Introduction
Preventing myocardial ischemia-reperfusion injury
(MIRI) is a frequent problem during the application of reperfusion
therapy, such as percutaneous coronary intervention in patients
with ST-segment elevation myocardial infarction (1). Accumulating evidence has
demonstrated that a short series of repetitive cycles of
reperfusion and ischemia performed at the onset of sustained
reperfusion, namely ischemic postconditioning (IPoC), attenuated
MIRI in several animal models (2-6)
and certain human studies (7,8).
However, recent studies have reported that the cardioprotective
effect of IPoC is usually abrogated by pathological conditions,
including hypercholesterolemia (9), although the mechanism underlying the
loss of cardioprotection in hypercholesterolemic animals remains
elusive.
It is widely accepted that hypercholesterolemia is
one of the most important risk factors for the development of
atherosclerosis, but there is evidence that hypercholesterolemia
also exerts direct negative effects on the myocardium itself
(10), including increased
endoplasmic reticulum (ER) stress (11), altered activation of signaling
pathways (12,13) and induction of apoptosis (13), and some researchers believe that
this may partly explain why the cardioprotective effect of IPoC is
suppressed by hypercholesterolemia (14). For example, Andreadou et al
(15) investigated whether
hypercholesterolemia suppresses the protective effect of IPoC
through a decrease in the phosphorylation of endothelial nitric
oxide synthase (eNOS) and the production of nitric oxide (NO). Our
previous studies demonstrated that the repression of IPoC by
hypercholesterolemia is associated with inactivation of the
reperfusion injury salvage kinase (RISK) pathway (13) and excessive mitochondrial
permeability transition pore (mPTP) opening (12). Of note, it was observed that
fasudil, a Rho-kinase inhibitor, restored the cardioprotective
effect of IPoC in hypercholesterolemic rats by activating the
PI3K/AKT/eNOS signaling pathway (16). Furthermore, it was demonstrated
that blocking of mPTP opening with cyclosporine A could restore the
benefits of IPoC in hypercholesterolemic rat hearts (17). Therefore, ameliorating the
negative impact of hypercholesterolemia on the myocardium by
pharmacological intervention appears to be a novel method for
preserving or restoring the cardioprotection of IPoC under
hypercholesterolemic conditions.
Lycopene (LP), a type of carotenoid present in
tomatoes and other red fruits and vegetables, has potent
antioxidant properties, and has been reported to reduce the risk of
multiple diseases, including coronary heart disease (18). More importantly, several recent
studies have demonstrated that LP protects against MIRI by
inhibiting the development of ER stress (19) and blocking mPTP opening (20). Given that LP can alleviate the
negative impact of hypercholesterolemia on the myocardium, we
hypothesized that LP treatment may preserve or restore the
protective effect of IPoC against MIRI in the presence of
hypercholesterolemia.
The aim of the present study was to determine
whether LP treatment could restore the cardioprotective effect of
IPoC in hypercholesterolemic rat hearts, and elucidate the
underlying mechanism.
Materials and methods
Animals and establishment of a
hypercholesterolemic rat model
A total of 110 male Wistar rats, weighing 100±10 g,
were purchased from Changsheng Biotechnology Co., Ltd. All rats
were housed in an environment of constant temperature (22-25°C) and
relative humidity (50-60%) on a 12/12 h light/dark cycle, and food
and water were provided ad libitum. To establish the
hypercholesterolemic rat model, the rats were divided in two groups
as follows: i) High-cholesterol diet (HCD) group (n=90), rats were
consecutively fed a cholesterol-enriched feedstuff, including 1.5%
cholesterol, 10% egg yolk powder, 10% lard, 0.5% sodium cholate, 3%
sugar and 75% normal feedstuff for 12 weeks; and ii) normal diet
(ND) group (n=20), rats were consecutively fed normal feedstuff for
the same time period. After 12 weeks, 1 ml blood samples were
collected from the tail vein to determine the serum values of total
cholesterol (TC), high-density lipoprotein-cholesterol and
low-density lipoprotein (LDL)-cholesterol using commercial kits
(Nanjing Jiancheng Bioengineering Institute, cat. nos. A111-1,
A112-2 and A113-2) according to the manufacturer's instructions.
All rats were handled according to the Guide for the Care and Use
of Laboratory Animals (National Institutes of Health) (21). The experimental protocol was
approved by the Institutional Ethics Committee of China Medical
University (Shenyang, China).
Establishment of MIRI and IPoC model in
isolated rat hearts
Heart preparation was performed as previously
described (22). Briefly, the
rats were anesthetized with intraperitoneal injection of
pentobarbital sodium (30 mg/kg). Each rat heart was rapidly
isolated and then placed into an iced heparinized K-H solution
(NaCl 0.15 mol/l, KCl 0.006 mol/l, CaCl2 0.002 mol/l and
NaHCO3 0.002 mol/l) for trimming. Subsequently, the
isolated rat heart was rapidly mounted to the Langendorff apparatus
through the aorta and perfused with K-H solution saturated with 95%
O2 and 5% CO2 at 75 mmHg and 37°C. A
water-filled latex balloon was inserted into the left ventricle
through the left atrium and connected to a pressure transducer for
pressure measurement. All isolated rat hearts were stabilized for
10 min before ischemia, and then subjected to 30 min global
ischemia followed by 60 min reperfusion to create the MIRI model.
As for the induction of IPoC, six cycles of brief
ischemia/reperfusion [(10/10 sec) x6] were performed at the onset
of sustained reperfusion.
Experimental protocol
Rats in the HCD group (n=90) were randomly divided
into six groups with 15 rats per group as follows: i) IR group,
isolated rat hearts were subjected to 30 min global ischemia
followed by 60 min reperfusion, as previously described; ii) IPoC
group, following 30 min of ischemia, six cycles of IPoC were
performed at the onset of reperfusion, followed by 58 min of
reperfusion; iii) 5 mg/kg/day LP treatment (LP-5) group, LP (CAS
no. 502-65-8, purity ≥90%), purchased from Meilun Biological
Technology Co. Ltd., was dissolved in corn oil (Sigma-Aldrich;
Merck KGaA), rats were treated with LP (5 mg/kg/day) for 5
consecutive days then other experimental procedures were as
described for the IR group; iv) IPoC + LP-5 group, rats were
treated with LP (5 mg/kg/day) for 5 consecutive days, and then IPoC
was performed as described for the IPoC group; v) 20 mg/kg/day LP
treatment (LP-20) group, rats were treated with LP (20 mg/kg/d) for
5 consecutive days, then other experimental procedures were as
described for the IR group; vi) sodium 4-phenylbutyrate treatment
(4-PBA) group, 4-PBA (Sigma-Aldrich; Merck KGaA), an ER stress
inhibitor, was intraperitoneally administered at a dose of 20 mg/kg
1 h prior to ischemia, then IR experimental procedures were as
described for the IR group.
According to previous studies (23-25), the dose of LP used for
intraperitoneal injection in rats ranged between 2.5 and 20
mg/kg/day; therefore, 5 mg/kg/day was selected as the low dose and
20 mg/kg/day as the high dose of LP treatment. Moreover, the period
of LP treatment through intraperitoneal injection ranged between 1
and 10 days in previous studies. Therefore, an intermediate
treatment period of 5 days was selected for this study. LP was
administered intraperitoneally at a dose of 5 mg/kg/day for 5
consecutive days. The dose of 4-PBA was based on a previous study
(26).
Measurement of changes in serum lipids
after LP treatment
To assess whether LP has an effect on the level of
serum lipids, 45 rats in the LP-5, IPoC + LP-5 and LP-20 group
(n=15 per group) were performed lipid profile as described earlier,
after received LP administration at a dose of 5 or 20 mg/kg/day for
5 consecutive days.
Observation of myocardial mitochondria by
electron microscopy
At the end of the reperfusion period, a 1×1×1-mm
piece of cardiac tissue was isolated from the left ventricle and
electron microscopic sections were prepared as previously described
(27). The morphological changes
of the mitochondria were observed by transmission electron
microscopy (JEM-1200EX; JEOL, Ltd.).
Hematoxylin and eosin staining
The hearts were removed from the perfusion apparatus
at the end of the reperfusion period. The left ventricular heart
tissues were fixed in 4% paraformaldehyde for 24 h at room
temperature, dehydrated and embedded in paraffin. The paraffin
blocks were then cut into 5-µm sections. The paraffin
sections were dewaxed, stained with hematoxylin for 5 min, followed
by 1% hydrochloric acid alcohol differentiation for 3 sec and eosin
staining for 3 min at room temperature. The pathological changes in
heart tissues were observed under a light microscope and imaged. To
evaluate myocardial damage quantitatively, the sections were scored
by two independent observers blinded to the experimental protocol,
as previously described (28): 0,
no damage; 1, interstitial edema and focal necrosis; 2, diffuse
myocardial cell swelling and necrosis; 3, necrosis with the
presence of contraction bands and neutrophil infiltrate; and 4,
widespread necrosis with the presence of contraction bands,
neutrophil infiltrate and hemorrhage.
Measurement of infarct size
Myocardial infarct size was measured by
triphenyltetrazolium chloride (TTC) staining. The hearts were
harvested at the end of the reperfusion period and the heart
samples were frozen at −80°C for 30 min. The frozen hearts were cut
into 1-2-mm sections from the apex to the bottom and incubated in
1% TTC solution at 37°C for 30 min. Subsequently, the heart
sections were washed with 1X PBS and fixed in 4% paraformaldehyde
overnight at room temperature. The stained sections were
photographed by a digital camera and analyzed using ImageJ2x
analysis software (National Institutes of Health). The severity of
myocardial infarction was reflected by the ratio of infarct size to
total size.
Measurement of lactate dehydrogenase
(LDH) and creatine kinase muscle/brain isoenzyme (CK-MB)
activity
LDH and CK-MB are released from cardiomyocytes into
the coronary effluent during perfusion (29). The activities of LDH and CK-MB in
the coronary effluent were determined using the LDH and CK-MB assay
kits (Nanjing Jiancheng Bioengineering Institute; cat. nos. A020-1
and E006) according to the manufacturer's protocols. The absorbance
value was detected at 450 nm by ultramicro microporous plate
spectrophotometer (BioTek Instruments, Inc.).
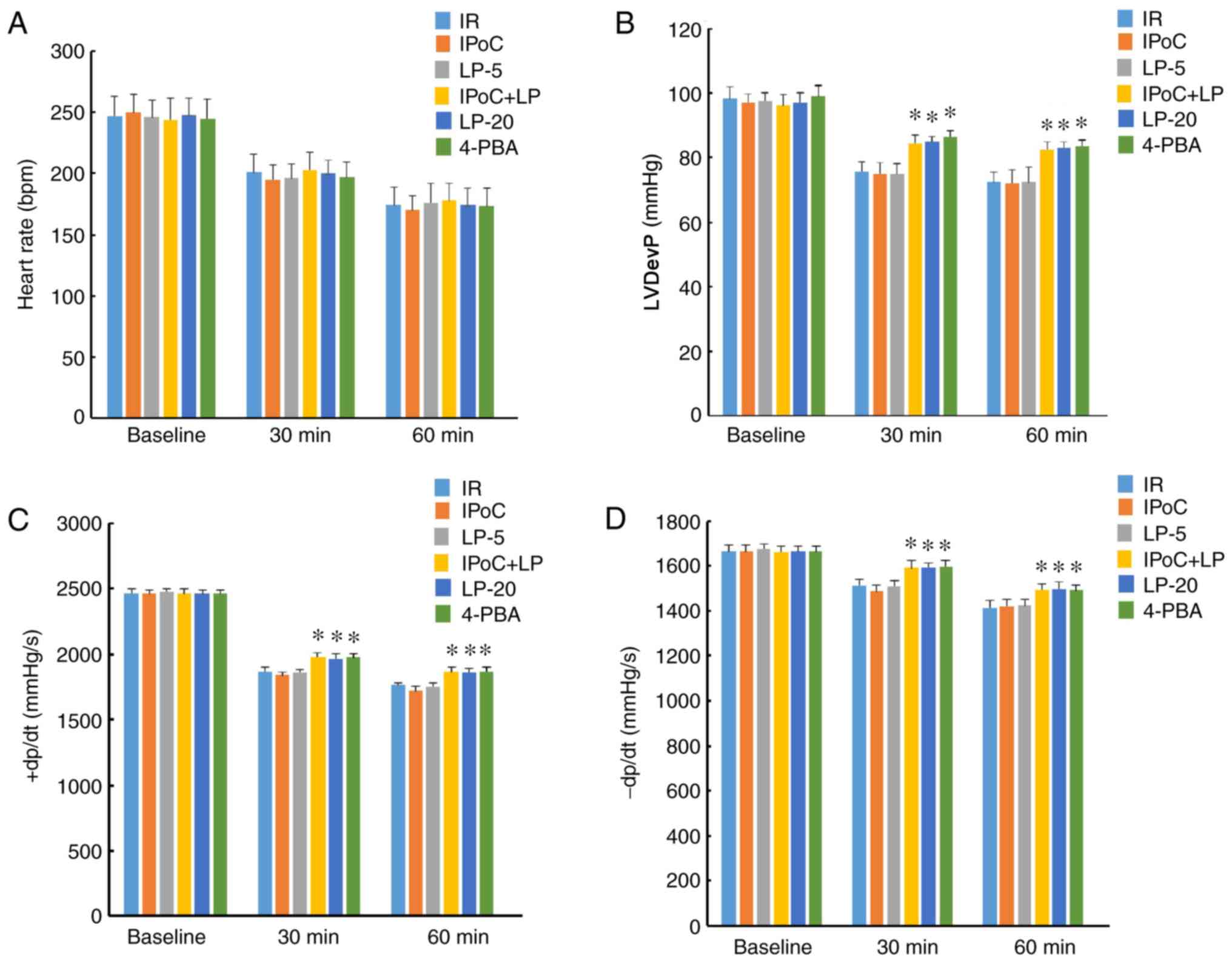
Cardiac function monitoring
To evaluate the change in cardiac function, heart
rate, left ventricular developed pressure (LVDevP), positive
first-order derivative of ventricular pressure (+dp/dt) and
negative first-order derivative of ventricular pressure (−dp/dt)
were continuously recorded using a hemodynamic system (MP150;
Biopac Systems, Inc.) during the IR and IPoC the experimental
procedure.
Measurement of cardiomyocyte
apoptosis
Myocardial cell apoptosis was detected by TUNEL
assay using the In Situ Cell Death Detection Kit (Roche
Diagnostics) according to the manufacturer's instructions. The
apoptotic cells were observed under a light microscope and
imaged.
Sensitivity of mPTP to calcium
The mitochondria were isolated from heart tissues
using the Mitochondrial Extraction Kit (BestBio Science) according
to the manufacturer's instructions. The sensitivity of mPTP to
calcium was determined using the Purified Mitochondrial Membrane
Pore Channel Colorimetric Assay kit (Shanghai Genmed Pharmaceutical
Technology Co., Ltd.) according to the manufacturer's
instructions.
Western blot analysis
The heart tissues were minced and homogenized with
RIPA buffer (Beyotime Institute of Biotechnology). Following
centrifugation, the supernatant was collected and protein
concentrations were measured using a bicinchoninic acid protein
concentration kit (Beyotime Institute of Biotechnology). After 30
µg protein was denatured at 100°C for 10 min, it was
separated by SDS-PAGE on 8 or 10% and transferred to a PVDF
membrane. The membrane was blocked using 5% skimmed milk for 1 h at
room temperature, and then incubated with specific antibodies,
including anti-glucose-regulated protein 78 (GRP78; 1:1,000; Abcam;
cat. no. ab108613), anti-C/EBP homologous protein (CHOP; 1:1,000;
Abcam; cat. no. ab11419), anti-AKT (1:1,000; Abcam; cat. no.
ab179463), anti-phospho (p)-AKT (phospho S473; 1:1,000; Abcam; cat.
no. ab81283), anti-ERK1/2 (1:1,000; Abcam; cat. no. ab17942),
anti-p-ERK1/2 (phospho T202 + Y204; 1:1,000; Abcam; cat. no.
ab214362), anti-glycogen synthase kinase-3β (GSK-3β; 1:1,000;
Abcam; cat. no. ab93926), anti-p-GSK-3β (phospho S9; 1:1,000;
Abcam; cat. no. ab107166), anti-cytochrome c (1:1,000;
Abcam; cat. no. ab133504), anti-cleaved caspase-9 (1:1,000; Abcam;
cat. no. ab2324), anti-cleaved caspase-3 (1:1,000; Abcam; cat. no.
ab2302) and anti-β-actin (1:1,000; Zhongshan Jinqiao Biotechnology;
cat. no. TA-09), at 4°C overnight. On the following day, the PVDF
membrane was incubated with horseradish peroxidase-conjugated goat
anti-rabbit IgG (1:4,000; EarthOx Life Science; cat. no.
E030120-01) or goat anti-mouse IgG (1:4,000; EarthOx Life Science;
cat. no. E030110-01) at room temperature for 30 min. Detection of
protein bands was performed using an ECL for western blotting kit
(Beyotime Institute of Biotechnology) according to the
manufacturer's instructions. Relative densitometry was calculated
with Image J2x analysis software.
Statistical analysis
All data are presented as mean ± SD and were
analyzed using SPSS version 17.0 (SPSS, Inc., Chicago, IL, USA).
Differences among three or more groups were first evaluated using
one-way analysis of variance, and if the differences were
significant, multiple comparison analysis was further performed
with Fisher's least significant difference test. Differences
between two groups were evaluated using independent samples t-test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Changes in serum lipids
The serum levels of TC and LDL were significantly
increased in the HCD group as compared with the ND group after 12
weeks (Table I). The result
suggested that the hypercholesterolemic rat model was successfully
established. Moreover, there was no significant difference in the
serum level of TC and LDL after a 5-day LP treatment either at the
dose of 5 or 20 mg/kg/day (Table
II), suggesting that short-term LP treatment did not affect the
serum levels of TC and LDL.
 | Table IAnalysis of serum lipid profile. |
Table I
Analysis of serum lipid profile.
| Group | TC (mg/dl) | HDL-C (mg/dl) | LDL-C (mg/dl) |
|---|
| Normal diet
(n=20) | 65.27±6.76 | 26.61±5.87 | 32.28±6.59 |
| High cholesterol
diet (n=90) |
132.08±17.42a | 27.19±5.23 | 92.96±19.08a |
| Table IIAnalysis of the change in the serum
lipid profile following lycopene (5 or 20 mg/kg/day) treatment. |
Table II
Analysis of the change in the serum
lipid profile following lycopene (5 or 20 mg/kg/day) treatment.
| Lycopene
treatment | TC (mg/dl) | HDL-C (mg/dl) | LDL-C (mg/dl) |
|---|
| Before | 132.08±17.42 | 27.19±5.23 | 92.96±19.08 |
| After | 131.37±16.21 | 26.97±4.27 | 92.40±16.83 |
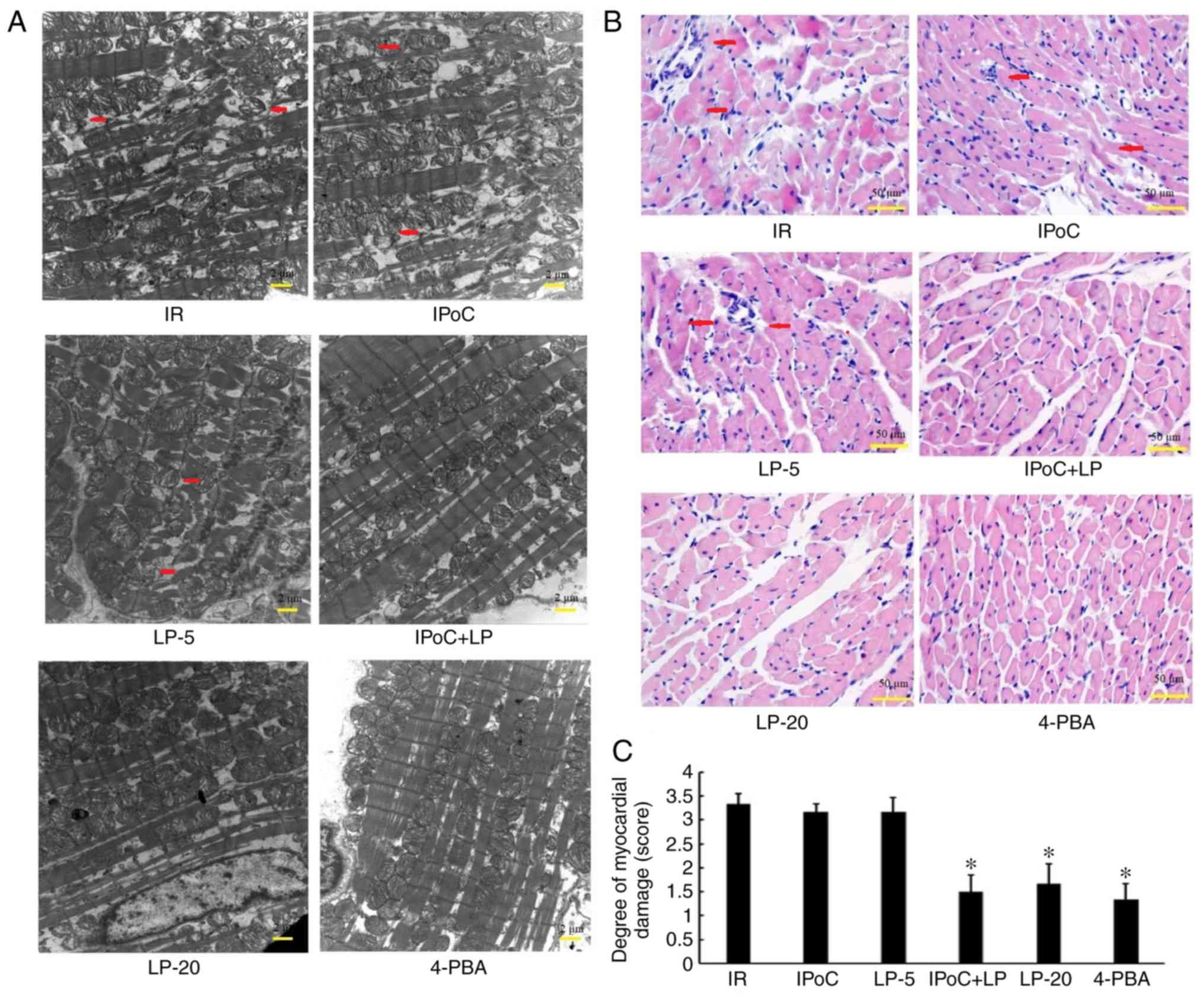
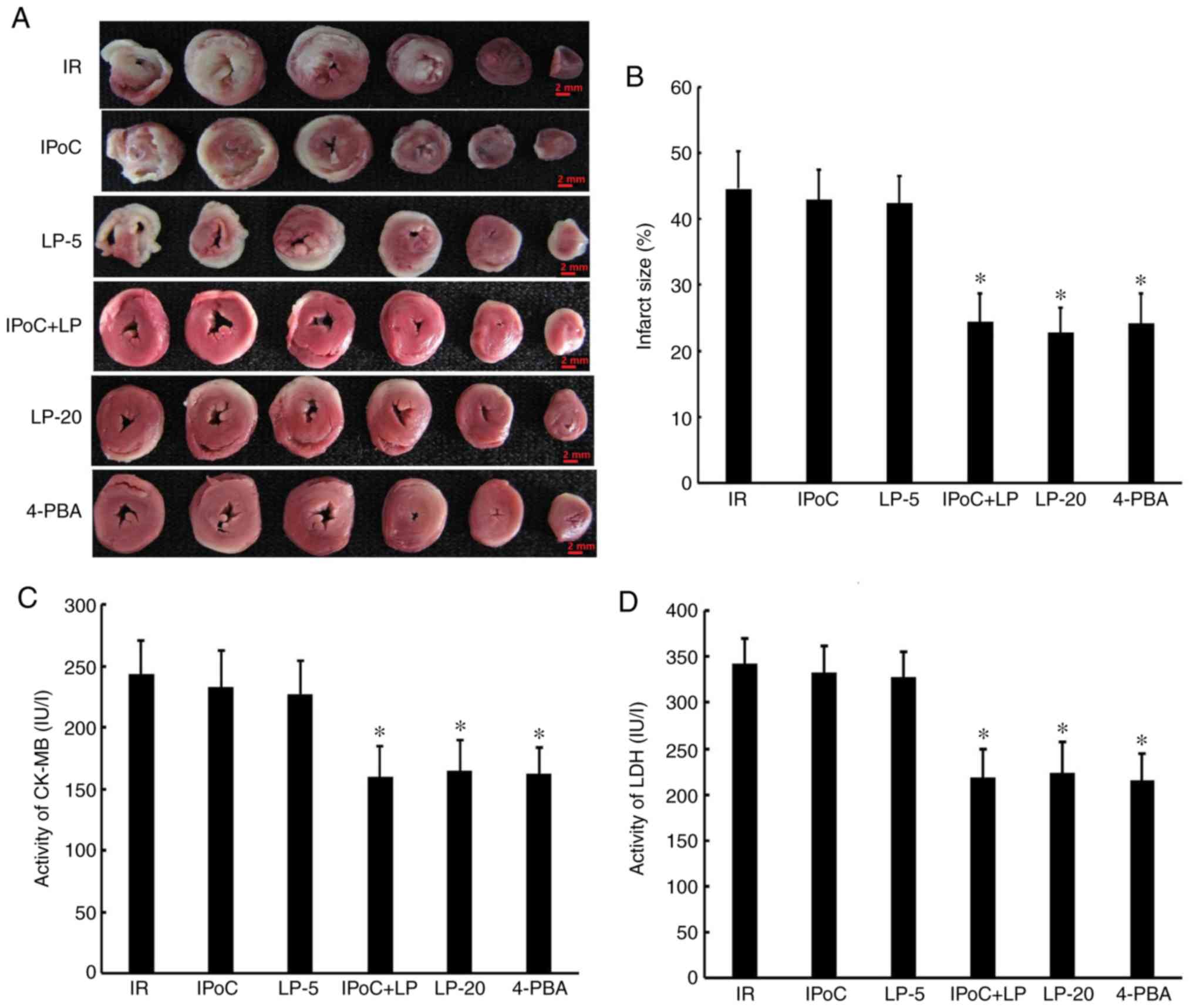
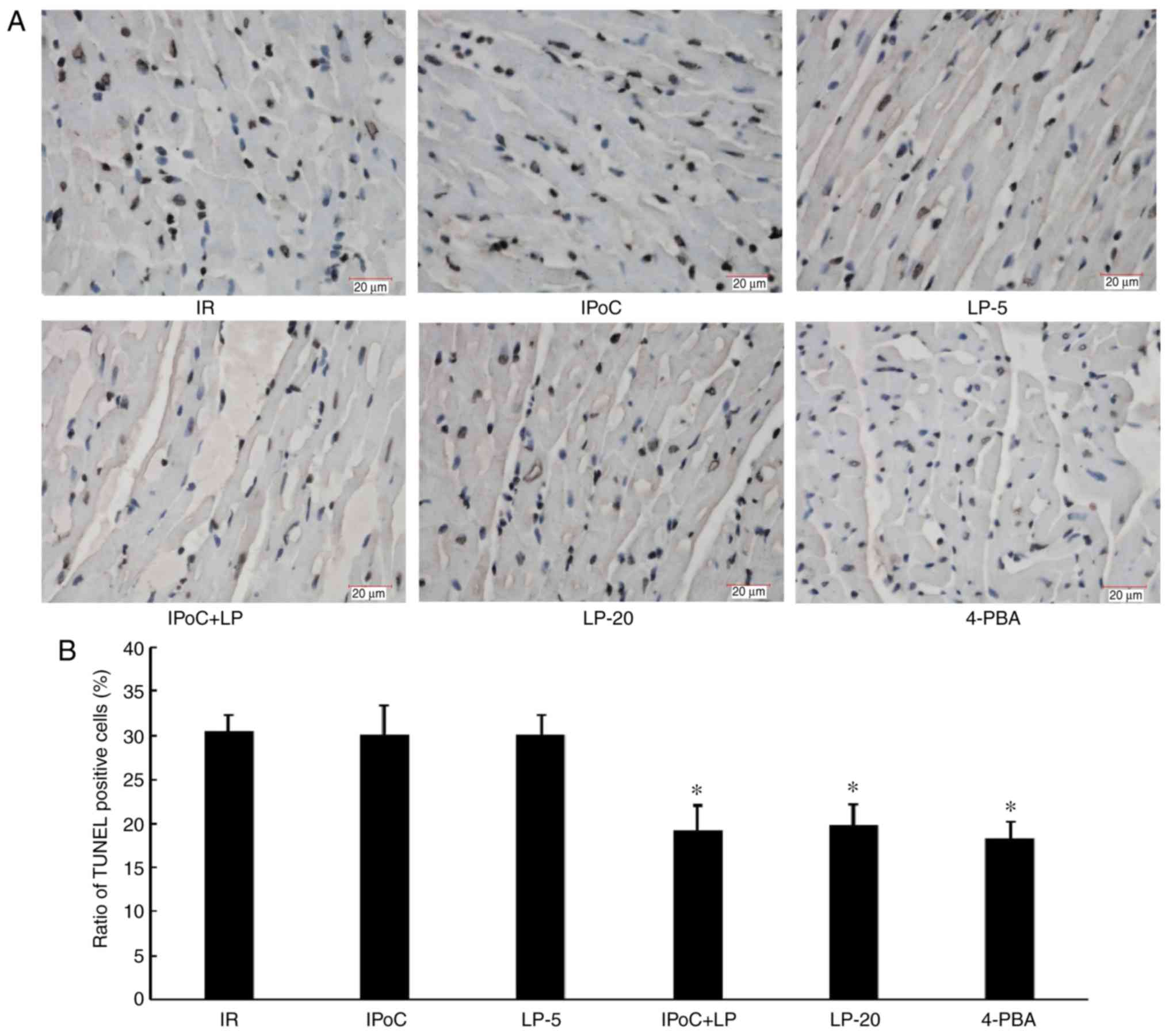
LP restores the protective effect of IPoC
in hypercholesterolemic rats
In the IPoC + LP-5 group, the swelling of the
mitochondria was reduced and mitochondrial cristae were denser, as
observed by electron microscopy (Fig.
1A); myocardial degeneration and inflammatory infiltration were
also less extensive, as observed by light microscopy (Fig. 1B and C); myocardial infarct size
(Fig. 2A and B) and the release
of CK-MB and LDH (Fig. 2C and D)
were reduced and the apoptosis rate was decreased (Fig. 3), whereas cardiac function, as
reflected by LVDevP and ±dp/dt, was improved (Fig. 4) in the IPoC + LP group compared
with the IR group under conditions of hypercholesterolemia,
although no beneficial effects were observed in the LP-5 and IPoC
only groups. These results suggest that LP pretreatment may
preserve or restore the cardioprotective effect of IPoC in
hypercholesterolemic rats. Moreover, the LP-20 and 4-PBA groups
displayed beneficial effects similar to the effects in the IPoC +
LP-5 group, suggesting that high-dose LP treatment or
pharmacological ER stress inhibition may protect
hypercholesterolemic rats against MIRI.
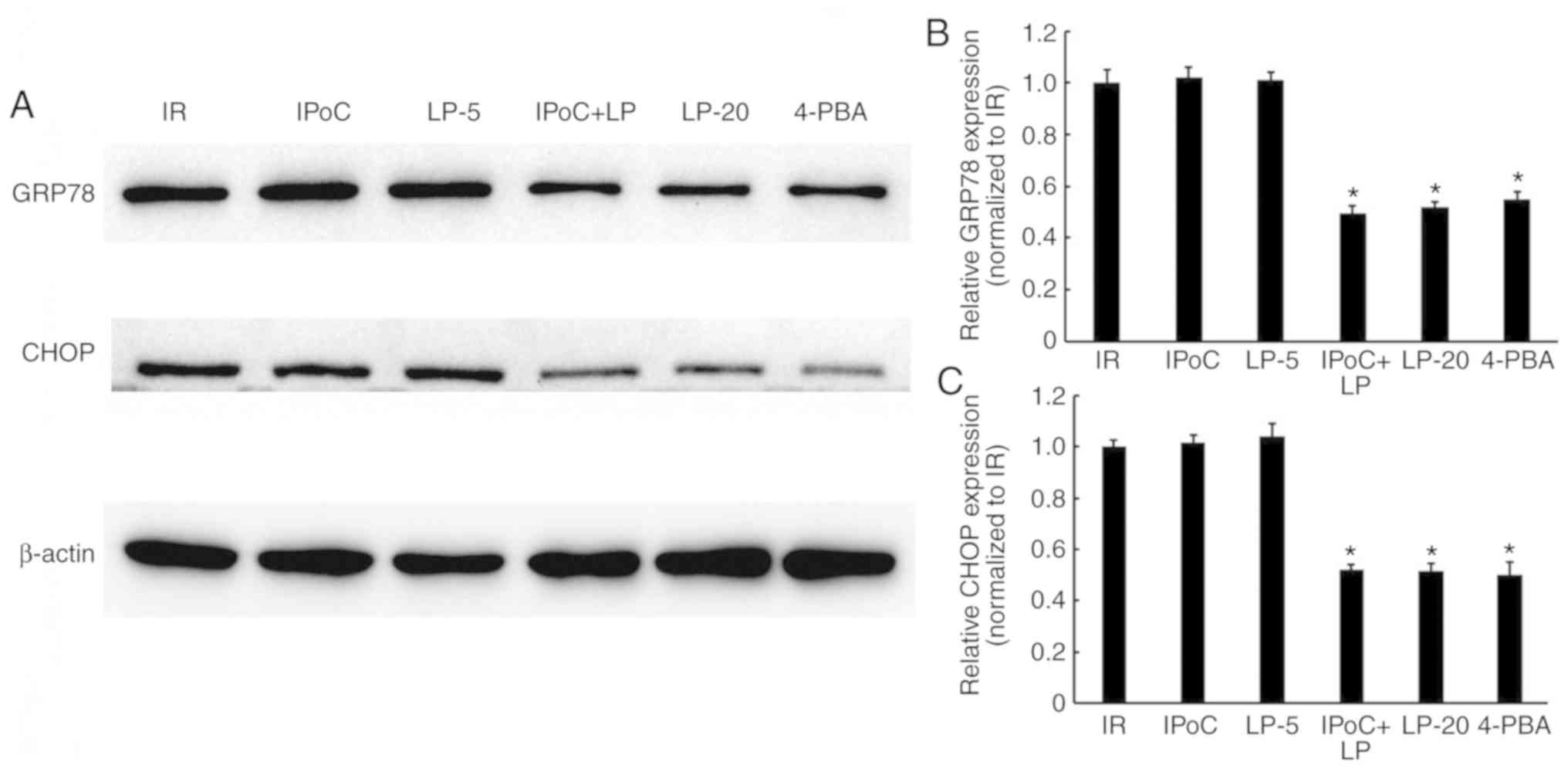
Effects of LP, alone or combined with
IPoC, on ER stress in hypercholesterolemic rats
As shown in Fig.
5, the protein expression of GRP78 and CHOP (markers of ER
stress) was markedly decreased in the IPoC + LP-5 and LP-20 groups
compared with IR and IPoC groups, which was similar to the effect
of the ER stress inhibitor 4-PBA. However, no significant change
was observed in the IPoC and LP-5 only groups. Therefore, high-dose
LP or the combination of low-dose LP and IPoC were successful in
reducing ER stress in hypercholesterolemic rats.
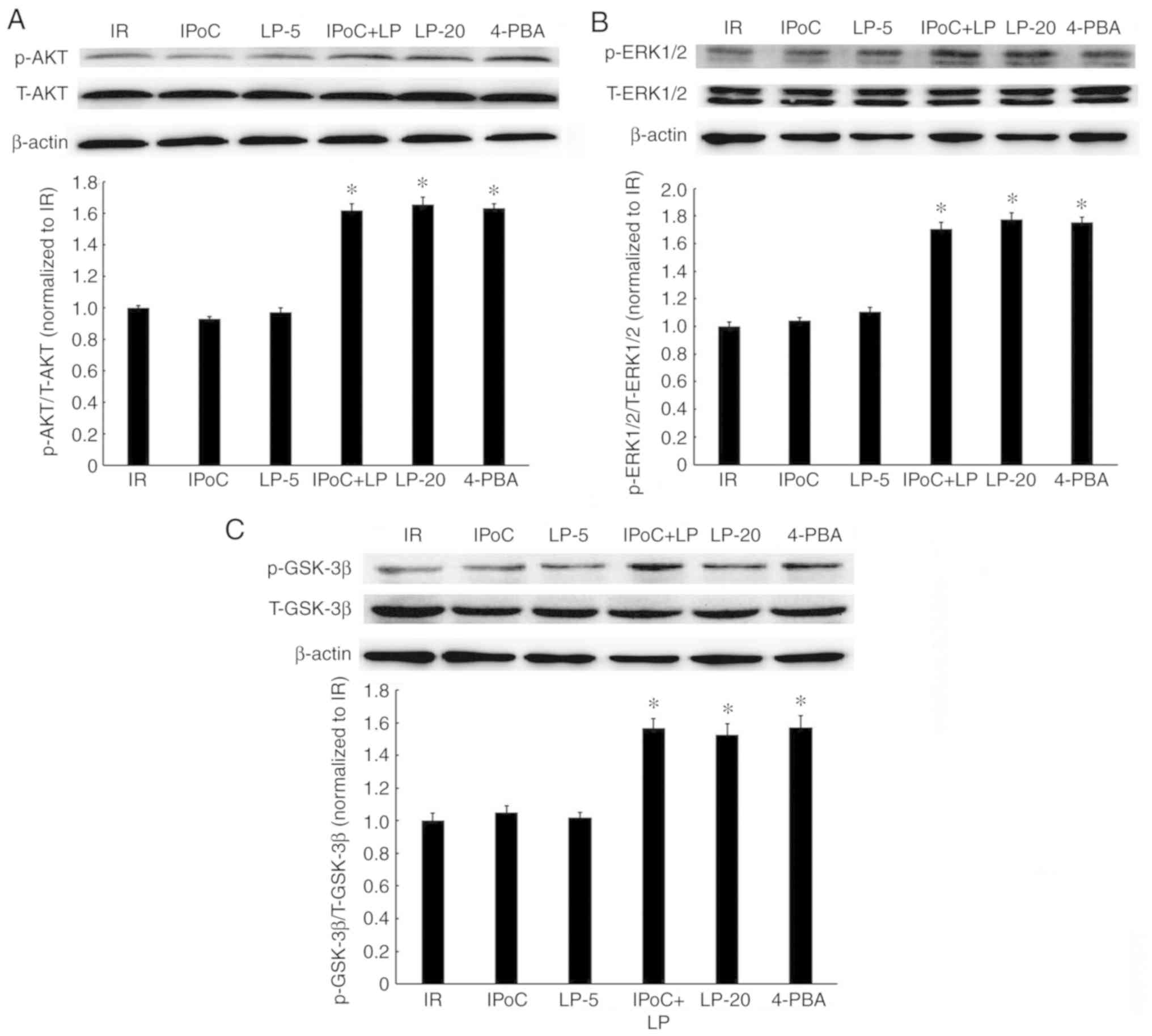
Effects of LP, alone or combined with
IPoC, on the RISK pathway in hypercholesterolemic rats
Considering that the RISK pathway is suppressed in
hypercholesterolemic rat hearts (12,13), the effect of LP, alone or combined
with IPoC, on the activation of AKT, ERK1/2 and GSK-3β was
investigated. The results revealed that the phosphorylation of AKT
(Fig. 6A), ERK1/2 (Fig. 6B) and GSK-3β (Fig. 6C) were enhanced in the IPoC +
LP-5, LP-20 and 4-PBA groups, compared with the IR group. However,
no significant change in the phosphorylation levels of AKT, ERK1/2
or GSK-3β was observed in the IPoC and LP-5 only groups. Thus,
high-dose LP or the combination of low-dose LP and IPoC
successfully activated the RISK pathway in hypercholesterolemic
rats.
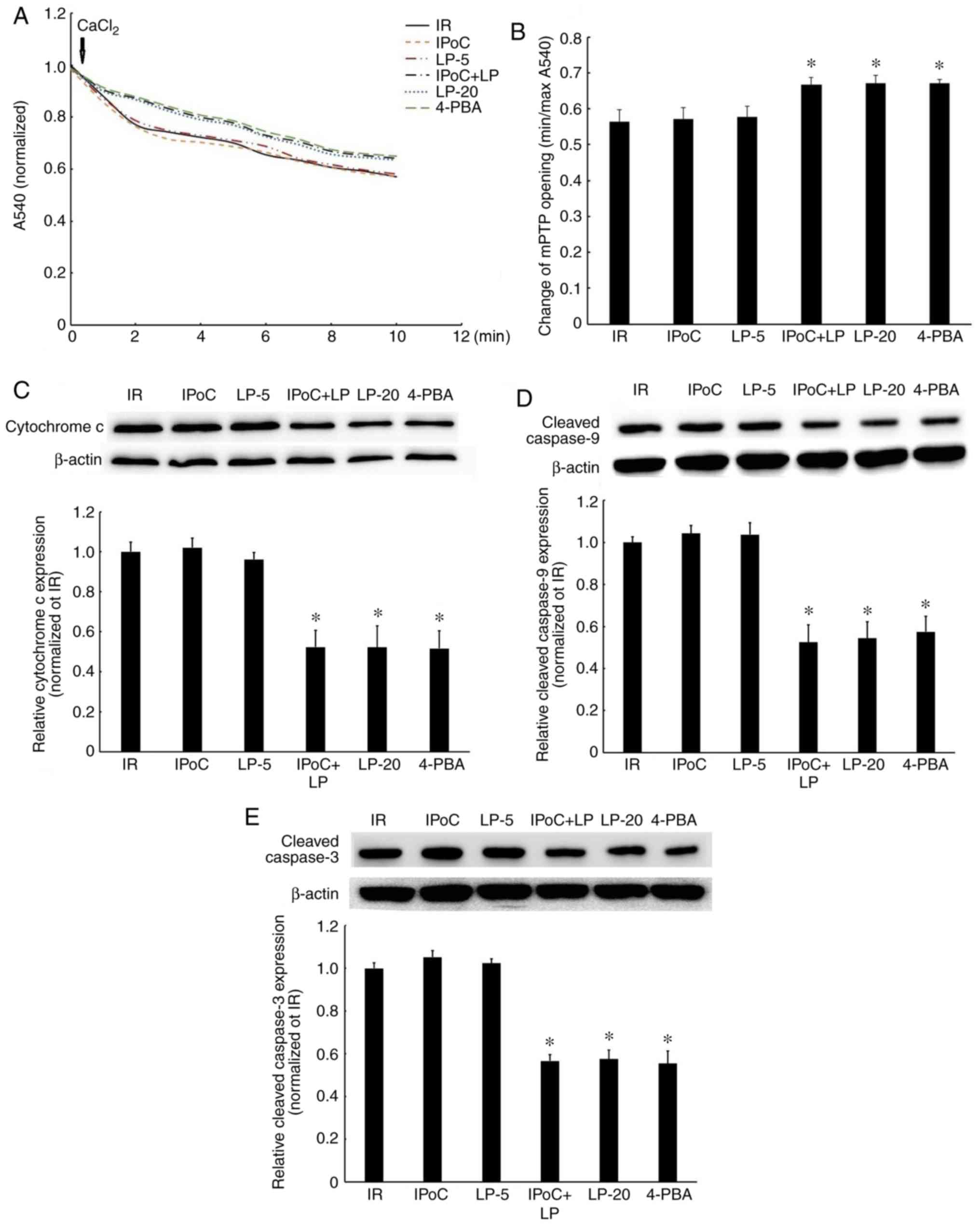
Effects of LP, alone or combined with
IPoC, on mPTP opening in hypercholesterolemic rats
mPTP is the end effector of IPoC and is regulated by
the RISK pathway (30). Thus, the
effect of LP, alone or combined with IPoC, on mPTP opening was
investigated. The results demonstrated that the sensitivity of mPTP
to calcium, an inducer of mPTP opening (31), was markedly decreased in the IPoC
+ LP-5, LP-20 and 4-PBA groups, compared with the IR group
(Fig. 7A and B), but no
significant change was observed in the IPoC and LP-5 groups. mPTP
opening may lead to the activation of the mitochondrial apoptosis
pathway. As expected, the expressions of mitochondrial
apoptosis-related proteins, including cytochrome c, cleaved
caspase-9 and cleaved caspase-3, were also reduced in the LP-5,
LP-20 and 4-PBA groups, compared with the IR group (Fig. 7C-E), but no significant change was
observed in the IPoC and LP-5 only groups, which was in accordance
with the extent of mPTP opening. Taken together, these results
confirmed that high-dose LP or the combination of low-dose LP and
IPoC inhibit mPTP opening in hypercholesterolemic rat hearts.
Discussion
Whether the cardioprotective effect of IPoC is
suppressed by hypercholesterolemia remains controversial (14). The results of the present study
demonstrated that the cardioprotection of IPoC is lost in rats fed
a high-cholesterol diet for 12 weeks, which is consistent with
previous findings in mini-swine fed a cholesterol-enriched diet for
4 weeks (32) and rabbits fed a
cholesterol-enriched diet for 6 weeks (33). However, Donato et al
(34) reported that the
cardioprotection of IPoC was preserved in rabbits after 4 weeks on
a cholesterol-enriched diet. It is hypothesized that this
discrepancy may be partly attributed to the differences in the
duration of the cholesterol-enriched diet and the animal
species.
Statins, a widely used class of lipid-lowering
drugs, were found to restore the loss of IPoC benefits in
hypercholesterolemic animals. Iliodromitis et al (35) first reported that the loss of the
IPoC benefits may be reversed in hypercholester-olemic rabbits that
received a 3-week simvastatin treatment due to the decrease in TC
and LDL plasma levels and attenuation of the oxidative and
nitrosative stress in the ischemic myocardium. Moreover, Andreadou
et al (15) reported that
a 3-day pravastatin treatment preserved the cardioprotective effect
of IPoC in hypercholesterolemic rabbits, independently of any
lipid-lowering effects, potentially through eNOS activation and the
attenuation of nitro-oxidative stress. In the present study, a
5-day LP treatment achieved no decrease in serum lipids, but the
combination of low-dose LP and IPoC attenuated MIRI in
hypercholesterolemic rats. Although LP has antihyperlipidemic
properties, its lipid-lowering effect usually requires long-term
treatment. Zeng et al (36) found that the values of TC and LDL
had not significantly changed when measured after 1 week of LP
treatment, but exhibited a significant decrease after ≥3 weeks of
LP treatment. This may explain why blood lipid levels had not
decreased after a 5-day LP treatment in the present study. Given
that the cardioprotective effect of IPoC is abolished by
hypercholesterolemia, LP administration may preserve or restore the
protective effect of IPoC in hypercholesterolemic rats,
independently of any lipid-lowering effects. Interestingly, it was
also observed that hypercholesterolemic rats treated with high-dose
LP alone exhibited similar cardioprotection as those receiving a
combination of low-dose LP and IPoC. Thus, it may be further
inferred that LP in combination with IPoC exerts synergistic or
additive protective effects against MIRI in hypercholesterolemic
rats. Although high-dose LP treatment exerted protective effects in
IR hearts under conditions of hypercholesterolemia, its side
effects on other organs could not be determined due to the isolated
rat heart model used in the present study, which is a potential
limitation for its use in clinical practice. In contrast to
high-dose LP treatment, low-dose LP combined with IPoC exerted
similar cardioprotective effects, but was associated with fewer
potential side effects due to the lower LP dose; for instance, high
doses of LP may increase the risk of lung cancer in smokers
(37,38). Therefore, combined treatment may
represent a safer method for preventing MIRI in patients with
hypercholesterolemia.
ER is responsible for the biosynthesis of all
secreted and membrane proteins. The accumulation and aggregation of
unfolded proteins in the ER leads to a condition referred to as ER
stress (39). The unfolded
protein response is initially activated to degrade unfolded
proteins and restore normal ER function. If ER stress is prolonged,
or if the adaptive response fails, the cells will die through
apoptosis (40). Thus, ER stress
is crucial for most cellular activities and survival. GRP78 and
CHOP, two major ER stress-related proteins, reflect the activation
of ER stress. The levels of GRP78 and CHOP were found to increase
in the ischemic myocardium by the combination of low-dose LP and
IPoC, which is similar to the effects of ER stress inhibitors, such
as 4-PBA. Considering that hypercholesterolemia increases ER stress
in the IR myocardium (11) and LP
reduces ER stress in hypoxic-reoxygenated cardiomyocytes (19), it may be deduced that the
restoration of IPoC in hypercholesterolemic rat by LP treatment may
be mediated via inhibition of ER stress.
IPoC protects against MIRI through activation of the
RISK pathway (AKT and ERK1/2) (41), and subsequently inhibiting GSK-3β
and mPTP opening (42); however,
hypercholesterolemia suppresses the activation of the RISK pathway
and induces the activation of GSK-3β and mPTP opening (12,13). In the present study, the
combination of LP and IPoC activated AKT and ERK1/2, subsequently
inhibiting GSK-3β and mPTP opening in hypercholesterolemic rat
hearts, suggesting that the restoration of IPoC by LP may be partly
attributed to the reactivation of the RISK pathway. ER stress has
been demonstrated to impair the activation of the RISK pathway
(43). Miki et al
(43) provided evidence that ER
stress may suppress the phosphorylation of AKT and ERK1/2, and
further impair GSK-3β-mediated suppression of mPTP opening.
Notably, the combination of LP and IPoC exerted the same effect on
the RISK pathway as that of an ER stress inhibitor. Thus, it may be
inferred that LP reactivates AKT and ERK1/2 in hypercholesterolemic
rat hearts partly through inhibition of ER stress.
There were certain limitations to the present study.
First, although LP restored the cardioprotection of IPoC in
hypercholesterolemic rats by inhibition of ER stress and activation
of the RISK pathway, whether activating ER stress or inhibiting the
RISK pathway by a pharmacological or genetic approach could reverse
the restoration of IPoC in hypercholesterolemic rats was not
determined. Therefore, these conclusions should be further
confirmed in future studies. Second, MIRI was mimicked in isolated
rat hearts using a Langendorff apparatus. In this model, the
ischemic heart was only subjected to 60-120 min reperfusion and
then harvested (44). Therefore,
cardiac fibrosis or remodeling, including changes in histology and
protein biomarkers (matrix metallopeptidase-2/-9 and cathepsins
S/K) (45-48), could not be observed in this model
within this short period of time. Finally, considerable evidence
indicates that reactive oxygen species (ROS) have a predominant
role in myocardial damage induced by reperfusion (49,50). However, the present study mainly
focused on the role of the regulation of ER stress and the RISK
signaling pathway in the restoration of IPoC by LP under
hypercholesterolemic conditions and the lack of ROS detection is a
limitation of the present study. Additional mechanisms that may be
involved in this process, such as oxidative stress mediated by ROS,
require further investigation.
Collectively, to the best of our knowledge, the
findings of the present study are the first to demonstrate that LP
restores the cardioprotective effect of IPoC in
hypercholesterolemic rats. Furthermore, the restoration of IPoC by
LP in hypercholesterolemic rat hearts was found to be associated
with the inhibition of ER stress and activation of the RISK
pathway. Given that LP is non-toxic and has fewer adverse effects
compared with statins, the combination of LP and IPoC may be a
novel therapy for preventing MIRI in patients with
hypercholesterolemia that receive percutaneous coronary
intervention.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81670320 and 81800232) and
the Natural Science Foundation of Liaoning Province (grant no.
201602826).
Availability of data and materials
The datasets generated and analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
DJ and NW conceived and designed the experiments. LD
conducted the experiments. CL, XL, ZH and SL participated in the
completion of the experiments. LD and NW analyzed the data. LD
wrote the paper. All authors read and approved the final version of
this manuscript.
Ethics approval and consent to
participate
The experimental protocol was approved by the
Institutional Ethics Committee of China Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Bulluck H, Yellon DM and Hausenloy DJ:
Reducing myocardial infarct size: Challenges and future
opportunities. Heart. 102:341–348. 2016. View Article : Google Scholar :
|
|
2
|
Zhao ZQ, Corvera JS, Halkos ME, Kerendi F,
Wang NP, Guyton RA and Vinten-Johansen J: Inhibition of myocardial
injury by ischemic postconditioning during reperfusion: Comparison
with ischemic preconditioning. Am J Physiol Heart Circ Physiol.
285:H579–H588. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shinohara G, Morita K, Nagahori R, Koh Y,
Kinouchi K, Abe T and Hashimoto K: Ischemic postconditioning
promotes left ventricular functional recovery after cardioplegic
arrest in an in vivo piglet model of global ischemia reperfusion
injury on cardiopulmonary bypass. J Thorac Cardiovasc Surg.
142:926–932. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
You L, Li L, Xu Q, Ren J and Zhang F:
Postconditioning reduces infarct size and cardiac myocyte apoptosis
via the opioid receptor and JAK-STAT signaling pathway. Mol Biol
Rep. 38:437–443. 2011. View Article : Google Scholar
|
|
5
|
Cao S, Liu Y, Wang H, Mao X, Chen J, Liu
J, Xia Z, Zhang L, Liu X and Yu T: Ischemic postconditioning
influences electron transport chain protein turnover in
Langendorff-perfused rat hearts. Peer J. 4:e17062016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tong G, Aponte AM, Kohr MJ, Steenbergen C,
Murphy E and Sun J: Postconditioning leads to an increase in
protein S-nitrosylation. Am J Physiol Heart Circ Physiol.
306:H825–H832. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Araszkiewicz A, Grygier M, Pyda M,
Rajewska J, Michalak M, Lesiak M and Grajek S: Postconditioning
reduces enzymatic infarct size and improves microvascular
reperfusion in patients with ST-segment elevation myocardial
infarction. Cardiology. 129:250–257. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Araszkiewicz A, Grygier M, Pyda M,
Rajewska J, Lesiak M and Grajek S: Postconditioning attenuates
early ventricular arrhythmias in patients with high-risk ST-segment
elevation myocardial infarction. J Cardiol. 65:459–465. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ferdinandy P, Hausenloy DJ, Heusch G,
Baxter GF and Schulz R: Interaction of risk factors, comorbidities,
and comedications with ischemia/reperfusion injury and
cardioprotection by preconditioning, postconditioning, and remote
conditioning. Pharmacol Rev. 66:1142–1174. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
D'Annunzio V, Donato M, Buchholz B, Pérez
V, Miksztowicz V, Berg G and Gelpi RJ: High cholesterol diet
effects on ischemia-reperfusion injury of the heart. Can J Physiol
Pharmacol. 90:1185–1196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu N, Zhang X, Jia P and Jia D:
Hypercholesterolemia aggravates myocardial ischemia reperfusion
injury via activating endoplasmic reticulum stress-mediated
apoptosis. Exp Mol Pathol. 99:449–454. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu N, Zhang X, Guan Y, Shu W, Jia P and
Jia D: Hypercholesterolemia abrogates the cardioprotection of
ischemic postconditioning in isolated rat heart: Roles of glycogen
synthase kinase-3β and the mitochondrial permeability transition
pore. Cell Biochem Biophys. 69:123–130. 2014. View Article : Google Scholar
|
|
13
|
Wu N, Zhang X, Jia P and Jia D:
Hypercholesterolemia abrogates the protective effect of ischemic
postconditioning by induction of apoptosis and impairment of
activation of reperfusion injury salvage kinase pathway. Biochem
Biophys Res Commun. 458:148–153. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Andreadou I, Iliodromitis EK, Lazou A,
Görbe A, Giricz Z, Schulz R and Ferdinandy P: Effect of
hypercholesterolaemia on myocardial function, ischaemia-reperfusion
injury and cardio-protection by preconditioning, postconditioning
and remote conditioning. Br J Pharmacol. 174:1555–1569. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Andreadou I, Farmakis D, Prokovas E,
Sigala F, Zoga A, Spyridaki K, Papalois A, Papapetropoulos A,
Anastasiou-Nana M, Kremastinos DT and Iliodromitis EK: Short-term
statin administration in hypercholesterolaemic rabbits resistant to
postconditioning: Effects on infarct size, endothelial nitric oxide
synthase, and nitro-oxidative stress. Cardiovasc Res. 94:501–509.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu N, Li W, Shu W, Lv Y and Jia D:
Inhibition of Rho-kinase by fasudil restores the cardioprotection
of ischemic postcondi-tioninng in hypercholesterolemic rat heart.
Mol Med Rep. 10:2517–2524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu N, Li WN, Shu WQ, Lv Y and Jia DL:
Blocking the mitochondrial permeability transition pore with
cyclosporine-A can restore cardioprotection of ischemic
postconditioning in hypercholesterolemic rat heart. Eur Rev Med
Pharmacol Sci. 19:446–454. 2015.PubMed/NCBI
|
|
18
|
Riccioni G, Mancini B, Di Ilio E,
Bucciarelli T and D'Orazio N: Protective effect of lycopene in
cardiovascular disease. Eur Rev Med Pharmacol Sci. 12:183–190.
2008.PubMed/NCBI
|
|
19
|
Xu J, Hu H, Chen B, Yue R, Zhou Z, Liu Y,
Zhang S, Xu L, Wang H and Yu Z: Lycopene protects against
hypoxia/reoxygenation injury by alleviating ER stress induced
apoptosis in neonatal mouse cardiomyocytes. PLoS One.
10:e01364432015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yue R, Hu H, Yiu KH, Luo T, Zhou Z, Xu L,
Zhang S, Li K and Yu Z: Lycopene protects against
hypoxia/reoxygenation-induced apoptosis by preventing mitochondrial
dysfunction in primary neonatal mouse cardiomyocytes. PLoS One.
7:e507782012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kastenmayer RJ, Moore RM, Bright AL,
Torres-Cruz R and Elkins WR: Select agent and toxin regulations:
Beyond the eighth edition of the guide for the care and use of
laboratory animals. J Am Assoc Lab Anim Sci. 51:333–338.
2012.PubMed/NCBI
|
|
22
|
Jia P, Liu C, Wu N, Jia D and Sun Y:
Agomelatine protects against myocardial ischemia reperfusion injury
by inhibiting mitochondrial permeability transition pore opening.
Am J Transl Res. 10:1310–1323. 2018.PubMed/NCBI
|
|
23
|
Bayramoglu G, Bayramoglu A, Altuner Y,
Uyanoglu M and Colak S: The effects of lycopene on hepatic
ischemia/reperfusion injury in rats. Cytotechnology. 67:487–491.
2015. View Article : Google Scholar :
|
|
24
|
Göncü T, Oğuz E, Sezen H, Koçarslan S,
Oğuz H, Akal A, Adıbelli FM, Çakmak S and Aksoy N:
Anti-inflammatory effect of lycopene on endotoxin-induced uveitis
in rats. Arq Bras Oftalmol. 79:357–362. 2016. View Article : Google Scholar
|
|
25
|
Güzel M, Sönmez MF, Baştuğ O, Aras NF,
Öztürk AB, Küçükaydın M and Turan C: Effectiveness of lycopene on
experimental testicular torsion. J Pediatr Surg. 51:1187–1191.
2016. View Article : Google Scholar
|
|
26
|
Grall S, Prunier-Mirebeau D, Tamareille S,
Mateus V, Lamon D, Furber A and Prunier F: Endoplasmic reticulum
stress pathway involvement in local and remote myocardial ischemic
conditioning. Shock. 39:433–439. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jia D: The protective effect of
mitochondrial ATP-sensitive K+ channel opener, nicorandil, combined
with Na+/Ca2+ exchange blocker KB-R7943 on myocardial
ischemia-reperfusion injury in rat. Cell Biochem Biophys.
60:219–224. 2011. View Article : Google Scholar
|
|
28
|
Zingarelli B, Salzman AL and Szabó C:
Genetic disruption of poly (ADP-ribose) synthetase inhibits the
expression of P-selectin and intercellular adhesion molecule-1 in
myocardial ischemia/reperfusion injury. Circ Res. 83:85–94. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu N, Zhang X and Jia D: High-dose fasudil
preconditioning and postconditioning attenuate myocardial
ischemia-reperfusion injury in hypercholesterolemic rats. Mol Med
Rep. 9:560–566. 2014. View Article : Google Scholar
|
|
30
|
Hausenloy DJ, Tsang A and Yellon DM: The
reperfusion injury salvage kinase pathway: A common target for both
ischemic preconditioning and postconditioning. Trends Cardiovasc
Med. 15:69–75. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhu H, Ding Y, Xu X, Li M, Fang Y, Gao B,
Mao H, Tong G, Zhou L and Huang J: Prostaglandin E1 protects
coronary microvascular function via the glycogen synthase kinase
3β-mitochondrial permeability transition pore pathway in rat hearts
subjected to sodium laurate-induced coronary microembolization. Am
J Transl Res. 9:2520–2534. 2017.
|
|
32
|
Zhao JL, Yang YJ, You SJ, Cui CJ and Gao
RL: Different effects of postconditioning on myocardial no-reflow
in the normal and hypercholesterolemic mini-swines. Microvasc Res.
73:137–142. 2007. View Article : Google Scholar
|
|
33
|
Iliodromitis EK, Zoga A, Vrettou A,
Andreadou I, Paraskevaidis IA, Kaklamanis L and Kremastinos DT: The
effectiveness of postconditioning and preconditioning on infarct
size in hypercho-lesterolemic and normal anesthetized rabbits.
Atherosclerosis. 188:356–362. 2006. View Article : Google Scholar
|
|
34
|
Donato M, D'Annunzio V, Berg G, Gonzalez
G, Schreier L, Morales C, Wikinski RL and Gelpi RJ: Ischemic
postconditioning reduces infarct size by activation of A1 receptors
and K+(ATP) channels in both normal and hypercholesterolemic
rabbits. J Cardiovasc Pharmacol. 49:287–292. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Iliodromitis EK, Andreadou I, Prokovas E,
Zoga A, Farmakis D, Fotopoulou T, Ioannidis K, Paraskevaidis IA and
Kremastinos DT: Simvastatin in contrast to postconditioning reduces
infarct size in hyperlipidemic rabbits: Possible role of
oxidative/nitrosative stress attenuation. Basic Res Cardiol.
105:193–203. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zeng YC, Hu MY, Qu SL and Zhou GY: Effects
of lycopene on blood lipid and red blood cell of rat with
hypercholesterolemia. Zhonghua Yu Fang Yi Xue Za Zhi. 43:1064–1068.
2009.In Chinese.
|
|
37
|
Paolini M, Abdel-Rahman SZ, Sapone A,
Pedulli GF, Perocco P, Cantelli-Forti G and Legator MS:
Beta-carotene: A cancer chemo-preventive agent or a co-carcinogen.
Mutat Res. 543:195–200. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yeh SL and Hu ML: Induction of oxidative
DNA damage in human foreskin fibroblast Hs68 cells by oxidized
beta-Carotene and lycopene. Free Radic Res. 35:203–213. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rasheva VI and Domingos PM: Cellular
responses to endoplasmic reticulum stress and apoptosis. Apoptosis.
14:996–1007. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Szegezdi E, Logue SE, Gorman AM and Samali
A: Mediators of endoplasmic reticulum stress-induced apoptosis.
EMBO Rep. 7:880–885. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sivaraman V, Mudalagiri NR, Di Salvo C,
Kolvekar S, Hayward M, Yap J, Keogh B, Hausenloy DJ and Yellon DM:
Postconditioning protects human atrial muscle through the
activation of the RISK pathway. Basic Res Cardiol. 102:453–459.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gomez L, Paillard M, Thibault H, Derumeaux
G and Ovize M: Inhibition of GSK3beta by postconditioning is
required to prevent opening of the mitochondrial permeability
transition pore during reperfusion. Circulation. 117:2761–2768.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Miki T, Miura T, Hotta H, Tanno M, Yano T,
Sato T, Terashima Y, Takada A, Ishikawa S and Shimamoto K:
Endoplasmic reticulum stress in diabetic hearts abolishes
erythropoietin-induced myocardial protection by impairment of
phosphoglycogen synthase kinase-3beta-mediated suppression of
mitochondrial permeability transition. Diabetes. 58:2863–2872.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bell RM, Mocanu MM and Yellon DM:
Retrograde heart perfusion: The Langendorff technique of isolated
heart perfusion. J Mol Cell Cardiol. 50:940–950. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hu L, Cheng XW, Song H, Inoue A, Jiang H,
Li X, Shi GP, Kozawa E, Okumura K and Kuzuya M: Cathepsin K
activity controls injury-related vascular repair in mice.
Hypertension. 63:607–615. 2014. View Article : Google Scholar :
|
|
46
|
Cheng Wu H, Hu XW, Takeshita L, Hu K, Du
C, Li Q, Zhu X, Huang E, Yisireyili ZM, et al: Cathepsin S activity
controls injury-related vascular repair in mice via the
TLR2-mediated p38MAPK and PI3K-Akt/p-HDAC6 signaling pathway.
Arterioscler Thromb Vasc Biol. 36:1549–1557. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cheng XW, Shi GP, Kuzuya M, Sasaki T,
Okumura K and Murohara T: Role for cysteine protease cathepsins in
heart disease: Focus on biology and mechanisms with clinical
implication. Circulation. 125:1551–1562. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ogasawara S, Cheng XW, Inoue A, Hu L, Piao
L, Yu C, Goto H, Xu W, Zhao G, Lei Y, et al: Cathepsin K activity
controls cardiotoxin-induced skeletal muscle repair in mice. J
Cachexia Sarcopenia Muscle. 9:160–175. 2018. View Article : Google Scholar
|
|
49
|
Cadenas S: ROS and redox signaling in
myocardial ischemia-reperfusion injury and cardioprotection. Free
Radic Biol Med. 117:76–89. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Andrienko TN, Pasdois P, Pereira GC, Ovens
MJ and Halestrap AP: The role of succinate and ROS in reperfusion
injury-A critical appraisal. J Mol Cell Cardiol. 110:1–14. 2017.
View Article : Google Scholar : PubMed/NCBI
|