Introduction
Recurrent miscarriage (RM) is a common reproductive
complication, which affects 1-3% of women during their reproductive
years (1). Recent studies on the
pathological factors of RM have made substantial progress and
immunological factors have drawn the attention of researchers.
Numerous studies have reported associations between immune cells
and RM, among which the role of macrophages has been confirmed.
Macrophages account for 20-25% of all decidual leukocytes at the
site of implantation, serving vital roles in regulating the
maternal immune microenvironment by linking the adaptive and innate
immune systems (2). It has
already been confirmed that aberrant macrophage infiltration at the
maternal-fetal interface is associated with RM (3-5).
Macrophages may be polarized into the M1/M2 subtype in specific
environments; M1 macrophages produce pro-inflammatory cytokines and
regulate inflammatory responses, whereas M2 macrophages promote
tissue remodeling and repair (6).
Early reports have indicated that decidual macrophages (dMΦ) may be
polarized into the M2 subtype in normal pregnancy (7), whereas those in pregnancy
complications, including RM or preeclampsia, polarize into the M1
subtype and mediate inflammatory responses (4,8).
Apoptosis is important for normal development of the
placenta and, in a normal pregnancy, apoptosis is observed in the
trophoblast layer of the placenta (9). Several studies have reported an
association between the deregulation of apoptosis and RM (10,11). The Fas/Fas ligand (FasL) system
resembles one of the major apoptotic pathways in tissues and cells.
The expression of FasL has been identified in several types of
immune cell, including activated CD8+ cells,
CD4+ T-helper type 1 cells, natural killer cells and
monocytes (12). A previous study
suggested that Fas/FasL-mediated cell death was associated with
cyclic changes in the bovine endometrium (13). Furthermore, excess macrophages in
the preeclamptic decidua may induce trophoblast apoptosis (14), and patients with RM have been
reported to have elevated expression of FasL (15-17). In addition, FasL can function as a
cytokine to induce apoptosis in susceptible cells in vitro
(18), and macrophages have been
reported to induce FasL-mediated apoptosis in a model of pulmonary
silicosis (19) and in chronic
demyelinated neuropathy (20).
Although higher incidences of trophoblast apoptosis and macrophage
infiltration have been detected in the decidua and placental
villous tissues from pregnancies in women with RM (3-5,10,11), the effect of macrophages on
trophoblast apoptosis, the involvement of FasL in this process, and
its potential role in the pathophysiology of RM remain to be fully
elucidated.
In the present study, the distribution pattern of
macrophages and the expression of FasL on macrophages were assessed
in clinical decidua samples. Furthermore, the role of FasL in the
process of macrophage-mediated trophoblast apoptosis was examined
in a macrophage and trophoblast co-culture model. The results
indicated that CD86+ macrophage populations in decidual
tissues were significantly increased, accompanied by the reduced
expression of CD163+ macrophages in RM and spontaneous
abortion groups of patients. Furthermore, the distribution of
CD68+ macrophages was significantly altered in RM, with
infiltration into the trophoblast cells. In addition, the elevated
expression of FasL on CD68+ and CD86+
macrophages was observed in the decidua of RM and spontaneous
abortion groups of patients, and FasL was reported to affect
trophoblast apoptosis mediated by co-culture with macrophages.
These results indicate that the aberration of macrophage-induced
FasL-mediated apoptosis may represent one of the causes of
spontaneous abortion, even for RM, suggesting that the unfortunate
pregnancy outcomes of these two groups may partly result from the
same causes.
Patients and methods
Patients and tissue samples
Women attending the Reproductive Center of Renmin
Hospital of Wuhan University (Wuhan, China) were enrolled in the
present study between January 2016 and January 2019. A total of 81
patients at between 7-9 weeks of gestation were enrolled, including
21 RM patients (RM group), 34 women undergoing electively induced
abortion of a pregnancy (control group) and 26 women who underwent
spontaneous abortion (abortion group). The inclusion criteria were
an age of <35 years, a normal body mass index (BMI) and a
regular menstrual cycle. Women with any endometrial or uterine
pathology, endometriosis genetic abnormalities or anovulation were
excluded. RM was defined as clinically spontaneous abortion
occurring prior to 20 weeks of gestation on at least two occasions.
Decidual and placental villous tissues were washed in PBS and
collected soon following curettage under sterile conditions, and
were divided into two aliquots. One aliquot was fixed in 10%
formaldehyde for terminal deoxynucleotidyl transferase deoxyuridine
triphosphate nick end labelling (TUNEL), immunohistochemistry and
immunofluorescence, and the other was stored at −80°C for western
blot analysis. All samples were collected following the provision
of informed consent from the patients, and all associated
procedures were performed with the approval of the internal Review
and Ethics Boards of Renmin Hospital of Wuhan University.
Cell culture
The human THP-1 peripheral blood acute mono-cytic
leukemia cell line was obtained from the Institute of Biochemistry
and Cell Biology, Chinese Academy of Sciences (Shanghai, China).
The THP-1 cells were grown in RPMI-1640 medium (Hyclone; GE
Healthcare Life Sciences, Logan, UT, USA) supplemented with 10%
fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), 100 U/ml penicillin and 100 mg/ml streptomycin
in an atmosphere containing 5% CO2 at 37°C. The
HTR-8/SVneo cell line, derived from human extravillous
trophoblasts, was obtained from the China Center for Type Culture
Collection (Wuhan, China). These cells were cultured in Dulbecco's
modified Eagle's medium (DMEM; high glucose; Hyclone; GE Healthcare
Life Sciences) supplemented with 10% FBS and antibiotics.
Macrophage polarization and co-culture
model
The THP-1 cells were differentiated into macrophages
by incubation with 50 ng/ml phorbol 12-myristate 13-acetate (PMA;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for 48 h.
Macrophages were confirmed using an immunofluorescence assay with
primary antibody against CD68 (Novus Biologicals, LLC, Littleton,
CO, USA), and five fields (magnification, ×400) were selected for
analysis using ImageJ software (version 1.46; National Institutes
of Health, Bethesda, MD, USA). A trophoblast and macrophage
co-culture model was established to determine the effect of
macrophage-derived cytokines on the apoptosis of
trophoblast-derived cells. In brief, 1.5×105 HTR-8/SVneo
cells in 2 ml DMEM containing 10% FBS were seeded into the lower
chambers and 5×105 differentiated macrophages (M0 in 1
ml RPMI-1640 with 10% FBS) were placed into the upper chambers of a
Transwell plate (Corning Incorporated, Corning, NY, USA), and
cultured for 24 h at 37°C for cell attachment. Subsequently, the
upper chambers containing M0 macrophages were placed on top of the
lower chambers containing HTR-8/SVneo, followed by incubation at
37°C for another 48 h.
Cell apoptosis assessment
Cell apoptosis was assessed using a fluorescein
isothiocyanate (FITC)-Annexin V Apoptosis Detection kit (Dojindo
Molecular Technologies, Inc., Kumamoto, Japan) and flow cytometry
using FACS Canto II (BD Biosciences, Franklin Lakes, NJ, USA). In
brief, the trophoblasts were collected and stained with propidium
iodide/FITC-Annexin V for 15 min at 4°C in the dark, followed by
the addition of binding buffer and analysis by flow cytometry
within 10 min. The data were analyzed using the FlowJo VX10
software (Tree Star, Inc., Ashland, OR, USA). All experiments were
performed in triplicate.
Western blot analysis
The cells and tissues were lysed with lysis buffer
containing 1 mM phenyl methane sulfonyl fluoride (both from
Beyotime Institute of Biotechnology, Haimen, China) and protease
and phosphatase inhibitors (21).
Total protein was extracted from the supernatants of the cell
lysates following incubation in lysis buffer for 30 min on ice,
followed by centrifugation at 4°C for 15 min at 13,000 g. The
supernatant was collected and the protein concentrations were
quantified using a bicinchoninic acid protein assay kit (Beyotime
Institute of Biotechnology). Equal quantities of protein (40-60
µg) were separated by 10% SDS-PAGE and then transferred onto
polyvinylidene difluoride membranes (EMD Millipore, Bedford, MA,
USA) at 200 mA. The membranes were blocked in 5% low-fat milk for 1
h at room temperature and incubated overnight at 4°C with the
following primary antibodies: Rabbit anti-B-cell lymphoma 2
(Bcl-2)-associated X protein (Bax; cat. no. 2774; 1:1,000
dilution), rabbit anti-Bcl-2 (cat. no. 2876; 1:1,000 dilution; both
from Cell Signaling Technology, Inc., Danvers, MA, USA) and mouse
anti-GAPDH (cat. no. 60004-1-Ig; 1:5,000 dilution; ProteinTech
Group, Inc., Chicago, IL, USA). Following incubation, the membranes
were washed with PBS with 0.1% Tween-20 (PBST) and incubated with
horseradish peroxidase-conjugated secondary antibodies (cat. no.
SA00001-1 and SA00001-2; 1:5,000 dilution; goat anti-mouse or goat
anti-rabbit; ProteinTech Group, Inc.) for 1 h at 37°C. The proteins
were detected using a Chemiluminescence Western Detection system
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) and visualized
using autoradiography. Densitometric analysis of band intensities
was performed using ImageJ software (version 1.46).
Immunohistochemistry
Immunohistochemical staining was performed as
previously described (22). In
brief, formalin-fixed, paraffin-embedded decidual and placental
villous tissues were cut into 4-µm sections, dewaxed in
xylene and rehydrated with an ethanol series of descending grades
followed by PBS. Subsequently, the samples were incubated in 3%
hydrogen peroxide for 15 min at room temperature to block the
endogenous peroxidase activity. Antigen retrieval was performed by
heating in a microwave oven with sodium citrate buffer. The
sections were then incubated in 5% bovine serum albumin
(Sigma-Aldrich; Merck KGaA) for 20 min at room temperature to block
any non-specific binding, followed by incubation with primary
antibodies against CD68 (cat. no. NB100-683; 1:100 dilution; Novus
Biologicals, LLC), CD86 (cat. no. ab53004; 1:100 dilution), CD163
(cat. no. ab87099; 1:50 dilution; both Abcam, Cambridge, MA, USA),
Bax (1:400 dilution) and Bcl-2 (1:400 dilution; both Cell Signaling
Technology, Inc.) at 37°C for 1 h. The sections were then washed
and incubated with goat anti-mouse/rabbit secondary antibody
(1:2,000 dilution; ProteinTech Group, Inc.) for 30 min at 37°C.
Antibody binding was identified as a brown precipitate following
staining with 3,3-diaminoben-zidine tetrahydrochloride (Dako;
Agilent Technologies, Inc., Santa Clara, CA, USA). Images were
acquired using an optical microscope at a magnification of ×400.
Five visual fields with tissue occupying >80% of the area were
selected for analysis and the number of immuno-positive cells was
calculated using ImageJ software (version 1.46).
Immunofluorescence
Double immunofluorescence was performed to evaluate
the expression of FasL on CD68+ and CD86+
macrophages and the distribution of CD68+ macrophages
and CK7+ trophoblasts. In brief, the sections were
incubated with primary antibodies against CD68 (1:200 dilution;
Novus Biologicals, LLC) or CD86 (1:500 dilution; Abcam) overnight
at 4°C, followed by incubation with cyanine-3 (Cy3)-conjugated goat
anti-mouse antibody (cat. no. BA1031; 1:100 dilution; Boster
Biological Technology, Wuhan, China) for 30 min at 37°C, followed
by incubation with bovine serum albumin for 30 min at room
temperature and then with primary antibodies against FasL (cat. no.
sc-33716; 1:100 dilution; Santa Cruz Biotechnology, Inc., Dallas,
TX, USA) or CK7 (cat. no. 17513-1-AP; 1:100 dilution; ProteinTech
Group, Inc.) at 4°C overnight. The sections were then incubated
with FITC-conjugated goat anti-rabbit antibody (cat. no. BA1105;
1:100 dilution; Boster Biological Technology) for 30 min at 37°C.
The sections were washed with TBST and visualized under a
fluorescence microscope at a magnification of ×400. Cy3 was
detected as red and FITC as green fluorescence, and the blue
fluorescence signal represented the nuclear DNA stained by
4′,6-diamidino-2-phenylindole (DAPI). Five random fields from
stained sections were selected. The proportion of CD68+
and CD86+ macrophages with FasL was calculated as the
number of CD68+FasL+ or
CD86+FasL+ cells divided by the number of
CD68+ or CD86+ cells in the field using
ImageJ software (version 1.46).
TUNEL
Apoptosis in the decidual and placental villous
tissues was detected using an in situ TUNEL detection kit
(Promega Corporation, Madison, WI, USA). In brief, the paraffinized
sections were dewaxed in xylene, rehydrated and washed in PBS, and
then stained according to the manufacturer's protocol. The nuclei
were visualized with 0.1 µg/ml DAPI (Beyotime Institute of
Biotechnology) in PBS at 30°C for 30 min. Cells containing the
characteristic fragmented nuclear chromatin and brown nuclear
staining were considered to be apoptotic. The apoptotic index was
calculated as follows: Apoptotic index = (number of TUNEL -
positive nuclei)/(number of total nuclei) × 100%. Images were
captured with a fluorescence microscope at a magnification of ×400
and analyzed using ImageJ software (version 1.46).
Flow cytometry
The THP-1 cells stimulated with PMA or PBS were
stained with anti-FasL antibody to determine the expression of FasL
on macrophages. In brief, THP-1 cells were collected following
stimulation with PMA or PBS, centrifuged at 300 × g for 10
min at room temperature and resuspended in PBS. The cells
(1×106/tube) were stained with primary antibody against
FasL (cat. no. sc-33716; 1:100 dilution; Santa Cruz Biotechnology,
Inc.) overnight at 4°C, and then incubated with FITC-conjugated
Affinipure goat anti-rabbit immunoglobulin G (H+L; cat. no.
SA00003-2; 1:100 dilution; ProteinTech Group, Inc.) for 1 h at
37°C. Finally, the fluorescence of the cells was analyzed by flow
cytometry (FACS Canto II; BD Biosciences).
Statistical analysis
Values were expressed as the mean ± standard error
of the mean. Statistical analyses were performed using SPSS 19.0
(IBM Corp., Armonk, NY, USA) using Student's t-test or one-way
analysis of variance with Bonferroni's post hoc test. The clinical
characteristics were compared between the normal and RM patients
using the Kruskal-Wallis test. The experiments were repeated at
least three times for each group. P<0.05 was considered to
indicate a statistically significant difference.
Results
Baseline characteristics of the control,
abortion and RM groups
To examine the expression and distribution of
macrophages in spontaneous abortion and in miscarriages in terms of
recurrence, three groups were included in the present study. The
clinical characteristics of the recruited patients are listed in
Table I. No significant
differences were observed among the three groups in terms of
maternal age, BMI and gestational age. However, the number of
miscarriages in the RM group and abortion group were significantly
higher and the numbers of live births were significantly lower
compared with those in the control group (P<0.01 for both).
 | Table IDemographic and clinical
characteristics of the study population. |
Table I
Demographic and clinical
characteristics of the study population.
|
Characteristics | Control group
(n=34) | Abortion group
(n=26) | RM group
(n=21) |
|---|
| Age (years) | 29.86±4.03 | 28.98±3.59 | 30.08±4.23 |
| BMI
(kg/m2) | 21.38±3.13 | 22.09±2.68 | 21.28±3.52 |
| Gestation age
(weeks) | 8.49±1.45 | 8.43±1.28 | 8.33±1.78 |
| No. of
miscarriages | 0.65±0.12 | 1.43±0.25a | 2.63±0.41a |
| No. of live
births | 1.51±0.34 | 0.96±0.23a | 0.00±0.00a |
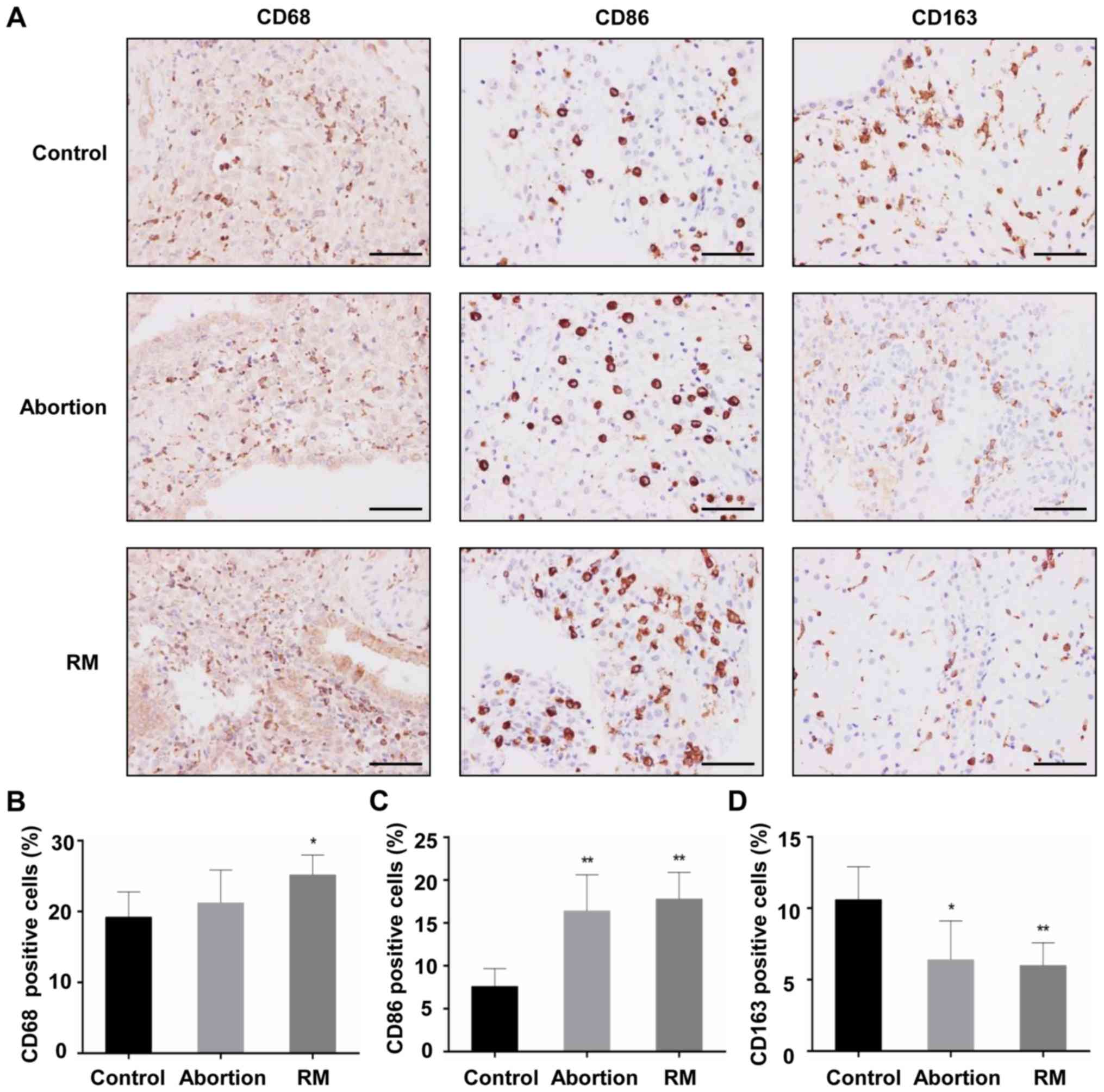
Comparison of macrophage population
between the control, abortion and RM groups
The decidual tissues from the three groups were
found to contain macrophages that were stained with the primary
antibody against CD68. The majority of the CD68+
macrophages from subjects with RM were distributed in the cytoplasm
of decidual stromal cells and certain macrophages dispersed around
blood vessels and glands, and the localization of CD68+
macrophages in the decidua of the control subjects and abortion
group was similar to that of the subjects from the RM group
(Fig. 1A). Cells positive for
CD163 (an M2 marker) and CD86 (an M1 marker) were apparent in the
three groups, mainly in the cytoplasm of stromal cells (Fig. 1A). In the RM patients, the
CD68+ macrophage population in the decidual tissues was
significantly increased, accompanied by an increased
CD86+ macrophage population (Fig. 1B and C) and a reduced
CD163+ macrophage population (Fig. 1D), compared with the control
group. In the abortion group, the CD68+ macrophage
population did not differ significantly, and the expression levels
of the CD86+ and CD163+ macrophage
populations were similar to those in the RM group (Fig. 1C). These results indicated that
increased macrophage infiltration and an aberrant macrophage
population in the decidua may be associated with miscarriage.
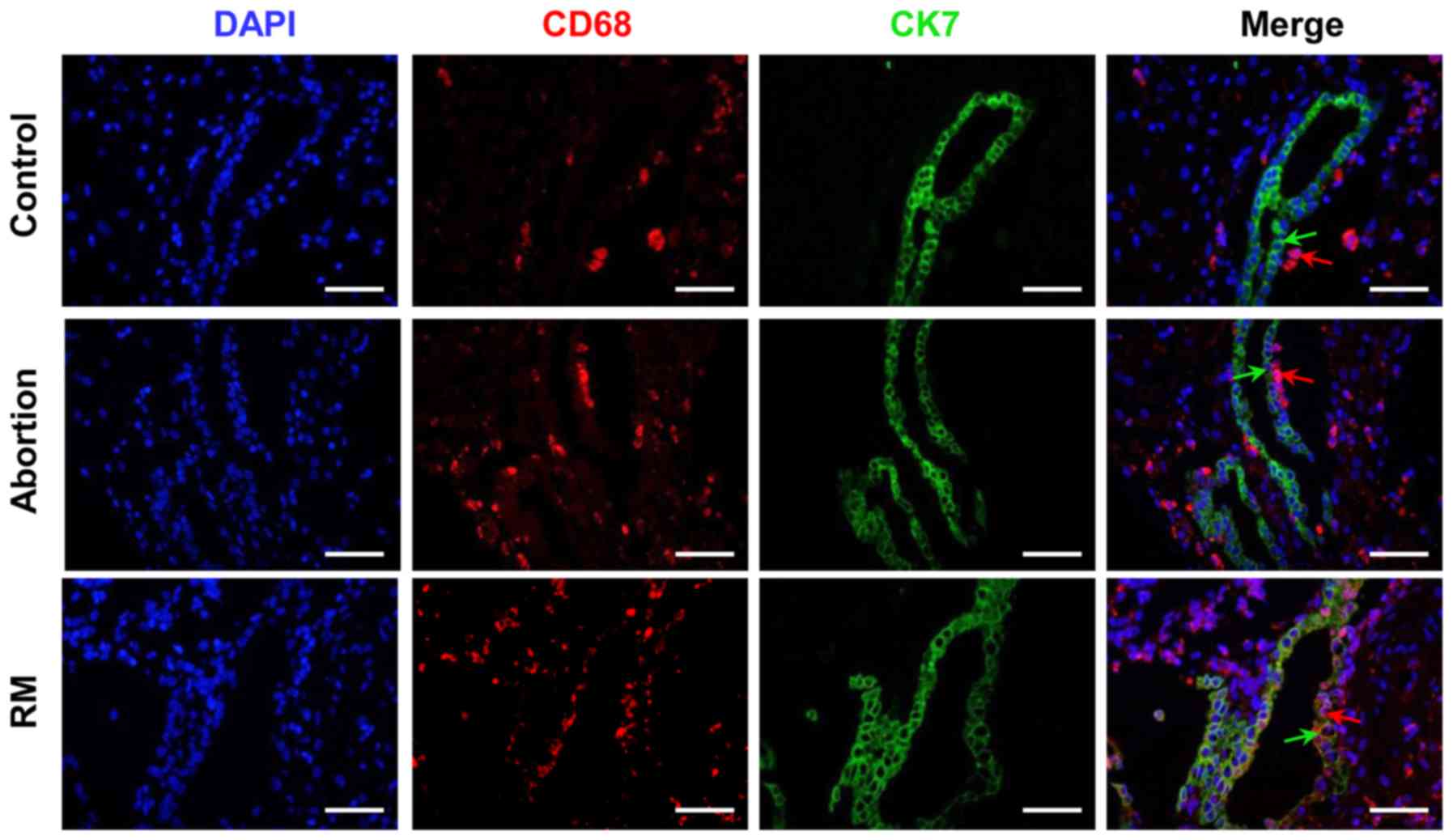
Comparison of macrophage distribution
among the control, abortion and RM groups
In the present study, CK7 was selected as a
trophoblast marker. As presented in Fig. 2, the immunofluorescence analysis
indicated that the number of CD68+ cells was
significantly higher in the RM patients compared with that in
control subjects, which is in accordance with the results of the
immunohistochemical analysis (Fig. 1A
and B). Furthermore, CD68+ macrophages were located
in the periphery of the trophoblast cells from the subjects with
normal pregnancies, whereas in the decidual tissues of abortion and
RM patients, significant infiltration of CD68+
macrophages into CK7+ trophoblast cells was observed
(Fig. 2). These results
demonstrated that the macrophage distribution was significantly
altered in the women in the abortion and RM groups.
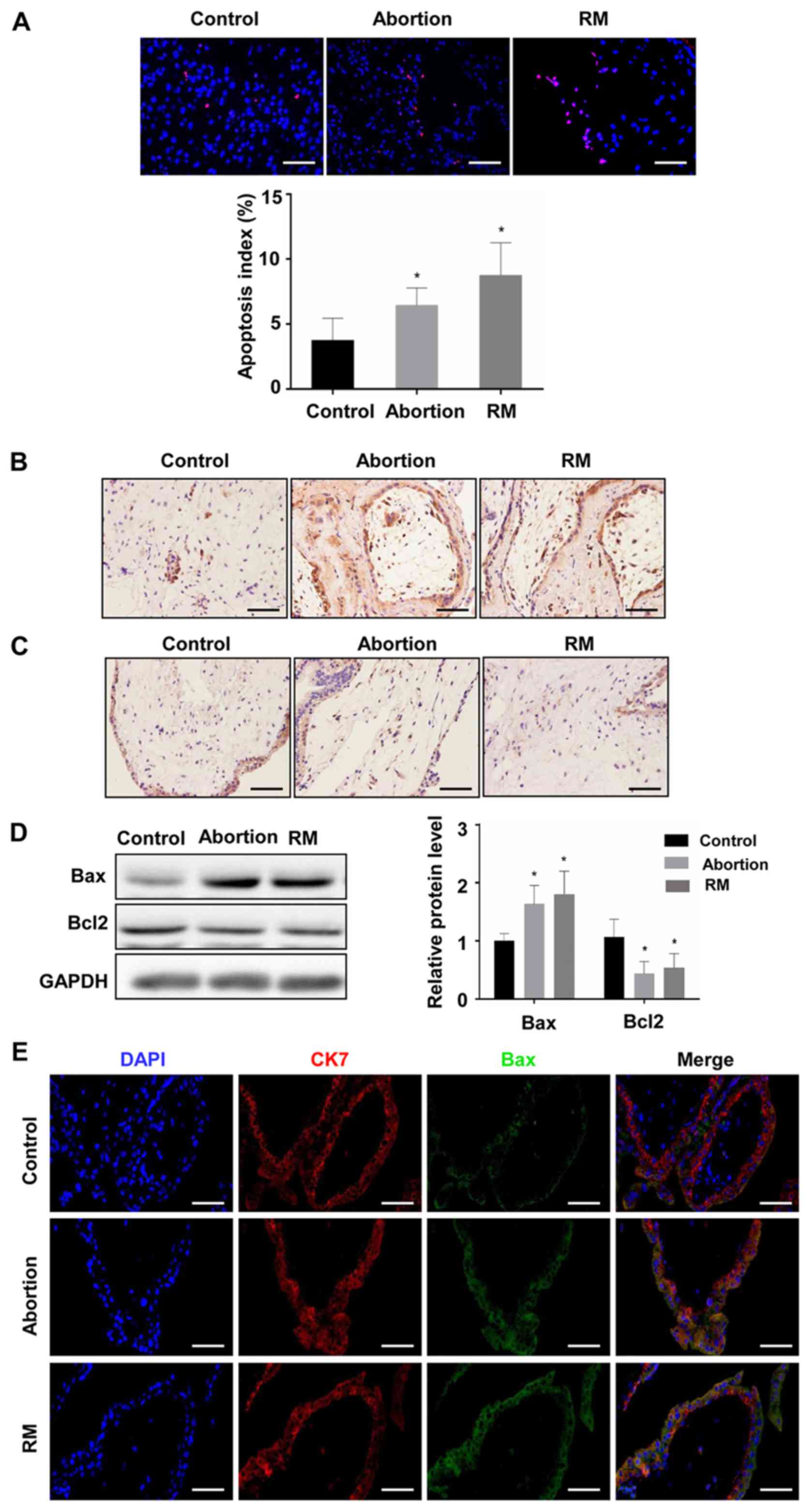
Comparison of apoptosis among the
control, abortion and RM groups
To investigate the association between apoptosis and
fetal loss, TUNEL was used to detect cell apoptosis and western
blot analysis was used to determine the expression of
apoptosis-promoting protein Bax and the anti-apoptotic protein
Bcl-2 in the decidual and placental villous tissues. The TUNEL
assay revealed that the proportion of cells with brown nuclear
staining (apoptotic cells) and the associated apoptotic index was
significantly higher in the abortion and RM groups compared with
that in the control group (Fig. 3A
and B), indicating that apoptosis in the decidua from the
abortion and RM patients was higher than that from the controls.
Furthermore, the results of the western blot analysis revealed that
the protein level of Bcl-2 was reduced and the protein level of Bax
was increased compared with levels in the control group, which was
further confirmed by the immunohistochemical and immunofluorescence
staining (Fig. 3C-F). As
expected, similar results were observed in the placental villous
tissues (Fig. 4A-E). These
results demonstrated that elevated apoptosis in the decidual and
placental villous tissues is associated with fetal loss.
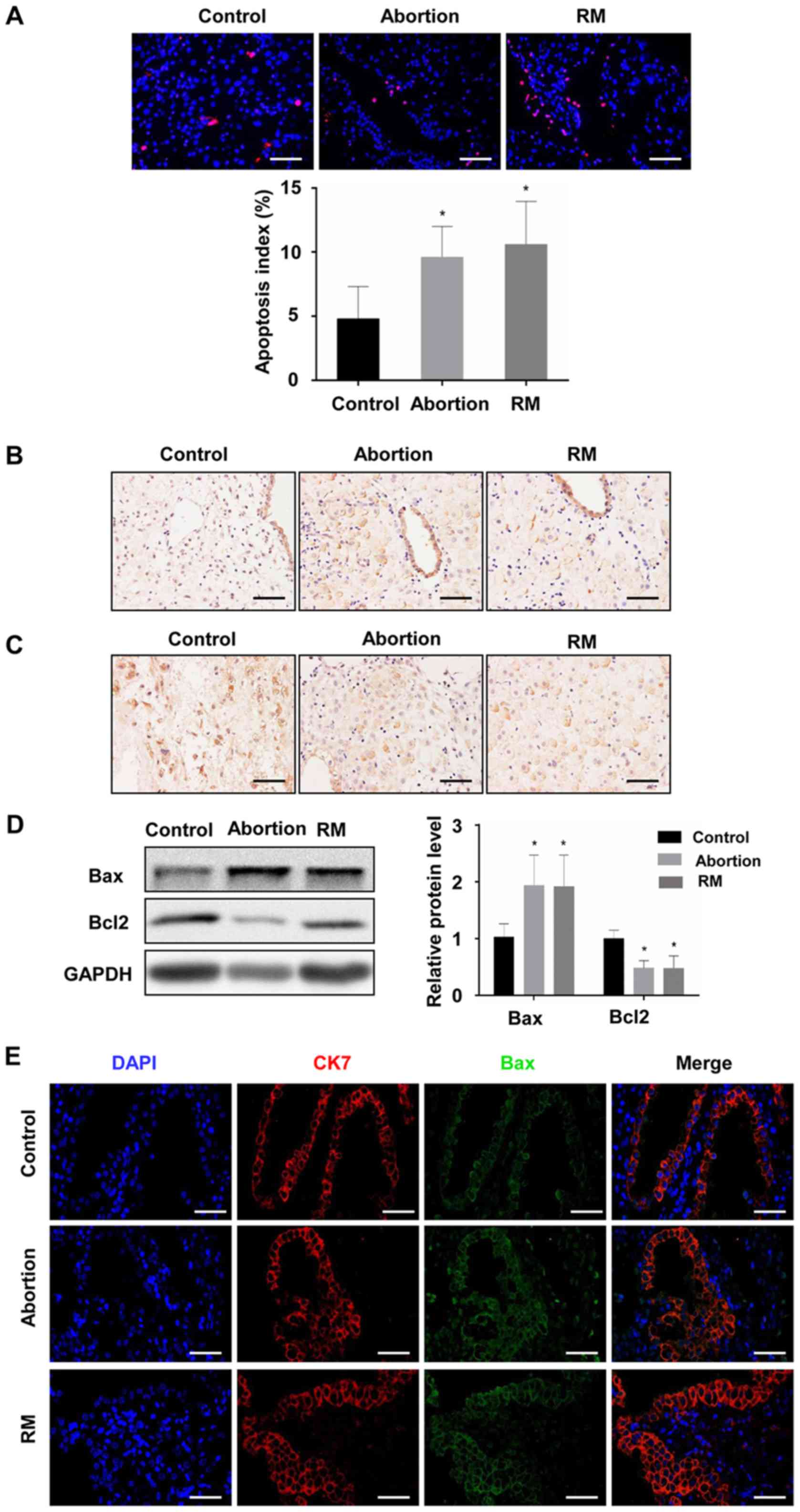 | Figure 3Comparison of levels of apoptosis in
decidual tissues among the control, abortion and RM groups. (A)
Apoptosis was determined by terminal deoxynucleotidyl transferase
deoxyuridine triphosphate nick end labelling in decidual tissues
from women in the normal pregnancy (n=34), abortion (n=26) and RM
(n=21) groups, respectively. Distributions of (B) Bax and (C) Bcl-2
in decidual tissues were detected by immunohistochemistry
(magnification, ×400). (D) Protein levels of Bax and Bcl-2 in
decidual tissues were measured by western blotting. (E) The
expression of Bax on trophoblasts was analyzed by dual
immunofluorescence (magnification, ×400). (F) The expression of
Bcl-2 on trophoblasts was analyzed by dual immunofluorescence
(magnification, ×400). Scale bars, 100 µm.
*P<0.05 vs. the control group. RM, recurrent
miscarriage; Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-associated X
protein; DAPI, 4′,6-diamidino-2-phenylindole. |
 | Figure 4Comparison of levels of apoptosis in
the placental villous tissues among control, abortion and RM
groups. (A) Apoptosis was determined by terminal deoxynucleotidyl
transferase deoxyuridine triphosphate nick end labelling in
placental villous tissues from women in the normal pregnancy
(n=34), abortion (n=26) and RM (n=21) groups, respectively.
Distributions of (B) Bax and (C) Bcl-2 in placental villous tissue
were detected by immunohistochemistry (magnification, ×400). (D)
Protein levels of Bax and Bcl-2 in placental villous tissues were
measured by western blotting. (E) The expression of Bax on
trophoblasts was analyzed by dual immunofluorescence
(magnification, ×400). (F) The expression of Bcl-2 on trophoblasts
was analyzed by dual immunofluorescence (magnification, ×400).
Scale bars, 100 µm. *P<0.05 vs. the control
group. RM, recurrent miscarriage; Bcl-2, B-cell lymphoma 2; Bax,
Bcl-2-associated X protein; DAPI,
4′,6-diamidino-2-phenylindole. |
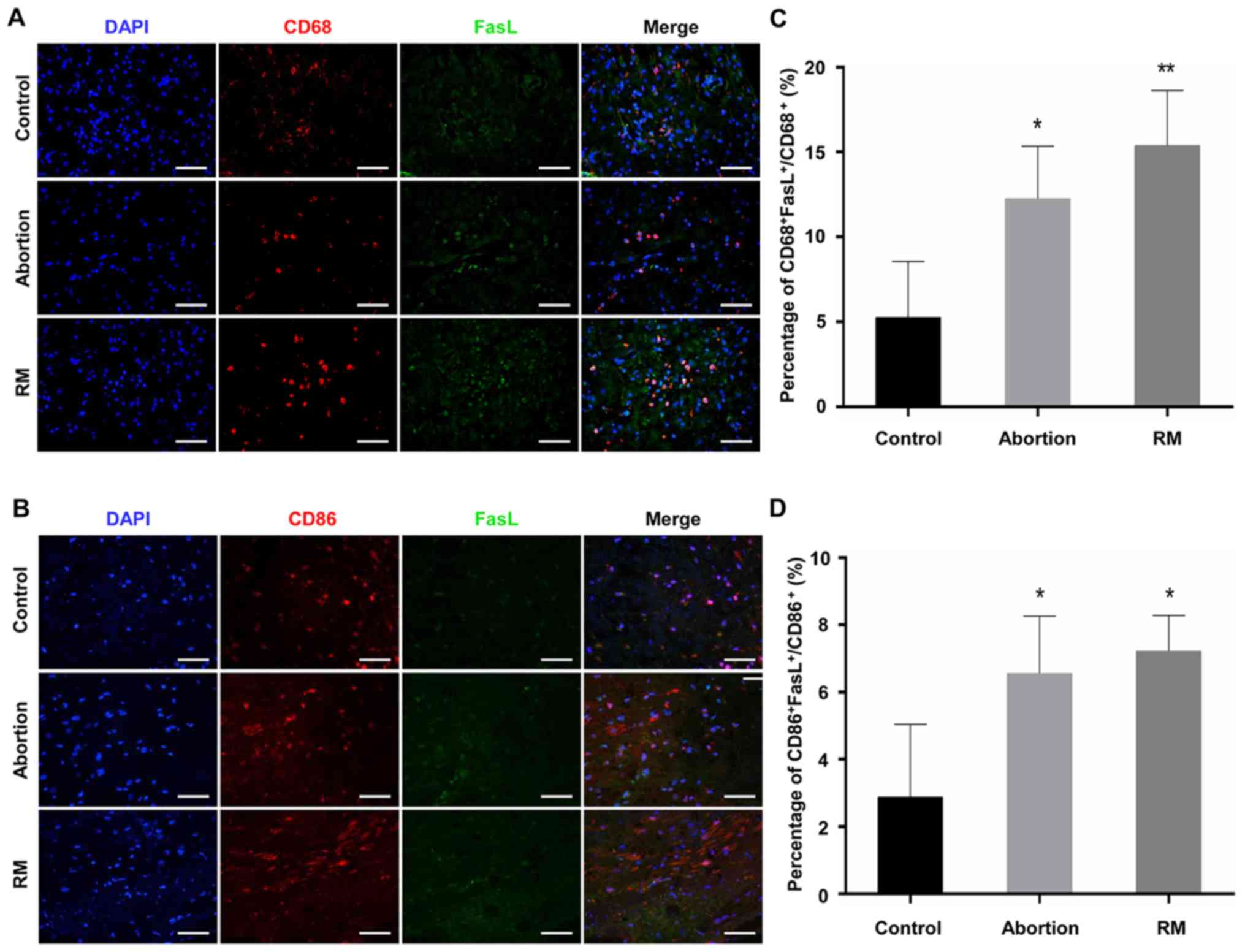
Comparison of the expression levels of
FasL on macrophages among the control, abortion and RM groups
As FasL is able to induce apoptosis (18), the expression of FasL has been
previously detected in macrophages, with patients with RM
exhibiting elevated expression of FasL (15-17). The present study performed dual
immunofluorescence analysis of CD68 and FasL to identify the
expression of FasL on macrophages in the decidua. The results
(Fig. 5A-D) indicated that
CD68+ dMΦ expressed minimal FasL protein, with only a
weak signal in normal women; by contrast, the decidua was rich in
FasL+ macrophages in the RM patients (Fig. 5A and C). In addition, the
expression of FasL on CD86+ macrophages was detected by
dual immunofluorescence. The proportion of
CD86+FasL+ macrophages in the abortion and RM
groups were significantly higher than that in the control group
(Fig. 5B and D). These results
demonstrated that the abnormal expression of FasL on dMΦ may be
associated with miscarriage, and even RM.
Effect of macrophages on the apoptosis of
trophoblast cells
As significantly increased macrophage infiltration
was observed in the abortion and RM patients (Fig. 1A and B), and as macrophages are
able to induce apoptosis (23), a
tropho-blast and macrophage co-culture model was used to determine
whether macrophages induce apoptosis in trophoblasts. As presented
in Fig. 6A, both the inactivated
and activated THP-1 cells (M0) expressed CD68, although the
expression of CD68 in the activated THP-1 cells was increased
(Fig. 6A). In addition, the
expression level of FasL was increased in the M0 macrophages
(Fig. 6B and C), and the
co-culture model demonstrated that M0 macrophages promoted the
apoptosis of trophoblast-derived cells (Fig. 6D and E). In order to verify the
role of FasL in macrophage-induced trophoblast apoptosis, an
intervention experiment was performed, in which anti-FasL blocking
antibody or isotype control antibody (R&D Systems, Inc.,
Minneapolis, MN, USA) was added to the co-culture model.
Macrophages induced significant cell apoptosis, whereas the
addition of anti-FasL blocking antibody significantly inhibited
this effect (Fig. 6D and E).
These results indicated that macrophages mediate the apoptosis of
trophoblast cells through FasL-dependent pathways.
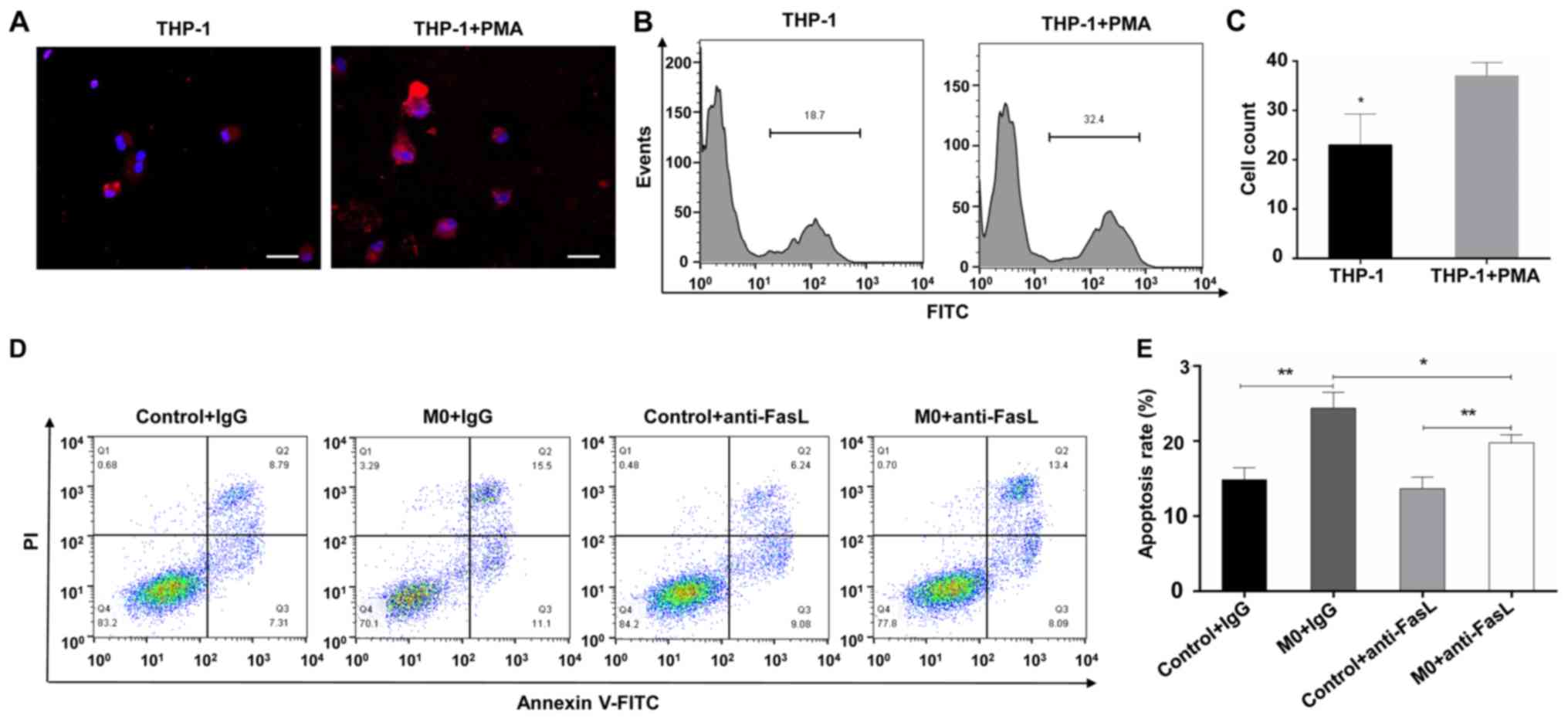 | Figure 6Effect of macrophages on the
apoptosis of trophoblast cells. (A) Trophoblast and macrophage
co-culture model to determine whether macrophage induce the
apoptosis of trophoblasts. Immunofluorescence staining of CD68 was
performed in THP-1 cells stimulated with PMA or PBS, DAPI staining
(blue) was performed to visualize cell nuclei (magnification,
×400). (B) Flow cytometry and (C) quantification of the expression
of FasL on THP-1 cells stimulated with PMA or PBS. Trophoblast
cells were co-cultured with macrophages in the presence of
anti-FasL antibody or isotype control. (D) Flow cytometry and (E)
quantification of apoptosis of the HTR-8/SVneo cell line. Scale
bars, 100 µm; *P<0.05; **P<0.01.
PMA, phorbol 12-myristate 13-acetate; PI, propidium iodide; FITC,
fluorescein isothiocyanate; DAPI, 4′,6-diamidino-2-phenylindole;
M0, differentiated macrophages; FasL, Fas ligand. |
Discussion
It is generally considered that macrophages are
essential for the establishment and maintenance of pregnancy, as
they are involved in diverse processes, including blood vessel
remodeling, immune tolerance, immunomodulation of maternal decidual
lymphocytes and parturition initiation (24-27). Comprising 20-25% of the decidual
leukocyte population in early pregnancy (2), dMΦ are involved in vascular
remodeling, inducing the apoptosis of damaged cells, removal of
apoptotic cell debris and the elimination of invading pathogens
(28). A previous study reported
that dMΦ may be polarized to the M2 subtype to exert important
roles in maintaining immune tolerance in early pregnancy (7), whereas macrophages polarized into
the M1 subtype may promote RM (8). In the present study, CD68 (a marker
for macrophages), CD163 (M2 marker) and CD86 (M1 marker) were used
to characterize macrophage populations in the decidua, and the
results indicated that M1 macrophages were significantly higher and
M2 macrophages were decreased in the RM and abortion groups
compared with those in the control group, which was in accordance
with the results of earlier studies (3-5,8).
During pregnancy, the invading trophoblast cell is in close contact
with maternal immune cells, including T lymphocytes, natural killer
cells and macrophages (29). A
previous study reported that macrophages significantly infiltrated
the trophoblasts in spontaneous miscarriage (8), although the distribution of
macrophages in RM was unclear. In the present study, dual
immunofluorescence staining against CD68 and CK7 revealed the
presence of macrophage infiltration into trophoblast cells in the
RM group. These results indicated that aberrant infiltration and
the distribution of dMΦ may be associated with the unfortunate
pregnancy outcomes of RM and abortion in women.
During pregnancy, the turnover of placenta depends
on the functional loss of trophoblast cells by apoptosis, and the
trophoblast cells are removed and replaced with a younger
population in this physiological process (30,31); however, decidual apoptosis may
result in a series of cellular dysfunctions, which may impair the
course of pregnancy and lead to RM (32). Several studies have reported on
the association between apoptosis deregulation and RM (10,11). In the present study, the apoptosis
of decidual and placental villous tissues was confirmed by TUNEL
staining. The results demonstrated a significant increase in the
apoptotic index in the decidual and placental villous tissues of
the abortion and RM groups, accompanied by decreased protein levels
of Bcl-2 and increased protein levels of Bax, compared with those
in the control group, which was in accordance with previous studies
(33,34). These results suggest that aberrant
apoptosis in decidual and placental villous tissues may be a cause
of embryo loss.
The Fas/FasL system is one of the major apoptotic
pathways in cells and tissues, and the binding of the Fas receptor
by FasL activates a cascade of intracellular proteolytic enzymes,
leading to the apoptosis of Fas-bearing cells (35). A previous study indicated that
excess macrophages in the preeclamptic decidua induced trophoblast
apoptosis (14), and patients
with RM exhibit higher expression of FasL (15-17). In addition, FasL is known to
induce apoptosis (18), and
macrophages induced FasL-mediated apoptosis in a model of pulmonary
silicosis (19) and chronic
demyelinated neuropathy (20). A
previous study by Guenther et al (8) suggested the potential role of FasL
on macrophages in trophoblast apoptosis in spontaneous miscarriage;
however, the role of aberrant expression of FasL on macrophages in
miscarriages, in terms of recurrence, and its exact effect on the
apoptosis of trophoblast cells remain to be fully elucidated. In
the present study, double immunofluorescence staining for CD68 and
FasL was performed in the decidua. The results indicated that dMΦ
became strongly positive for FasL, a ligand which induces apoptosis
in Fas-expressing extravillous trophoblasts (17). The co-localization of CD68 and
FasL suggested that macrophages may contribute to the production
and/or release of FasL. In addition, the co-culture experiment
revealed that macrophages promoted the apoptosis of trophoblast
cells, which was consistent with a study by Guirelli et al
(23), and this effect was
reversed by FasL neutralization antibody, which was in agreement
with previous studies (19,20), supporting the hypothesis that
macrophages induce the apoptosis of trophoblast cells through the
release of FasL (23).
In conclusion, the results of the present study
indicate that abnormal macrophage infiltration and distribution, in
addition to the aberrant expression of FasL on dMΦ are associated
with spontaneous abortion and RM. In vitro assays further
demonstrated that macrophages induced apoptosis in trophoblast
cells by FasL mediation. Therefore, it is possible to conclude that
the aberration of macrophage-induced, FasL-mediated apoptosis may
represent one of the mechanisms leading to spontaneous abortion and
RM. However, it was not possible to conclude whether the abnormal
macrophage profile and trophoblast apoptosis were a cause or a
result of the miscarriage process. In addition, flow cytometric
analysis provides an effective way of precisely detecting FasL on
dMΦ in single cell suspensions made from decidua, and this warrants
further investigation. Additional investigations are also required
to clarify the specific effect of FasL on the interaction between
macrophages and trophoblasts.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81771618, 81771662
and 81701412) and the National Key Research and Development Program
of China (grant nos. 2016YFC1000600 and 2018YFC1002804).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YC and JY were involved in study design and revision
of the manuscript. JD and TY wrote the manuscript. JD, TY and NY
performed the experiments. JD, TY and NY analyzed and interpreted
the data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Written informed consent was obtained from all the
patients and the research protocols were approved by the Ethics
Committee of Renmin Hospital of Wuhan University.
Patient consent for publication
Informed consent was obtained from the subjects
participating in the present study.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
RM
|
recurrent miscarriage
|
|
FasL
|
Fas ligand
|
|
PMA
|
phorbol 12-myristate 13-acetate
|
|
dMΦ
|
decidual macrophage
|
|
BMI
|
body mass index
|
|
FBS
|
fetal bovine serum
|
|
DAPI
|
4′,6-diamidino-2-phenylindole
|
Acknowledgments
Not applicable.
References
|
1
|
Rai R and Regan L: Recurrent miscarriage.
Lancet. 368:601–611. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Heikkinen J, Möttönen M, Komi J, Alanen A
and Lassila O: Phenotypic characterization of human decidual
macrophages. Clin Exp Immunol. 131:498–505. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kolben TM, Rogatsch E, Vattai A, Hester A,
Kuhn C, Schmoeckel E, Mahner S, Jeschke U and Kolben T: PPARγ
expression is diminished in macrophages of recurrent miscarriage
placentas. Int J Mol Sci. 19:18722018. View Article : Google Scholar
|
|
4
|
Tsao FY, Wu MY, Chang YL, Wu CT and Ho HN:
M1 macrophages decrease in the deciduae from normal pregnancies but
not from spontaneous abortions or unexplained recurrent spontaneous
abortions. J Formos Med Assoc. 117:204–211. 2018. View Article : Google Scholar
|
|
5
|
Quack KC, Vassiliadou N, Pudney J,
Anderson DJ and Hill JA: Leukocyte activation in the decidua of
chromosomally normal and abnormal fetuses from women with recurrent
abortion. Hum Reprod. 16:949–955. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Colin S, Chinetti-Gbaguidi G and Staels B:
Macrophage phenotypes in atherosclerosis. Immunol Rev. 262:153–166.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gustafsson C, Mjösberg J, Matussek A,
Geffers R, Matthiesen L, Berg G, Sharma S, Buer J and Ernerudh J:
Gene expression profiling of human decidual macrophages: Evidence
for immunosuppressive phenotype. PLoS One. 3:e20782008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guenther S, Vrekoussis T, Heublein S,
Bayer B, Anz D, Knabl J, Navrozoglou I, Dian D, Friese K,
Makrigiannakis A and Jeschke U: Decidual macrophages are
significantly increased in spontaneous miscarriages and overexpress
FasL: A potential role for macrophages in trophoblast apoptosis.
Int J Mol Sci. 13:9069–9080. 2012. View Article : Google Scholar :
|
|
9
|
Jerzak M and Bischof P: Apoptosis in the
first trimester human placenta: The role in maintaining immune
privilege at the maternal-foetal interface and in the trophoblast
remodelling. Eur J Obstet Gynecol Reprod Biol. 100:138–142. 2002.
View Article : Google Scholar
|
|
10
|
Abrahams VM, Kim YM, Straszewski SL,
Romero R and Mor G: Macrophages and apoptotic cell clearance during
pregnancy. Am J Reprod Immunol. 51:275–282. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Crocker IP, Cooper S, Ong SC and Baker PN:
Differences in apoptotic susceptibility of cytotrophoblasts and
syncytiotropho-blasts in normal pregnancy to those complicated with
preeclampsia and intrauterine growth restriction. Am J Pathol.
162:637–643. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lettau M, Paulsen M, Kabelitz D and
Janssen O: FasL expression and reverse signalling. Results Probl
Cell Differ. 49:49–61. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Arai M, Yoshioka S, Nishimura R and Okuda
K: FAS/FASL-mediated cell death in the bovine endometrium. Anim
Reprod Sci. 151:97–104. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang SJ, Chen CP, Schatz F, Rahman M,
Abrahams VM and Lockwood CJ: Pre-eclampsia is associated with
dendritic cell recruitment into the uterine decidua. J Pathol.
214:328–336. 2008. View Article : Google Scholar
|
|
15
|
Banzato PC, Daher S, Traina E, Torloni MR,
Gueuvoghlanian-Silva BY, Puccini RF, Pendeloski KP and Mattar R:
FAS and FAS-L genotype and expression in patients with recurrent
pregnancy loss. Reprod Sci. 20:1111–1115. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Choi HK, Choi BC, Lee SH, Kim JW, Cha KY
and Baek KH: Expression of angiogenesis- and apoptosis-related
genes in chorionic villi derived from recurrent pregnancy loss
patients. Mol Reprod Dev. 66:24–31. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Minas V, Jeschke U, Kalantaridou SN,
Richter DU, Reimer T, Mylonas I, Friese K and Makrigiannakis A:
Abortion is associated with increased expression of FasL in
decidual leukocytes and apoptosis of extravillous trophoblasts: A
role for CRH and urocortin. Mol Hum Reprod. 13:663–673. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kiener PA, Davis PM, Rankin BM, Klebanoff
SJ, Ledbetter JA, Starling GC and Liles WC: Human monocytic cells
contain high levels of intracellular Fas ligand: Rapid release
following cellular activation. J Immunol. 159:1594–1598.
1997.PubMed/NCBI
|
|
19
|
Borges VM, Falcão H, Leite-Júnior JH,
Alvim L, Teixeira GP, Russo M, Nóbrega AF, Lopes MF, Rocco PM,
Davidson WF, et al: Fas ligand triggers pulmonary silicosis. J Exp
Med. 194:155–164. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dace DS, Khan AA, Stark JL, Kelly J, Cross
AH and Apte RS: Interleukin-10 overexpression promotes
Fas-ligand-dependent chronic macrophage-mediated demyelinating
polyneuropathy. PLoS One. 4:e71212009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Maugey-Laulom B, Commenges-Ducos M,
Jullien V, Papaxanthos-Roche A, Scotet V and Commenges D:
Endometrial vascularity and ongoing pregnancy after IVF. Eur J
Obstet Gynecol Reprod Biol. 104:137–143. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ding JL, Diao LH, Yin TL, Huang CY, Yin B,
Chen C, Zhang Y, Li J, Cheng YX, Zeng Y and Yang J: Aberrant
expressions of endometrial Id3 and CTLA-4 are associated with
unexplained repeated implantation failure and recurrent
miscarriage. Am J Reprod Immunol. 78:782017. View Article : Google Scholar
|
|
23
|
Guirelli PM, Angeloni MB, Barbosa BF,
Gomes AO, Castro AS, Franco PS, Silva RJ, Oliveira JG,
Martins-Filho OA, Mineo JR, et al: Trophoblast-macrophage crosstalk
on human extravillous under Toxoplasma gondii infection. Placenta.
36:1106–1114. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nagamatsu T and Schust DJ: The
immunomodulatory roles of macrophages at the maternal-fetal
interface. Reprod Sci. 17:209–218. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hamilton S, Oomomian Y, Stephen G,
Shynlova O, Tower CL, Garrod A, Lye SJ and Jones RL: Macrophages
infiltrate the human and rat decidua during term and preterm labor:
Evidence that decidual inflammation precedes labor. Biol Reprod.
86:392012. View Article : Google Scholar
|
|
26
|
Co EC, Gormley M, Kapidzic M, Rosen DB,
Scott MA, Stolp HA, McMaster M, Lanier LL, Bárcena A and Fisher SJ:
Maternal decidual macrophages inhibit NK cell killing of invasive
cytotrophoblasts during human pregnancy. Biol Reprod. 88:1552013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shynlova O, Nedd-Roderique T, Li Y,
Dorogin A and Lye SJ: Myometrial immune cells contribute to term
parturition, preterm labour and post-partum involution in mice. J
Cell Mol Med. 17:90–102. 2013. View Article : Google Scholar
|
|
28
|
Mor G and Abrahams VM: Potential role of
macrophages as immunoregulators of pregnancy. Reprod Biol
Endocrinol. 1:1192003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hemberger M, Hughes M and Cross JC:
Trophoblast stem cells differentiate in vitro into invasive
trophoblast giant cells. Dev Biol. 271:362–371. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huppertz B, Kadyrov M and Kingdom JC:
Apoptosis and its role in the trophoblast. Am J Obstet Gynecol.
195:29–39. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Straszewski-Chavez SL, Abrahams VM and Mor
G: The role of apoptosis in the regulation of trophoblast survival
and differentiation during pregnancy. Endocr Rev. 26:877–897. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cinar O, Kara F and Can A: Potential role
of decidual apoptosis in the pathogenesis of miscarriages. Gynecol
Endocrinol. 28:382–385. 2012. View Article : Google Scholar
|
|
33
|
Pang W, Zhang Y, Zhao N, Darwiche SS, Fu X
and Xiang W: Low expression of Mfn2 is associated with
mitochondrial damage and apoptosis in the placental villi of early
unexplained miscarriage. Placenta. 34:613–618. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lv X, Cai Z and Li S: Increased apoptosis
rate of human decidual cells and cytotrophoblasts in patients with
recurrent spontaneous abortion as a result of abnormal expression
of CDKN1A and Bax. Exp Ther Med. 12:2865–2868. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Aschkenazi S, Straszewski S, Verwer KM,
Foellmer H, Rutherford T and Mor G: Differential regulation and
function of the Fas/Fas ligand system in human trophoblast cells.
Biol Reprod. 66:1853–1861. 2002. View Article : Google Scholar : PubMed/NCBI
|