Introduction
Gastric cancer is one of the most common
malignancies, with a substantial impact on global health (1-3).
During its early stages, it is either asymptomatic or presents with
nonspecific symptoms. Therefore, gastric cancer often reaches an
advanced stage prior to diagnosis and poses a major clinical
challenge with poor patient outcomes (4-6).
In addition, there are limited efficacious therapeutic strategies
for treating advanced gastric cancer (7,8),
and the molecular mechanisms underlying gastric cancer progression
remain unclear. Thus, elucidating the key molecular mechanisms will
aid in the development of novel therapeutic targets for this
disease.
Disintegrin and metalloproteinase domain-containing
proteins (ADAMs) are a family of multidomain transmembrane
glycoproteins, which are widely implicated in cell adhesion,
cell-cell signaling and cell migration (9,10).
The diverse roles of ADAMs in the pathological processes of various
human cancers have gained increasing attention. In gastric cancer,
ADAM33 affects cell migration and proliferation by regulating the
secretion of interleukin-18 (11). ADAM9, 12, and 15 are expressed at
higher levels in gastric cancer cells, compared with non-neoplastic
foveolar epithelial cells, and have been implicated in the
malignant growth of gastric cancer (12). ADAM10 may serve as a useful
prognostic marker for predicting gastric cancer progression, as
ADAM10 expression correlates with lymph node and distant metastases
(13). Although efforts have been
made, a higher priority must be given to further investigate key
members of the ADAM family that are associated with tumor
metastasis and are of prognostic significance for patients with
gastric cancer.
ADAM17 (also known as TNF-α-converting-enzyme) has
been identified to function as a signaling scissor in the tumor
microenvironment (14) and thus
contributes to tumorigenesis and tumor progression (15,16). The dysregulation of ADAM17
expression and its crucial role in the pathology of various cancers
has been widely revealed. For instance, increased ADAM17 expression
is associated with aggressive progression of non-small cell lung
cancer (17). ADAM17 silencing
suppresses cancer cell growth autonomy and inhibits tissue invasion
(18). Further, ADAM17 promotes
breast cancer tumorigenesis and progression by regulating cell
proliferation, angiogenesis, invasion and apoptosis (19). Notably, ADAM17 was found to be
upregulated in gastric cancer, and this increased expression
contributed to tumor progression and resulted in poor prognosis;
ADAM17 may therefore be an independent factor to predict tumor
prognosis (20,21). In addition, microRNA (miR)-338-3p
has been shown to suppress the proliferation, migration and
invasion of gastric cancer cells through downregulation of ADAM17
(22). ADAM17 expression is
upregulated by forkhead box protein M1 and consequently promotes
the proliferation and tumor growth of gastric cancer cells
(23). Additionally, ADAM17
promotes epithelial-mesenchymal transition (EMT) in gastric cancer
cells through the transforming growth factor (TGF)-β/mothers
against decapentaplegic homolog (Smad) signaling pathway (24). Nevertheless, the role and
potential regulatory mechanism of ADAM17 in gastric cancer
progression has not been fully elucidated.
The objective of the present study was to identify
the role of ADAM17 in regulating gastric cancer metastasis, to
investigate its mechanism of action, and to evaluate its clinical
significance. In the present study, key members of the ADAM protein
family associated with lymph node metastasis in gastric cancers
were identified, and the correlation between survival times of
patients and their clinicopathological features was investigated.
Additionally, the expression of ADAM17 in gastric cancer cells with
different metastatic potential was determined and the effects of
ADAM17 dysregulation on cell viability, migration, and invasion
were explored. Finally, key pathways correlated with ADAM17 were
identified using gene set enrichment analysis (GSEA) and confirmed
by western blot analysis.
Patients and methods
Patients
From January 2011 to December 2012, 193 patients
(150 males and 43 females) were enrolled at The First Hospital of
Jilin University with gastric cancer and positive lymph node
metastasis. Patients underwent gastrectomy with D2 lymph node
dissection. The diagnosis of was confirmed by histopathological
analysis. Patients with distant metastasis, incomplete clinical or
pathological data, impaired organ function, other malignant tumors
in the previous year, or were undergoing preoperative neoadjuvant
chemotherapy/emergency surgery for obstruction and perforation were
excluded. A summary of the patient clinical data is presented in
Table I. Primary gastric cancer
tissues, adjacent non-cancerous gastric tissues, positive
metastatic lymph node tissues and corresponding negative metastatic
lymph node tissues were obtained, fixed in 10% formalin at room
temperature for 48 h, and embedded in paraffin. The present study
was approved by the Research Ethics Boards of The First Hospital of
Jilin University, and all patients provided informed consent.
 | Table IUnivariate analysis of the
correlation between clinicopathological factors and survival time
of patients with gastric cancer (n=193). |
Table I
Univariate analysis of the
correlation between clinicopathological factors and survival time
of patients with gastric cancer (n=193).
| Clinicopathological
factors | Category | Cases | Average survival
time (days) | Log-rank test | P-value |
|---|
| Sex | Male | 150 |
1,250.625±71.768 | 0.303 | 0.582 |
| Female | 43 |
1,347.992±136.535 | | |
| Age (years) | ≤60 | 84 |
1,371.239±96.847 | 1.239 | 0.266 |
| >60 | 109 |
1,203.797±84.352 | | |
| Location of
tumor | Lower third | 111 |
1,256.182±84.888 | 0.033 | 0.984 |
| Middle third | 48 |
1,293.933±131.450 | | |
| Upper third | 34 |
1,284.501±132.457 | | |
| Tumor cell
differentiation | Poorly
differentiated | 118 |
1,188.670±82.583 | 2.154 | 0.142 |
| Moderately | 75 |
1,409.714±97.930 | | |
| differentiated | | | | |
| Vascular
invasion | No | 23 |
1,975.831±130.803 | 11.517 | 0.001 |
| Yes | 170 |
1,176.990±66.925 | | |
| Neural
invasion | No | 82 |
1,551.325±94.354 | 13.877 | <0.001 |
| Yes | 111 |
1,066.643±80.522 | | |
| T staging | T1 | 10 |
2,150.600±173.040 | 18.978 | <0.001 |
| T2 | 14 |
1,839.429±157.060 | | |
| T3 | 155 |
1,188.106±69.482 | | |
| T4 | 14 |
854.327±199.427 | | |
| N staging | N0 | 25 |
2,045.247±109.148 | 46.631 | <0.001 |
| N1 | 59 |
1,513.435±107.510 | | |
| N2 | 57 |
1,152.316±114.874 | | |
| N3 | 52 | 711.307±84.118 | | |
| Drinking | No | 147 |
1,290.952±73.459 | 0.190 | 0.663 |
| Yes | 46 |
1,202.467±124.052 | | |
| Smoking | No | 120 |
1,303.457±82.993 | 0.520 | 0.471 |
| Yes | 73 | 1,210.292
±95.941 | | |
| Tumor size | ≤10 | 57 |
1,402.576±118.432 | 1.809 | 0.179 |
| >10 | 136 |
1,223.221±75.350 | | |
| Family history | No | 173 |
1,280.990±66.943 | 0.476 | 0.788 |
| Yes | 19 |
1,232.173±216.603 | | |
| Obesity | No | 10 |
1,609.167±245.612 | 1.923 | 0.166 |
| Yes | 178 |
1,255.652±66.649 | | |
| ADAM9 | Negative | 2 |
556.500±123.500 | 2.524 | 0.112 |
| Positive | 191 |
1,284.413±64.362 | | |
| ADAM17 | Negative | 80 |
1,629.540±89.506 | 19.844 | <0.001 |
| Positive | 113 | 996.790±75.842 | | |
Immunohistochemistry and evaluation of
immunostaining intensity
Tissue sections (4 µm) cut from the
paraffin-embedded samples mentioned above were deparaffinized with
xylene and rehydrated with graded xylene and serial ethanol
concentrations. For antigen retrieval, the sections were incubated
with citrate buffer (0.01 M, pH 6.0) and microwaved at 95°C for 10
min. After rinsing thrice with PBS (pH 7.2), 3%
H2O2 in methanol was used to block endogenous
peroxidase activity at room temperature for 15 min. To reduce
non-specific reactions, 5% bovine serum albumin (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) and 0.3% Triton X-100
were added and incubated with the sections at room temperature for
1 h. Subsequently, the sections were probed with appropriate
antibodies against ADAM8 (1:100; cat. no. 23778-1-AP; ProteinTech
Group, Inc., Chicago, IL, USA), ADAM9 (1:75; cat. no. PA5-25959;
Thermo Fisher Scientific, Inc.), ADAM10 (1:250; cat no. PA5-28161;
Thermo Fisher Scientific, Inc.), ADAM12 (1:100; cat. no.
14139-1-AP), ADAM17 (1:300; cat. no. 20259-1-AP) and GAPDH (1:100;
cat. no. 10494-1-AP; all ProteinTech Group, Inc.) overnight at 4°C.
PBS was used as the negative control. Following three rinses with
PBS (pH 7.2), the sections were probed with horseradish peroxidase
(HRP)-labeled Peroxidase AffiniPure goat anti-rabbit IgG (1:300;
cat. no. 111-035-045) or Peroxidase AffiniPure HRP-labeled goat
anti-mouse IgG (1:400; cat. no. 115-035-003; both Jackson
ImmunoResearch Laboratories, Inc., West Grove, PA, USA) secondary
antibodies for 30 min at room temperature. Following three washes,
the sections were stained with 3,3′-diaminobenzidine (cat. no.
DA1010; Beijing Solarbio Science & Technology Co., Ltd.,
Beijing, China) at room temperature for 1 min, counterstained with
hematoxylin (cat. no. 517-28-2; Beijing Solarbio Science &
Technology Co., Ltd.) at room temperature for 50 sec, dehydrated
with graded concentrations of ethanol and xylene, and
coverslipped.
Under a light microscope (magnification, ×40), the
immunostaining intensity for each protein was reviewed and
independently scored by two pathologists who were blinded to the
clinical data and scored independently according to the staining
intensity and the proportion of stained tumor cells, as previously
described (20) with minor
modifications. According to the staining intensity, they were
scored as follows: No staining =0; light yellow (weak staining) =1;
yellow brown (moderate staining) =2; and brown (strong staining)
=3. The scores were expressed in terms of the proportion of cell
staining as follows: scores of 0, 1, 2, 3 and 4 indicated 0, ≤25,
26-50, 51-75, and ≥75% positive cells, respectively. Thus, the two
combined scores (from the two independent pathologists) were taken
as the final score: 0-1, 2-3, 4-5 and 6-7 indicated negat ive (−),
weak positive (+), strong positive (++), and very strong positive
(+++), respectively. In the statistical analysis, (+ +) and (+ + +)
were classified as the positive group, while (−) and (+) were
classified as the negative group.
Cell culture
Human gastric cancer cell lines KATO III and AGS
were purchased from the China Center for Type Culture Collection
(Wuhan University, Wuhan, China). SGC-7901 and BGC-823 cell lines
were obtained from the Shanghai Institute of Cell Biology of the
Chinese Academy of Science (Shanghai, China). All cells were grown
in RPMI-1640 medium (cat. no. 11995500BT) containing 10% fetal
bovine serum (cat. no. 10099-141; both Gibco; Thermo Fisher
Scientific, Inc.) and a mixture of penicillin (100 U/ml) and
streptomycin (100 µg/ml) at 37°C until 80-90%
confluence.
Cell transfection
The coding sequence of ADAM17 was synthesized by
Genewiz, Inc. (Suzhou, China). The overexpressed vector
pcDNA-3-ADAM17 was then constructed by inserting the coding
sequence of ADAM17 into the pcDNA-3 vector (Invitrogen; Thermo
Fisher Scientific, Inc.) with the restriction sites KpnI
(GGTACC) and EcoRI (GAATTC). The empty pcDNA-3 vector served
as the control. Small interfering RNAs (siRNAs) targeting ADAM17
(siRNA-ADAM17) and negative control siRNAs (siRNA-NC) were
synthesized by Shanghai GenePharma Co., Ltd. (Shanghai, China). The
sequences were as follows: siRNA-ADAM17 forward, 5′-GGG CCG AAU AUA
ACA UAG AdTdT-3′ and reverse, 5′-UCU AUG UUA UAU UCG GCC CdTdT-3′;
siRNA-NC forward, 5′-UUC UCC GAA CGU GUC ACG UdTdT-3′ and reverse,
5′-ACG UGA CAC GUU CGG AGA AdTdT-3′.
These siRNAs were designed using BLOCK-iT™ RNAi
Designer (Thermo Fisher Scientific, Inc.). For cell transfection,
SGC-7901 and BGC-823 cells (2.4×105/well) were seeded
into a six-well plate and incubated for 24 h at 37°C. Next, 2
µg pcDNA-3-ADAM17 and 2 µg pcDNA-3 were transfected
into BGC-823 cells using Lipofectamine® 2000 reagent
(cat. no. 11668-027; Invitrogen; Thermo Fisher Scientific, Inc.).
si-RNA-ADAM17 (50 nM) BS siRNA-NC (50 nM) were transfected into
SGC-7901 cells using the same method. Cells were incubated for
another 48 h at 37°C before performing further experiments.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total mRNA was extracted from transfected cells
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
Reverse transcription of mRNA to cDNA was performed using the
PrimeScript RT Master Mix kit (cat. no. RR036A; Takara Bio, Inc.,
Otsu, Japan) with the following conditions: 37°C for 15 sec, 85°C
for 5 sec; then held at 4°C. To detect the expression levels of
ADAM17 mRNA, qPCR was performed using a SYBR Green kit (cat. no.
4367659; Invitrogen) on an ABI ViiA7 PCR system (both Thermo Fisher
Scientific, Inc.). Primers used for the amplification of targets
were as follows: ADAM17 forward, 5′-ATC AAA CCC TTT CCT GCG-3′ and
reverse, 5′-CAA ACC CAT CCT CGT CCA-3′; GAPDH forward, 5′-TGA CAA
CTT TGG TAT CGT GGA AGG-3′ and reverse, 5′-AGG CAG GGA TGA TGT TCT
GGA GAG-3′. The thermocycling conditions were as follows: 95°C for
10 min, followed by 40 cycles at 95°C for 15 sec and 60°C for 1
min, and finally 95°C for 15 sec, 60°C for 1 min, and 95°C for 15
sec. The relative expression of ADAM17 mRNA was normalized to GAPDH
and was quantified using the 2−ΔΔCq method (25).
Cell Counting kit-8 (CCK-8) assay
Cells (5×106/well) were seeded in a
96-well plate and incubated for 24 h and were subsequently
transfected with pcDNA-3-ADAM17, pcDNA-3, siRNA-ADAM17 or siRNA-NC.
Each treatment was performed in triplicate wells. After
transfection for 48 h, cell viability of each group was assessed
using a CCK-8 kit (cat. no. CK04, Dojindo Molecular Technologies,
Inc., Kumamoto, Japan) following the manufacturers’ recommended
protocols. Absorbance at 450 nm was measured on a microplate reader
(BioTek Instruments, Inc., Winooski, VT, USA).
Scratch wound healing assay
The cell migratory capacity of each group was
evaluated using the scratch wound healing assay, as described
previously (26). Briefly,
transfected cells were seeded in culture dishes in triplicate and
were incubated at 37°C until a confluent monolayer was formed
(>90%). With a 10 µl sterile pipette tip, a ‘scratch’ of
the cell monolayer was created in a straight line. The cells were
washed thrice with PBS (pH 7.2) to remove cell fragments, and the
cells were incubated with serum-free RPMI-1640 medium for 24 h at
37°C. The migrated cells were observed under an inverted light
microscope (Nikon Corporation, Tokyo, Japan).
GSEA of gastric cancer expression
GSEA (www.broadin-stitute.org/gsea/index.jsp ) (27) is a computational method for
interpreting gene expression data; it creates a molecular signature
database based on known information on the positions,
characteristics and biological functions of genes. To investigate
the role of ADAM17 in gastric cancer, mRNA sequence data of stomach
adenocarcinoma from The Cancer Genome Atlas (TCGA; tcga-data.nci.nih.gov/) was downloaded, which included
415 stomach adenocarcinoma samples. Key pathways that were enriched
by gene sets significantly associated with ADAM17 were then
analyzed by GSEA, and P<0.05 was set as the cut-off value.
Western blot analysis
Total protein was extracted from gastric cancer
tissues and cells using radioimmunoprecipitation assay buffer (cat.
no. PL007; Sangon Biotech Co., Ltd., Shanghai, China) for 20 min on
ice. Protein concentration was determined with bicinchoninic acid
protein assay kit (cat no. 23223; Pierce; Thermo Fisher Scientific,
Inc.). For western blotting, protein extracts (30 µg/lane)
were subjected to 10% SDS-PAGE and transferred onto a
polyvinylidene fluoride membrane. Following blocking in 1X
Tris-buffered saline/0.05% Tween-20 (TBST) supplemented with 5%
non-fat milk at room temperature for 1 h, primary antibodies
against ADAM17 (1:1,000; cat. no. 20259-1-AP; ProteinTech Group,
Inc.), ADAM9 (1:1,000; cat. no. PA5-25959; Thermo Fisher
Scientific, Inc.), neurogenic locus notch homolog protein (Notch)2
(1:1,000; cat. no. WL02409; Wanleibio Co., Ltd., Shanghai, China);
glycogen synthase kinase (GSK)-3β (1:1,000; cat. no. 22104-1-AP;
ProteinTech Group, Inc.), β-catenin (1:1,000; cat. no. 8480S; Cell
Signaling Technology, Inc., Danvers, MA, USA) or β-actin (1:2,000;
cat. no. 60008-1-Ig; ProteinTech Group, Inc.) were added to the
membranes and incubated overnight at 4°C. β-actin was used as the
internal control. After washing with 1X PBST thrice, HRP-labeled
Peroxidase AffiniPure goat anti-rabbit IgG (1:5,000; cat. no.
111-035-045; Jackson ImmunoResearch Laboratories, Inc.) or
Peroxidase AffiniPure goat anti-mouse IgG HRP (1:5,000; cat. no.
115-035-003; Jackson ImmunoResearch Laboratories, Inc.) was added
to the membranes and incubated at room temperature for another 2 h.
After rinsing five times with 1X PBST, protein bands were
visualized using chemiluminescent HRP substrate (cat. no.
WBKLS0500; Merck KGaA, Darmstadt, Germany). The intensity of each
protein band was quantified and analyzed using Tanon Image Software
version 1.10 (Tanon 1600R; Tanon Science and Technology Co., Ltd.,
Shanghai, China).
Statistical analysis
Univariate survival analysis for the comparison of
survival times was conducted using the Kaplan-Meier method, and the
log-rank test was applied to analyze the significance of difference
between survival times of patients and their clinicopathological
features, as well as between survival times and ADAM9 or ADAM17
expression. Multivariate survival analysis was performed for
identifying significant variables associated with survival times
using the Cox regression proportional hazards model. In addition,
the risk scores of death were calculated with multivariate logistic
regression analysis (28)
According to the T staging, N staging and ADAM17 values, the risk
score of each patient (ranging from 0 to 100) was calculated. The
receiver operating characteristic (ROC) curve was applied, and the
area under the curve (AUC) was determined by the MedCalc
statistical software version 11.0 (MedCalc, Mariakerke, Belgium).
Data were presented as the mean ± standard deviation, and the
differences between two groups were compared using the Student’s
t-test and among three or more groups were analyzed by one-way
analysis of variance, followed by Fisher’s least significant
difference test. Statistical analyses were carried out using SPSS
17.0 (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
ADAM9 and ADAM17 are significantly
upregulated in gastric cancer and positive metastatic lymph node
tissues
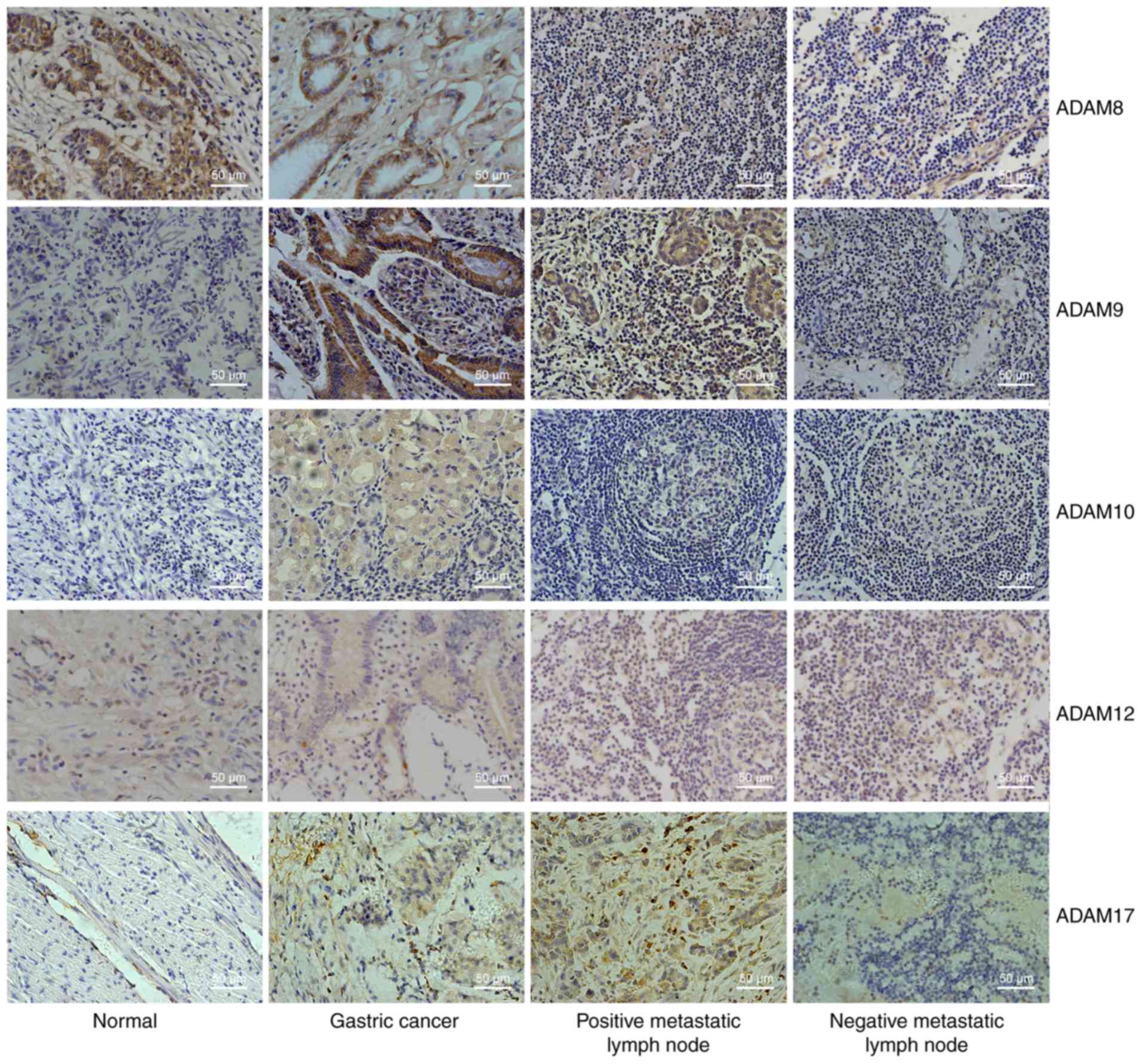
Fig. 1 shows the
immunohistochemical staining intensity for ADAM8, ADAM9, ADAM10,
ADAM12 and ADAM17 in adjacent non-cancerous gastric tissues,
gastric cancer tissues, positive metastatic lymph node tissues and
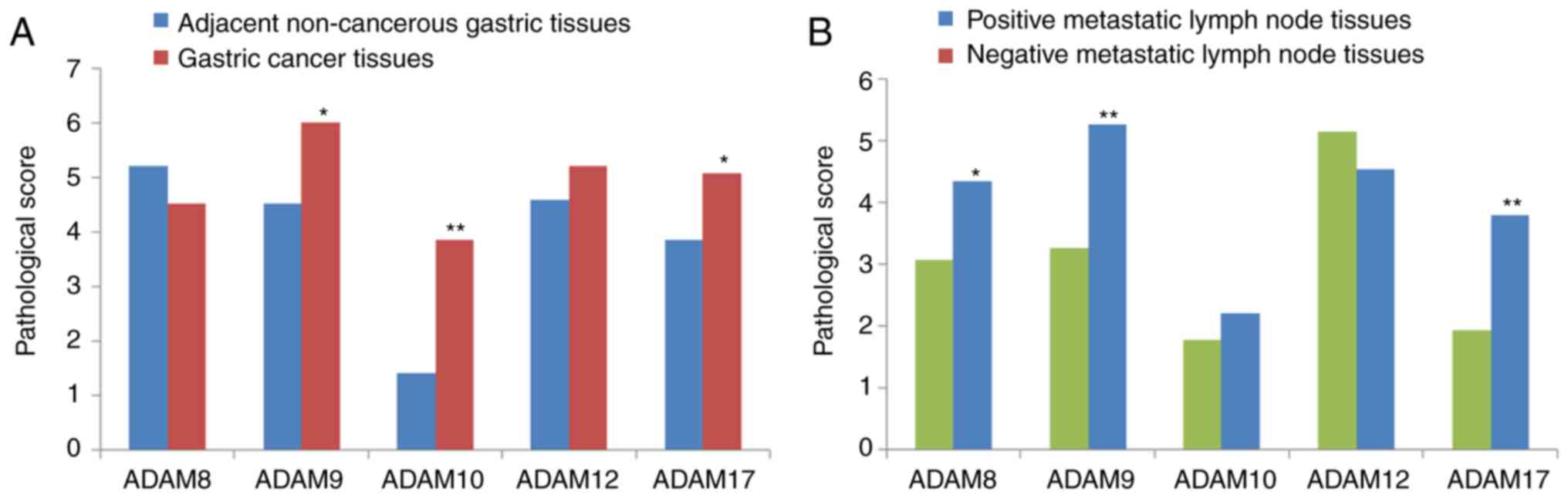
negative metastatic lymph node tissues. From the combined score
that was obtained from the staining intensity and the percentage of
cell staining, the expression of ADAM9, ADAM10, and ADAM17 in
gastric tumor tissues was significantly upregu-lated compared to
those in adjacent normal tissues (P<0.05; Fig. 2A). Furthermore, the expression of
ADAM8, ADAM9 and ADAM17 in positive metastatic lymph node tissues
was also upregulated relative to those in the corresponding
negative tissues (P<0.05; Fig.
2B). These data indicated that ADAM9 and ADAM17 were
significantly upregulated in both gastric cancer and positive
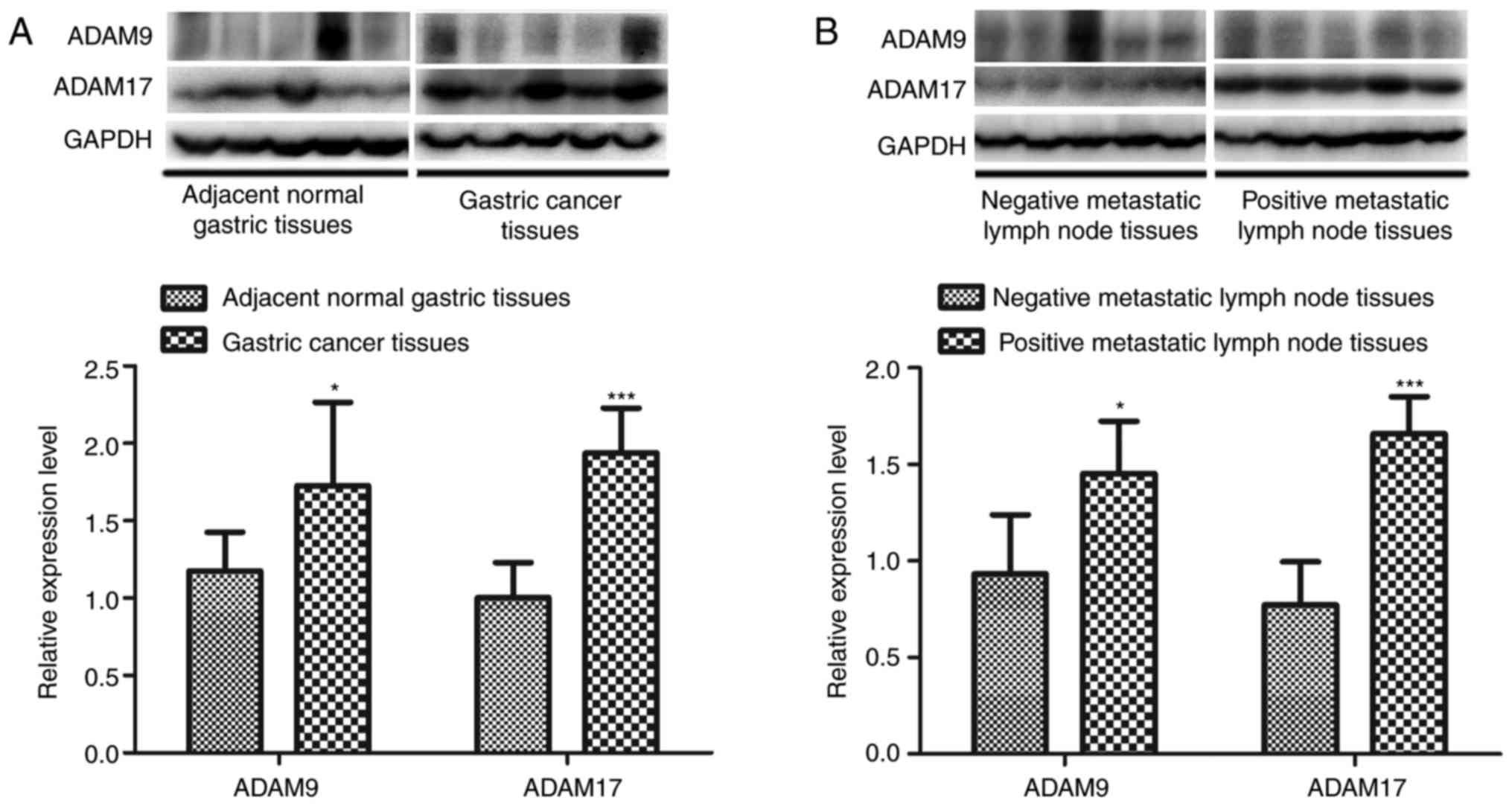
metastatic lymph node tissues. To further verify the
immunohistochemistry results, the expression of ADAM9 and ADAM17
was detected by western blotting. As expected, ADAM9 and ADAM17
expression was significantly upregulated in primary gastric tumor
tissues and positive metastatic lymph node tissues (P<0.05;
Fig. 3).
Survival times of patients correlated
with T staging, N staging, and ADAM17 expression
Univariate survival analysis for the comparison of
survival times between patients with clinicopathological features
was performed. As presented in Table
I, the survival times of patients were significantly associated
with vascular invasion, neural invasion, T staging, N staging and
ADAM17 expression (P<0.05). Therefore, multivariate analysis was
used to identify significant clini-copathological features
associated with survival times. The results demonstrated that the
survival times of patients strongly correlated with T staging, N
staging and ADAM17 expression (P<0.05; Table II). Furthermore, multivariate
logistic regression analysis showed similar results that the above
three variables were independent predictors (P<0.05; Table III). The above three variables
were then included in the ‘risk score’ calculation to measure the
‘mortality risk score’ of any given patients, as follows:
Probability =1/[1+exp [5.454-0.993 (T staging)-0.720 (N
staging)-0.771 (ADAM17)]].
| Table IIMultivariate survival analysis of
gastric cancer (Cox proportional hazards model). |
Table II
Multivariate survival analysis of
gastric cancer (Cox proportional hazards model).
| Covariates | Variable
coefficient | Standard error | Wald | P-value | HR | 95% CI |
|---|
| ADAM17 | 0.806 | 0.199 | 16.430 | <0.001 | 2.239 | 1.516-3.305 |
| T staging (T2/3/4
vs. T1) | 0.598 | 0.192 | 9.677 | 0.002 | 1.818 | 1.248-2.650 |
| N staging (N1/2/3
vs. N0) | 0.543 | 0.104 | 27.147 | <0.001 | 1.722 | 1.404-2.113 |
| Vascular
invasion | 0.408 | 0.406 | 1.009 | 0.315 | 1.504 | 0.678-3.336 |
| Neural
invasion | 0.092 | 0.211 | 0.193 | 0.661 | 1.097 | 0.726-1.658 |
| Table IIIIndependent predictors of lymph node
metastasis based on the multivariate logistic regression
analysis. |
Table III
Independent predictors of lymph node
metastasis based on the multivariate logistic regression
analysis.
| Variable(s) | Regression
coefficients | Standard error | Wald | P-value | OR | 95% CI for OR
|
|---|
| Lower | Upper |
|---|
| T staging | 0.993 | 0.350 | 8.052 | 0.005 | 2.700 | 1.360 | 5.362 |
| N staging | 0.720 | 0.184 | 15.338 | <0.001 | 2.055 | 1.433 | 2.947 |
| ADAM17 | 0.771 | 0.338 | 5.207 | 0.022 | 2.161 | 1.115 | 4.190 |
| Constant | −5.454 | 1.203 | 20.569 | <0.001 | 0.004 | | |
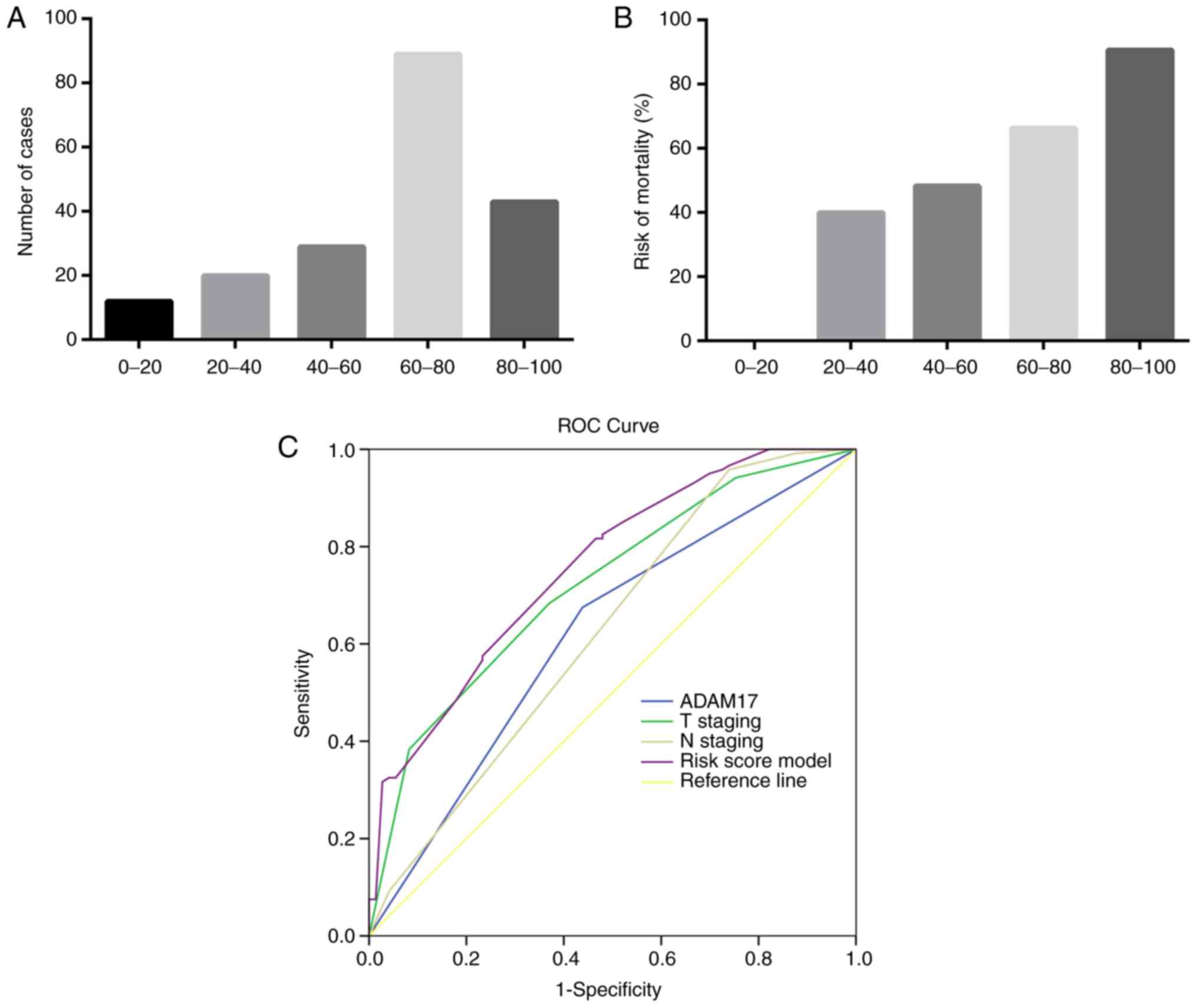
The distribution of the risk score of 193 patients
is presented in Fig. 4. The
majority of scores were distributed between 60 and 100 (60-80,
n=89; 80-100, n=43; Fig. 4A),
suggesting that this model can separate the low- and high-risk
groups, and the risk of mortality increased with the increase in
risk scores (Fig. 4B). To further
determine the optimal model to predict the risk of death, ROC curve
analysis was performed. As shown in Table IV and Fig. 4C, the AUCs for the risk score
model, T staging, N staging, and ADAM17 expression were 0.757,
0.625, 0.720 and 0.618, respectively. These data indicated that T
staging, N staging and ADAM17 expression had independent prognostic
value for predicting the risk of patients with gastric cancer.
| Table IVAnalysis of AUC of different
predictors for mortality risk. |
Table IV
Analysis of AUC of different
predictors for mortality risk.
| Test variable | AUC | Standard error | P-value | 95% Confidence
Interval
|
|---|
| Lower bound | Upper bound |
|---|
| ADAM17 | 0.618 | 0.042 | 0.006 | 0.536 | 0.701 |
| T staging | 0.625 | 0.043 | 0.004 | 0.541 | 0.709 |
| N staging | 0.720 | 0.037 | 0.000 | 0.648 | 0.793 |
| Risk score
model | 0.757 | 0.035 | <0.001 | 0.688 | 0.826 |
ADAM17 was highly expressed in gastric
cancer cells with high metastatic potential
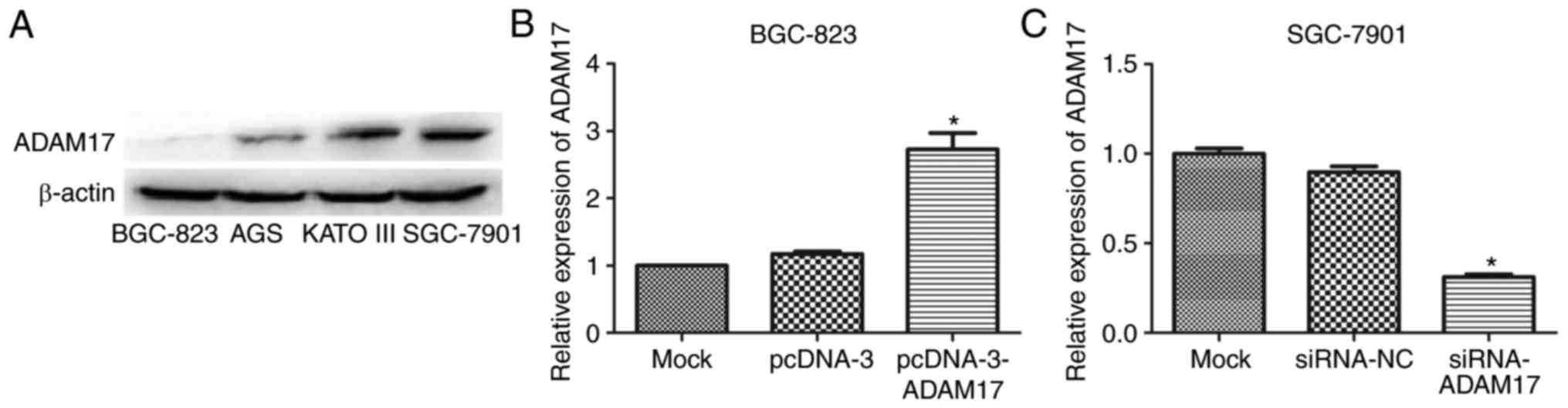
Next, the expression of ADAM7 in gastric cancer
cells was determined. As shown in Fig. 5A, ADAM17 expression in KATO III
and SGC-7901 cells that had high metastatic potential was
significantly higher than that in BGC-823 and AGS cells with low
metastatic potential, suggesting that ADAM17 may be associated with
tumor cell metastasis. Among the above four cell lines, ADAM17
expression was lowest in BGC-823 cells and highest in SGC-7901
cells. Therefore, BGC-823 and SGC-7901 cells were used for the
overexpression and repression of ADAM17 expression, respectively.
As expected, ADAM17 expression was significantly increased in
pcDNA-3-ADAM17-transfected BGC-823 cells, compared with mock cells
or pcDNA-3-transfected cells (P<0.05; Fig. 5B), while its expression was
markedly reduced in siRNA-ADAM17-trans-fected SGC-7901 cells
compared with either mock cells or siRNA-NC-transfected cells
(P<0.05; Fig. 5C).
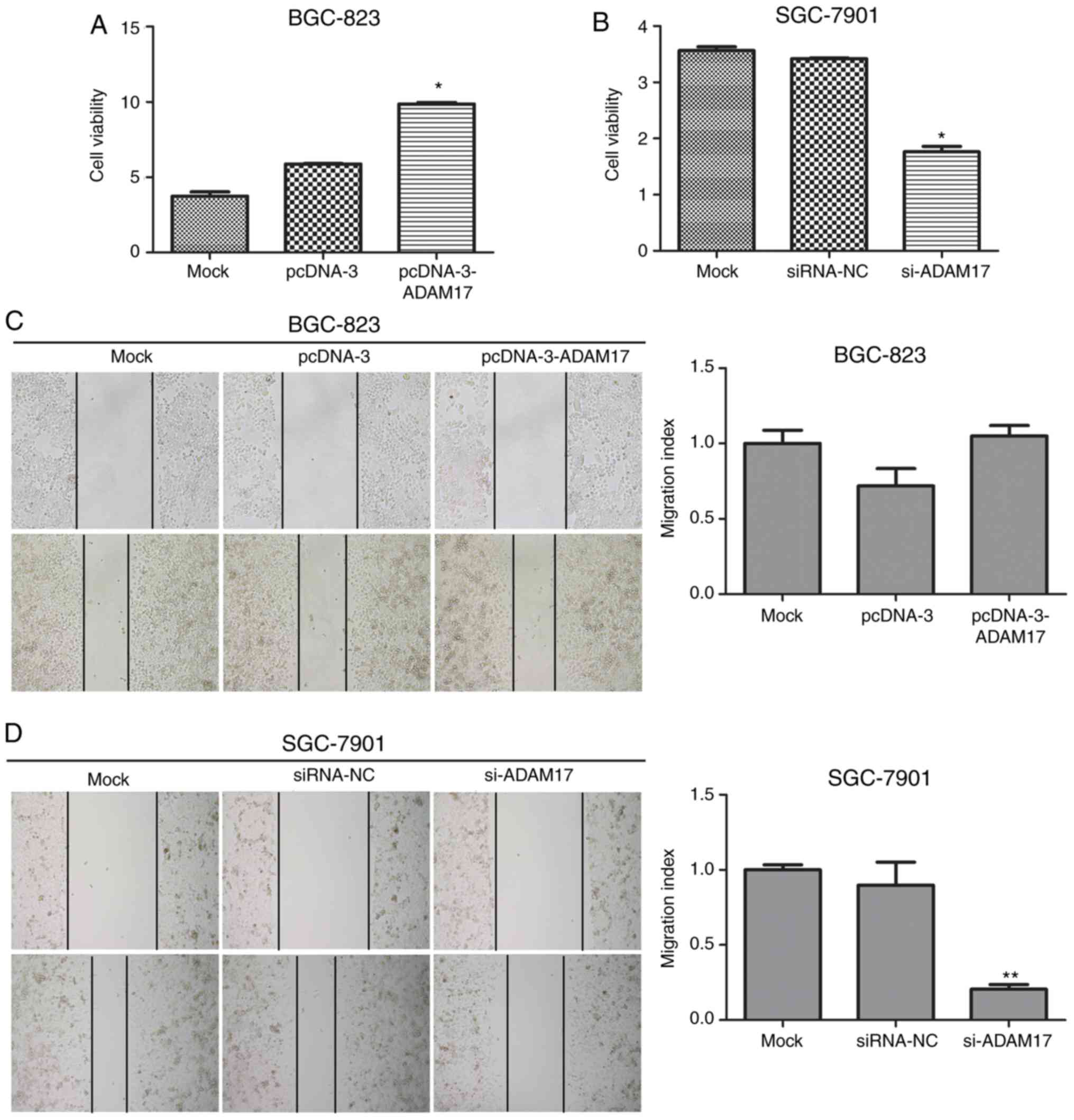
ADAM17 promotes the viability and
migration of gastric cancer cells
The effects of ADAM17 on cell viability in each
group was assessed using the CCK-8 assay. As presented in Fig. 6A, the overexpression of ADAM17 in
pcDNA-3-ADAM17-transfected BGC-823 cells resulted in a significant
increase in cell viability, compared with mock cells or
pcDNA-3-transfected cells (P<0.05). However, the cell viability
of siRNA-ADAM17-transfected SGC-7901 cells was significantly
reduced compared with the mock or siRNA-NC transfected cells
(P<0.05; Fig. 6B). Next, the
scratch wound healing assay was performed to study the effects of
ADAM17 on cell migration. It was revealed that the number of
migrated SGC-7901 cells was significantly decreased following
ADAM17 silencing (P<0.05; Fig.
6D). However, the migratory capacity between
pcDNA-3-ADAM17-transfected BGC-823 cells, mock cells and
pcDNA-3-transfected cells did not show significant difference
(P>0.05; Fig. 6C). Taken
together, these results suggested that ADAM17 promoted gastric
cancer cell viability and migration.
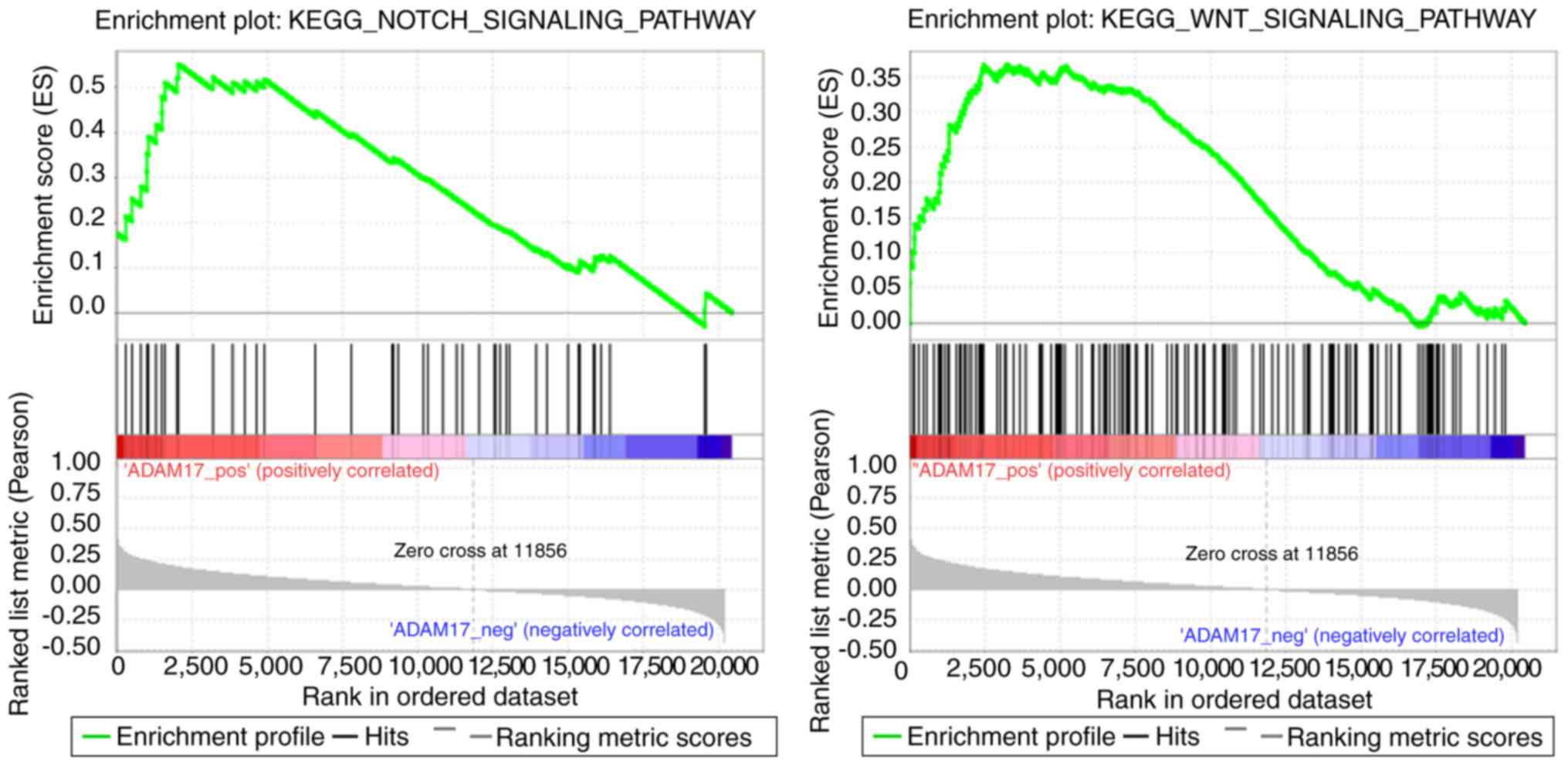
GSEA for analyzing ADAM17 function in
gastric cancer
The results of GSEA revealed 12 pathways, including
the Notch and Wnt signaling pathways (Fig. 7), that positively correlated with
ADAM17 expression (Table V).
Furthermore, 19 pathways associated with metabolism, including
oxidative phosphorylation and phenylalanine metabolism, were
negatively correlated with ADAM17 expression (Table V).
| Table VResults of gene set enrichment
analysis showed key pathways correlated with ADAM17 expression in
gastric cancer. |
Table V
Results of gene set enrichment
analysis showed key pathways correlated with ADAM17 expression in
gastric cancer.
| Name | Size | ES | NES | P-value |
|---|
|
|---|
| A, Positively
correlation with ADAM17 expression | | | | |
|---|
| Notch signaling
pathway | 47 | 0.55082 | 1.91005 | 0.00178 |
| Ubiquitin-mediated
proteolysis | 134 | 0.45800 | 1.82770 | 0.00180 |
| Small cell lung
cancer | 84 | 0.44413 | 1.66057 | 0.00361 |
| Adherens
junction | 73 | 0.43867 | 1.62956 | 0.01235 |
| Glycosaminoglycan
biosynthesis heparan sulfate | 26 | 0.53938 | 1.60466 | 0.02632 |
| ErbB signaling
pathway | 87 | 0.41745 | 1.58098 | 0.02330 |
| Prostate
cancer | 89 | 0.40135 | 1.57703 | 0.01431 |
| Pathways in
cancer | 324 | 0.35638 | 1.56122 | 0.01165 |
|
Progesterone-mediated oocyte
maturation | 85 | 0.40244 | 1.54025 | 0.01783 |
| Renal cell
carcinoma | 70 | 0.40003 | 1.51718 | 0.03209 |
| Axon guidance | 128 | 0.39454 | 1.51392 | 0.03375 |
| Wnt signaling
pathway | 149 | 0.36756 | 1.49736 | 0.02993 |
|
| B, Negative
correlation with ADAM17 expression |
|
| Parkinson’s
disease | 113 | −0.78448 | −2.33212 | <0.001 |
| Oxidative
phosphorylation | 116 | −0.77444 | −2.23988 | <0.0001 |
| Huntington’s
disease | 172 | −0.62239 | −2.23308 | 0.00217 |
| Alzheimer’s
disease | 156 | −0.61537 | −2.22474 | 0.00221 |
| Cardiac muscle
contraction | 73 | −0.54398 | −2.06387 | 0.00236 |
| Ribosome | 87 | −0.84358 - | 2.01567 | 0.00408 |
| Primary bile acid
biosynthesis | 16 | −0.62983 | −1.80464 | 0.01307 |
| Phenylalanine
metabolism | 18 | −0.56414 | −1.72816 | 0.01659 |
| Peroxisome | 78 | −0.4785 | −1.72701 | 0.01379 |
| Glutathione
metabolism | 50 | −0.5088 | −1.70551 | 0.01235 |
| Linoleic acid
metabolism | 28 | −0.50486 | −1.69893 | 0.01566 |
| Tryptophan
metabolism | 39 | −0.46789 | −1.68343 | 0.02103 |
| Citric acid
cycle | 30 | −0.61788 | −1.67656 | 0.04651 |
| Drug metabolism
cytochrome p450 | 71 | −0.47023 | −1.67642 | 0.02727 |
| Tyrosine
metabolism | 42 | −0.43363 | −1.64599 | 0.01232 |
| Metabolism of
xenobiotics by cytochrome p450 | 69 | −0.46883 | −1.63288 | 0.03529 |
| Histidine
metabolism | 28 | −0.45888 | −1.58055 | 0.02778 |
| Fatty acid
metabolism | 42 | −0.49172 | −1.57598 | 0.04292 |
| Arachidonic acid
metabolism | 57 | −0.39434 | −1.50295 | 0.04798 |
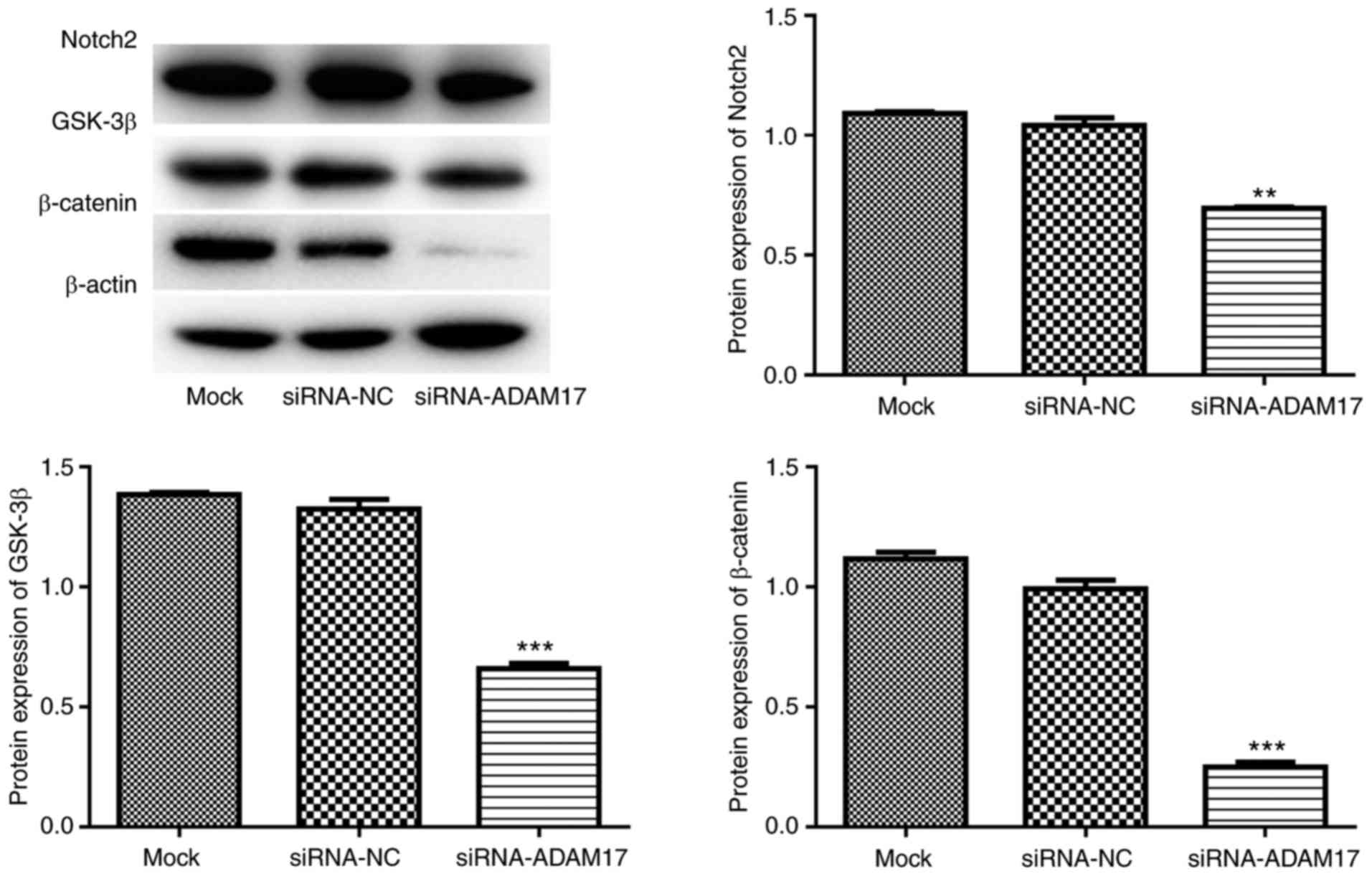
Suppression of ADAM17 in SGC-7901 cells
inhibits Notch and Wnt signaling pathways
To further verify the results of GSEA, western blot
analysis was performed to investigate the expression of key
proteins involving in Notch and Wnt signaling pathways in SGC-7901
cells following the suppression of ADAM17 expression. The results
demonstrated that suppression of ADAM17 in SGC-7901 cells resulted
in a significant downregulation of Notch2, GSK-3β and β-catenin
expression (Fig. 8), suggesting
that suppression of ADAM17 in SGC-7901 cells inhibited the Notch
and/or Wnt signaling pathways.
Discussion
Tumor metastasis is one of the main causes of
treatment failure among patients with cancer, and ADAMs have been
implicated in tumor metastasis and progression (29). Therefore, a better understanding
of the key members of the ADAM family that are involved in gastric
cancer progression will facilitate the development of a promising
therapeutic regimen.
In the present study, ADAM17 was found to be
significantly upregulated in gastric cancer and positive metastatic
lymph node tissues. Consistent with previous findings reporting
that ADAM17 is an independent prognostic factor for patients with
gastric cancer (23), a strong
correlation was identified between the survival time of patients
and ADAM17 expression, confirming that ADAM17 could be used as an
independent prognostic factor. In addition, it was shown that
ADAM17 was upregulated in gastric cancer cells with high metastatic
potential. Notably, ADAM17 overexpression significantly increased
BGC-823 cell viability with low metastatic potential, while the
knockdown of ADAM17 reduced the viability and migratory capacity of
SGC-7901 cells, which have high metastatic potential. Furthermore,
GSEA revealed a strong positive correlation between ADAM17
expression and the Notch/Wnt signaling pathways. Further
experiments confirmed that suppression of ADAM17 in SGC-7901 cells
inhibited the Notch and Wnt signaling pathways.
Metastasis is a multistep process mediating the
progression of malignant tumors, in which cell-cell and cell-matrix
interactions are implicated. These interactions result in the
activation of numerous cytokines and growth factors and the
subsequent generation of signals to promote tumor growth and
survival (30). ADAM17 affects
various growth factors, cytokines, receptors, and cell adhesion
molecules, which are all broadly involved in regulating the
proliferation, migration and invasion of tumor cells (31). Because angiogenesis is essential
for tumor growth and metastasis (32,33), and ADAM17 is a central regulator
of angiogenesis (34), ADAM17 may
also influence tumor angiogenesis and invasion (35). ADAM17 promotes the malignant
phenotype of U87 human glioma cells by increasing their
proliferation and invasion (36),
and the silencing of ADAM17 significantly suppresses the
proliferation and invasion of A549 cells in vitro (37). Furthermore, a previous report
confirmed that ADAM17 is highly expressed in lymph node-positive
breast cancer tissues in comparison with lymph node-negative
tissues (38), implying the
association between ADAM17 and lymph node metastasis. Notably,
ADAM17 has been shown to prevent the miR-338-3p-mediated inhibition
of gastric cancer cell migration and invasion (22). Xu et al (24) demonstrated that ADAM17 promotes
EMT in gastric cancer cells through the TGF-β/Smad pathway, and an
increasing number of studies have indicated that EMT is a
phenotypic conversion associated with cancer metastasis (39,40). In the present study, ADAM17 was
found to be upregulated in gastric cancer and lymph node-positive
gastric cancer tissues. In addition, ADAM17 was upregulated in
gastric cancer cells with high metastatic potential. Importantly,
ADAM17 silencing decreased the viability and migratory capacity of
SGC-7901 cells with high metastatic potential. However, ADAM17
overexpression only increased the viability of BGC-823 cells, and
did not promote the migratory capacity, which may be due to their
low metastatic potential. Taken together, it was speculated that
the ADAM17 upregulation facilitated the metastasis of gastric
cancer by promoting cell viability and migration.
The increased expression of ADAM17 is closely
correlated with poor prognosis in many human cancers, such as
gallbladder carcinoma (41) and
breast cancer (42) and may serve
as a poor prognostic factor (43,44). ADAM17 expression also has
prognostic significance in patients with gastric cancer (21,45). Furthermore, ADAM17 is considered
to be a potential target in anticancer treatment or as an indicator
for predicting therapeutic outcomes (42). Kyula et al (46) demonstrated that chemotherapy with
fluorouracil acutely activates ADAM17 and consequently results in
drug resistance in colorectal cancer. In line with these previous
findings, the results of the multivariate analysis performed in the
present study showed a strong correlation between the survival time
of patients and the expression of ADAM17. Therefore, it was
hypothesized that increased expression of ADAM17 may have been
involved in poor gastric cancer prognosis, and that it may serve as
an independent prognostic marker. Targeting ADAM17 may have
prospects in cancer prognosis and therapy.
GSEA and western blotting revealed a positive
correlation between ADAM17 expression and the Notch and Wnt
signaling pathways. An aberration in Notch signaling results in the
metastasis of various cancers via the regulation of EMT or tumor
angiogenesis (47). A
meta-analysis confirmed that the Notch signaling pathway is a key
pathway for mediating tumor progression in gastric cancer (48). Brzozowa et al (49) revealed that the Notch signaling
pathway serves a key role in the pathogenesis of gastric cancer. In
addition, the Wnt signaling pathway is closely correlated with lung
cancer and bone metastasis (50),
and Wnt/β-catenin signaling regulates tumor metastasis in breast
cancer (51,52). Yanaka et al (53) revealed that the activation of the
Wnt signaling pathway is a key mechanism in mediating
miR-544a-induced EMT to regulate gastric cancer progression. Tan
et al (54) confirmed that
dixin promotes the metastasis of gastric cancer via activation of
the Wnt signaling pathway. Given the key role of the Notch and Wnt
signaling pathways in the metastasis and progression of gastric
cancer, it was speculated that these pathways are key mechanisms by
which ADAM17 mediates its effects in gastric cancer.
However, the current study had several limitations.
First, the clinical sample size used was relatively small and not
sufficient. More samples should be collected to confirm the
results. Second, almost all patients were on adjuvant therapy;
thus, it was difficult to clearly verify whether ADAM17 was a pure
prognostic factor. Third, an invasion assay was not performed to
evaluate the function of ADAM17 in the cell invasion ability. An
invasion assay should be performed to evaluate the function of
ADAM17 in cell invasion ability in future studies, which will
provide strong evidence supporting the role of ADAM17 in gastric
cancer metastasis and progression. Lastly, the relationship between
ADAM17 and Notch or Wnt signaling pathway was only preliminarily
confirmed in SGC-7901 cells. More studies are still required to
verify these findings.
In conclusion, the data in the present study
demonstrated that increased ADAM17 expression may have contributed
to gastric cancer metastasis and progression, potentially via
activation of the Notch or Wnt signaling pathways, and that ADAM17
may serve as a useful prognostic marker in the treatment of gastric
cancer.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81402374 and
81372295) and Excellent Young Foundation of Jilin Scientific and
Technological Development Program (grant no. 20170520002JH).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors’ contributions
WL, JS and DW contributed to the study design. WL,
LW and YZ collected the data and performed the experiments. WL and
XS interpreted and discussed the data. WL and DW analyzed the data.
WL drafted the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Boards of The First Hospital of Jilin University, and all
patients provided informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Carcas LP: Gastric cancer review. J
Carcinogenesis. 13:142013. View Article : Google Scholar
|
|
2
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guggenheim DE and Shah MA: Gastric cancer
epidemiology and risk factors. J Surg Oncol. 107:230–236. 2013.
View Article : Google Scholar
|
|
4
|
Wadhwa R, Taketa T, Sudo K, Blum MA and
Ajani JA: Modern oncological approaches to gastric adenocarcinoma.
Gastroenterol Clin North Am. 42:359–369. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee JH, Kim Y, Choi JW and Kim YS: Genetic
variants and risk of gastric cancer: A pathway analysis of a
genome-wide association study. Springerplus. 4:2152015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ang TL and Fock KM: Clinical epidemiology
of gastric cancer. Singapore Med J. 55:621–628. 2014. View Article : Google Scholar
|
|
7
|
Lordick F, Kang YK, Chung HC, Salman P, Oh
SC, Bodoky G, Kurteva G, Volovat C, Moiseyenko VM, Gorbunova V, et
al: Capecitabine and cisplatin with or without cetuximab for
patients with previously untreated advanced gastric cancer
(EXPAND): A randomised, open-label phase 3 trial. Lancet Oncol.
14:490–499. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Coburn NG: Lymph nodes and gastric cancer.
J Surg Oncol. 99:199–206. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wildeboer D, Naus S, Amy Sang QX, Bartsch
JW and Pagenstecher A: Metalloproteinase disintegrins ADAM8 and
ADAM19 are highly regulated in human primary brain tumors and their
expression levels and activities are associated with invasiveness.
J Neuropathol Exp Neurol. 65:516–527. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Reiss K and Saftig P: The ‘a disintegrin
and metalloprotease’ (ADAM) family of sheddases: Physiological and
cellular functions. Semin Cell Dev Biol. 20:126–137. 2009.
View Article : Google Scholar
|
|
11
|
Kim KE, Song H, Hahm C, Yoon SY, Park S,
Lee HR, Hur DY, Kim T, Kim CH, Bang SI, et al: Expression of ADAM33
is a novel regulatory mechanism in IL-18-secreted process in
gastric cancer. J Immunol. 182:3548–3555. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Carl-Mcgrath S, Lendeckel U, Ebert M,
Roessner A and Röcken C: The disintegrin-metalloproteinases ADAM9,
ADAM12, and ADAM15 are upregulated in gastric cancer. Int J Oncol.
26:17–24. 2005.
|
|
13
|
Wang YY, Ye ZY, Li L, Zhao ZS, Shao QS and
Tao HQ: ADAM 10 is associated with gastric cancer progression and
prognosis of patients. J Surg Oncol. 103:116–123. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Murphy G: The ADAMs: Signalling scissors
in the tumour microenvironment. Nat Rev Cancer. 8:929–941. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gall SML, Bobé P, Reiss K, Horiuchi K, Niu
XD, Lundell D, Gibb DR, Conrad D, Saftig P and Blopel CP: ADAMs 10
and 17 represent differentially regulated components of a general
shedding machinery for membrane proteins such as transforming
growth factor alpha, L-selectin, and tumor necrosis factor alpha.
Mol Biol Cell. 20:1785–1794. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lorenzen I, Trad A and Grötzinger J:
Multimerisation of A disintegrin and metalloprotease protein-17
(ADAM17) is mediated by its EGF-like domain. Biochem Biophys Res
Commun. 415:330–336. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ni SS, Zhang J, Zhao WL, Dong XC and Wang
JL: ADAM17 is overexpressed in non-small cell lung cancer and its
expression correlates with poor patient survival. Tumour Biol.
34:1813–1818. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Franovic A, Robert I, Smith K, Kurban G,
Pause A, Gunaratnam L and Lee S: Multiple acquired renal carcinoma
tumor capabilities abolished upon silencing of ADAM17. Cancer Res.
66:8083–8090. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shen H, Li L, Zhou S, Yu D, Yang S, Chen
X, Wang D, Zhong S, Zhao J and Tang J: The role of ADAM17 in
tumorigenesis and progression of breast cancer. Tumour Biol. 2016.
View Article : Google Scholar
|
|
20
|
Shou ZX, Jin X and Zhao ZS: Upregulated
expression of ADAM17 is a prognostic marker for patients with
gastric cancer. Ann Surg. 256:1014–1022. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang TC, Zhu WG, Huang MD, Fan RH and
Chen XF: Prognostic value of ADAM17 in human gastric cancer. Med
Oncol. 29:2684–2690. 2012. View Article : Google Scholar
|
|
22
|
Chen JT, Yao KH, Hua L, Zhang LP, Wang CY
and Zhang JJ: MiR-338-3p inhibits the proliferation and migration
of gastric cancer cells by targeting ADAM17. Int J Clin Exp Pathol.
8:10922–10928. 2015.PubMed/NCBI
|
|
23
|
Fang W, Qian J, Wu Q, Chen Y and Yu G:
ADAM-17 expression is enhanced by FoxM1 and is a poor prognostic
sign in gastric carcinoma. J Surg Res. 220:223–233. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu M, Zhou H, Zhang C, He J, Wei H, Zhou
M, Lu Y, Sun Y, Ding JW, Zeng J, et al: ADAM17 promotes
epithelial-mesenchymal transition via TGF-β/Smad pathway in gastric
carcinoma cells. Int J Oncol. 49:2520–2528. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
26
|
Cory G: Scratch-wound assay. Methods Mol
Biol. 769:25–30. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES and Mesirov JP: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Marrelli D, De Stefano A, de Manzoni G,
Morgagni P, Di Leo A and Roviello F: Prediction of recurrence after
radical surgery for gastric cancer: A scoring system obtained from
a prospective multicenter study. Ann Surg. 241:247–255. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Torres-Collado AX and Iruela-Arispe ML:
Contribution of ADAMs and ADAMTSs to tumor expansion and
metastasis. Cancer Genome and Tumor Microenvironment.
Thomas-Tikhonenko Andrei: Springer; New York, NY: pp. 293–314.
2010, View Article : Google Scholar
|
|
30
|
Yu CC, Tsai LL, Wang ML, Yu CH, Lo WL,
Chang YC, Chiou GY, Chou MY and Chiou SH: miR145 targets the
SOX9/ADAM17 axis to inhibit tumor-initiating cells and
IL-6-mediated paracrine effects in head and neck cancer. Cancer
Res. 73:3425–3440. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mężyk-Kopeć R, Wyroba B, Stalińska K,
Próchnicki T, Wiatrowska K, Kilarski WW, Swartz MA and Bereta J:
ADAM17 promotes motility, invasion, and sprouting of lymphatic
endo-thelial cells. PLoS One. 10:e01326612015. View Article : Google Scholar
|
|
32
|
Saaristo A, Karpanen T and Alitalo K:
Mechanisms of angio-genesis and their use in the inhibition of
tumor growth and metastasis. Oncogene. 19:6122–6129. 2000.
View Article : Google Scholar
|
|
33
|
Jiang BH and Liu LZ: PI3K/PTEN signaling
in tumorigenesis and angiogenesis. Biochim Biophys Acta.
1784:150–158. 2008. View Article : Google Scholar
|
|
34
|
Gooz P, Gooz M, Baldys A and Hoffman S:
ADAM_17: A central regulator of angiogenesis. Matrix Biology.
25:S55–S56. 2006. View Article : Google Scholar
|
|
35
|
Klein T and Bischoff R: Active
metalloproteases of the A Disintegrin and Metalloprotease (ADAM)
family: Biological function and structure. J Proteome Res.
10:17–33. 2011. View Article : Google Scholar
|
|
36
|
Zheng X, Jiang F, Katakowski M, Lu Y and
Chopp M: ADAM17 promotes glioma cell malignant phenotype. Mol
Carcinog. 51:150–164. 2012. View Article : Google Scholar
|
|
37
|
Lv X, Li Y, Qian M, Ma C, Jing H, Wen Z
and Qian D: ADAM17 silencing suppresses the migration and invasion
of non-small cell lung cancer. Mol Med Rep. 9:1935–1940. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mcgowan PM, Ryan BM, Hill AD, Mcdermott E,
O’Higgins N and Duffy MJ: ADAM-17 expression in breast cancer
correlates with variables of tumor progression. Clin Cancer Res.
13:2335–2343. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xiang J, Fu X, Ran W and Wang Z: Grhl2
reduces invasion and migration through inhibition of TGFβ-induced
EMT in gastric cancer. Oncogenesis. 6:e2842017. View Article : Google Scholar
|
|
41
|
Wu K, Liao M, Liu B and Deng Z: ADAM-17
over-expression in gallbladder carcinoma correlates with poor
prognosis of patients. Med Oncol. 28:4752011. View Article : Google Scholar
|
|
42
|
McGowan PM, Mckiernan E, Bolster F, Ryan
BM, Hill AD, Mcdermott EW, Evoy D, O’Higgins N, Crown J and Duffy
MJ: ADAM-17 predicts adverse outcome in patients with breast
cancer. Ann Oncol. 19:1075–1081. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gooz M: ADAM-17: The enzyme that does it
all. Crit Rev Biochem Mol Biol. 45:146–169. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rose-John S: ADAM17, shedding, TACE as
therapeutic targets. Pharmacol Res. 71:19–22. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Aydin D, Bilici A, Yavuzer D, Kefeli U,
Tan A, Ercelep O, Mert A, Yuksel S, Ozcelik M, Isik D, et al:
Prognostic significance of ADAM17 expression in patients with
gastric cancer who underwent curative gastrectomy. Clin Transl
Oncol. 17:604–611. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kyula JN, Schaeybroeck SV, Doherty J,
Fenning CS, Longley DB and Johnston PG: Chemotherapy-induced
activation of ADAM-17: A novel mechanism of drug resistance in
colorectal cancer. Clin Cancer Res. 16:3378–3389. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hu YY, Zheng MH, Zhang R, Liang YM and Han
H: Notch signaling pathway and cancer metastasis. Adv Exp Med Biol.
727:186–198. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Du X, Cheng Z, Wang YH, Guo ZH, Zhang SQ,
Hu JK and Zhou ZG: Role of Notch signaling pathway in gastric
cancer: A meta-analysis of the literature. World J Gastroenterol.
20:9191–9199. 2014.PubMed/NCBI
|
|
49
|
Brzozowa M, Mielańczyk Ł, Michalski M,
Malinowski Ł, Kowalczyk-Ziomek G, Helewski K, Harabin-Słowińska M
and Wojnicz R: Role of Notch signaling pathway in gastric cancer
pathogenesis. Contemp Oncol (Pozn). 17:1–5. 2013.
|
|
50
|
Xi Y and Chen Y: Wnt signaling pathway:
Implications for therapy in lung cancer and bone metastasis. Cancer
Lett. 353:8–16. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Cai J, Guan H, Fang L, Yang Y, Zhu X, Yuan
J, Wu J and Li M: MicroRNA-374a activates Wnt/β-catenin signaling
to promote breast cancer metastasis. J Clin Invest. 123:566–579.
2013.PubMed/NCBI
|
|
52
|
Jang GB, Kim JY, Cho SD, Park KS, Jung JY,
Lee HY, Hong IS and Nam JS: Blockade of Wnt/β-catenin signaling
suppresses breast cancer metastasis by inhibiting CSC-like
phenotype. Sci Rep. 5:124652015. View Article : Google Scholar
|
|
53
|
Yanaka Y, Muramatsu T, Uetake H, Kozaki KI
and Inazawa J: miR-544a induces epithelial-mesenchymal transition
through the activation of WNT signaling pathway in gastric cancer.
Carcinogenesis. 36:1363–1371. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Tan C, Qiao F, Wei P, Chi Y, Wang W, Ni S,
Wang Q, Chen T, Sheng W, Du X and Wang L: DIXDC1 activates the Wnt
signaling pathway and promotes gastric cancer cell invasion and
metastasis. Mol Carcinog. 55:397–408. 2016. View Article : Google Scholar
|