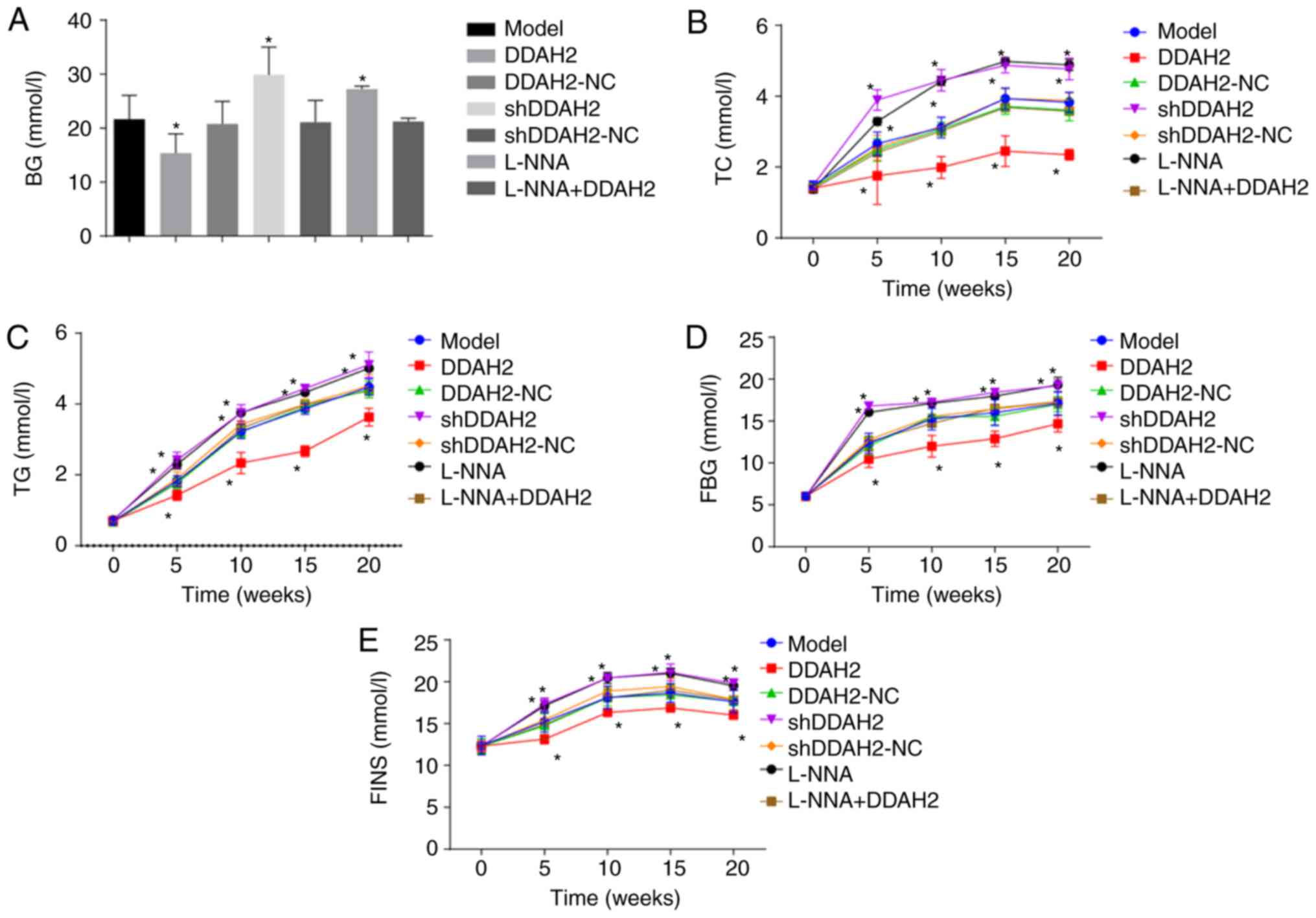 | Figure 2DDAH2 positively regulates blood
biochemical indicators in diabetic rats with myocardial fibrosis.
(A) BG level in diabetic rats following 5-20 weeks of treatment
with lentivirus expressing DDAH2 or shDDAH2, and/or L-NNA. (B) TC
level in diabetic rats following 5-20 weeks of treatment with
lentivirus expressing DDAH2 or shDDAH2, and/or L-NNA. (C) TG level
in diabetic rats following 5-20 weeks of treatment with lentivirus
expressing DDAH2 or shDDAH2, and/or L-NNA. (D) FBG level in
diabetic rats following 5-20 weeks of treatment with lentivirus
expressing DDAH2 or shDDAH2, and/or L-NNA. (E) FINS in diabetic
rats following 5-20 weeks of treatment with lentivirus expressing
DDAH2 or shDDAH2, and/or L-NNA. *P<0.05. vs. model
group. Data are presented as the mean ± standard deviation; data at
the same time point in different groups were analyzed by one-way
ANOVA; data at different time points were analyzed by repeated
measure ANOVA. All data are representative of three independent
experiments. n=5. DDAH2, dimethylarginine dimethylaminohydrolase 2;
NC, negative control; sh, sort hairpin RNA; BG, blood glucose; TC,
total cholesterol; TG, triglyceride; FBG, fasting blood glucose;
FINS, fasting insulin; ANOVA, analysis of variance. |
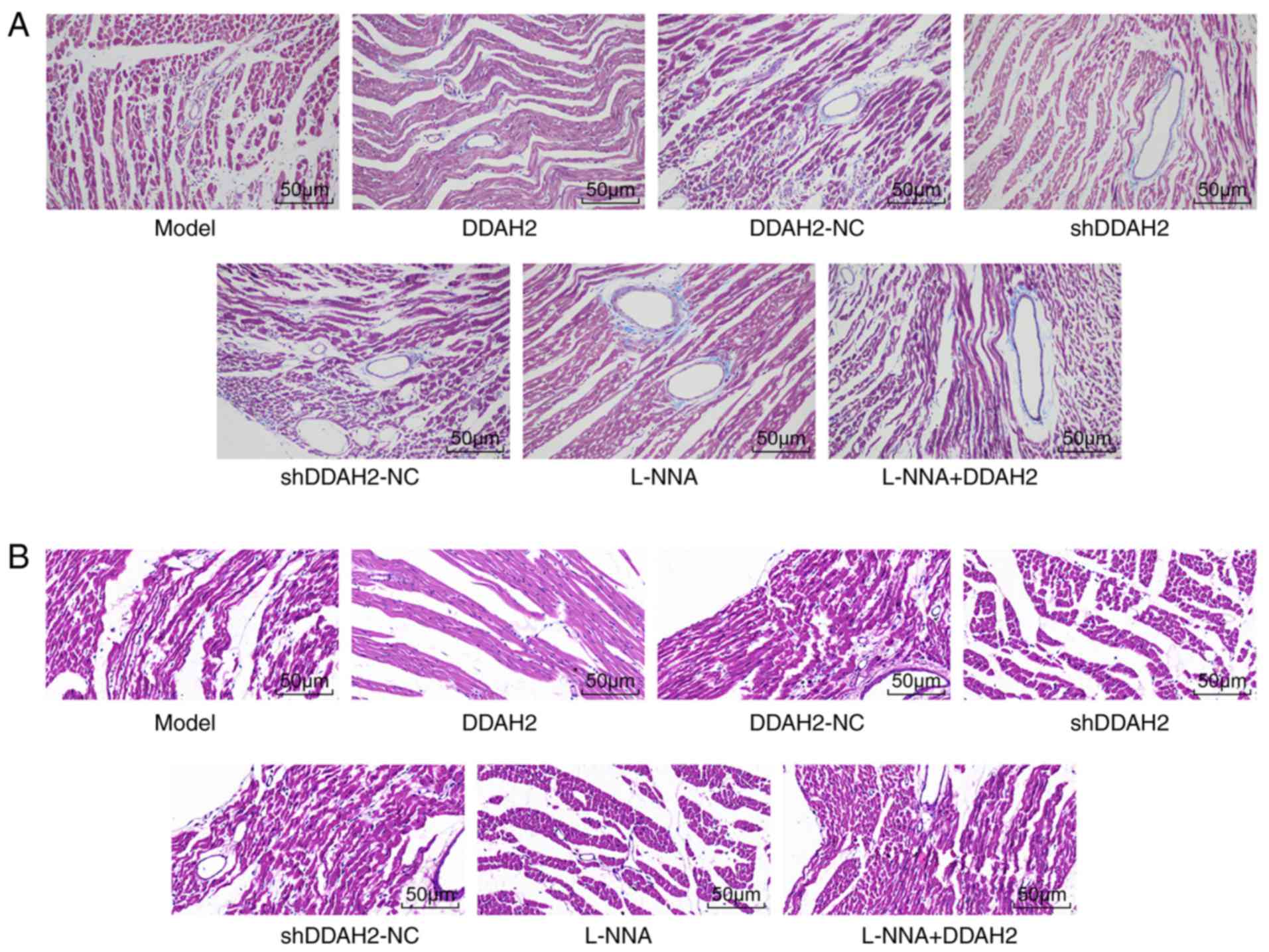
DDAH2 attenuates myocardial fibrosis in
diabetic rats
Masson staining and H&E staining were performed
to further determine whether DDAH2 was involved in myocardial
fibrosis in diabetic rats. Following Masson staining, collagen
fibers turned blue and normal cells were stained red. In the
diabetic rats of the model, DDAH2-NC, shDDAH2-NC, and L-NNA + DDAH2
groups, the myocardial cells were arranged loosely and disorderly
with hypertrophied and deformed cardiac muscle cells. In addition,
collagen fibers were distributed densely in the myocardial cells
and blood vessels. Compared with the diabetic rats of model group,
the myocardial tissues in the diabetic rats of the DDAH2 group were
arranged regularly with reduced swelling and collagen fibers,
whereas the arrangement of myocardial cells in the diabetic rats of
the shDDAH2 and L-NNA groups was disordered with aggravated
swelling and increased collagen fibers (Fig. 3A).
Following H&E staining, the myocardial cells in
the diabetic rats of the model, DDAH2-NC, shDDAH2-NC, and L-NNA +
DDAH2 groups were hypertrophied with obvious myocardial fiber
damage and rupture. Less myocardial hypertrophy and necrosis, and
generally complete cell morphology were observed in the DDAH2 group
when compared with the model group. The diabetic rats in the
shDDAH2 and L-NNA groups showed more hypertrophic and distorted
myocardial cells, with disordered arrangement and an increase of
intercellular space compared with the model group (Fig. 3B). These results showed DDAH2
attenuated myocardial fibrosis by reducing swelling and decreasing
collagen fibers in diabetic rats.
DDAH2 inhibits migration of myocardial
cells under high glucose conditions
Transwell, RT-qPCR and western blot assays were
performed to evaluate the function of DDAH2 in the migration of
H9C2 myocardial cells under high glucose conditions. The results of
the Transwell assay (Fig. 4A)
showed that the migration ability of the H9C2 cells in the DDAH2
group was significantly decreased compared with that in the model
group (P<0.05), whereas the migration ability of H9C2 cells in
the shDDAH2 and L-NNA groups was significantly enhanced
(P<0.05). No significant differences in H9C2 cell migration were
observed in the DDAH2-NC, shDDAH2-NC and L-NNA + DDAH2 groups
(P>0.05).
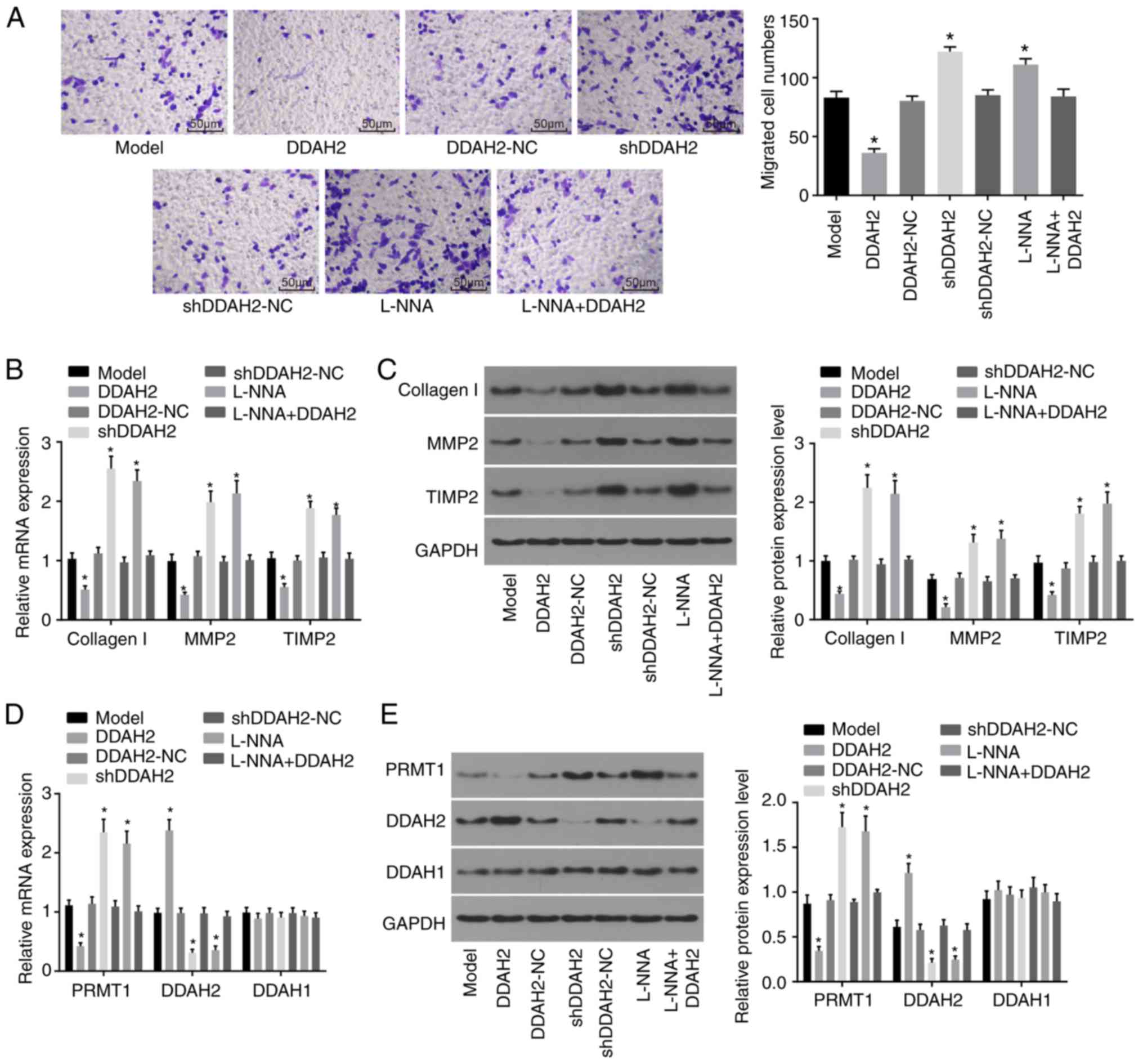 | Figure 4DDAH2 inhibits myocardial cell
migration. (A) Migration ability of H9C2 myocardial cells under
high glucose conditions following 48 h of treatment with lentivirus
expressing DDAH2 or shDDAH2, and/or L-NNA (magnification, ×200).
(B) mRNA levels of collagen I, MMP2 and TIMP in H9C2 myocardial
cells under high glucose conditions following 48 h of treatment
with lentivirus expressing DDAH2 or shDDAH2, and/or L-NNA. (C)
Protein levels of collagen I, MMP2 and TIMP in H9C2 myocardial
cells under high glucose conditions following 48 h of treatment
with lentivirus expressing DDAH2 or shDDAH2, and/or L-NNA. (D) mRNA
expression of PRMT1, DDAH2 and DDAH1 in H9C2 myocardial cells under
high glucose condition following 48 h of treatment with lentivirus
expressing DDAH2 or shDDAH2, and/or L-NNA (E) Protein expression of
PRMT1, DDAH2 and DDAH1 in H9C2 myocardial cells under high glucose
conditions following 48 h of treatment with lentivirus expressing
DDAH2 or shDDAH2, and/or L-NNA. Data are presented as the mean ±
standard deviation and analyzed by one-way analysis of variance.
All data are representative of three independent experiments.
*P<0.05, vs. model group. DDAH, dimethylarginine
dimethylaminohydrolase; PRMT1, protein arginine N-methyltransferase
1; NC, negative control; sh, short hairpin RNA; MMP2, matrix
metalloproteinase 2; TIMP2, tissue inhibitor of metalloproteinase
2; GAPDH, glyceraldehyde-3-phosphate dehydrogenase. |
In addition, the results of the RT-qPCR and western
blot analyses (Fig. 4B and C)
showed that, compared with the H9C2 cells of the model group, the
mRNA and protein levels of collagen I, MMP2 and TIMP2 in the H9C2
cells of the DDAH2 group were significantly decreased (all
P<0.05). The mRNA and protein levels of collagen I, MMP2 and
TIMP2 in the H9C2 cells of the shDDAH2 and L-NNA groups were
significantly increased, compared with those in the H9C2 cells of
the model group (all P<0.05). No noticeable differences
in the mRNA and protein levels of collagen I, MMP2, and TIMP2 were
found in the model, DDAH2-NC, shDDAH2-NC, and L-NNA + DDAH2 groups
(P>0.05). These result indicated that DDAH2 negatively affected
myocardial cell migration.
In addition, the results (Fig. 4D and E) showed that, compared with
the H9C2 cells in the model group, the H9C2 cells in the DDAH2
group showed significantly decreased mRNA and protein expression
levels of PRMT1 but increased mRNA and protein expression levels of
DDAH2 (all P<0.05). The H9C2 cells in the shDDAH2 and L-NNA
groups exhibited increased mRNA and protein expression levels of
PRMT1 but decreased mRNA and protein expression levels of DDAH2
compared with those in the model group (all P<0.05). No
significant differences in the mRNA and protein expression levels
of PRMT1 and DDAH2 were observed among the H9C2 cells in the model,
DDAH2-NC, shDDAH2-NC and L-NNA+DDAH2 groups (P>0.05). No
significant differences in the mRNA and protein expression levels
of DDAH1 were found among the groups (P>0.05). Taken together,
these results suggested that DDAH2 may induce the activation of
DDAH in myocardial cells (H9C2) under high glucose conditions.
DDAH2 activates the DDAH/ADMA/NOS/NO
pathway in diabetic rats with myocardial fibrosis
Finally, the regulatory association between DDAH2
and the DDAH/ADMA/NOS/NO pathway was investigated in diabetic rats
with myocardial fibrosis. Compared with the rats in the model
group, the rats in the DDAH2 group exhibited lower serum ADMA
content but higher serum DDAH activity, NOS activity and NO content
(all P<0.05); an opposite trend was observed in the shDDAH2 and
L-NNA groups (all P<0.05). No notable differences in serum ADMA
content, DDAH activity, NOS activity or NO content were observed
among the model, DDAH2-NC, shDDAH2-NC, and L-NNA + DDAH2 groups
(P>0.05) (Fig. 5A-D).
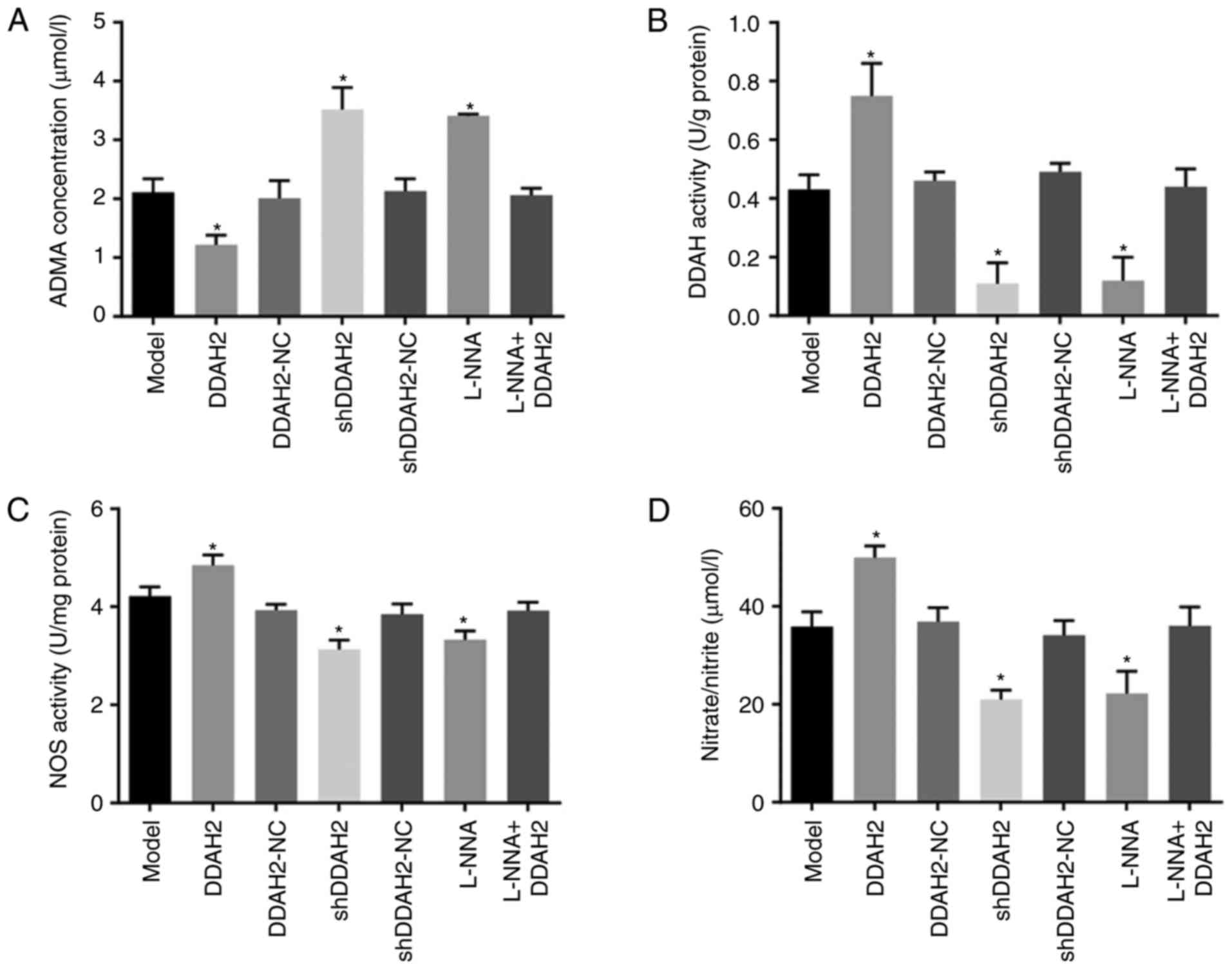 | Figure 5DDAH2 activates the DDAH/ADMA/NOS/NO
pathway in diabetic rats with myocardial fibrosis. (A) Serum ADMA
content in diabetic rats following 20 weeks of treatment with
lentivirus expressing DDAH2 or shDDAH2, and/or L-NNA. (B) DDAH
activity in myocardial tissues of diabetic rats following 20 weeks
of treatment with lentivirus expressing DDAH2 or shDDAH2, and/or
L-NNA. (C) NOS activity in myocardial tissues of diabetic rats
following 20 weeks of treatment with lentivirus expressing DDAH2 or
shDDAH2, and/or L-NNA. (D) NO content in myocardial tissues of
diabetic rats following 20 weeks of treatment with lentivirus
expressing DDAH2 or shDDAH2, and/or L-NNA. (E) mRNA and (F) protein
levels of PRMT1, DDAH2 and DDAH1 in myocardial tissues of rats
following 20 weeks of treatment with lentivirus expressing DDAH2 or
shDDAH2, and/or L-NNA. Data are presented as the mean ± standard
deviation and were analyzed by one-way analysis of variance.
*P<0.05, vs. model group All data are representative
of three independent experiments. DDAH, dimethylarginine
dimethylaminohydrolase; ADMA, asymmetric NG,
NGdimethyl-L-arginine; PRMT1, protein arginine
N-methyltransferase 1; NC, negative control; shRNA, short hairpin
RNA; ADMA, asymmetric dimethylarginine; NOS, nitric oxide synthase;
NO, nitric oxide; GAPDH, glyceraldehyde-3-phosphate
dehydrogenase. |
The results of the RT-qPCR and western blot analyses
(Fig. 5E and F) showed that,
compared with the diabetic rats in the model group, the mRNA and
protein levels of PRMT1 were decreased whereas those of DDAH2 were
increased in the myocardial tissues of diabetic rats in the DDAH2
group (all P<0.05). By contrast, the opposite trend was found in
the shDDAH2 and L-NNA groups (all P<0.05). No significant
changes in the mRNA and protein levels of PRMT1 or DDAH2 were shown
among the model, DDAH2-NC, shDDAH2-NC and L-NNA + DDAH2 groups
(P>0.05). No significant differences in the mRNA and protein
levels of DDAH1 were observed among any groups (P>0.05). The
above findings revealed that DDAH2 activated the DDAH/ADMA/NOS/NO
pathway in diabetic rats with myocardial fibrosis.
Discussion
DCM features molecular, metabolic and structural
heart changes (22), and is
considered to be the most common cause of high morbidity and
mortality rates in diabetic patients (23). In an effort to identify a novel
treatment method for DCM, the present study examined the mechanism
underlying the effect of DDAH2 in DCM by regulating the
DDAH/ADMA/NOS/NO pathway.
The first finding of the present study was that the
DCM rats treated with DDAH2 exhibited reduced LVEDP, BG, TC, TG,
FBG, FINS levels but increased LVSP and LV ± dp/dt levels in
myocardial tissues, indicating that DDAH2 improved cardiac function
in diabetic rats. The overexpression of DDAH has been reported to
delay the development of graft coronary artery disease (24). DDAH is capable of altering key
fibrotic signaling cascades and is considered a promising treatment
for patients with idiopathic pulmonary fibrosis (25,26). The sequence variation in DDAH1 and
DDAH2 genes is reported to be implicated in type 2 diabetes
(27-29). In line with the findings of the
present study, a previous study indicated that the upregulation of
DDAH2 improved endothelial dysfunction and can be regarded as a
positive factor for diabetes treatment (30). The above findings indicate that
DDAH2 is a positive factor for cardiac function in diabetes.
In addition, the present study showed that DDAH2
exerted inhibitory effects on myocardial cell migration under high
glucose conditions and myocardial fibrosis in diabetic rats. This
result is in accordance with higher expression levels of collagen
I, MMP2 and TIMP2. Collagen is regarded as the major component of
the extracellular matrix and mainly appears in the mammalian body
(31). Former studies have
reported that excessive synthesis of collagen can lead to
myocardial fibrosis (32,33). It was suggested in a previous
study that MMP positively correlates with type I collagen (34). DDAH is a vital enzyme which may
have a positive correlation with MMP-2 and collagen (35-37). MMPs constitute a group of
zinc-binding proteolytic enzymes which commonly remodel the
extracellular matrix and are involved in the inflammatory response.
MMP-2, as one member of the MMP family, is closely associated with
diabetes (38,39). Furthermore, it has been revealed
that MMP2 and MMP9 are implicated in the migration of human
cardiomyocyte progenitor cells (40). TIMPs are endogenous inhibitors of
MMPs and are involved in the common biological behaviors of cells
(41). In addition, TIMP2 has
been found to be responsible for enhancing the migration of
epidermal keratinocytes and dermal fibroblasts (42), and myocardial fibrosis can be
suppressed by the rosuvastatin-induced upregulation of DDAH2
(43). These results reveal that
the migration of myocardial cells and myocardial fibrosis can be
disrupted by DDAH2.
Of note, the present study found that DDAH2
positively regulates the DDAH/ADMA/NOS/NO pathway in myocardial
cells under high glucose conditions and in diabetic rats with
myocardial fibrosis, which resulted in a decrease in PRMT1 and ADMA
content, together with increases in NOS activity and NO content. In
detail, silencing or the overexpression of DDAH2 in diabetic rats
showed effects on the expression of key factors associated with the
DDAH/ADMA/NOS/NO pathway, aggravating or alleviating the myocardial
fibrosis of diabetic rats. ADMA can be catabolized into citrulline
and methylamines in an active manner under physiological
conditions, and this effect is modulated by the alteration of DDAH
(44). ADMA, which appears during
the hydrolysis of methylated proteins, is produced by consistent
protein turnover and catalyzed by S-adenosylmethionine PRMT-1
(45). PRMT-1 is downregulated by
DDAH, and ADMA is negatively associated with DDAH (46,47). In addition, DDAH is positively
correlated with NOS and NO through an inverse association with ADM
(48). Similar to the findings of
the present study, several studies have suggested that ADMA is
negatively associated with DDAH1, whereas NOS and NO are positively
associated with DDAH1. In these regulatory processes, ADMA is a
primary endogenous factor inhibiting the production of NOS and NO
(47,49,50). Previously, it has been indicated
that a lower expression of ADMA is essential for reducing the
cardiovascular risk in diabetic patients (51). The suppression of endothelial NOS
has been revealed to enhance myocardial fibrosis (52). In addition, the expression of
DDAH2 is decreased and that of ADMA is increased under high glucose
conditions, and the high glucose-induced cell senescence and
upregulated expression of ADMA may be promoted in the absence of
DDAH2 (53). Taken together, it
is possible to suggest a model in which DDAH2 affects
diabetes-induced cardiac damage and myocardial fibrosis via
modulation of the DDAH/ADMA/NOS/NO pathway.
In conclusion, the results of the present study
suggest that the treatment of diabetic model rats with DDAH2 is
able to improve impaired cardiac function and myocardial fibrosis,
thus delaying the progression of DCM and inhibiting subsequent
undesirable consequences. The DDAH/ADMA/NOS/NO pathway was
stimulated by DDAH2 treatment, therefore, DDAH2 is considered to be
a novel therapeutic approach specific to DCM. However, additional
in vitro investigations are required to definitively
identify the molecular mechanisms and target genes of DDAH2.
Funding
The present study was supported by the Health
Science and Technology Project of Yunnan Province (grant no.
2014NS243), the Applied Yunnan Local Colleges Basic Research Joint
Special Key Projects in 2017 (grant no. 2017FH001-009) and the
Basic Research Program of Science and Technology Plan of Yunnan
Province [grant no. 2017FF117(-064)].
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors’ contributions
ZDZ JMY XMF and XCW contributed to conception and
design of the study. JYY, XRW and PH contributed to acquisition,
analysis, and interpretation of the data and was the major
contributor in writing the manuscript. YQL WX and JLD each
contributed to acquisition of the data, WYL, HF, ZHX and XZ
collected the clinical specimen and carried out the animal
experiments. ZDZ, JMY, XMF, XCW and XZ contributed to revision of
the manuscript. JYY, XRW, PH, YQL and WX contributed to analysis
and interpretation of the data and to revision of the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by The First People’s
Hospital of Yunnan Province, Affiliated Hospital of Kunming Science
and Technology University. All experimental procedures and
protocols were in line with the recommendations of the Guide for
the Care and Use of Laboratory Animals of the American National
Institutes of Health.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Nouhjah S, Shahbazian H, Amoori N,
Jahanfar S, Shahbazian N, Jahanshahi A and Cheraghian B: Postpartum
screening practices, progression to abnormal glucose tolerance and
its related risk factors in Asian women with a known history of
gestational diabetes: A systematic review and meta-analysis.
Diabetes Metab Syndr. 2(Suppl 11): S703–S712. 2017. View Article : Google Scholar
|
|
2
|
Han K, Yao J, Yin X, Zhao M and Sun Q:
Review on the prevalence of diabetes and risk factors and situation
of disease management in floating population in China. Glob Health
Res Policy. 2:332017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhu WW, Yang HX, Wang C, Su RN, Feng H and
Kapur A: High prevalence of gestational diabetes mellitus in
beijing: Effect of maternal birth weight and other risk factors.
Chin Med J (Engl). 130:1019–1025. 2017. View Article : Google Scholar
|
|
4
|
Xu Y, Wang L, He J, Bi Y, Li M, Wang T,
Wang L, Jiang Y, Dai M, Lu J, et al: Prevalence and control of
diabetes in Chinese adults. JAMA. 310:948–959. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu F, Song R, Feng Y, Guo J, Chen Y,
Zhang Y, Chen T, Wang Y, Huang Y, Li CY, et al: Upregulation of
MG53 induces diabetic cardiomyopathy through transcriptional
activation of peroxisome proliferation-activated receptor α.
Circulation. 131:795–804. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Berclaz PY, Gao H, Tobian JA, Swanson DL,
Webb DM, Crapo RO and Jensen RL: The impact of diabetes and age on
pulmonary function: Data from the National Health and Nutrition
Examination Survey. Diabetes Res Clin Pract. 83:e1-e32009.
View Article : Google Scholar
|
|
7
|
Jellis C, Martin J, Narula J and Marwick
TH: Assessment of nonischemic myocardial fibrosis. J Am Coll
Cardiol. 56:89–97. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ares-Carrasco S, Picatoste B,
Benito-Martin A, Zubiri I, Sanz AB, Sanchez-Nino MD, Ortiz A, Egido
J, Tunon J and Lorenzo O: Myocardial fibrosis and apoptosis, but
not inflammation, are present in long-term experimental diabetes.
Am J Physiol Heart Circ Physiol. 297:H2109–H2119. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hollekim-Strand SM, Bjorgaas MR,
Albrektsen G, Tjonna AE, Wisloff U and Ingul CB: High-intensity
interval exercise effectively improves cardiac function in patients
with type 2 diabetes mellitus and diastolic dysfunction: A
randomized controlled trial. J Am Coll Cardiol. 64:1758–1760. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tappia PS, Thliveris J, Xu YJ,
Aroutiounova N and Dhalla NS: Effects of amino acid supplementation
on myocardial cell damage and cardiac function in diabetes. Exp
Clin Cardiol. 16:e17-e222011.
|
|
11
|
Karamitsos TD: Vascular and myocardial
fibrosis in diabetes mellitus. Cardiology. 114:105–106. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lai YL, Aoyama S, Ohata M, Otsuka N,
Shiokawa H, Tomono S, Fujiwara Y, Kanazawa H, Miyoshi N and Ohshima
H: Dysregulation of
dimethylargininedimethylaminohydrolase/asymmetric dimethylarginine
pathway in rat type II diabetic nephropathy. J Clin Biochem Nutr.
51:143–149. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sakurada M, Shichiri M, Imamura M, Azuma H
and Hirata Y: Nitric oxide upregulates dimethylarginine
dimethylamino-hydrolase-2 via cyclic GMP induction in endothelial
cells. Hypertension. 52:903–909. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Torondel B, Nandi M, Kelly P,
Wojciak-Stothard B, Fleming I and Leiper J: Adenoviral-mediated
overexpression of DDAH improves vascular tone regulation. Vasc Med.
15:205–213. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zheng J, Wang K, Jin P, Dong C, Yuan Q, Li
Y and Yang Z: The association of adipose-derived dimethylarginine
dimethylaminohydrolase-2 with insulin sensitivity in experimental
type 2 diabetes mellitus. Acta Biochim Biophys Sin (Shanghai).
45:641–648. 2013. View Article : Google Scholar
|
|
16
|
Thum T, Fraccarollo D, Schultheiss M,
Froese S, Galuppo P, Widder JD, Tsikas D, Ertl G and Bauersachs J:
Endothelial nitric oxide synthase uncoupling impairs endothelial
progenitor cell mobilization and function in diabetes. Diabetes.
56:666–674. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen D, Zhang KQ, Li B, Sun DQ, Zhang H
and Fu Q: Epigallocatechin-3-gallate ameliorates erectile function
in aged rats via regulation of PRMT1/DDAH/ADMA/NOS metabolism
pathway. Asian J Androl. 19:291–297. 2017. View Article : Google Scholar :
|
|
18
|
Wang JH, Chen D, Zhang KQ, Zhang H and Fu
Q: Effect of DDAH/ADMA/NOS regulation pathway on cavernae corporum
cavernosorum rat penis of different age. Andrologia. 48:262–267.
2016. View Article : Google Scholar
|
|
19
|
Yuan Q, Hu CP, Gong ZC, Bai YP, Liu SY, Li
YJ and Jiang JL: Accelerated onset of senescence of endothelial
progenitor cells in patients with type 2 diabetes mellitus: Role of
dimethylarginine dimethylaminohydrolase 2 and asymmetric
dimethylarginine. Biochem Biophys Res Commun. 458:869–876. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu CW, Lin Y, Lei YP, Wang L, He ZM and
Xiong Y: Pyrrolidine dithiocarbamate ameliorates endothelial
dysfunction in thoracic aorta of diabetic rats by preserving
vascular DDAH activity. PLoS One. 12:e01799082017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ayuk SM, Abrahamse H and Houreld NN: The
role of photobiomodulation on gene expression of cell adhesion
molecules in diabetic wounded fibroblasts in vitro. J Photochem
Photobiol B. 161:368–374. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mizamtsidi M, Paschou SA, Grapsa J and
Vryonidou A: Diabetic cardiomyopathy: A clinical entity or a
cluster of molecular heart changes. Eur J Clin Invest. 46:947–953.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tang SG, Liu XY, Ye JM, Hu TT, Yang YY,
Han T and Tan W: Isosteviol ameliorates diabetic cardiomyopathy in
rats by inhibiting ERK and NF-κB signaling pathways. J Endocrinol.
238:47–60. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tanaka M, Sydow K, Gunawan F, Jacobi J,
Tsao PS, Robbins RC and Cooke JP: Dimethylarginine
dimethylaminohydrolase overexpression suppresses graft coronary
artery disease. Circulation. 112:1549–1556. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Janssen W, Pullamsetti SS, Cooke J,
Weissmann N, Guenther A and Schermuly RT: The role of
dimethylarginine dimethylaminohydrolase (DDAH) in pulmonary
fibrosis. J Pathol. 229:242–249. 2013. View Article : Google Scholar
|
|
26
|
Pullamsetti SS, Savai R, Dumitrascu R,
Dahal BK, Wilhelm J, Konigshoff M, Zakrzewicz D, Ghofrani HA,
Weissmann N, Eickelberg O, et al: The role of dimethylarginine
dimethylaminohydrolase in idiopathic pulmonary fibrosis. Sci Transl
Med. 3:87ra532011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Abhary S, Burdon KP, Kuot A, Javadiyan S,
Whiting MJ, Kasmeridis N, Petrovsky N and Craig JE: Sequence
variation in DDAH1 and DDAH2 genes is strongly and additively
associated with serum ADMA concentrations in individuals with type
2 diabetes. PLoS One. 5:e94622010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Seo HA, Kim SW, Jeon EJ, Jeong JY, Moon
SS, Lee WK, Kim JG, Lee IK and Park KG: Association of the DDAH2
gene polymorphism with type 2 diabetes and hypertension. Diabetes
Res Clin Pract. 98:125–131. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lu TM, Lin SJ, Lin MW, Hsu CP and Chung
MY: The association of dimethylarginine dimethylaminohydrolase 1
gene polymorphism with type 2 diabetes: A cohort study. Cardiovasc
Diabetol. 10:162011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu CW, Guo Z, Feng M, Wu ZZ, He ZM and
Xiong Y: Ex vivo gene transferring of human dimethylarginine
dimethylaminohy-drolase-2 improved endothelial dysfunction in
diabetic rat aortas and high glucose-treated endothelial cells.
Atherosclerosis. 209:66–73. 2010. View Article : Google Scholar
|
|
31
|
Rajan N, Habermehl J, Coté MF, Doillon CJ
and Mantovani D: Preparation of ready-to-use, storable and
reconstituted type I collagen from rat tail tendon for tissue
engineering applications. Nat Protoc. 1:2753–2758. 2006. View Article : Google Scholar
|
|
32
|
Challa AA, Vukmirovic M, Blackmon J and
Stefanovic B: Withaferin-A reduces type I collagen expression in
vitro and inhibits development of myocardial fibrosis in vivo. PLoS
One. 7:e429892012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Querejeta R, López B, González A, Sánchez
E, Larman M, Martínez Ubago JL and Díez J: Increased collagen type
I synthesis in patients with heart failure of hypertensive origin:
Relation to myocardial fibrosis. Circulation. 110:1263–1268. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yucel T, Mutnal A, Fay K, Fligiel SE, Wang
T, Johnson T, Baker SR and Varani J: Matrix metalloproteinase
expression in basal cell carcinoma: Relationship between enzyme
profile and collagen fragmentation pattern. Exp Mol Pathol.
79:151–160. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hu X, Atzler D, Xu X, Zhang P, Guo H, Lu
Z, Fassett J, Schwedhelm E, Böger RH, Bache RJ and Chen Y:
Dimethylarginine dimethylaminohydrolase-1 is the critical enzyme
for degrading the cardiovascular risk factor asymmetrical
dimethylarginine. Arterioscler Thromb Vasc Biol. 31:1540–1546.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ko CH, Shen SC, Lee TJ and Chen YC:
Myricetin inhibits matrix metalloproteinase 2 protein expression
and enzyme activity in colorectal carcinoma cells. Mol Cancer Ther.
4:281–290. 2005.PubMed/NCBI
|
|
37
|
Hasegawa K, Wakino S, Tanaka T, Kimoto M,
Tatematsu S, Kanda T, Yoshioka K, Homma K, Sugano N, Kurabayashi M,
et al: Dimethylarginine dimethylaminohydrolase 2 increases vascular
endothelial growth factor expression through Sp1 transcription
factor in endothelial cells. Arterioscler Thromb Vasc Biol.
26:1488–1494. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lewandowski KC, Banach E, Bieńkiewicz M
and Lewiński A: Matrix metalloproteinases in type 2 diabetes and
non-diabetic controls: Effects of short-term and chronic
hyperglycaemia. Arch Med Sci. 7:294–303. 2011. View Article : Google Scholar
|
|
39
|
Van Linthout S, Seeland U, Riad A,
Eckhardt O, Hohl M, Dhayat N, Richter U, Fischer JW, Böhm M,
Pauschinger M, et al: Reduced MMP-2 activity contributes to cardiac
fibrosis in experimental diabetic cardiomyopathy. Basic Res
Cardiol. 103:319–327. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu J, van Mil A, Aguor EN, Siddiqi S,
Vrijsen K, Jaksani S, Metz C, Zhao J, Strijkers GJ, Doevendans PA
and Sluijter JP: MiR-155 inhibits cell migration of human
cardiomyocyte progenitor cells (hCMPCs) via targeting of MMP-16. J
Cell Mol Med. 16:2379–2386. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Peterson NB, Beeghly-Fadiel A, Gao YT,
Long J, Cai Q, Shu XO and Zheng W: Polymorphisms in tissue
inhibitors of metalloproteinases-2 and -3 and breast cancer
susceptibility and survival. Int J Cancer. 125:844–850. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Miyoshi H, Kanekura T, Aoki T and Kanzaki
T: Beneficial effects of tissue inhibitor of metalloproteinases-2
(TIMP-2) on chronic dermatitis. J Dermatol. 32:346–353. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhou R, Ma P, Xiong A, Xu Y, Wang Y and Xu
Q: Protective effects of low-dose rosuvastatin on
isoproterenol-induced chronic heart failure in rats by regulation
of DDAH-ADMA-NO pathway. Cardiovasc Ther. 35:2017. View Article : Google Scholar
|
|
44
|
Frombaum M, Therond P, Djelidi R, Beaudeux
JL, Bonnefont-Rousselot D and Borderie D: Piceatannol is more
effective than resveratrol in restoring endothelial cell
dimethylarginine dimethylaminohydrolase expression and activity
after high-glucose oxidative stress. Free Radic Res. 45:293–302.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Novella S, Laguna-Fernández A,
Lázaro-Franco M, Sobrino A, Bueno-Betí C, Tarín JJ, Monsalve E,
Sanchís J and Hermenegildo C: Estradiol, acting through estrogen
receptor alpha, restores dimethylarginine dimethylaminohydrolase
activity and nitric oxide production in oxLDL-treated human
arterial endothelial cells. Mol Cell Endocrinol. 365:11–16. 2013.
View Article : Google Scholar
|
|
46
|
Pope AJ, Karuppiah K and Cardounel AJ:
Role of the PRMT-DDAH-ADMA axis in the regulation of endothelial
nitric oxide production. Pharmacol Res. 60:461–465. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jiang DJ, Jia SJ, Yan J, Zhou Z, Yuan Q
and Li YJ: Involvement of DDAH/ADMA/NOS pathway in nicotine-induced
endothelial dysfunction. Biochem Biophys Res Commun. 349:683–693.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Palm F, Onozato ML, Luo Z and Wilcox CS:
Dimethylarginine dimethylaminohydrolase (DDAH): Expression,
regulation, and function in the cardiovascular and renal systems.
Am J Physiol Heart Circ Physiol. 293:H3227–H3245. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Jiang DJ, Jia SJ, Dai Z and Li YJ:
Asymmetric dimethylarginine induces apoptosis via p38
MAPK/caspase-3-dependent signaling pathway in endothelial cells. J
Mol Cell Cardiol. 40:529–539. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Greco R, Ferrigno A, Demartini C, Zanaboni
A, Mangione AS, Blandini F, Nappi G, Vairetti M and Tassorelli C:
Evaluation of ADMA-DDAH-NOS axis in specific brain areas following
nitroglycerin administration: Study in an animal model of migraine.
J Headache Pain. 16:5602015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
King DE, Player M and Everett CJ: The
impact of pioglitazone on ADMA and oxidative stress markers in
patients with type 2 diabetes. Prim Care Diabetes. 6:157–161. 2012.
View Article : Google Scholar
|
|
52
|
Kazakov A, Hall R, Jagoda P, Bachelier K,
Müller-Best P, Semenov A, Lammert F, Böhm M and Laufs U: Inhibition
of endothelial nitric oxide synthase induces and enhances
myocardial fibrosis. Cardiovasc Res. 100:211–221. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yuan Q, Peng J, Liu SY, Wang CJ, Xiang DX,
Xiong XM, Hu CP and Li YJ: Inhibitory effect of resveratrol
derivative BTM-0512 on high glucose-induced cell senescence
involves dimethylaminohydrolase/asymmetric dimethylarginine
pathway. Clin Exp Pharmacol Physiol. 37:630–635. 2010. View Article : Google Scholar : PubMed/NCBI
|