Introduction
Gastric cancer is one of the most common malignant
gastrointestinal tumor types globally (1). Numerous novel cases of gastric
cancer occur in developing countries, of which ~50% of patients
live in Eastern Asia and are mainly concentrated in China (2). There is no effective treatment for
the patients in those regions, which seriously affects the survival
of the patients.
Lysophosphatidic acid (LPA) is produced by the
autotoxin hydrolysis of lysophosphatidylcholine (3). As the simplest form of a
glycerophospholipid, LPA is associated with malignant behaviors
including tumor invasion and metastasis, which may inhibit tumor
cell apoptosis and promote its proliferation in situ
(4). LPA receptors are divided
into six subtypes, including LPA1-LPA6. LPA1-LPA3 are members of
the endothelial cell differentiation gene (EDG) family, while
LPA4-LPA6 belong to the purine receptor family (5). When a specific LPA receptor binds to
LPA, it may cause a corresponding biological effect (6). In the research of malignant tumor
types, LPA2 (also known as EDG4) has been widely studied and is
highly expressed in a number of different tumor tissue types,
including breast cancer, liver cancer, gastric cancer and
colorectal cancer (7-9). LPA2 is also involved in biological
behaviors, including proliferation, anti-apoptosis, drug
resistance, metastasis and the invasion of numerous cancer cells,
resulting in poor clinic outcomes (10-13). Furthermore, LPA upregulates the
expression of matrix metalloprotein-9 through activating the
nuclear factor-κB pathway in a LPA2 dependent manner (14).
The Notch signaling pathway is involved in cell
differentiation, proliferation, apoptosis and adhesion (15). It also serves a key function in
maintaining the function of normal cells and tissues. Abnormal
activation of the Notch signaling pathway is associated with the
pathogenesis of a number of different malignancies (16). There are four single-stranded
transmembrane receptors (Notch1-4) in the Notch signaling pathway.
These receptors may be cleaved by γ-secretase following binding to
the ligands Jagged1, Jagged2, Delta1, Delta3 and Delta4 (17). Following binding, the Notch
intracellular domain (NICD) is released and enters the nucleus,
where it stimulates the transcription of downstream target genes,
including Hes Family BHLH Transcription Factor 1 (Hes-1), protein
kinase B (Akt) and Cyclin D1, amongst others (18). Notch1 is abnormally expressed in a
variety of tumor cells and is associated with the poor biological
behavior of malignant tumor types, and the invasion and metastasis
of non-small cell lung cancer (NSCLC) cells. NSCLC cells are
regulated by Notch1, and the downregulation of the Notch1 gene in
SGC-7901 gastric cancer cells inhibits their proliferation and
invasion (19,20). Notch1 is also highly expressed in
gastric cancer tissues and is associated with a poor prognosis
(21).
Invasion and migration are the preconditions for the
metastasis of malignant tumor types (22). Previous studies focus on the
factors that regulate invasion and metastasis in the early stage of
cancer development, with the purpose of providing a reliable
foundation for early diagnosis and treatment (23-25). The epithelial-mesenchymal
transition (EMT) program of tumor cells is closely associated with
invasion and migration (26,27). Cells lose polarity during EMT,
undergo remodification of the cytoskeleton, alter their original
morphology and transform into cells with the capacity to relocate
during EMT (28). During this
process, the expression of the epithelial marker E-Cadherin is
decreased, while the expression of the mesenchymal markers
vimentin, N-cadherin and Snail Family Transcriptional Repressor 1
are increased (29,30).
A previous study has revealed that LPA2 is involved
in the apoptosis, invasion and migration of SGC-7901 cells, and
that the downregulation of LPA2 reduces the expression of Notch1 in
those cells (31). However, the
association between LPA2 and Notch1 remains unclear. The present
study aimed to investigate whether LPA2 and Notch1 were able to
coregulate the invasion and migration of SGC-7901 gastric cancer
cells, to providing novel insight for the study of molecular
pathological diagnosis of gastric cancer cells.
Materials and methods
Cell culture and treatment
GES-1 and SGC-7901 cell lines were obtained from the
Laboratory of Pathology, School of Basic Medicine, Lanzhou
University (Lanzhou, China) (32). Cells were maintained in RPMI-1640
medium (Hyclone; GE Healthcare Life Sciences, Logan, UT, USA) and
supplemented with 1% penicillin/streptomycin (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany), supplemented with 10% fetal bovine serum
(FBS; Biological Industries, Kibbutz Beit Haemek, Israel). All
cells were incubated at 37°C in 5% CO2 and saturated
humidity.
LPA treatment
GES-1 and SGC-7901 cells were seeded in 6-well
plates at a density of 2×105 cells/well, and the
monolayer cells were treated with or without different doses of
Oleoy-L-Alpha-Lysobisphosphatidic acid (Sigma-Aldrich; Merck KGaA)
dissolved in dimethyl sulfoxide. Cells were then collected after
incubation with LPA for 24 h, and analyzed using western
blotting.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from GES-1 and SGC-7901
cells using TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) according to the protocol, and the steps of
extraction were performed on ice. RT-qPCR was performed using
Moloney murine leukemia virus reverse transcriptase (Promega
Corporation, Madison, WI, USA) and random hexamer primers (Takara
Bio, Inc., Otsu, Japan). The conditons for reverse transcription
were set as: 25°C for 10 min; 37°C for 60 min; and 70°C for 15 min.
The resulting cDNAs were used for qPCR. The transcription levels of
mRNAs were quantified by qPCR using SYBR Premix Ex Taq reagents
(Takara Bio, Inc.) in the Mx3005P qPCR System (Agilent
Technologies, Inc., Santa Clara, CA, USA). GAPDH was used as an
internal control. The qPCR primers are listed in Table I. The theromocyling conditions
were set as: 95°C for 2 min (hold stage); 95°C for 10 sec, 60°C for
34 sec (40 cycles, PCR stage); 95°C for 15 sec, 60°C for 1 min,
95°C for 1 sec (melt curve stage). The relative fold changes of
mRNA were calculated using the comparative cycle threshold
(2−ΔΔCq) method (33).
All experiments were repeated three times with similar results. The
data presented represent the results of one of the triplicate
experiments.
 | Table IReverse transcription-quantitative
polymerase chain reaction primers used. |
Table I
Reverse transcription-quantitative
polymerase chain reaction primers used.
| Primers | Sequences | Target gene |
|---|
| LPA1-F |
5′-TGCTTGGGGCCTTTATCATC-3′ | LPA1 |
| LPA1-R |
5′-TTCTCATAGGCCAGCACGTC-3′ | |
| LPA2-F |
5′-ACACTTCTGGCACTGCCTCT-3′ | LPA2 |
| LPA2-R | 5′-AGGCTGAGTGT
GTCTCTCG-3′ | |
| LPA3-F |
5′-TAGGGGCGTTTGTGGTATGC-3′ | LPA3 |
| LPA3-R |
5′-CACCTTTTCACATGCTGCAC-3′ | |
| LPA4-F |
5′-CCATGGGTGACAGAAGATTCA-3′ | LPA4 |
| LPA4-R |
5′-GGCAGTAGCATTGCCCAAC-3′ | |
| LPA5-F |
5′-TCTCTGCTGCTGATGAAGCTG-3′ | LPA5 |
| LPA5-R |
5′-AGGGAGGTCATGGGAATGTG-3′ | |
| LPA6-F |
5′-CCAGCGGAAATTTTACAGCA-3′ | LPA6 |
| LPA6-R |
5′-GCAAATTATCTGGATCTTTGGATG-3′ | |
| Notch1-F |
5′-GCTTGTGGTAGCAAGGAAGC-3′ | Notch1 |
| Notch1-R |
5′-CCACATTCAAGTGGCTGATG-3′ | |
| GAPDH-F |
5′-AAGGTGAAGGTCGGAGTC-3′ | GAPDH |
| GAPDH-R | 5′-TGTAGTTGAGGTCAA
TGAAGG-3′ | |
Western blotting and
co-immunoprecipitation (co-IP) assay
GES-1 and SGC-7901 cells were harvested and lysed
using protein loading buffer (Beijing Solarbio Science &
Technology Co., Ltd., Beijing, China) containing 1% PMSF protease
inhibitors. Firstly, the cells were lysed for 10 min on ice, and
then transferred to centrifuge tubes and heated for 10 min in
boiling water at 100°C. The total protein concentration was
measured using a BCA Protein assay kit (Pierce; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. In
total, 300 μg protein was loaded per lane and separated on
10% SDS-PAGE for 1 h at room temperature. The resolved proteins
were transferred to polyvinylidene difluoride membranes. After 2 h
transfer in cold transfer buffer, the membranes were blocked for 2
h at room temperature with 5% skimmed milk and incubated with
primary antibodies [LPA2 (additionally termed EDG-4), Notch1,
Hes-1, Akt, p-Akt, E-Cadherin, vimentin, F-actin, β-actin]
overnight at 4°C. Membranes were then incubated with secondary
horseradish peroxidase (HRP)-labeled antibodies (goat anti-rabbit
IgG or goat anti-mouse IgG) for 2 h at room temperature and
visual-ized using Electro-Chemi-Luminescence detection reagent
(Thermo Fisher Scientific, Inc.) and Image Lab™ software 4.1
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) was used to select
the optimum exposure blot. The changes in the abundance of proteins
were determined by densitometric analysis using ImageJ Software
1.4.3.67 (National Institutes of Health, Bethesda, MD, USA) and
normalized to β-actin, as previously described (34).
For co-IP assays, SGC-7901 cells were cultured in 6
cm dishes, and the monolayer cells were treated with or without 15
μM LPA. Cells were lysed at 4°C using RIPA Lysis buffer
(Beijing Solarbio Science & Technology Co., Ltd.) for 20 min,
and the lysates were centrifuged at 12,000 × g and 4°C for 10 min.
Co-IP assays were performed using a Co-IP kit (Pierce; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
Briefly, a total of 200 μg protein was incubated with LPA2
and Notch1 antibodies (detailed below) at 4°C for 12 h. Immune
complexes were precipitated with protein A/G sepharose beads and
the complexes were centrifuged at 400 × g at 4°C for 10 min.
Finally, immune complexes were analyzed by western blotting.
Antibodies
The commercial primary antibodies used in the
present study are as follows: Anti-EDG4 antibody (cat. no. 135980;
Abcam, Cambridge, MA, USA), anti-Notch1 antibody (cat. no. 3608;
Cell Signaling Technology, Inc., Danvers, MA, USA), anti-Hes-1
antibody (cat. no. 11988; Cell Signaling Technology, Inc.),
anti-Akt antibody (cat. no. 4691; Cell Signaling Technology, Inc.),
anti-p-Akt antibody (cat. no. 13038; Cell Signaling Technology,
Inc.), anti-E-Cadherin antibody (cat. no. A42; Cell Signaling
Technology, Inc.), anti-vimentin antibody (cat. no. 5741; Cell
Signaling Technology, Inc.), anti-F-actin antibody (cat. no.
130935; Abcam) and anti-β-actin antibody (cat. no. 608407; Thermo
Fisher Scientific, Inc.). All the primary antibodies were diluted
to 1:1,000. The secondary antibodies used in the present study were
as follows: HRP-labeled goat anti-rabbit IgG (cat. no. 97051;
Abcam) and HRP-labeled goat anti-mouse IgG (cat. no. 205719;
Abcam). The secondary antibodies were diluted to 1:5,000.
Wound scratch assay
GES-1 and SGC-7901 cells were seeded in 6-well
plates and cultured to 70-80% confluence. Scrape lines were made
using 200 μl pipette tips, as described previously (35); the initial wound width was at
least 500 nm. Plates were washed twice with phosphate buffered
saline (PBS) and re-incubated with different doses of LPA.
Photographs were taken at 0, 6, 12 and 24 h post-LPA treatment. The
wound healing rate = [Edge distance (0 h)-Edge distance (n h)]/Edge
distance (0 h) ×100%.
Transwell migration and invasion
assays
A total of 2×105 GES-1 or SGC-7901 cells
were suspended in serum-free RPMI-1640 medium with or without
Matrigel (BD Biosciences, Franklin Lakes, NJ, USA; diluted in a 1:8
proprotion) in the upper chambers (Corning Inc., Corning, NY, USA)
and LPA or RPMI 1640 medium containing 5% FBS were also added into
24-well plates (lower chambers), and the incubation time was 24 h.
Cells were then washed with PBS, fixed using 4% paraformaldehyde
for 15 min at the room temperature and stained using 0.1% crystal
violet for 10 min at the room temperature. The cells that were
unattached to the trans membrane were removed using cotton swabs,
as previously described (25).
The cells that passed through the membrane and were located in five
randomly selected light microscopic fields (magnification, ×200)
per chamber were then imaged and counted.
MTS assay
GES-1 and SGC-7901 cells were seeded in 96-well
plates at a density of 1×103 per well. Following
incubation for 24 h, the cells were washed using PBS and treated
with different concentrations of LPA. Cell Titer96®AQueous One
Solution reagent (Promega Corporation) was added continuously,
according to the manufacturer's protocol. Following incubation for
3 h at 37°C and 5% CO2, the absorbance value (optical
density value) at 490 nm were recorded on a microplate reader and
cell viability was calculated, using the following formula: [Cell
viability (%)=(A experiment-A blank)/(A control-A blank) ×100].
Transfection with small interfering RNA
(siRNA)
SGC-7901 cells cultured in 6-well plates at a
density of 2×105 cells per well. When the cells reached
70-80% confluence, 150 nM LPA2 or Notch1 siRNA (Shanghai GenePharma
Co., Ltd., Shanghai, China) were transfected into cells, and
Lipofectamine 2000® (Thermo Fisher Scientific, Inc.) was used,
according to the manufacturer's protocol. The cells were incubated
at 37°C in 5% CO2 and saturated humidity for 24 h. In
total, 150 nM negative control (NC) siRNA was used as the negative
control. The sequences of the siRNAs are listed in Table II.
| Table IIsiRNAs used in the present study. |
Table II
siRNAs used in the present study.
| siRNA | Sequence |
|---|
| Negative
control | Sense:
5′-AUUGACCAGUGAGUUGGCCTT-3′ |
| Antisense:
5′-GCGAGUCUGUCCACUAUACTT-3′ |
| si-LPA2 | Sense:
5′-AUUGACCAGUGAGUUGGCCTT-3′ |
| Antisense:
5′-GCGAGUCUGUCCACUAUACTT-3′ |
| si-Notch1 | Sense:
5′-UCGCAUUGACCAUUCAAACUGGUGG-3′ |
| Antisense:
5′-CCACCAGUUUGAAUGGUCAAUGCGA-3′ |
Indirect immunofluorescence microscopy
assay (IFA)
The IFA assay was performed as described previously
(36). Briefly, SGC-7901 cells
were plated into Nunc glass-bottom dishes (Thermo Fisher
Scientific, Inc.) and treated with LPA for 24 h or fresh RPMI-1640
medium containing 10% FBS. Cells were washed at 37°C with PBS and
fixed using 4% paraformaldehyde for 15 min at room temperature.
Cells were then incubated with fluorescein isothiocyanate
(FITC)-phalloidin (1:200; Beijing Solarbio Science & Technology
Co., Ltd.) overnight at 4°C. Then, cells were incubated with DAPI
for 30 sec at the room temperature. Dishes were analyzed and imaged
on a fluorescent microscope (magnification, ×63).
Statistical analysis
Statistical analyses were performed using GraphPad
Prism software 7 (GraphPad Software, Inc., La Jolla, CA, USA).
Differences between experimental groups were assessed using one-way
analysis of variance and Tukey's post-hoc test. Student's t-test
was used to examine the statistical differences between two groups
P<0.05 was considered to indicate a statistically significant
difference. The results are presented as the mean ± the standard
error of the mean.
Results
Determination of the experimental LPA
concentration
To select the concentration of LPA that is able to
induce metastasis and invasion, wound scratch assays, transwell
migration and invasion assays were performed. SGC-7901 gastric
cancer cells were selected as the experimental cells, and the
normal gastric epithelial cells (GES-1) were selected as the
control. The wound healing rate reached the maximum at an LPA
concentration of 15 μM (Fig.
1A and B). The number of SGC-7901 cells that traversed the
membrane were significantly increased at 15 μM LPA in the
transwell migration and invasion assays compared with the control
(untreated) group (P<0.01; Fig.
1C-H). MTS assays were also performed to confirm the impact of
LPA on the viability of cells (Fig.
1I) and it was revealed that 15 μM LPA did not affect
the viability of the cells. Therefore, the 15 μM LPA and a
duration time is 24 h were used for subsequent experiments.
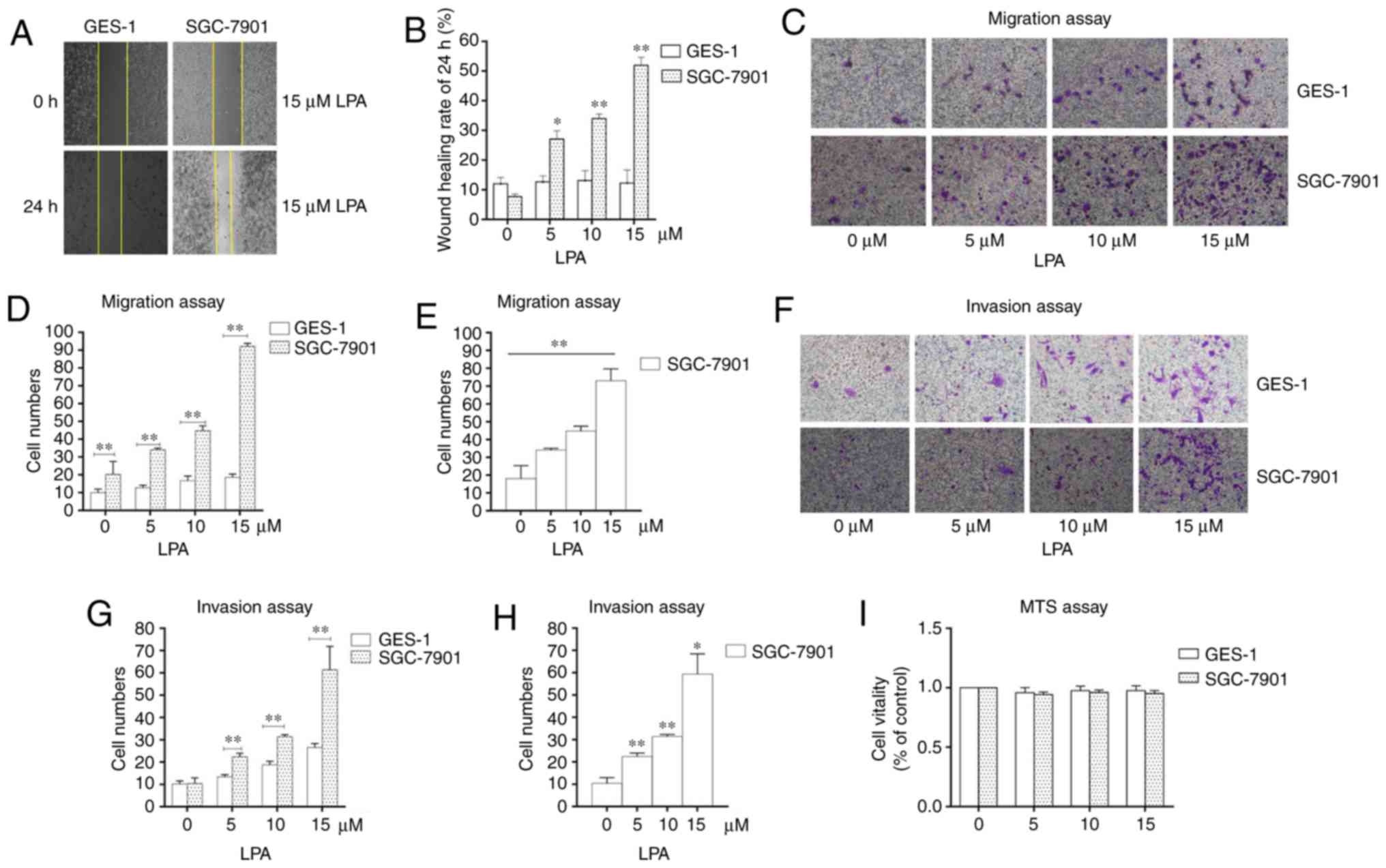 | Figure 1Identifying the experimental
concentration of LPA. (A) Wound healing assays were performed using
SGC-7901 and GES-1 cells which were incubated with 0, 5, 10 and 15
μM LPA. Photographs were obtained at 0, 6, 12 and 24 h after
LPA treatment. (B) Quantified wound healing rate. (C) Migration
assay for SGC-7901 and GES-1 cells traeted with different doses (0,
5, 10 and 15 μM) of LPA. (D and E) Counting the number of
transmembrane cells in the migration assay. (F) Invasion assay for
SGC-7901 and GES-1 cells traeted with different doses (0, 5, 10 and
15 μM) of LPA. (G and H) Counting the number of
transmembrane cells in the invasion assay. (I) Cells were seeded in
96-well plates, and the monolayer cells were treated with different
doses of LPA for 24 h. The cytotoxicity of LPA was measured using
MTS assays. These results are representative of three independent
experiments. *P<0.05 and **P<0.01 with
comparisons shown by lines. LPA, lysophosphatidic acid. |
LPA stimulates LPA2, activating the Notch
signaling pathway in SGC-7901 cells
A previous study revealed that the migration
behavior of SGC-7901 gastric cancer cells was regulated by LPA via
the LPA/LPA2/Gq/11/p38 pathway (37). However, whether LPA induced the
migration and invasion of SGC-7901 cells by LPA2 and Notch pathways
remained unknown. The present study therefore evaluated the mRNA
expression levels of LPA1-6 in SGC-7901 and GES-1 cells using
RT-qPCR. The expression of LPA2 was most significantly enhanced in
SGC-7901 cells compared with GES-1 cells (P<0.05; Fig. S1).
To identify whether the Notch pathway was involved
in the LPA-induced migration and invasion of SGC-7901 cells, the
mRNA levels and the abundance of LPA2 and Notch1 in SGC-7901 and
GES-1 cells were compared. It was revealed that the expression of
LPA2 and Notch1 mRNA and protein levels were significantly higher
in SGC-7901 cells compared with GES-1 cells (P<0.05; Fig. 2A and B). In addition, the SGC-7901
cells were incubated with LPA for 24 h, cells were collected and
analyzed by western blotting. The results revealed that the
abundance of LPA2, Notch1, Hes-1 and phosphorylated-Akt were
significantly enhanced in LPA-treated cells compared with that in
LPA-untreated cells (P<0.05; Fig.
2C). In addition, the mRNA levels of LPA2 and Notch1 were also
signficantly increased following LPA treatment (P<0.05; Fig. 2D). Collectively, these results
indicate that LPA activates LPA2 and Notch signaling pathways.
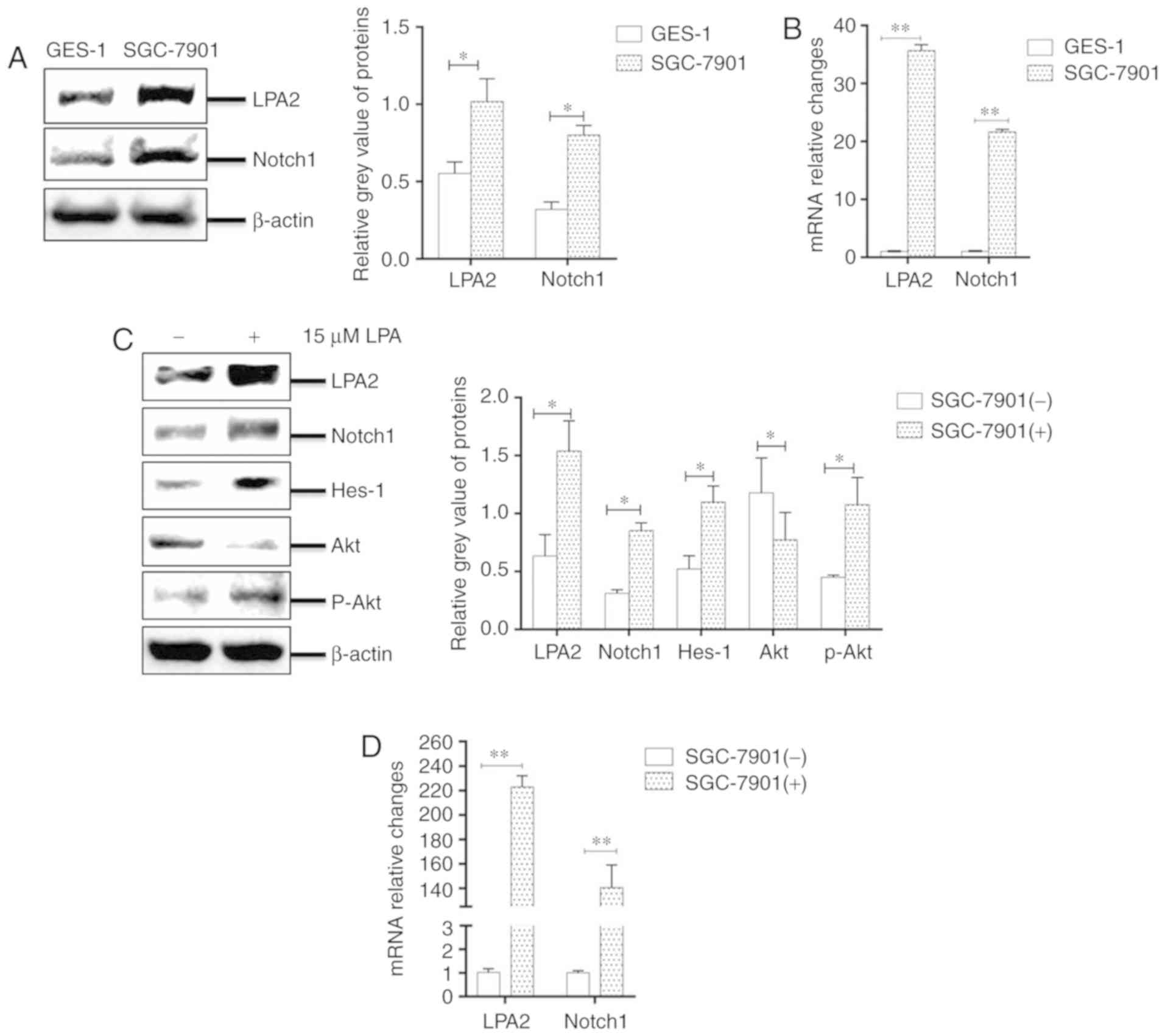 | Figure 2LPA stimuluates LPA2, activating the
Notch signaling pathway in SGC-7901 cells. Expression of LPA2 and
Notch1 were determined in GES-1 and SGC-7901 cells by (A) western
blotting and (B) RT-qPCR. Expression of LPA2, Notch1, Hes-1, Akt
and p-Akt were determined in SGC-7901 cells by (C) western
blotting, and (D) the mRNA levels of LPA2 and Notch1 were
determined by RT-qPCR. (-), cells treated without 15 μM LPA;
(+), cells treated with 15 μM LPA. The blots with tracks
form different exposures. These results are representative of three
independent experiments. *P<0.05 and
**P<0.01 with comparisons shown by lines. LPA,
lysophosphatidic acid; RT-qPCR, reverse transcription-quantitative
polymerase chain reaction; Hes-1, Hes Family BHLH Transcription
Factor 1; Akt, protein kinase B; p-, phosphorylated. |
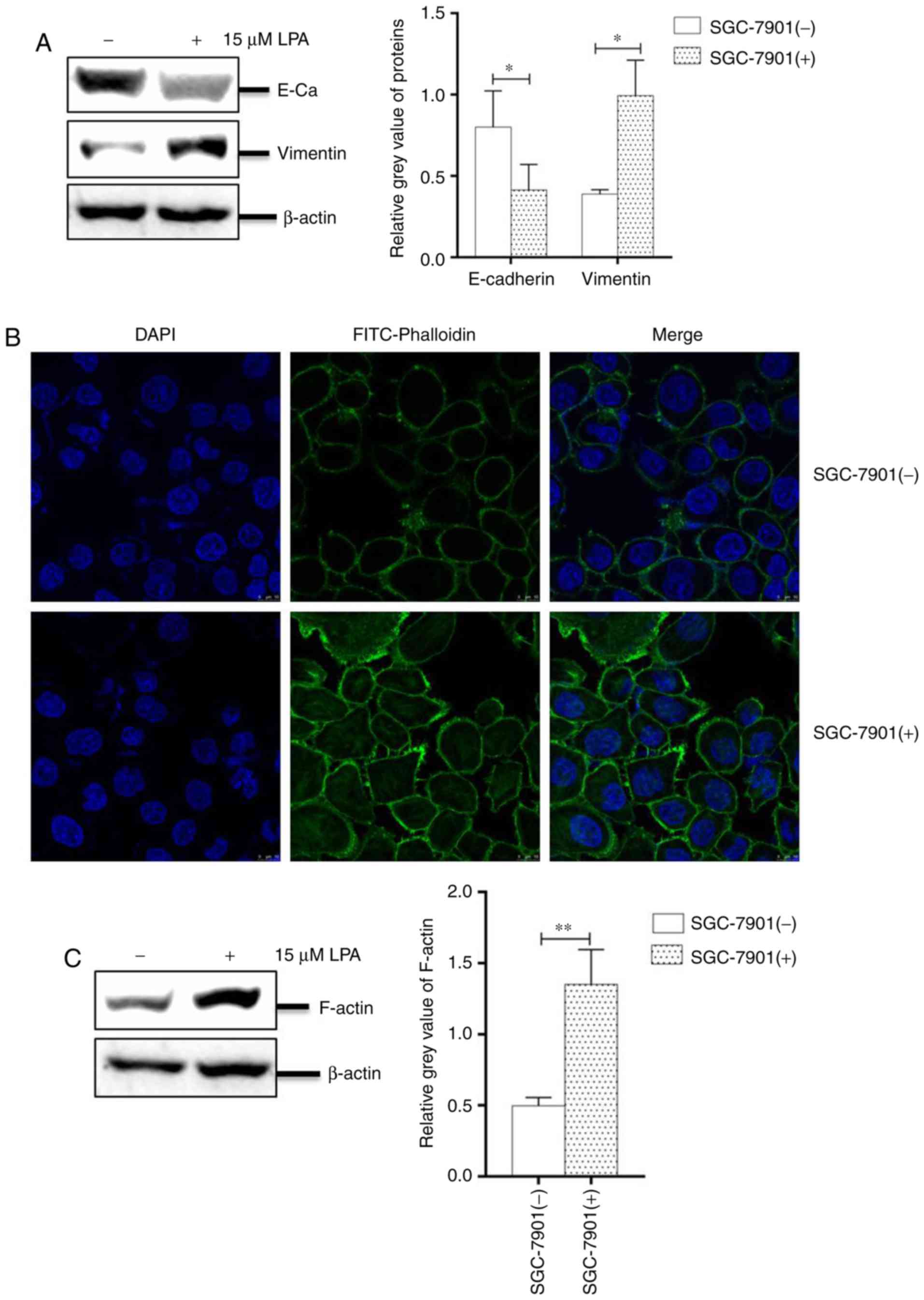
LPA induced EMT and remodeling of the
cytoskeleton in SGC-7901 cells
The progress of EMT is crucial for the migration and
invasion in numerous malignant tumor types (38). Under an oncogenic stimulant, the
expression of E-cadherin is decreased, whereas the expression of
vimentin is increased, resulting in an untethering of cell-cell
adhesion. In addition, cellular morphology and fibrous actin
(F-actin) will be altered during EMT (39).
To investigate the EMT progress, SGC-7901 cells
cultured in 6-well plates were treated with or without LPA for 24
h. The collected cells were analyzed by western blotting and IFA.
The results revealed that the abundance of E-cadherin was
significantly reduced, while the expression of vimentin was
significantly increased compared with untreated cells (P<0.05;
Fig. 3A). The IFA assay also
revealed numerous filaments (cytoskeletal fibers) on the surface of
cells and filamentous bulges (pseudopodium) surrounding LPA-treated
cells (Fig. 3B). To confirm this
effect, the present study analyzed the expression of F-actin and
revealed that its levels were significantly increased in
LPA-treated SGC-7901 cells (P<0.01; Fig. 3C). Collectively, these results
indicated that LPA induces EMT progression and remodels the
cytoskeleton of SGC-7901 cells.
Effects of LPA2 knockdown on cellular
functions induced by LPA treatment in SGC-7901 cells
LPA2 is a mediator of the biological behaviors of
numerous cancer cell types (40,41). To investigate the functions of
LPA2 on LPA-induced SGC-7901 cells, cells were transfected with
LPA2 or NC siRNA for 24 h, then treated with or without LPA. The
cells were collected and analyzed by western blotting, qPCR, IFA,
transwell migration and invasion assays. The results revealed that
the expression of LPA2, Notch1, Hes-1 and Akt were significantly
decreased in the LPA2 siRNA-transfected cells compared with that in
the NC siRNA-transfected cells (P<0.05). In addition, the
expression of LPA2, Notch1, Hes-1 and p-Akt were enhanced following
LPA treatment (Fig. 4A and B).
The transwell migration (Fig. 4C)
and invasion assays (Fig. 4D)
revealed that the cells that transited membranes were significantly
fewer in the si-LPA2 group compared with that in the NC group
(P<0.01). When treated with LPA, the cell numbers that transited
the chambers were higher in the NC group compared with the si-LPA2
group.
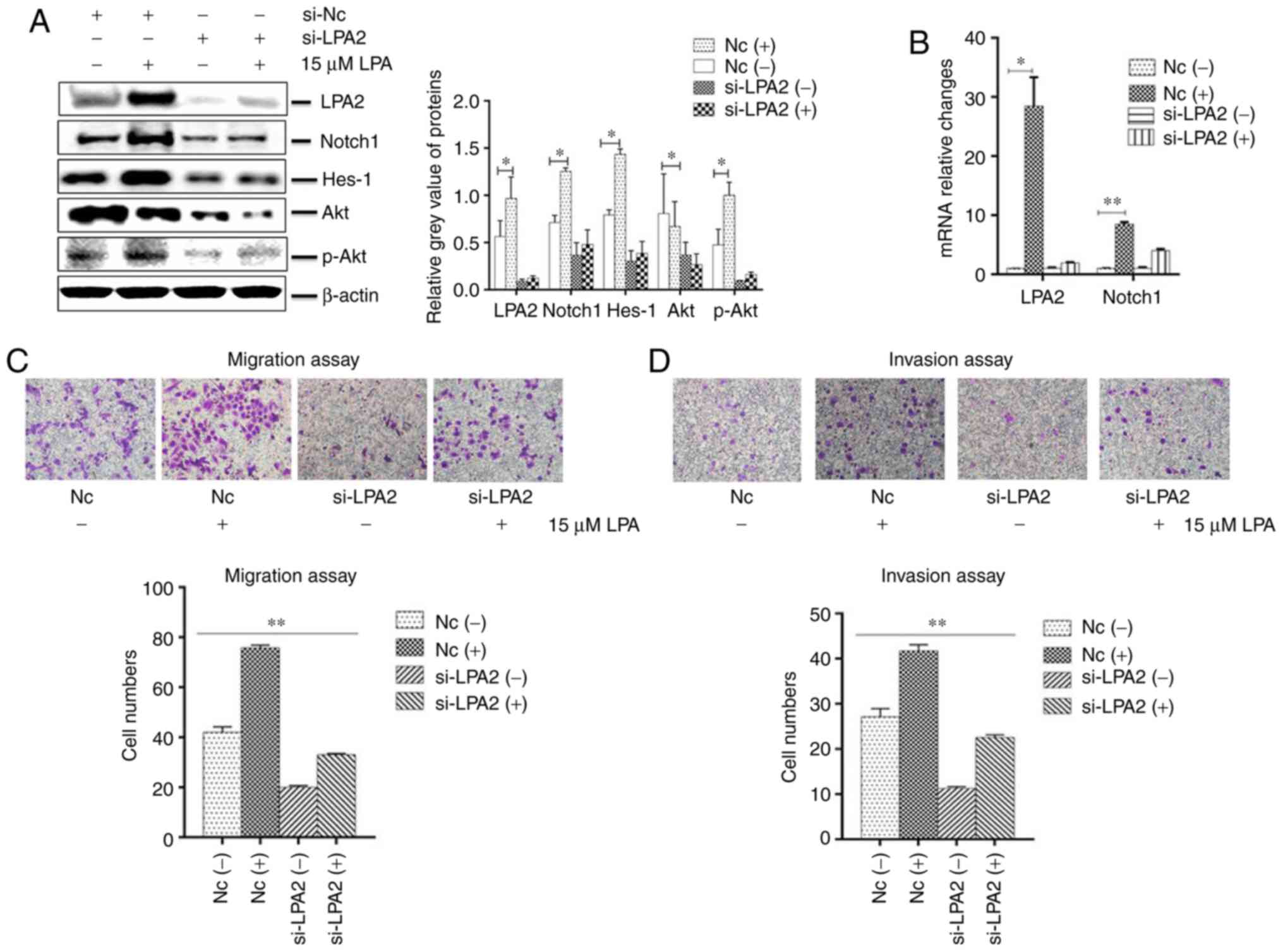 | Figure 4Effects of LPA2 knockdown on cellular
functions induced by LPA treatment in SGC-7901 cells. SGC-7901
cells were seeded in 6-well plates, and the monolayer cells were
transfected with 150 nM LPA2 or NC siRNA for 24 h. Following
transfection cells were then treated with or without 15 μM
LPA for 24 h. Cells were then collected and analysed by (A) western
blotting and (B) reverse transcription-quantitative polymerase
chain reaction. (C) Transwell migration and (D) invasion assays
were performed. (E) Expression of E-Cadherin and vimentin by
western blotting. (F and G) Similar RNA transfection was performed
as described above. Following 24 h transfection, cells were treated
with or without 15 μM LPA for 24 h. Then indirect
immunofluorescence microscopy assay experiments were performed. (H)
Similar transfections were performed as described above, and cells
cultured in 6-well plates were treated with or without 15 μM
LPA for 24 h. Cells were then collected and the expression of
F-actin was analysed by western blotting. (-), cells treated
without 15 μM LPA; (+), cells treated with 15 μM LPA.
The blots with tracks form different exposures. These results are
representative of three independent experiments.
*P<0.05 and **P<0.01 with comparisons
shown by lines. LPA, lysophosphatidic acid; NC, negative control;
Hes-1, Hes Family BHLH Transcription Factor 1; Akt, protein kinase
B; p-, phosphorylated; si-, small interfering RNA; FITC,
fluorescein isothiocyanate. |
These results also demonstrated that the abundance
of E-cadherin in the si-LPA2 group was markedly increased
(P<0.05), while vimentin was decreased compared with the NC
group (P<0.05). In addition, E-cadherin was decreased and
vimentin was increased following LPA treatment (Fig. 4E). IFA revealed that the
cytoskeleton (Fig. 4F and G) and
expression of F-actin (Fig. 4H)
were significantly enhanced in the NC siRNA-transfected cells
following LPA treatment compared with the LPA un-treated group
(P<0.05), and no clear changes to the cytoskeleton (Fig. 4F and G) or in the expression of
F-actin (Fig. 4H) were observed
in the LPA2 siRNA-transfected cells when treated with or without
LPA. Altogether, these results indicated that LPA2 served a key
function in mediating EMT progression and regulating the
cytoskeleton of SGC-7901 cells.
Effects of Notch1 knockdown on cellular
functions induced by LPA treatment in SGC-7901 cells
The expression of Notch1 was reduced in SGC-7901
cells with downregulated LPA2. Therefore, the present study
investigated whether Notch1 may affect the migration and invasion
of SGC-7901 cells. To do so, Notch1 or NC siRNA was transfected
into SGC-7901 cells and then treated with or without LPA. The
collected cells were analyzed by western blotting, qPCR, IFA,
transwell migration and invasion assays. The results revealed that
the expression of LPA2, Notch1, Hes-1 and Akt were decreased in the
Notch1 siRNA-transfected cells compared with that in the NC-siRNA
transfected cells. In addition, the expression of LPA2, Notch1,
Hes-1 and p-Akt were enhanced following LPA treatment (P<0.05;
Fig. 5A and B). The present study
additionally investigated the function of Notch1 on migration and
invasion in vitro. The cells which transited the bottom of
chambers were significantly fewer in si-Notch1 group compared with
the NC group (P<0.01), and the trans-membrane cells that were
counted were elevated in the NC group once LPA was added (Fig. 5C and D).
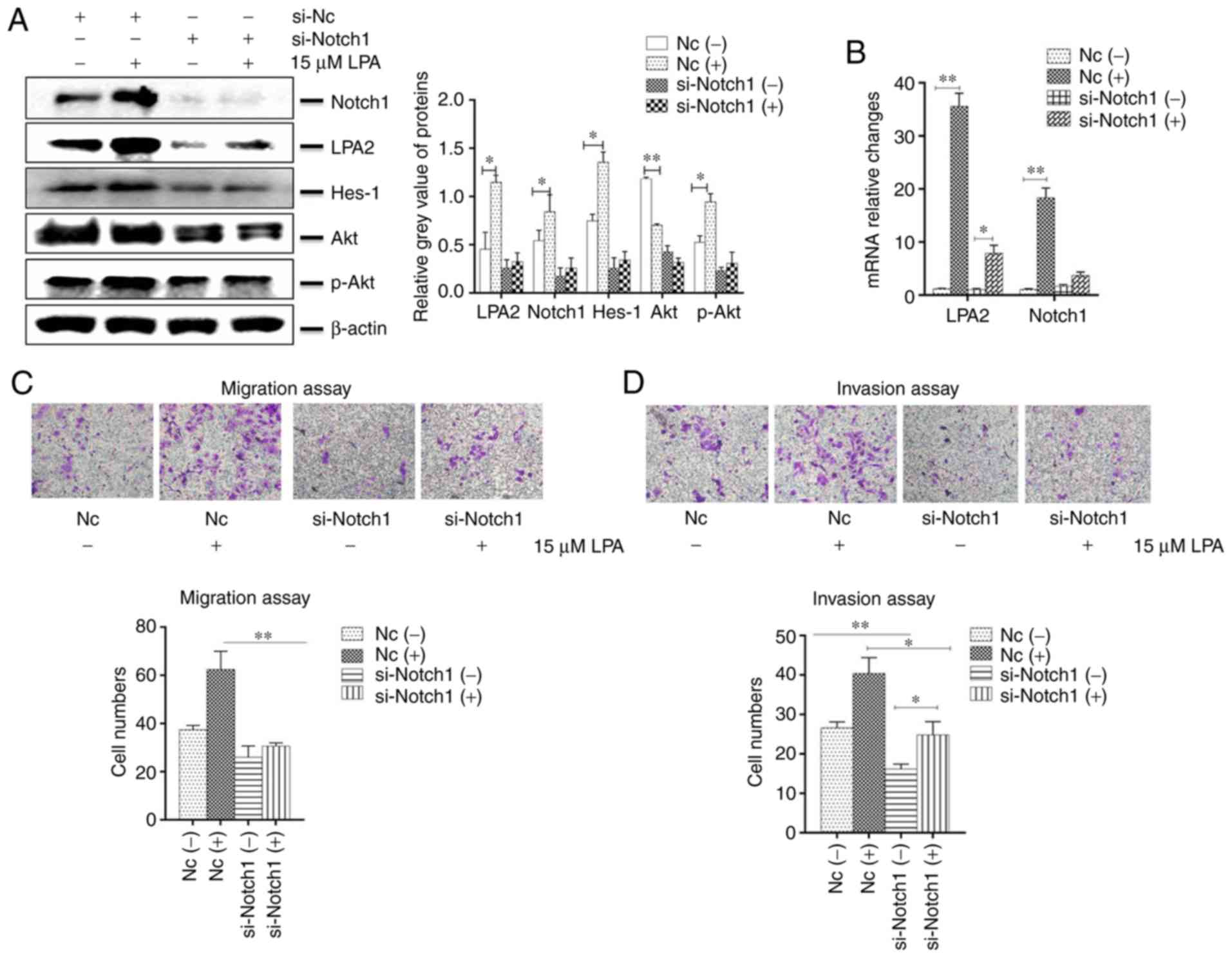 | Figure 5Effects of Notch1 knockdown on
cellular functions induced by LPA treatment in SGC-7901 cells.
SGC-7901 cells were seeded in 6-well plates, and the monolayer
cells were transfected with 150 nM Notch1 or NC siRNA for 24 h.
Transfected cells were then treated with or without 15 μM
LPA for 24 h, and collected for analysis by (A) western blotting
and (B) reverse transcription-quantitative polymerase chain
reaction. Transfections were performed as described above and (C)
transwell migration and (D) invasion assays were performed. (E)
Expression of E-Cadherin and vimentin were determined by western
blotting. (F and G) Similar RNA transfection was performed as
described above. Following 24 h transfection, cells were treated
with or without 15 μM LPA for 24 h. Then indirect
immunofluorescence microscopy assay experiments were performed. (H)
Expression of F-actin by western blotting. (-), cells treated
without 15 μM LPA; (+), cells treated with 15 μM LPA.
The blots with tracks form different exposures. These results are
representative of three independent experiments.
*P<0.05 and **P<0.01 with comparisons
shown by lines. LPA, lysophosphatidic acid; NC, negative control;
Hes-1, Hes Family BHLH Transcription Factor 1; Akt, protein kinase
B; p-, phosphorylated; si-, small interfering RNA; E-Ca,
E-cadherin; FITC, fluorescein isothiocyanate. |
The abundance of E-cadherin in the si-Notch1 group
was increased and vimentin was decreased, compared with the NC
group, while the expression of E-cadherin was decreased and
vimentin was increased, following treatment with LPA (P<0.05;
Fig. 5E). IFA revealed that the
cytoskeleton (Fig. 5F and G) and
expression of F-actin (Fig. 5H)
were significantly increased in the NC siRNA-transfected cells
following LPA treatment (P<0.05), and no substantial changes to
the cytoskeleton (Fig. 5F and G)
or F-actin expression (Fig. 5H)
in the si-Notch1 group occured. Altogether, these results suggest
that Notch1 functions as a mediator in regulating the EMT progress
and regulating the cytoskeleton of SGC-7901 cells.
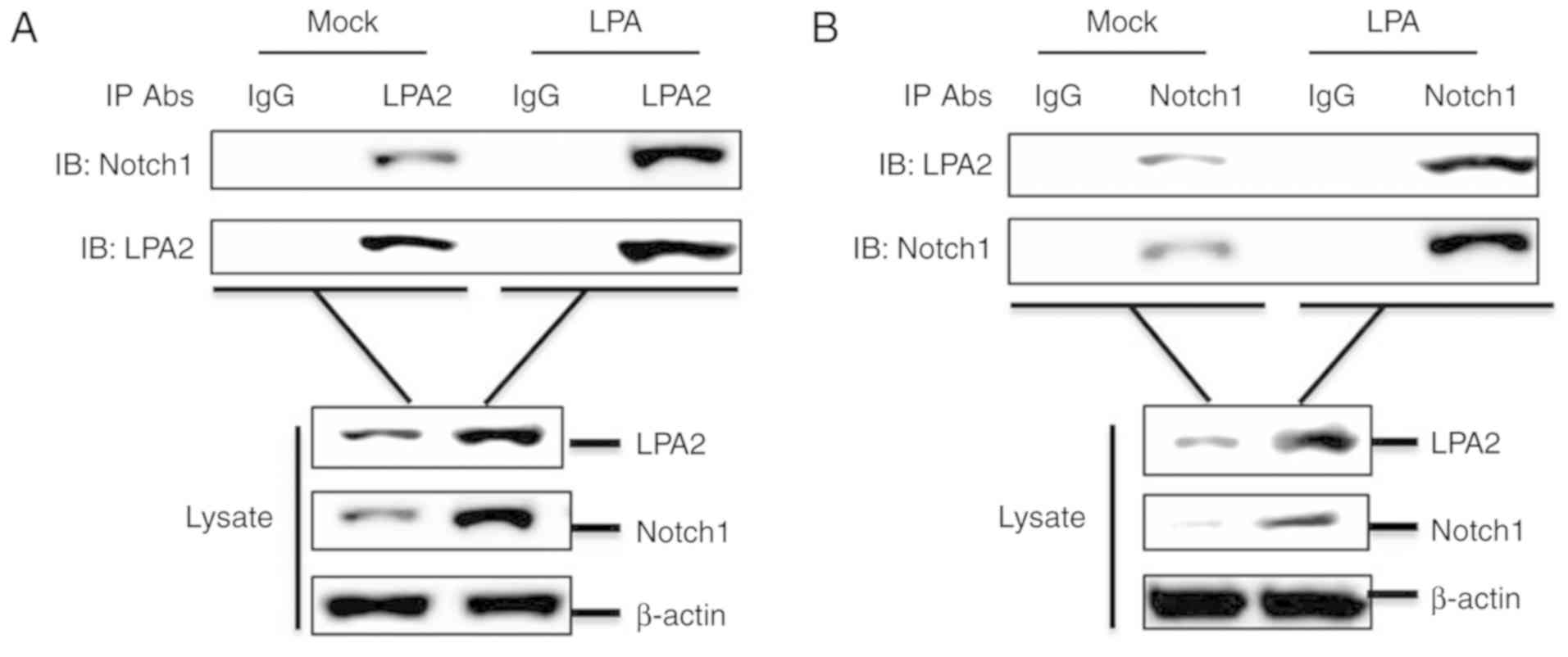
LPA2 interacts with Notch1
To investigate a potential interaction between LPA2
and Notch1, a co-IP assay was performed. SGC-7901 cells cultured in
60-mm-dishes were treated with or without 15 μM LPA for 24
h. The cells were lysed and the lysates were collected, then the
compounds were immunoprecipitated with anti-LPA2 antibody and
analyzed by western blotting. LPA2 pulled down Notch1 (Fig. 6A). A reverse immunoprecipitation
experiment was also performed using an anti-Notch1 antibody, which
revealed that Notch1 also pulled down LPA2 (Fig. 6B). In addition, LPA promoted the
interaction between LPA2 and Notch1 (Fig. 6A and B). Altogether, these results
indicate that the interaction between LPA2 and Notch1 is involved
in the migration and invasion of SGC-7901 cells.
Discussion
To date, numerous studies have indicated that LPA is
at high levels in a multitude of malignant tumor types, including
ovarian cancer, liver cancer and oral squamous cell carcinoma
(42-44). LPA may also promote tumor
angiogenesis, providing nutrients to tumor tissues, rebuild the
cytoskeleton and promote the motor ability of tumor cells. LPA also
functions as a biomarker for gastric cancer cases with metastasis
(45).
LPA2 is involved in numerous oncologic signal
pathways, particularly in the microenvironment of rich-LPA
(46). A previous study
demonstrated that over-expressing LPA2 (with no treatment with LPA)
may accelerate the proliferation, invasion, migration and
anti-apoptosis of SGC-7901 cells (31). In the present study, the results
indicated that the reduction of LPA2 (with no treatment with LPA)
also affected the invasion and migration of SGC-7901 cells.
Altogether, the results indicate that LPA2 alone may affect the
aggressiveness of cells. Notch1 is a key element of the Notch
pathway, and regulates the functions of numerous cancer cell
phenotypes (47,48). The Notch pathway is also involved
in the drug-resistance of gastric cancer cells (49). Numerous factors are associated
with the risk of gastric cancer disease-specific mortality, which
may result from interacting or opposing cell signaling pathways.
However, there are no clear links between LPA2 and Notch1.
EMT progression is a multi-step biological process
and is characterized by a rebuilding of the cytoskeleton, allowing
cancer cells to migrate and invade. The regulation of EMT relies on
a complex network of signaling pathways (50). During EMT progression, cell-cell
adhesion is affected by cytoskeleton alterations and cellular
morphology is altered. Previous studies have revealed that Notch1
controls the EMT of breast cancer cells in a snail family
transcriptional repressor 2-dependent manner (51), and that LPA2 meditates the
cell-cell and cell-matrix of HEC1A cells (52). Nevertheless, the mechanisms by
which LPA2 and Notch1 co-regulate EMT progression are not well
known.
The LPA-LPA2 axis has been implicated in the
pathology of human gastric cancer cells (37,38,53), and the present study focused on
the pathways via LPA2 and Notch1 induced migration and invasion of
gastric cancer cells. In the present study, the results revealed
that following LPA treatment, the expression of LPA2, Notch1 and
Hes-1 were enhanced in SGC-7901 cells, and the phosphorylation of
Akt was increased, which indicated that LPA may activate Notch
pathway through LPA2. There are multiple chemokines in the
microenvironment in which cells existed, which drives cells to
transform into the mesenchymal phenotypes (54). Western blotting and IFA assays
revealed that LPA triggered EMT progression by regulating cellular
morphology, the cytoskeleton and pseudopods of SGC-7901 cells. In
colon cancer cells, LPA2 was highly expressed in HCT116 cells, and
when RNA interference was used to knockdown the LPA2 in HCT116
cells, the proliferation and tumor formation were attenuated
(55). Another study revealed
that the migration and invasion of ovarian cancer cells SKOV-3 were
determined by LPA2, and the aggressiveness of SKOV-3 cells may be
strengthened by the stimulation of LPA (56). Sun et al (57) indicated that overexpression Notch1
affected the progression of cervical cancer, and the reduction of
Notch1 protein in the Hela, SiHa, C33A, Caski and HT-3 cells
decreased the malignant behaviors of these cells. The present study
contributed to the use of si-LPA2 and si-Notch1 siRNAs to identify
the mechanisms and functions of LPA2 and Notch1. SGC-7901 cell
migration and invasion were suppressed by LPA2 knockdown, similar
to that observed with Notch1 downregulation. In the
siRNA-transfected SGC-7901 cells, the expression of Hes-1 was
reduced, and the process of Akt phsophorylation was inhibited, and
there were no changes when LPA was added. The function of the Notch
pathway was affected in the siRNAs transfected cells, and a series
of detailed analyses indicated that LPA2 and Notch1 function as
molecular switches in controlling the malignant behaviors of
SGC-7901 gastric cancer cells.
Zang et al (49) revealed that the cleaved form of
Notch1 may interact with β-catenin and promote the proliferation,
migration and inhibition of cell apoptosis of gastric cancer cells.
Another study revealed that LPA2-mediated signal transduction may
be achieved through G protein-activated signaling cascades and the
interacting partner-meditated signaling pathways (58). In the present study, Notch
signaling was predicted to be a potential target of LPA by the
endogenous gastric cancer cells network model, and the regulation
of the LPA-LPA2-Notch pathway was hypothesized to serve a crucial
function in the malignant behavior of SGC-7901 cells. So the
present study focus on the association between LPA2 and Notch1 in
SGC-7901 cells. To further verify our results, co-IP assays were
used to confirm the interaction between LPA2 and Notch1. The
results confirm that when exogenous LPA was added as a
chemoattractant, the invasion and migration behav-iors of SGC-7901
cells may be substantially promoted. The specific mechanism is that
LPA binds to its receptor LPA2, and recruits Notch1 to accelerate
EMT. In addition, the Notch pathway was also activated following
LPA treatment, thus the malignant behaviors of the cells were
further strengthened, and in the SGC-7901 cells in which the dual
molecules were downregulated also demonstrated increased migration
and invasion abilities, which may be attributed to the interaction
between LPA2 and Notch1.
In the present study, the transwell migration and
invasion assays revealed that the cells which transited chambers
were substantially fewer in si-LPA2 and si-Notch1 groups compared
with si-LPA2 or si-Notch1 groups without treatment (Fig. S2A-B), which indicated that LPA2
and Notch1 may co-regulate the migration and invasion of SGC-7901
cells.
In conclusion, the present results provide a
theoretical basis for the research of gastric cancer cells in
molecular pathology, and further studies are required to determine
the comprehensive mechanisms by which the combination of LPA2 with
Notch1 function as diagnostic makers of gastric cancer.
Supplementary Materials
Funding
The present study was supported by grants from the
Gansu Science Foundation (grant nos. 1606RJDA313 and 18JR3RA293)
and the Lanzhou Science and Technology planning project (grant nos.
2017-4-64 and 2018-3-44).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
ZR, CZ, LM, XiZ, SS, DT, JX, YH, BW and FZ performed
the experiments. ZR and CZ contributed to the data analysis and
wrote the manuscript. XuZ and HZ contributed to the study design
and concept, and contributed to the experiment materials. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare no that they have are no
competing interests.
Acknowledgments
Not applicable.
References
|
1
|
Charalampakis N, Economopoulou P,
Kotsantis I, Tolia M, Schizas D, Liakakos T, Elimova E, Ajani JA
and Psyrri A: Medical management of gastric cancer: A 2017 update.
Cancer Med. 7:123–133. 2018. View Article : Google Scholar :
|
|
2
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar
|
|
3
|
Tabuchi S: The autotaxin-lysophosphatidic
acid-lysophosphatidic acid receptor cascade: Proposal of a novel
potential therapeutic target for treating glioblastoma multiforme.
Lipids Health Dis. 14:562015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mills GB and Moolenaar WH: The emerging
role of lysophosphatidic acid in cancer. Nat Rev Cancer. 3:582–591.
2003. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kihara Y, Maceyka M, Spiegel S and Chun J:
Lysophospholipid receptor nomenclature review: IUPHAR review 8. Br
J Pharmacol. 171:3575–3594. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee SC, Fujiwara Y, Liu J, Yue J, Shimizu
Y, Norman DD, Wang Y, Tsukahara R, Szabo E, Patil R, et al:
Autotaxin and LPA1 and LPA5 receptors exert disparate functions in
tumor cells versus the host tissue microenvironment in melanoma
invasion and metastasis. Mol Cancer Res. 13:174–185. 2015.
View Article : Google Scholar
|
|
7
|
Li M, Xiao D, Zhang J, Qu H, Yang Y, Yan
Y, Liu X, Wang J, Liu L, Wang J and Duan X: Expression of LPA2 is
associated with poor prognosis in human breast cancer and regulates
HIF-1α expression and breast cancer cell growth. Oncol Rep.
36:3479–3487. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Enooku K, Uranbileg B, Ikeda H, Kurano M,
Sato M, Kudo H, Maki H, Koike K, Hasegawa K, Kokudo N and Yatomi Y:
Higher LPA2 and LPA6 mRNA levels in hepatocellular carcinoma are
associated with poorer differentiation, microvascular invasion and
earlier recurrence with higher serum autotaxin levels. PLoS One.
11:e01618252016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yun CC, Sun H, Wang D, Rusovici R,
Castleberry A, Hall RA and Shim H: LPA2 receptor mediates mitogenic
signals in human colon cancer cells. Am J Physiol Cell Physiol.
289:C2–C11. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mukherjee A, Ma Y, Yuan F, Gong Y, Fang Z,
Mohamed EM, Berrios E, Shao H and Fang X: Lysophosphatidic acid
up-regulates hexokinase II and glycolysis to promote proliferation
of ovarian cancer cells. Neoplasia. 17:723–734. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang R, Wang J, Ma S, Huang Z and Zhang
G: Requirement of Osteopontin in the migration and protection
against Taxol-induced apoptosis via the ATX-LPA axis in SGC7901
cells. BMC Cell Biol. 12:112011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Venkatraman G, Benesch MG, Tang X, Dewald
J, McMullen TP and Brindley DN: Lysophosphatidate signaling
stabilizes Nrf2 and increases the expression of genes involved in
drug resistance and oxidative stress responses: Implications for
cancer treatment. FASEB J. 29:772–785. 2015. View Article : Google Scholar
|
|
13
|
Yamashita H, Kitayama J, Shida D, Ishikawa
M, Hama K, Aoki J, Arai H and Nagawa H: Differential expression of
lysophosphatidic acid receptor-2 in intestinal and diffuse type
gastric cancer. J Surg Oncol. 93:30–35. 2006. View Article : Google Scholar
|
|
14
|
Gu C, Wang F, Zhao Z, Wang H, Cong X and
Chen X: Lysophosphatidic acid is associated with atherosclerotic
plaque instability by regulating NF-κB dependent matrix
metallopro-teinase-9 expression via LPA2 in macrophages.
Front Physiol. 8:2662017. View Article : Google Scholar
|
|
15
|
Dang TP: Notch, apoptosis and cancer. Adv
Exp Med Biol. 727:199–209. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hu YY, Zheng MH, Zhang R, Liang YM and Han
H: Notch signaling pathway and cancer metastasis. Adv Exp Med Biol.
727:186–198. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rizzo P, Osipo C, Foreman K, Golde T,
Osborne B and Miele L: Rational targeting of Notch signaling in
cancer. Oncogene. 27:5124–5131. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yamamoto S, Schulze KL and Bellen HJ:
Introduction to Notch signaling. Methods Mol Biol. 1187:1–14. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yuan Q, Chen X, Han Y, Lei T, Wu Q, Yu X,
Wang L, Fan Z and Wang S: Modification of α2,6-sialylation mediates
the invasiveness and tumorigenicity of non-small cell lung cancer
cells in vitro and in vivo via Notch1/Hes1/MMPs pathway. Int J
Cancer. 143:2319–2330. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wei G, Chang Y, Zheng J, He S, Chen N,
Wang X and Sun X: Notch1 silencing inhibits proliferation and
invasion in SGC7901 gastric cancer cells. Mol Med Rep. 9:1153–1158.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang H, Wang X, Xu J and Sun Y: Notch1
activation is a poor prognostic factor in patients with gastric
cancer. Br J Cancer. 110:2283–2290. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu XX, Hu Z, Shen X, Dong LY, Zhou WZ and
Hu WH: IL- 33 p romotes gastric cancer cell invasion and migration
Via ST2 ERK1/2 p athway. Dig Dis Sci. 60:1265–1272. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li X, Liu S, Yan J, Peng L, Chen M, Yang J
and Zhang G: The characteristics, prognosis, and risk factors of
lymph node metastasis in early gastric cancer. Gastroenterol Res
Pract. 2018:69457432018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Song Q, Ji Q and Li Q: The role and
mechanism of β-arrestins in cancer invasion and metastasis
(Review). Int J Mol Med. 41:631–639. 2018.
|
|
25
|
Tian D, Li Y, Li X and Tian Z: Aloperine
inhibits proliferation, migration and invasion and induces
apoptosis by blocking the Ras signaling pathway in human breast
cancer cells. Mol Med Rep. 18:3699–3710. 2018.PubMed/NCBI
|
|
26
|
Osborne CC, Perry KJ, Shankland M and
Henry JQ: Ectomesoderm and EMT-related genes in spiralian
development. Dev Dyn. 247:2018.Doi: 1002/dvdy.24667. View Article : Google Scholar
|
|
27
|
Nantajit D, Lin D and Li JJ: The network
of epithelial-mesen-chymal transition: Potential new targets for
tumor resistance. J Cancer Res Clin Oncol. 141:1697–1713. 2015.
View Article : Google Scholar
|
|
28
|
Yilmaz M and Christofori G: EMT, the
cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev.
28:15–33. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen Z, He J, Xing X, Li P, Zhang W, Tong
Z, Jing X, Li L, Liu D, Wu Q and Ju H: Mn12Ac inhibits the
migration, invasion and epithelial-mesenchymal transition of lung
cancer cells by downregulating the Wnt/β-catenin and PI3K/AKT
signaling pathways. Oncol Lett. 16:3943–3948. 2018.PubMed/NCBI
|
|
30
|
Zhao L, Pang A and Li Y: Function of GCN5
in the TGF-β1-induced epithelial-to-mesenchymal transition in
breast cancer. Oncol Lett. 16:3955–3963. 2018.PubMed/NCBI
|
|
31
|
Linna MA, Chen Z, Zhi-Heng R, Xiao Z and
Xu Z: Expression of LPA2 regulates the migration, invasion,
proliferation, and apoptosis of gastric cancer SGC-7901 cells. Sci
Technol Eng. 18:26–32. 2018.
|
|
32
|
Liu X, Sun K, Song A, Zhang X, Zhang X and
He X: Curcumin inhibits proliferation of gastric cancer cells by
impairing ATP-sensitive potassium channel opening. World J Surg
Oncol. 12:3892014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhu Z, Wang G, Yang F, Cao W, Mao R, Du X,
Zhang X, Li C, Li D, Zhang K, et al: Foot-and-mouth disease virus
viroporin 2B antagonizes RIG-I-mediated antiviral effects by
inhibition of its protein expression. J Virol. 90:11106–11121.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Arranz-Valsero I, Soriano-Romaní L,
García-Posadas L, López-García A and Diebold Y: IL-6 as a corneal
wound healing mediator in an in vitro scratch assay. Exp Eye Res.
125:183–192. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu H, Xue Q, Cao W, Yang F, Ma L, Liu W,
Zhang K, Liu X, Zhu Z and Zheng H: Foot-and-mouth disease virus
nonstructural protein 2B interacts with cyclophilin A, modulating
virus replication. FASEB J. June 15–2018.Epub ahead of print.
View Article : Google Scholar
|
|
37
|
Yang D, Yang W, Zhang Q, Hu Y, Bao L and
Damirin A: Migration of gastric cancer cells in response to
lysophosphatidic acid is mediated by LPA receptor 2. Oncol Lett.
5:1048–1052. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
De Craene B and Berx G: Regulatory
networks defining EMT during cancer initiation and progression. Nat
Rev Cancer. 13:97–110. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Clay MR and Halloran MC: Cadherin 6
promotes neural crest cell detachment via F-actin regulation and
influences active Rho distribution during epithelial-to-mesenchymal
transition. Development. 141:2506–2515. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Takahashi K, Fukushima K, Fukushima N,
Honoki K and Tsujiuchi T: Enhanced cellular functions through
induction of LPA2 by cisplatin in fibrosarcoma HT1080
cells. Mol Cell Biochem. 431:29–35. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Takahashi K, Fukushima K, Tanaka K, Minami
K, Ishimoto K, Otagaki S, Fukushima N, Honoki K and Tsujiuchi T:
Involvement of LPA signaling via LPA receptor-2 in the promotion of
malignant properties in osteosarcoma cells. Exp Cell Res.
369:316–324. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang H, Liu W, Wei D, Hu K, Wu X and Yao
Y: Effect of the LPA-mediated CXCL12-CXCR4 axis in the tumor
proliferation, migration and invasion of ovarian cancer cell lines.
Oncol Lett. 7:1581–1585. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Erstad DJ, Tager AM, Hoshida Y and Fuchs
BC: The autotaxinlysophosphatidic acid pathway emerges as a
therapeutic target to prevent liver cancer. Mol Cell Oncol.
4:e13118272017. View Article : Google Scholar
|
|
44
|
Aasrum M, Tjomsland V, Thoresen GH, De
Angelis PM, Christoffersen T and Brusevold IJ: PI3K is required for
both basal and LPA-induced DNA synthesis in oral carcinoma cells. J
Oral Pathol Med. 45:425–432. 2016. View Article : Google Scholar
|
|
45
|
Zeng R, Li B, Huang J, Zhong M, Li L, Duan
C, Zeng S, Huang J, Liu W, Lu J, et al: Lysophosphatidic acid is a
biomarker for peritoneal carcinomatosis of gastric cancer and
correlates with poor prognosis. Genet Test Mol Biomarkers.
21:641–648. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Singla A, Kumar A, Priyamvada S, Tahniyath
M, Saksena S, Gill RK, Alrefai WA and Dudeja PK: LPA stimulates
intestinal DRA gene transcription via LPA2 receptor, PI3K/AKT, and
c-Fos-dependent pathway. Am J Physiol Gastrointest Liver Physiol.
302:G618–G627. 2012. View Article : Google Scholar :
|
|
47
|
Brooks YS, Ostano P, Jo SH, Dai J, Getsios
S, Dziunycz P, Hofbauer GF, Cerveny K, Chiorino G, Lefort K and
Dotto GP: Multifactorial ERβ and NOTCH1 control of squamous
differentiation and cancer. J Clin Invest. 124:2260–2276. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kang L, Mao J, Tao Y, Song B, Ma W, Lu Y,
Zhao L, Li J, Yang B and Li L: MicroRNA-34a suppresses the breast
cancer stem cell-like characteristics by downregulating Notch 1
pathway. Cancer Sci. 106:700–708. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zang MD, Hu L, Fan ZY, Wang HX, Zhu ZL,
Cao S, Wu XY, Li JF, Su LP, Li C, et al: Luteolin suppresses
gastric cancer progression by reversing epithelial-mesenchymal
transition via suppression of the Notch signaling pathway. J Transl
Med. 15:522017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liu T, Nie F, Yang X, Wang X, Yuan Y, Lv
Z, Zhou L, Peng R, Ni D, Gu Y, et al: MicroRNA-590 is an
EMT-suppressive microRNA involved in the TGFβ signaling pathway.
Mol Med Rep. 12:7403–7411. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Shao S and Zhao X, Zhang X, Luo M, Zuo X,
Huang S, Wang Y, Gu S and Zhao X: Notch1 signaling regulates the
epithelial-mesenchymal transition and invasion of breast cancer in
a Slug-dependent manner. Mol Cancer. 14:282015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Hope JM, Wang FQ, Whyte JS, Ariztia EV,
Abdalla W, Long K and Fishman DA: LPA receptor 2 mediates
LPA-induced endometrial cancer invasion. Gynecol Oncol.
112:215–223. 2009. View Article : Google Scholar
|
|
53
|
Shida D, Kitayama J, Yamaguchi H, Hama K,
Aoki J, Arai H, Yamashita H, Mori K, Sako A, Konishi T, et al: Dual
mode regulation of migration by lysophosphatidic acid in human
gastric cancer cells. Exp Cell Res. 301:168–178. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Suzuki HI, Horie M, Mihira H and Saito A:
Molecular analysis of endothelial-mesenchymal transition induced by
transforming growth factor-β signaling. J Vis Exp. 2018. View Article : Google Scholar
|
|
55
|
Yang M, Zhong WW, Srivastava N, Slavin A,
Yang J, Hoey T and An S: G protein-coupled lysophosphatidic acid
receptors stimulate proliferation of colon cancer cells through the
{beta}-catenin pathway. Proc Natl Acad Sci USA. 102:6027–6032.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yu S, Murph MM, Lu Y, Liu S, Hall HS, Liu
J, Stephens C, Fang X and Mills GB: Lysophosphatidic acid receptors
determine tumorigenicity and aggressiveness of ovarian cancer
cells. J Natl Cancer Inst. 100:1630–1642. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Sun Y, Zhang R, Zhou S and Ji Y:
Overexpression of Notch1 is associated with the progression of
cervical cancer. Oncol Lett. 9:2750–2756. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Lin FT and Lai YJ: Regulation of the LPA2
receptor signaling through the carboxyl-terminal tail-mediated
protein-protein interactions. Biochim Biophys Acta. 1781:558–562.
2008. View Article : Google Scholar : PubMed/NCBI
|