Introduction
Acute lung injury (ALI) is a critical complication
of some clinical illnesses with high rates of morbidity and
mortality, and it can lead to acute respiratory distress syndrome
(1). ALI is characterized by an
excessive inflammatory response, excessive neutrophil infiltration
into the lung tissues, the release of pro-inflammatory cytokines,
and lung endothelial and epithelial injuries, which cause edema and
gas exchange deterioration (2-4).
Therefore, the inhibition of the excessive inflammatory response
may be an effective approach for the prevention and treatment of
ALI.
Numerous studies have reported that volatile
anesthetic can provide protective effects in various tissues and
organs (5-7). Minguet et al (8) demonstrated that volatile anesthetics
could protect against ischemia-reperfusion (IR) injury in the
heart. Sevoflurane, a volatile anesthetic, is most commonly used as
first line anesthesia (9).
Despite being an inhaled agent, only a few studies have been
reported using sevoflurane to attenuate lung injury (10-12). For example, Suter et al
(13) revealed that sevoflurane
can reduce the lung tissue edema and inflammatory cell infiltration
induced by endotoxin. Ohsumi et al (14) demonstrated that sevoflurane
exhibited significant protective effects against lung IR injury
(IRI) via its anti-inflammatory activities in a rat lung
transplantation model. In addition, sevoflurane was found to
improve LPS-induced ALI by inhibiting lung inflammation in a series
of in vivo and in vitro experiments (15,16). However, the mechanism of such
effects on ALI is also still unclear.
MicroRNAs (miRNAs/miRs) are a class of endogenous
small noncoding RNAs of ~21-23 nucleotides in length that repress
the translation or induce the degradation of target mRNA by binding
to the 3′-untranslated region (UTR) of target mRNAs (17,18). An increasing body of evidence has
supported that miRNAs are implicated in certain inflammatory lung
diseases. For example, Guo et al (19) reported that the upregulation of
miR-125b significantly reduced lipopolysaccharide (LPS)-induced
pulmonary inflammation in mice. Chen et al (20) demonstrated that miR-212-3p
inhibited the LPS-induced inflammatory response by targeting high
mobility group box-1 protein in murine macrophages. Recently,
several studies have revealed that sevoflurane exerts its
biological effects via modulation of miRNAs (21-23). Wu et al (22) revealed that sevoflurane protected
against hepatic IRI by modulating miRNA-200c regulation in mice.
Furthermore Wenlan et al (23) demonstrated that miR-34a-5p
attenuated the protective effect of sevoflurane in
hypoxia/reoxygenation-induced cardiomyocyte injury by targeting
Syntaxin 1A. In another study, Lv et al (21) reported that the downregulation of
miR-27a-3p expression ameliorated sevoflurane-induced neurotoxicity
as well as learning and memory impairment in neonatal mice.
However, whether miRNAs participate in the anti-inflammatory
activity of sevoflurane in ALI remains unclear.
In the present study, the aim was to assess the
possible protective effects of sevoflurane against LPS-induced lung
injury and to elucidate the possible underlying mechanisms. The
results from the in vivo experiments suggest that
sevoflurane attenuates LPS-induced ALI through the inhibition of
miR-27a/Toll-like receptor 4 (TLR4)/MyD88/NF-κB signaling pathway
activation. Therefore, the present study suggests that sevoflurane
may be a potential approach for ALI treatment.
Materials and methods
Animals
Male BALB/c mice (n=84), weighing 20-30 g, aged 8-12
weeks, were purchased from the Shanghai Experimental Animal Center.
All surgical and care procedures were approved by the Animal Care
and Use Committee of the First Affiliated Hospital of Xinxiang
Medical University. All mice were maintained in a
temperature-controlled room (22±2°C) with a 12-h light/dark cycle,
a relative humidity of 40-60% and free access to food and
water.
Experimental design and LPS
induction
The mice were randomized into 7 experimental groups
(n=6/group): Group 1, the Sham group who received saline (0.3 ml,
intragastrically); group 2, the ALI group who only received LPS (5
mg/kg body weight, intranasally; Escherichia coli serotype
055:B5; Sigma-Aldrich; Merck KGaA) in 300 µl PBS for
sensitization (24); group 3, the
sevoflurane group (SEVO group), in which mice inhaled 3%
sevoflurane (Sigma-Aldrich; Merck KGaA) for 4 h; group 4, ALI +
SEVO group (3% sevoflurane + LPS), in which mice were treated the
same as the ALI group but were administered 3% sevoflurane for 4 h,
starting 2 h after LPS treatment (25); group 5, ALI + agomir-27a group (8
mg/kg agomir-27a + LPS), in which mice were injected with
agomir-27a (8 mg/kg) by tail intravenous injection 24 h prior to
LPS treatment, and then repeatedly injected every 24 h for 3 day;
group 6, ALI + SEVO + antagomir-27a group (3% sevoflurane + 8 mg/kg
antagomir-27a + LPS), in which mice were treated in the same way as
group 5 (8 mg/kg antagomir-27a + LPS) but were also administered 3%
sevoflurane for 4 h, starting 2 h after LPS treatment; group 7, ALI
+ SEVO + antagomir-negative control (NC) group (3% sevoflurane + 8
mg/kg antagomir-NC + LPS), in which mice were treated in the same
way as group 5 (8 mg/kg antagomir-NC + LPS) but were also
administered 3% sevoflurane for 4 h, starting at 2 h after LPS
treatment. Animals were sacrificed 3 days after treatment with LPS
or saline, and the lung tissues at 6, 12 and 24 h, and
bronchoalveolar lavage fluid (BALF) at 12 h were collected for
analysis. Antagomir-27a and agomir-27a were designed and
synthesized by Guangzhou RiboBio Co., Ltd. The doses of miRNA were
selected based on previous studies (26-29). The lung tissues were snap-frozen
and stored at -80°C until subsequent use.
To evaluate the transfection efficiency of miR-27a
in vivo, 12 mice were randomized into another 4 experimental
groups (n=3/group). Agomir-27a/agomir-NC groups
(Antagomir-27a/antagomir-NC groups), in which all mice were given
agomir-27a/agomir-NC (Antagomir-27a/antagomir-NC groups; 8 mg/kg)
via tail intravenous administration every 24 h for 3 days; after a
further 3 days, animals were sacrificed and the lung tissues were
extracted.
The survival experiment
The survival experiments were performed in all 7
groups of mice (the Sham, ALI, SEVO, ALI + SEVO, ALI + agomir-27a,
ALI + SEVO + antagomir-27a and ALI + SEVO + antagomir-NC groups;
n=10/each group). Mice were treated as aforementioned. Mice were
monitored regularly, and survival was recorded over a period of 7
days.
Bronchoalveolar lavage collection
BALF was obtained by flushing the right lung lobes
with 1.0 ml PBS. In total, ~0.8 ml of BALF was recovered without
significant differences between groups. After centrifugation for 10
min at 600 g at 4°C, the pellet was re-suspended in PBS. Total cell
numbers were counted using a hemocytometer. At least 200 cells were
counted per sample. The supernatants were collected and immediately
frozen on dry ice and stored at -80°C for cytokine measurements
using mouse ELISA kits (R&D Systems, Inc.) for interleukin
(IL)-1β, IL-6, and tumor necrosis factor (TNF)-α.
Evaluation of lung permeability
Lung permeability was assessed using the Evans blue
dye extravasation method, as described previously (30). Briefly, via the tail vein, 4 ml/kg
of 2% Evans blue (Sigma-Aldrich; Merck KGaA) in normal saline was
administered 10 h before the animals were euthanized. After
adequate perfusion with normal saline, Evans blue dye was extracted
from the lung using formamide for 18 h at 60°C and measured as the
absorbance of the supernatant at 620 nm on a microplate reader
(BioTek Instruments, Inc.) and was reported as the amount of Evans
blue per wet tissue weight (µg/g).
Lung wet/dry (W/D) ratio
Mice were sacrificed, and the right lungs were
excised and placed in an incubator at 80°C for 48 h to obtain the
dry weight. The ratio of wet lung to dry lung was then calculated
to assess tissue edema.
Histopathologic evaluation of lung
tissues
Upper and lower lobe lung samples were excised 24 h
after the LPS challenge, fixed with 10% formalin at room
temperature for 24 h and microsectioned at 5 µm. Then, the
tissues were embedded in paraffin and stained with hematoxylin and
eosin at 37°C for 5 min. Finally, the lung injury score was
assessed in a blinded fashion via semi-quantitative light
microscopy evaluation as previously described (31). The images were captured with a
Nikon E100 light microscope with ×200 magnification.
Measurement of pro-inflammatory
cytokines
The levels of TNF-α (cat. no. MTA00B), IL-6 (cat.
no. M6000B), and IL-1β (cat. no. MLB00C) in BALF were measured
using mouse ELISA kits (R&D Systems, Inc.) according to the
manufacturer's instructions protocol.
miRNA microarray
Total RNA was isolated from frozen lung tissue using
the miRNeasy isolation kit (Qiagen, Inc.) following the
manufacturer's protocol. The samples were analyzed via a miRCURY
LNA™ Array (v.18.0; Agilent Technologies, Inc.). The procedure and
images process method were conducted as described previously
(32).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from the left lung tissue
samples using a miRNeasy mini kit (Qiagen, Inc.) following the
manufacturer's protocol. RT was performed at 40°C for 45 min
followed by incubation at 95°C for 5 min using a miRNA reverse
transcription kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.). qPCR was performed using a standard protocol from the SYBR
Green PCR kit (Toyobo Life Science) on an ABI PRISM 7500 Real-time
PCR system (Thermo Fisher Scientific, Inc.). RT-qPCR was performed
at 95°C for 10 min, followed by 40 cycles of amplification at 95°C
for 2 sec and 60°C for 34 sec. The melting curve stage was
performed at 95°C for 30 sec, 60°C for 30 min and 95°C for 30 sec.
The primer sequences were as follows: miR-19a forward, 5′-TCA TCA
CGC TGT GCA AAT CT-3′ and reverse, 5′-TAT GGT TGT TCT GCT CTC TGT
CTC-3′; miR-149* forward, 5′-ACA CTC CAG CTG GGA GGG ACG GGG GC-3′
and reverse, 5′-CTC AAC TGG TGT CGT GGA-3′; miR-146a-5p forward,
5′-TGA GAA CTG AAT TCC ATG GGT-3′ and reverse, 5′-CGA GAA GCT TGC
ATC ACC AGA GAA CG-3′; miR-122 forward 5′-GTG ACA ATG GTG GAA TGT
GG-3′ and reverse, 5′-AAA GCA AAC GAT GCC AAG AC-3′; miR-27a
forward 5′-TGC GGT TCA CAG TGG CTA AG-3′ and reverse, 5′-CTC AAC
TGG TGT CGT GGA-3′; U6 forward, 5′-GCT TCG GCA GCA CAT ATA CTA AAA
T-3′ and reverse, 5′CGC TTC ACG AAT TTG CGT GTC AT-3′. The results
were quantified using the 2−ΔΔCq method (33).
Cells transfection
In order to up-or downregulate the expression of
miR-27a, 50 nM miR-27a mimics, 50 nM miR-27a inhibitor and 50 nM
their NC were synthesized by Shanghai GenePharma Co., Ltd. The
sequences for these oligonucleotides were as follows: 5′-UUC ACA
GUG GCU AAG UUC CGC-3′ for miR-27a mimics; 5′-GCG GAA CUU AGC CAC
UGU GAA-3′ for miR-27a inhibitor; 5′-UUC UCC GAA CGU GUC ACG UTT-3′
for NC mimics; and 5′-CAG UAC UUUUGU GUA GUA CAA-3′ for the NC
inhibitor. Cell transfection was conducted using Lipofectamine
2000™ reagent (Invitrogen; Thermo Fisher Scientific, Inc.) for 48 h
following the manufacturer's protocol. At 24 h post-transfection,
the cells were collected and used for the following analyses.
Bioinformatics
TargetScan (version 7.0; www.targetscan.org/), miRbase (version 21.0;
www.mirbase.org/) and PicTar (version 2006;
pictar.mdc-berlin.de) target gene prediction software were used to
select MIF as a target gene of miR-27a.
Luciferase reporter assay
The 3′-UTR of TLR4 and the mutated sequence were
inserted into the pGL3 control vector (Promega Corporation) to
construct the wild-type (wt) TLR4-3′-UTR vector and mutant
TLR4-3′-UTR vector, respectively. For luciferase reporter assay,
293T cells (American Type Culture Collection) in 24-well plates
(5.0×105/well) and were transfected with either the wt
or mut reporter vector, combined with miR-27a mimics or miR-27a
inhibitor using Lipofectamine 2000™ (Invitrogen; Thermo Fisher
Scientific, Inc.). At 48 h post-transfection, the dual-luciferase
reporter assay system (Promega Corporation) was used to measure the
luciferase activity. The pRL-TK plasmid (Promega Corporation) was
used as a normalizing control. All experiments were performed in
triplicate.
Western blot analysis
Lung tissue samples were lysed in RIPA buffer (Cell
Signaling Technology, Inc.) supplemented with protease inhibitors
(Roche Applied Science) for 30 min on ice. The protein
concentrations were measured using a BCA protein assay kit
(Beyotime Institute of Biotechnology). The protein samples (40
µg) were separated by 10% SDS-PAGE (Bio-Rad Laboratories,
Inc.) and transferred onto a PVDF membrane, which was then blocked
with 5% non-fat milk at room temperature for 1 h. Membranes were
incubated with primary antibodies against Bcl-2 (cat. no. ab32124;
1:2,000), cleaved caspase-3 (cat. no. ab2302; 1:2,000), TLR4 (cat.
no. ab22048; 1:2,000), nuclear-phospho (p)-p65 (Ser-536; cat. no.
ab86299; 1:2,000), total p-65 (cat. no. ab140751; 1:2,000), p-IκBα
(Ser-36; cat. no. ab133462; 1:5,000), total IκBα (cat. no. ab32518;
1:5,000), MyD88 (cat. no. ab199247; 1:2,000) and β-actin (cat. no.
ab8226; 1:2,000; all Abcam) at 4°C overnight. After washing with
PBS, the membrane was incubated with the appropriate horseradish
peroxidase-conjugated secondary antibodies including goat
anti-rabbit immunoglobulin (Ig)-G (cat. no. ab6721; 1:5,000; Abcam)
and goat anti-mouse IgG (cat. no. ab6789; 1:5,000; Abcam) for 1 h
at room temperature. A chemiluminescence detection system (EMD
Millipore) was used for visualization of the results and
quantification of the bands was performed using Quantity One
software (version 4.4; Bio-Rad Laboratories, Inc.).
Statistical analysis
GraphPad Prism 5.0 (GraphPad Software, Inc.) was
used to analyze the statistical analyses. Data are presented as the
mean ± standard deviation. Statistical differences were analyzed
using one-way analysis of variance with the Tukey's post hoc test.
Survival studies were analyzed using the log rank test and the
results are presented as the Kaplan-Meier curves. P<0.05 were
considered to indicate a statistically significant difference.
Results
Sevoflurane protects against LPS-induced
ALI in mice
Previous studies on rat and mice have revealed that
sevoflurane is able to improve different types of injuries,
including lung, liver and renal injuries (10,34-36). In the present study, a mouse model
of LPS-induced ALI was used to evaluate the therapeutic effects of
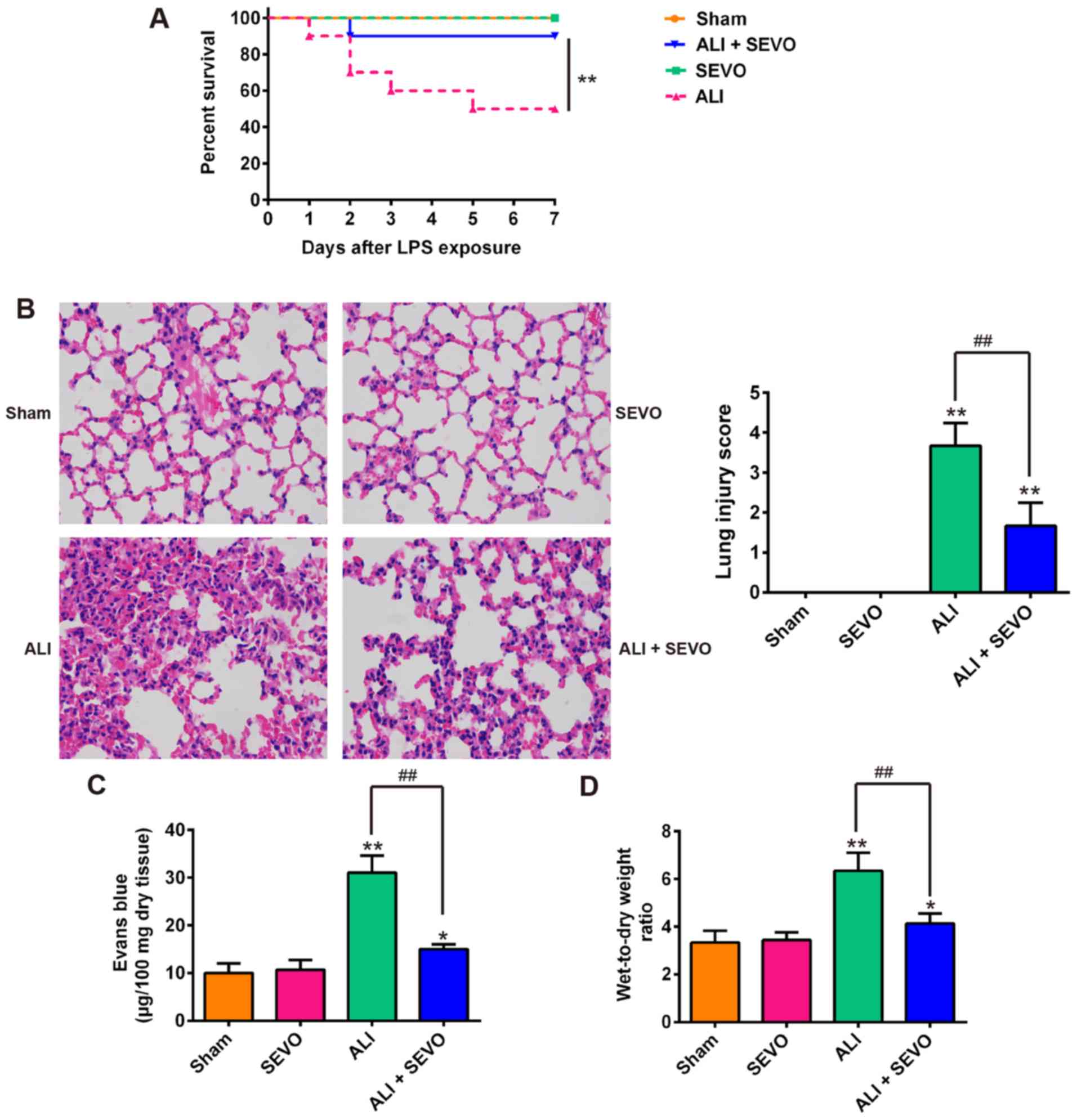
sevoflurane. As shown in Fig. 1A,
50% of the LPS-induced ALI mice succumbed within 7 days whilst the
survival rate of the ALI + sevoflurane mice was significantly
higher than that of the ALI group. Then, the present study analyzed
the histopathological changes in the lungs of ALI mice. Lung
inflammatory responses including interstitial infiltration of
inflammatory cells and thickening of the alveolar walls observed in
the lung tissues of LPS-induced ALI mice were markedly reduced
following sevoflurane treatment (Fig.
1B-D). Subsequently, the histopathological changes were scored
and the result revealed that sevoflurane clearly ameliorated the
histological damage (Fig. 1B). In
addition, Evans blue dye was employed to evaluate the permeability
of lung vasculature and it was observed that the LPS-induced
increase in capillary permeability was significantly abolished by
sevoflurane (Fig. 1C).
Furthermore, the lung W/D ratio (an indicator of the extent of lung
edema) of mice in the ALI group was higher than that of Sham mice,
whilst sevoflurane significantly reduced the W/D ratio of
LPS-stimulated ALI mice compared with the ALI group (Fig. 1D). These results indicated that
the mouse model of ALI was successfully established and that
sevoflurane treatment can protect LPS induced ALI in mice.
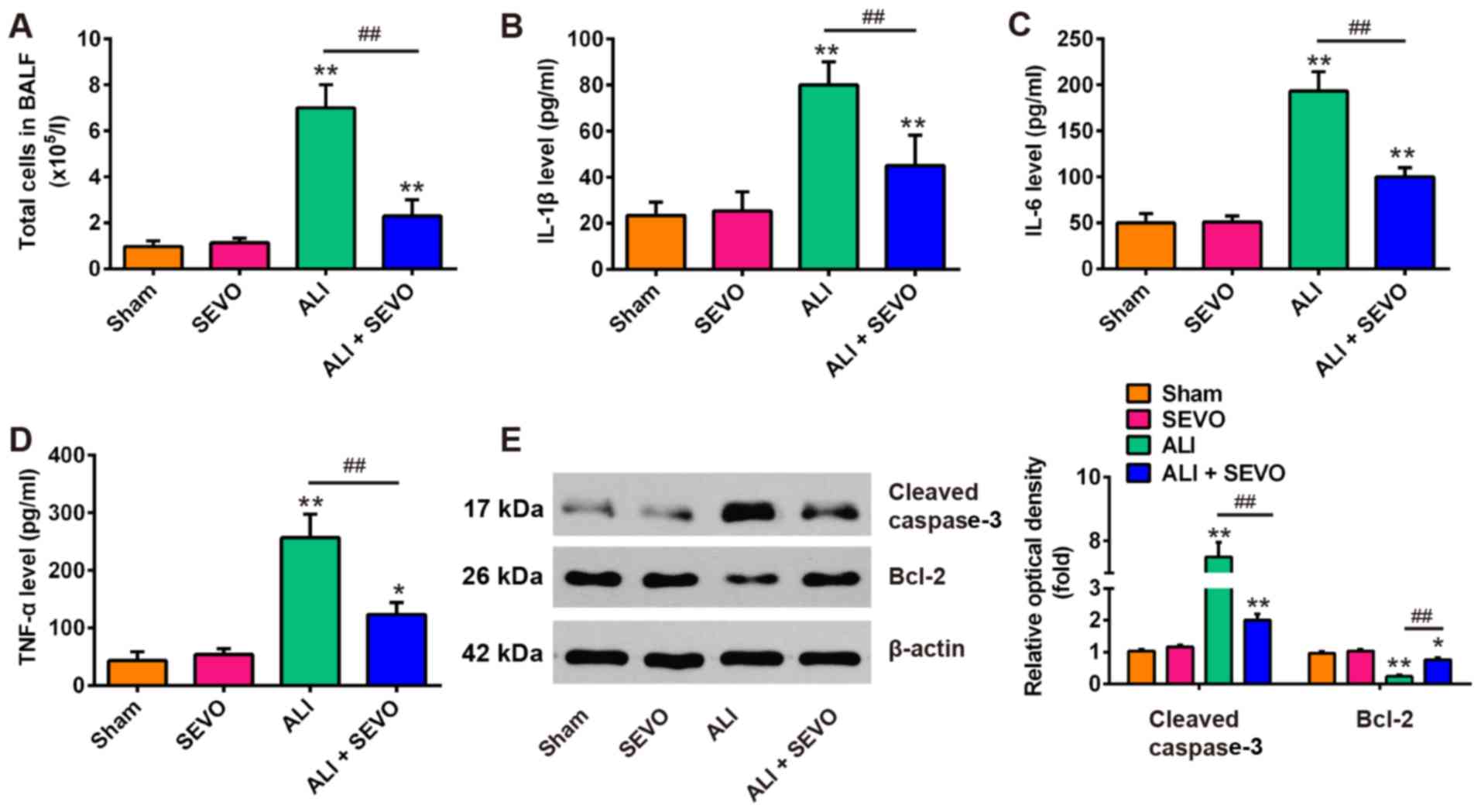
Sevoflurane inhibits LPS induced
inflammation and apoptosis in mice
An excessive inflammatory response and increased
endothelial cell permeability contribute to vascular leakage in
ALI, which is considered to be central to the pathogenesis of ALI
(37). Thus, the present study
then examined the protective effects of sevoflurane on vascular
leakage in ALI mice by measuring the total cell counts in BALF
using a hemocytometer. As shown in Fig. 2A, LPS caused a significant
increase in the total cell counts in BALF compared with the Sham
group, whereas sevoflurane lead to a reduction in the LPS-induced
total cell counts in the LPS and sevoflurane treated group. To
further investigate the anti-inflammatory effects of sevoflurane,
the present study measured the expression levels of inflammatory
factors (IL-1β, IL-6 and TNF-α) in BALF via ELISA. The results
demonstrated that the concentrations of these pro-inflammatory
factors in the ALI group were significantly increased compared with
the Sham group, whereas sevoflurane pretreatment significantly
ameliorated the levels of these inflammatory cytokines (Fig. 2B-D). Additionally, the levels of
the pro-apoptotic marker cleaved caspase-3 and anti-apoptotic
marker Bcl-2 were also analyzed by western blotting in BALF. As
presented in Fig. 2E, LPS
administration increased the levels of the pro-apoptotic protein,
cleaved caspase 3 and decreased the level of the anti-apoptotic
protein Bcl2 compared with the Sham group. On the other hand,
sevoflurane pretreatment induced a significant decrease in the
levels of cleaved caspase-3 as well as a marked increase in the
level of Bcl2 when compared with the ALI group. These results
suggest that sevoflurane attenuates LPS-induced lung injury by
suppressing the inflammatory response and reducing apoptosis.
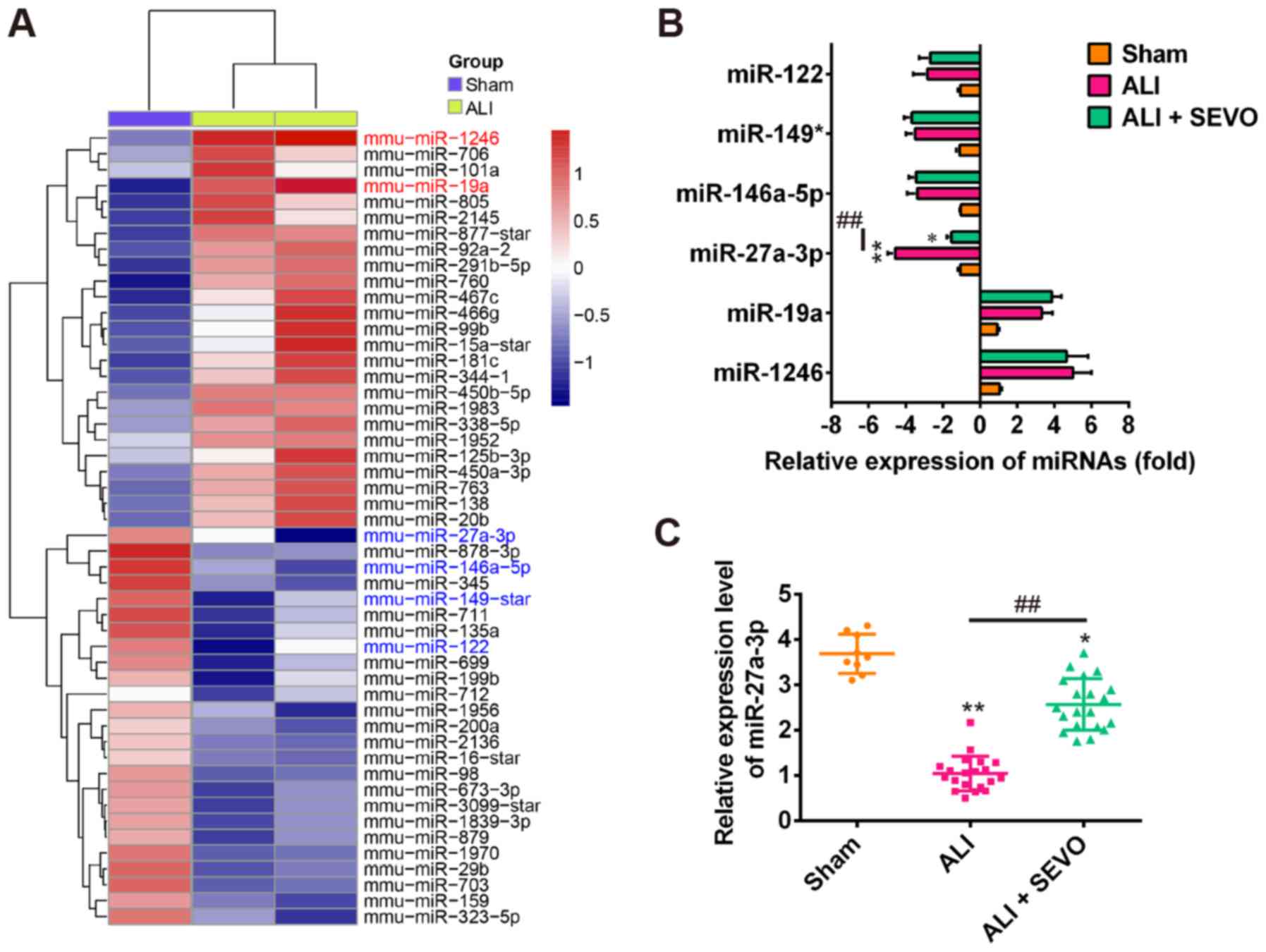
Sevoflurane upregulates the expression of
miR-27a
Previous studies have reported that various
aberrantly expressed miRNAs are associated with ALI and
inflammatory responses (38,39). To determine the potential
involvement of miRNAs in ALI in mice, the present study used
microarray analysis to determine the miRNA levels in lung tissues
between ALI and Sham groups. The miRNA microarray identified 25
miRNAs that were upregulated and 25 miRNAs that were downregulated
in the ALI group, compared with the Sham group (Fig. 3A). To determine whether
sevoflurane could alter the expressions of some miRNAs obtained
from the miRNA microarray data, a total of 6 miRNAs were selected
including miR-1246 (40),
miR-149* (41), miR-122 (42), miR-19a (43), miR-146a-5p (44) and miR-27a (45) as the candidate miRNAs as they have
been shown to play a critical role in inflammation regulation. As
shown in Fig. 3B sevoflurane only
upregulated the expression of miR-27a, while the other miRNAs
exhibited no evident changes in expression after sevoflurane
treatment. To further investigate the effect of sevoflurane on the
expression of miR-27a in ALI mice, RT-qPCR was performed in a
larger scale sample. As shown in Fig.
3C, miR-27a expression was significantly decreased after LPS
treatment (ALI group); however, in the sevoflurane pretreatment
group (ALI + SEVO) the level of miR-27a increased significantly
when compared with the ALI group. As miR-27a has previously been
reported to be upregulated by sevoflurane in a sevoflurane-induced
anesthetic mouse model (21), it
seems plausible that the observed protective effects of sevoflurane
may be exerted through upregulating the expression of miR-27a in
ALI.
miR-27a is involved in the therapeutic
effects of sevoflurane in ALI mice
Previous studies have shown that miR-27a play
important roles in several types of injuries, including traumatic
brain injury (46),
oxygen-glucose deprivation-induced injury (46) and LPS-induced sepsis injury
(47). These studies warranted
the investigation into whether the ectopic expression of miR-27a
contributes to the therapeutic effects of sevoflurane in ALI mice.
Since the stability of antagomir/agomir is stronger than
inhibitor/mimics, the agomir/antagomir were used in the in
vivo experiments. The present study overex-pressed and knocked
down the expression of miR-27a via an injection of
agomir-27a/antagomir-27a prior to LPS and sevoflurane treatment,
and then observed the alterations in the therapeutic effects of
sevoflurane in ALI mice. Firstly, it was observed that the
expression of miR-27a was significantly increased or decreased
following the injection of agomir-27a or antagomir-27a,
respectively (Fig. 4A). As shown
in Fig. 4B, mouse survival in the
ALI + agomir-27a group was markedly improved compared with ALI
group, which was similar to the effects of sevoflurane pretreatment
observed. However, the survival rate of ALI + SEVO + antagomir-27a
mice was significantly lower than that of the ALI + SEVO group.
Furthermore, the agomir-27a injection improved LPS-induced lung
injury, as evidenced by the reduced histological scores, capillary
permeability and W/D ratio of lung in the lung tissues (Fig. 4C-E), which is also similar to the
observed effects of sevoflurane. However, the improvement induced
by sevoflurane in lung injury was abrogated when miR-27a was
knocked down. In addition, the agomir-27a injection attenuated the
LPS-induced inflammatory response including decreasing the total
cell counts and reducing the levels of IL-1β, IL-6 and TNF-α in
BALF (Fig. 4F-I); treatment with
sevoflurane produced similar results. However, the inhibitory
effects of sevoflurane on inflammatory response were abolished
after the antagomir-27a injection. These results indicate that
sevoflurane ameliorates LPS-induced ALI in mice by upregulating
miR-27a.
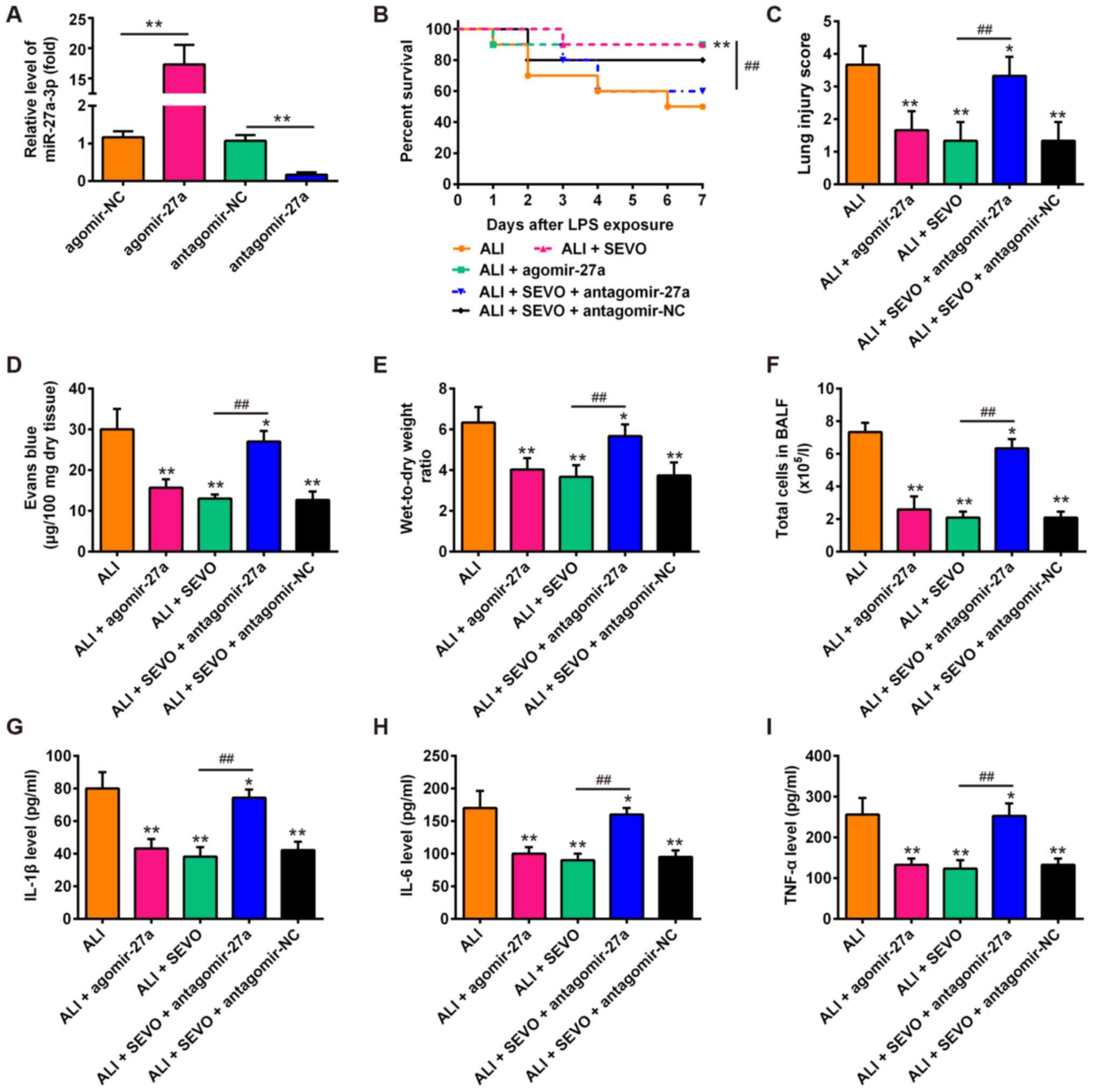 | Figure 4miR-27a is involved in the
therapeutic effects of sevoflurane in ALI mice. (A) The expression
of miR-27a was measured by reverse transcription-quantitative PCR
after agomir-27a and antagomir-27a (8 mg/kg) treatment via tail
intravenous injection in mice (n=3 mice/group).
**P<0.01, as indicated. Then, mice were divided into
5 groups: The Sham, ALI group, ALI + SEVO, ALI + agomir-27a and ALI
+ SEVO + antagomir-27a groups (n=6 mice/group). (B) Animal survival
was recorded at the indicated time points. (C) Lung slides were
stained with hematoxylin and eosin to review lung damage and the
histopathological changes were scored. (D) The capillary
permeability was evaluated by Evans Blue assay. (E) The ratio of
wet lung to dry lung was calculated to evaluate lung tissue edema.
(F) The total counts of cells from BALF were counted using a
hemocytometer. (G-I) The protein levels of (G) IL-1β, (H) IL-6 and
(I) TNF-α were assessed by ELISA. All data are presented as the
mean ± standard deviation of three independent experiments.
*P<0.05 and **P<0.01 vs. Sham group;
##P<0.01, as indicated. SEVO, the sevoflurane treated
group; ALI, acute lung injury; miR, microRNA; IL, interleukin; TNF,
tumor necrosis factor; BALF, bronchoalveolar lavage fluid; NC,
negative control. |
TLR4 is a direct target of miR-27a
To explore the molecular mechanism by which miR-27a
functions in the therapeutic effects of sevoflurane in ALI mice,
the potential targets of miR-27a were screened using the
TargetScan, miRBase and PicTar databases. According to the
bioinformatics analysis, the present study focused on TLR4, an
upstream positive regulator of the NF-κB signaling pathway. As
shown in Fig. 5A, the RNA
sequence alignment revealed that the 3′-UTR of TLR4 mRNA contained
a complementary site for the seed region of miR-27a. To confirm the
target reaction of miR-27a on the 3′-UTR of TLR4 mRNA, a luciferase
activity assay was performed. The reporters were co-transfected
with either miR-27a mimics/inhibitor or with NC mimics/inhibitor
into 239T cells, and luciferase activity was then measured. It was
observed that the miR-27a expression levels were significantly
reduced in cells transfected with the miR-27a inhibitor, and were
significantly increased in cells transfected with the miR-27a
mimics (Fig. 5B). Then, it was
revealed that the overexpression of miR-27a decreased the relative
luciferase activity in the presence of the wt 3′-UTR, whereas
knockdown of miR-27a increased the relative luciferase activity
(Fig. 5C). Similarly, the
luciferase activity did not change significantly when the targeted
sequence of TLR4 was mutated in the miR-27a-binding site. To
further confirm that TLR4 was negatively regulated by miR-27a, the
TLR4 protein expression levels were analyzed by western blot
analysis. The results demonstrated that the TLR4 levels were
markedly downregulated after miR-27a mimics transfection, while
transfection with the miR-27a inhibitor enhanced TLR4 expression
(Fig. 5D). In addition, the
present study also measured the expression of TLR4 in ALI mice
treated with sevoflurane and/or antagomir-27a. When sevoflurane was
applied to LPS treated mice, it was observed that the expression of
TLR4 protein decreased significantly compared with the ALI group.
However, antagomir-27a injection reversed the inhibitory effect of
sevoflurane on the protein levels of TLR4 (Fig. 5E). Taken together, these results
indicate that TLR4 is a functional target of miR-27a, suggesting
that the miR-27a/TLR4 signaling axis may be involved in the
therapeutic effects of sevoflurane in ALI mice.
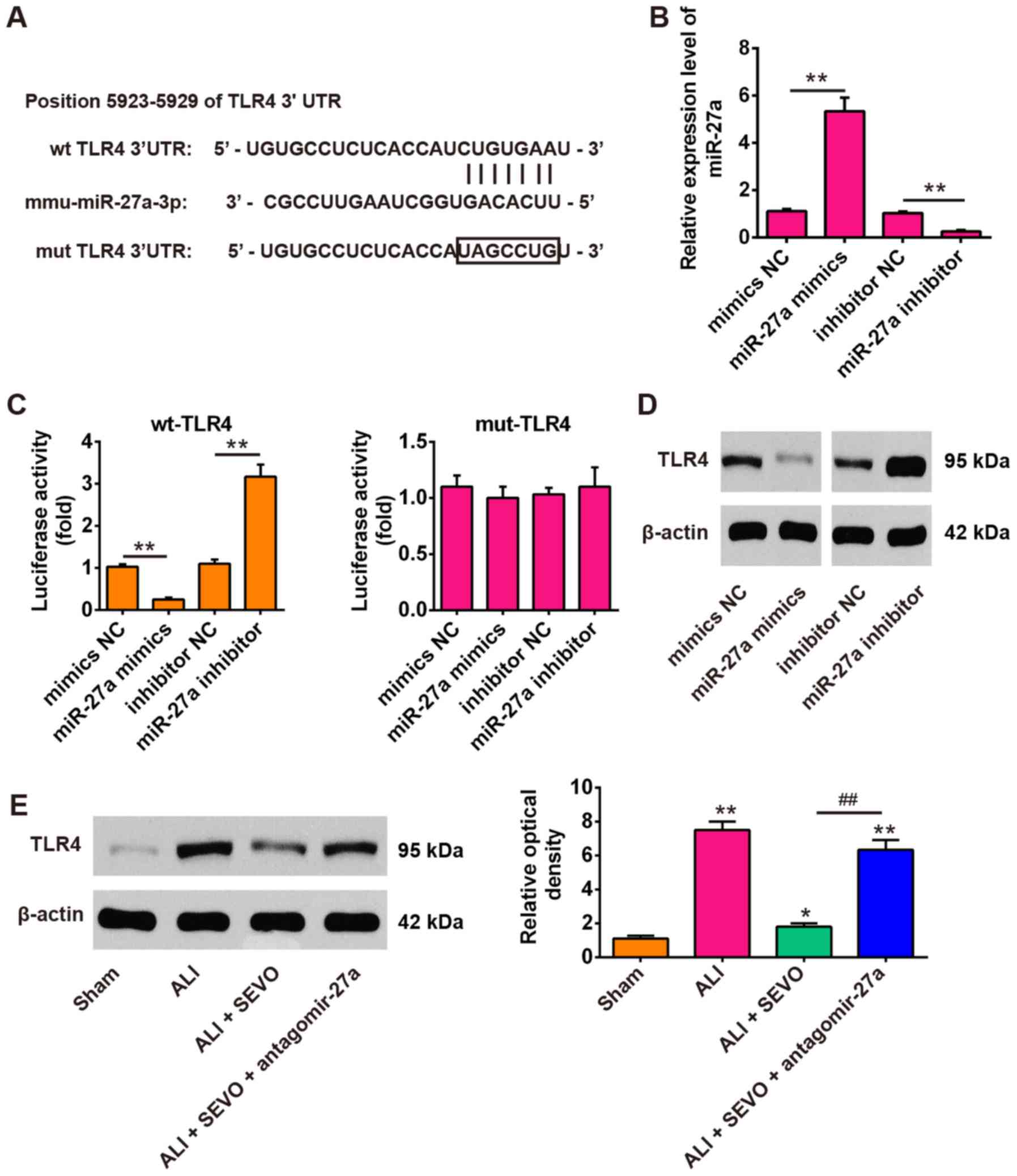 | Figure 5TLR4 was a direct target of miR-27a.
(A) miR-27a-binding sequences in the 3′-UTR of TLR4 and mutated
sites in the 3′-UTR of TLR4. (B) The transfection efficiency of
miR-27a mimics/inhibitor was determined by reverse
transcription-quantitative PCR. (C) miR-27a mimics suppressed the
lucif-erase activities of constructs containing the 3′-UTR segment
of TLR4, while miR-27a inhibitor significantly increased the
luciferase activities of constructs containing the 3′-UTR segment
of TLR4. All data are expressed as the mean ± standard deviation
(n=3). **P<0.01, as indicated. (D) The expression of
TLR4 was detected by western blotting after treatment with miR-27a
mimics or miR-27a inhibitor. (E) The expression of TLR4 was
detected by western blotting in 4 groups of mice: The Sham, ALI,
ALI + SEVO and ALI + SEVO + antagomir-27a groups (n=6 mice/group).
All data are presented as the mean ± standard deviation (n=6).
*P<0.05 and **P<0.01 vs. Sham group;
##P<0.01, as indicated. UTR, untranslated region;
TLF4, Toll-like receptor 4; SEVO, the sevoflurane treated group;
ALI, acute lung injury; miR, microRNA; NC, negative control; wt,
wild-type; mut, mutant. |
The role of the NF-κB signaling pathway
in the protective effects of sevoflurane
Previous studies have indicated that the activation
of the NF-κB signaling pathway plays a pivotal role in the
inflammatory response to ALI, whereby downregulation of NF-κB
expression reduces the release of pro-inflammatory cytokines and
attenuates ALI (48,49). Given that miR-27a directly targets
the TLR4, a well-known upstream positive regulator of the NF-κB
signaling pathway (50), the
present study determined whether the miR-27a/TLR4/NF-κB axis was
involved in the therapeutic effects of sevoflurane in ALI mice. The
protein expressions of the NF-κB signaling pathway transcription
factors including MyD88, nuclear p-p65, total p-65, p-IκB-α and
total IκB-α in the lung tissues obtained from the different groups
were determined by western blotting. The results revealed that the
levels of MyD88, nuclear p-p65 and p-IκB-α were increased in ALI
mice when compared with the Sham group, but these promotional
effects were inhibited by sevoflurane (Fig. 6). However, inhibition of miR-27a
by the antagomir-27a reversed the inhibitory effects of sevoflurane
on the expressions of MyD88, nuclear p-p65 and p-IκB-α (Fig. 6A and B). These results suggest
that sevoflurane blocked the activation of the NF-κB signaling
pathway by upregulating the miR-27a/TLR4 axis.
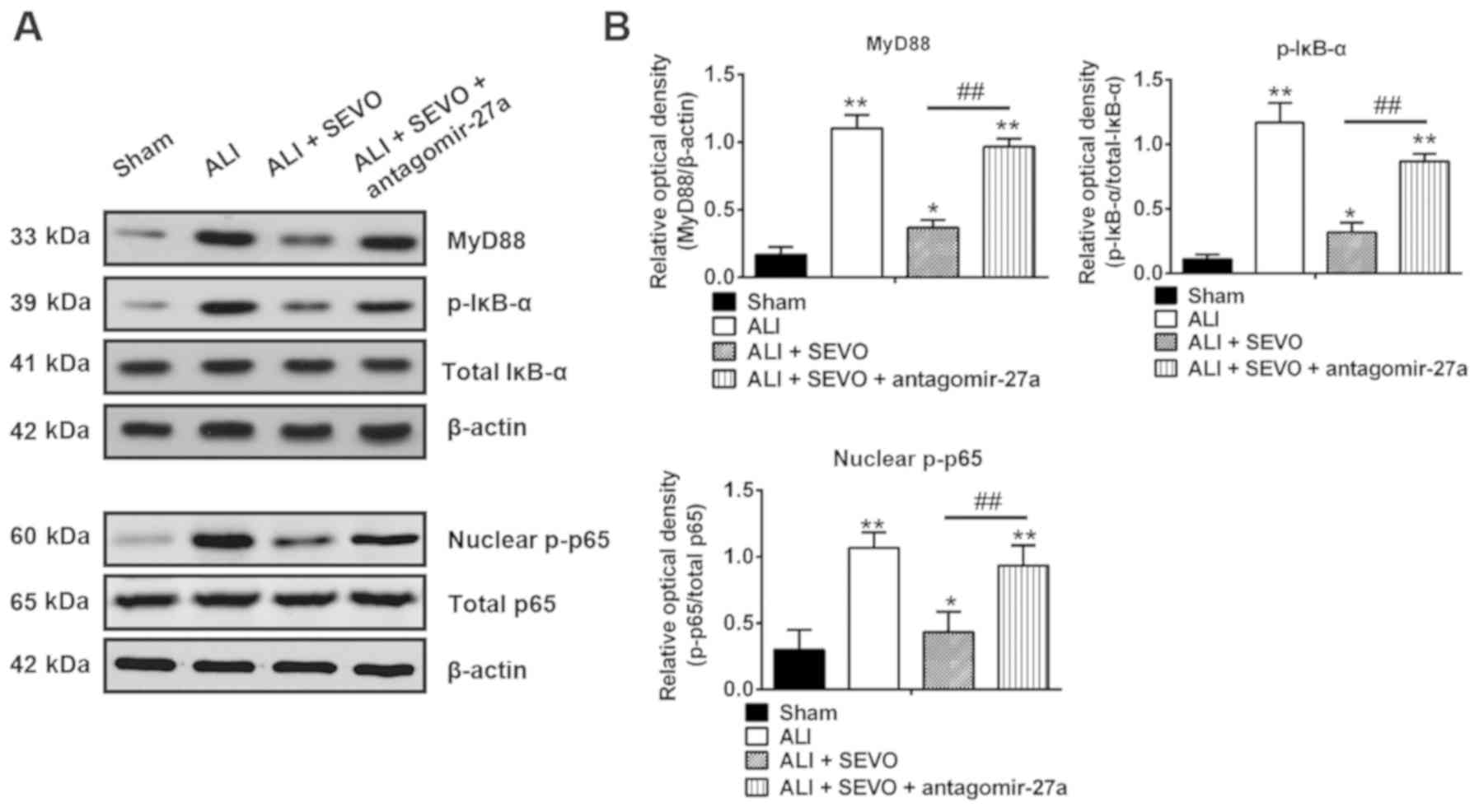 | Figure 6Sevoflurane blocks the activation of
the TLR4/MyD88/NF-κB signaling pathway. (A) The protein expressions
of the NF-κB signaling pathway transcription factors including
MyD88, nuclear p-p65, total p-65, total IκB-α and p-IκB-α were
detected by western blotting in four groups of mice: The Sham, ALI,
ALI + SEVO and ALI + SEVO + antagomir-27a-3p groups (n=6
mice/group). (B) The bands were semi-quantitatively analyzed using
Quantity One software, and normalized to β-actin, total p65 and
total IκB-α density, respectively. Data are presented as the mean ±
standard deviation of three independent experiments.
*P<0.05 and **P<0.01 vs. Sham group;
##P<0.01, as indicated. TLF4, Toll-like receptor 4;
SEVO, the sevoflurane treated group; ALI, acute lung injury; p-,
phosphorylated. |
Discussion
In the present study, it was demonstrated that
sevoflurane effectively protected against LPS-induced ALI by
inhibiting the inflammatory response in mice. Furthermore,
sevoflurane increased the expression of miR-27a in the lung tissues
of the LPS-induced ALI mice model. In addition, the present results
demonstrated that the overexpression of miR-27a ameliorated
LPS-induced ALI and inhibited the expression of the
pro-inflammatory cytokines, TNF-α, IL-1β and IL-6. In addition, it
was identified that miR-27a suppressed TLR4 by binding to its
3′-UTR, and it was also indicated that sevoflurane exerts an
anti-inflammatory effect on LPS-induced ALI by suppressing the
activation of the TLR4/MyD88/NF-κB signaling pathways.
To date, several in vivo studies have
explored the effects of sevoflurane on lung tissue. For example,
Steurer et al (51) showed
that sevoflurane could attenuate LPS-induced injury by suppressing
the inflammatory response in alveolar macrophages. In addition,
Voigtsberger et al (52)
reported that sevoflurane alleviated lung damage in an in
vivo model of LPS-induced lung injury. Tang et al
(53) demonstrated that
sevoflurane could improve lung function post-acute lung injury by
regulating immune homeostasis in mice. However, the relative
mechanisms by which sevoflurane improve ALI remain unknown. In the
present study, sevoflurane effectively reduced LPS-induced
pulmonary inflammation and resulted in the significant reduction of
LPS-induced increases in lung permeability and the W/D ratio of
lung tissue, accompanied by a significant reduction in the
histopathology changes of the lung. The present results
demonstrated that sevoflurane could protect the lung from
LPS-induced ALI by regulating the inflammatory responses. In
addition, it was also revealed that the expression of the
apoptosis-related protein cleaved caspase 3 was significantly
decreased, while Bcl2 was increased in the sevoflurane treated ALI
mice. However, the possible molecular mechanism requires further
investigation to be fully understood.
Previous studies had demonstrated that sevoflurane
could attenuate LPS-induced ALI via its anti-inflammatory
properties. Voigtsberger et al (52) reported that sevoflurane
ameliorated the gas exchange and attenuated lung damage; this
property appeared to be mediated via the inhibition of lung
inflammation as indicated by the lower levels of cytokines and less
recruitment of effector cells into the lung tissue. Schläpfer et
al (54) revealed that
sevoflurane exposure can positively influence the course of
LPS-induced lung injury with regard to oxygenation and this effect
was mediated by the anti-inflammatory properties of sevoflurane
leading to less edema formation. Hofstetter et al (55) examined the anti-inflammatory
effects of sevoflurane in an in vivo model of LPS-induced
endotoxemia in rats. In this study, the administration of
sevoflurane 15 min after stimulation with LPS resulted in a
decrease in TNF-α and IL-1β plasma levels (55). However, the molecular mechanisms
remain unknown.
Recently, an association has also been reported
between the aberrant expression of miRNAs and exposure to
anesthetics such as sevoflurane (56). For example, Tang et al
(57) reported that miR-29a
regulated LPS-induced inflammatory responses in murine macrophages
through the Akt1/NF-κB signaling pathway. Therefore, given the
important role of sevoflurane in the regulation of miRNAs, it is
possible that miRNAs are also involved in the lung protective
mechanisms of sevoflurane. In the present study, microarray
screening revealed miR-27a to be one of the major miRNAs that were
upregulated in ALI mice by sevoflurane treatment, and that this
upregulation was time-dependent. Notably, previous studies have
reported that downregulation of miR-27a expression attenuated
sevoflurane-induced neuronal apoptosis, and learning and memory
impairment via the upregulation of peroxisome
proliferator-activated receptor-γ, which play important roles in
sevoflurane mediated neurotoxicity and cognitive impairment effects
(21,55). Thus, the present study focused on
miR-27a for further study. Notably, it was confirmed that the
agomir-27a injection produced similar protective effects when
compared with sevoflurane, while the antagomir-27a injection
suppressed the therapeutic effects of sevoflurane in ALI mice.
Taken together, these results indicate that upregulation of miR-27a
may contribute to sevoflurane-induced protective effects against
LPS-induced ALI.
TLR4, a component of the primary innate immune
receptor-mediated inflammatory signaling pathway, has been
associated with various inflammation-related diseases including ALI
(58-60). A previous study has shown that LPS
could induce TLR4 activation and subsequently transmit a signal via
an adaptor molecule, MyD88, leading to the activation of NF-κB,
which in turn causes the upregulation of pro-inflammatory cytokines
such as IL-1β, IL-6 and TNF-α (61). The present results demonstrated
that TLR4 was a direct target of miR-27a. In addition, it was
revealed that LPS induced the upregulation of TLR4, but the
expression of TLR4 was markedly decreased after sevoflurane
treatment. Notably, increased MyD88, nuclear p-p65 and p-IκB-α, and
decreased IκB-α were detected in ALI mice, which was indicative of
the activation of the NF-κB signaling pathway in ALI mice. However,
pretreatment with sevoflurane decreased the expression of MyD88,
nuclear p-p65 and p-IκB-α, and increased the expression of IκB-α in
all ALI mice. Interestingly, the inactivation of NF-κB signaling in
sevoflurane treated ALI mice was re-activated by an injection of
the antagomir-27a. These results indicate that sevoflurane blocked
the activation of the NF-κB signaling pathway through the
miR-27a/TLR4 axis.
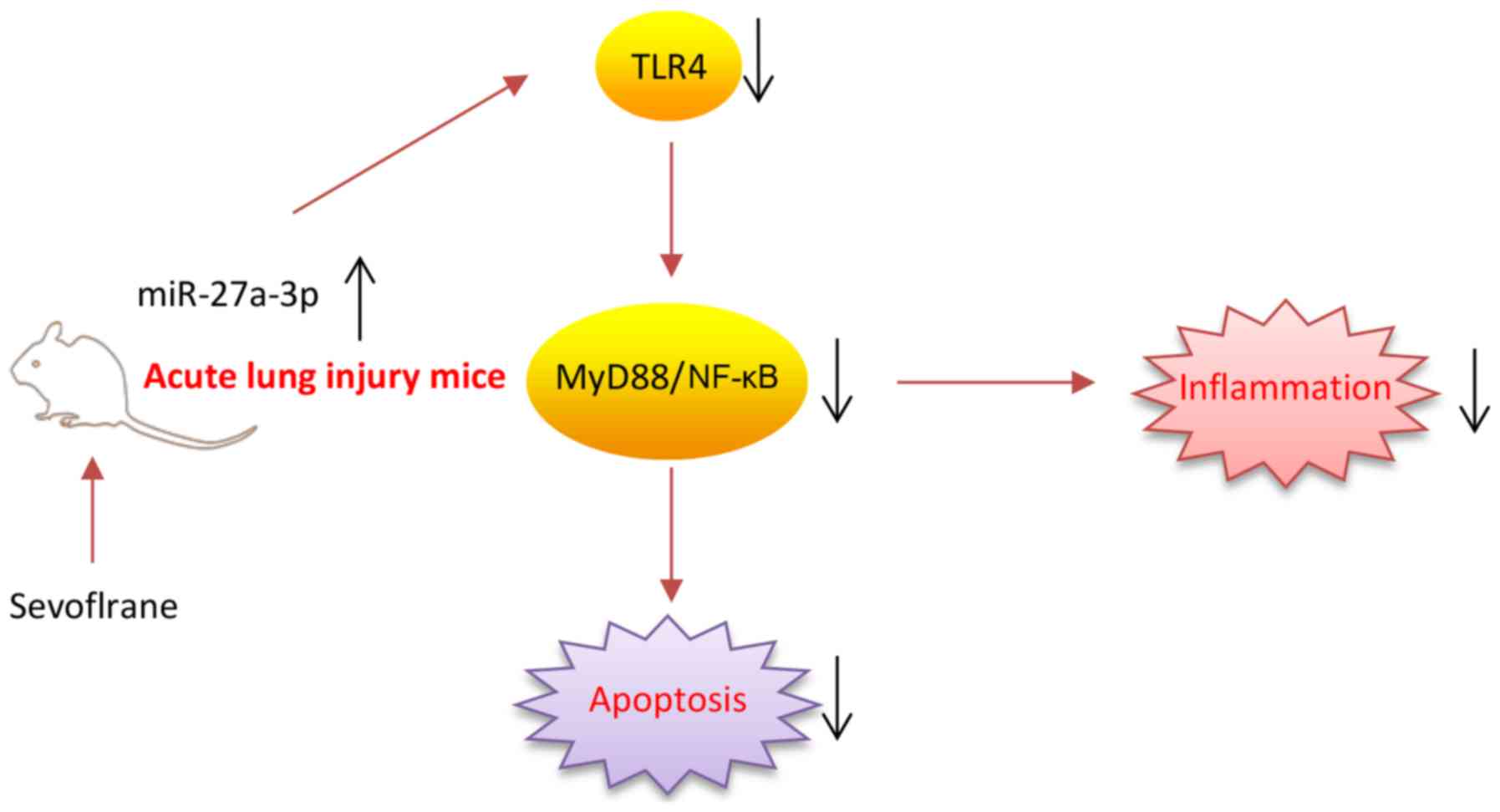
In conclusion, as illustrated in Fig. 7, the present results firstly
demonstrated that sevoflurane effectively protected ALI against
inflammation damage, which was largely dependent on the
upregulation of miR-27a via suppressing the LPS-activated
TLR4/MyD88/NF-κB signaling pathway. The present study provides
beneficial evidence for the application of sevoflurane in the
prevention of ALI. However, the application and efficacy of
sevoflurane in clinical practice remains to be studied.
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XZ, JT, GL, XL and DS performed the experiments,
analyzed the data and wrote the paper. YW conceived and designed
the study, and contributed to data analysis and experimental
materials. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by The First
Affiliated Hospital of Xinxiang Medical University Ethics
Committees (Henan, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rubenfeld GD, Caldwell E, Peabody E,
Weaver J, Martin DP, Neff M, Stern EJ and Hudson LD: Incidence and
outcomes of acute lung injury. N Engl J Med. 353:1685–1693. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brigham KL: Lower tidal volume ventilation
and plasma cytokine markers of inflammation in patients with acute
lung injury. Curr Infect Dis Rep. 7:327–328. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bhatia M and Moochhala S: Role of
inflammatory mediators in the pathophysiology of acute respiratory
distress syndrome. J Pathol. 202:145–156. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Grommes J and Soehnlein O: Contribution of
neutrophils to acute lung injury. Mol Med. 17:293–307. 2011.
View Article : Google Scholar :
|
|
5
|
Kersten JR, Schmeling TJ, Hettrick DA,
Pagel PS, Gross GJ and Warltier DC: Mechanism of myocardial
protection by isoflurane. Role of adenosine triphosphate-regulated
potassium (KATP) channels. Anesthesiology. 85:794–807; discussion
27A. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kersten JR, Schmeling TJ, Pagel PS, Gross
GJ and Warltier DC: Isoflurane mimics ischemic preconditioning via
activation of K(ATP) channels: Reduction of myocardial infarct size
with an acute memory phase. Anesthesiology. 87:361–370. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schlack W, Preckel B, Barthel H, Obal D
and Thamer V: Halothane reduces reperfusion injury after regional
ischaemia in the rabbit heart in vivo. Br J Anaesth. 79:88–96.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Minguet G, Joris J and Lamy M:
Preconditioning and protection against ischaemia-reperfusion in
non-cardiac organs: A place for volatile anaesthetics? Eur J
Anaesthesiol. 24:733–745. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kalimeris K, Zerva A, Matsota P, Nomikos
T, Fragopoulou E, Politi AN, Karamitopoulou E and Kostopanagiotou
G: Pretreatment with sevoflurane attenuates direct lung injury.
Minerva Anestesiol. 80:635–644. 2014.
|
|
10
|
Liu R, Ishibe Y and Ueda M:
Isoflurane-sevoflurane adminstration before ischemia attenuates
ischemia-reperfusion-induced injury in isolated rat lungs.
Anesthesiology. 92:833–840. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kalb R, Schober P, Schwarte LA, Weimann J
and Loer SA: Preconditioning, but not postconditioning, with
Sevoflurane reduces pulmonary neutrophil accumulation after lower
body ischaemia/reperfusion injury in rats. Eur J Anaesthesiol.
25:454–459. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Casanova J, Garutti I, Simon C, Giraldez
A, Martin B, Gonzalez G, Azcarate L, Garcia C and Vara E: The
effects of anesthetic preconditioning with sevoflurane in an
experimental lung autotransplant model in pigs. Anesth Analg.
113:742–748. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Suter D, Spahn DR, Blumenthal S, Reyes L,
Booy C, Z'graggen BR and Beck-Schimmer B: The immunomodulatory
effect of sevoflurane in endotoxin-injured alveolar epithelial
cells. Anesth Analg. 104:638–645. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ohsumi A, Marseu K, Slinger P, McRae K,
Kim H, Guan Z, Hwang DM, Liu M, Keshavjee S and Cypel M:
Sevoflurane attenuates ischemia-reperfusion injury in a rat lung
transplantation model. Ann Thorac Surg. 103:1578–1586. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun XJ, Li XQ, Wang XL, Tan WF and Wang
JK: Sevoflurane inhibits nuclear factor-κB activation in
lipopolysaccha-ride-induced acute inflammatory lung injury via
toll-like receptor 4 signaling. PLoS One. 10:e01227522015.
View Article : Google Scholar
|
|
16
|
Song SY, Zhou B, Yang SM, Liu GZ, Tian JM
and Yue XQ: Preventive effects of sevoflurane treatment on lung
inflammation in rats. Asian Pac J Trop Med. 6:53–56. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Stefani G and Slack FJ: Small non-coding
RNAs in animal development. Nat Rev Mol Cell Biol. 9:219–230. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guo Z, Gu Y, Wang C, Zhang J, Shan S, Gu
X, Wang K, Han Y and Ren T: Enforced expression of miR-125b
attenuates LPS-induced acute lung injury. Immunol Lett. 162:18–26.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen W, Ma X, Zhang P, Li Q, Liang X and
Liu J: MiR-212 3p inhibits LPS-induced inflammatory response
through targeting HMGB1 in murine macrophages. Exp Cell Res.
350:318–326. 2017. View Article : Google Scholar
|
|
21
|
Lv X, Yan J, Jiang J, Zhou X, Lu Y and
Jiang H: MicroRNA-27a 3p suppression of peroxisome
proliferator-activated receptor-γ contributes to cognitive
impairments resulting from sevoflurane treatment. J Neurochem.
143:306–319. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu Y, Gu C and Huang X: Sevoflurane
protects against hepatic ischemia/reperfusion injury by modulating
microRNA-200c regulation in mice. Biomed Pharmacother.
84:1126–1136. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wenlan L, Zhongyuan X, Shaoqing L, Liying
Z, Bo Z and Min L: MiR-34a-5p mediates sevoflurane preconditioning
induced inhibition of hypoxia/reoxygenation injury through STX1A in
cardiomyocytes. Biomed Pharmacother. 102:153–159. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Beck-Schimmer B, Madjdpour C, Kneller S,
Ziegler U, Pasch T, Wüthrich RP, Ward PA and Schimmer RC: Role of
alveolar epithelial ICAM-1 in lipopolysaccharide-induced lung
inflammation. Eur Respir J. 19:1142–1150. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang L, Ye Y, Su HB and Yang JP: The
anesthetic agent sevoflurane attenuates pulmonary acute lung injury
by modulating apoptotic pathways. Braz J Med Biol Res.
50:e57472017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cao YZ, Tu YY, Chen X, Wang BL, Zhong YX
and Liu MH: Protective effect of Ulinastatin against murine models
of sepsis: Inhibition of TNF-α and IL-6 and augmentation of IL-10
and IL-13. Exp Toxicol Pathol. 64:543–547. 2012. View Article : Google Scholar
|
|
27
|
Gao C, Li R and Wang S: Ulinastatin
protects pulmonary tissues from lipopolysaccharide-induced injury
as an immunomodulator. J Trauma Acute Care Surg. 72:169–176.
2012.
|
|
28
|
Luo Y, Che W and Zhao M: Ulinastatin
post-treatment attenuates lipopolysaccharide-induced acute lung
injury in rats and human alveolar epithelial cells. Int J Mol Med.
39:297–306. 2017. View Article : Google Scholar :
|
|
29
|
Xu Z, Zhang C, Cheng L, Hu M, Tao H and
Song L: The microRNA miR-17 regulates lung FoxA1 expression during
lipopolysaccharide-induced acute lung injury. Biochem Biophys Res
Commun. 445:48–53. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Duan Y, Learoyd J, Meliton AY, Leff AR and
Zhu X: Inhibition of Pyk2 blocks lung inflammation and injury in a
mouse model of acute lung injury. Respir Res. 13:42012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Barreto TR, Costola-de-Souza C, Margatho
RO, Queiroz-Hazarbassanov N, Rodrigues SC, Felício LF, Palermo-Neto
J and Zager A: Repeated Domperidone treatment modulates pulmonary
cytokines in LPS-induced acute lung injury in mice. Int
Immunopharmacol. 56:43–50. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang Q, Zhang D and Li Y and Li Y and Li
Y: Paclitaxel alleviated liver injury of septic mice by alleviating
inflammatory response via microRNA-27a/TAB3/NF-κB signaling
pathway. Biomed Pharmacother. 97:1424–1433. 2018. View Article : Google Scholar
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
34
|
Luo C, Yuan D, Zhao W, Chen H, Luo G, Su G
and Hei Z: Sevoflurane ameliorates intestinal
ischemia-reperfusion-induced lung injury by inhibiting the
synergistic action between mast cell activation and oxidative
stress. Mol Med Rep. 12:1082–1090. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu C, Shen Z, Liu Y, Peng J, Miao L, Zeng
W and Li Y: Sevoflurane protects against intestinal
ischemia-reperfusion injury partly by phosphatidylinositol 3
kinases/Akt pathway in rats. Surgery. 157:924–933. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhao W, Gan X, Su G, Wanling G, Li S, Hei
Z, Yang C and Wang H: The interaction between oxidative stress and
mast cell activation plays a role in acute lung injuries induced by
intestinal ischemia-reperfusion. J Surg Res. 187:542–552. 2014.
View Article : Google Scholar
|
|
37
|
Zhou T, Garcia JG and Zhang W: Integrating
microRNAs into a system biology approach to acute lung injury.
Transl Res. 157:180–190. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Martin GS: Sepsis, severe sepsis and
septic shock: Changes in incidence, pathogens and outcomes. Expert
Rev Anti Infect Ther. 10:701–706. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Huang RS, Hu GQ, Lin B, Lin ZY and Sun CC:
MicroRNA-155 silencing enhances inflammatory response and lipid
uptake in oxidized low-density lipoprotein-stimulated human THP-1
macrophages. J Investig Med. 58:961–967. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu DP, Zhang JL, Wang JY, Cui MX, Jia JL,
Liu XH and Liang QD: MiR-1246 promotes LPS-induced inflammatory
injury in chondrogenic cells ATDC5 by targeting HNF4γ. Cell Physiol
Biochem. 43:2010–2021. 2017. View Article : Google Scholar
|
|
41
|
Zhang Q, Su J, Wang Z, Qi H, Ge Z, Li Z,
Chen WD and Wang YD: MicroRNA-149* suppresses hepatic
inflammatory response through antagonizing STAT3 signaling pathway.
Oncotarget. 8:65397–65406. 2017.PubMed/NCBI
|
|
42
|
Satishchandran A, Ambade A, Rao S, Hsueh
YC, Iracheta-Vellve A, Tornai D, Lowe P, Gyongyosi B, Li J,
Catalano D, et al: MicroRNA 122, regulated by GRLH2, protects
livers of mice and patients from ethanol-induced liver disease.
Gastroenterology. 154:238–252.e7. 2018. View Article : Google Scholar
|
|
43
|
Chen H, Li X, Liu S, Gu L and Zhou X:
MircroRNA-19a promotes vascular inflammation and foam cell
formation by targeting HBP-1 in atherogenesis. Sci Rep.
7:120892017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lo WY, Peng CT and Wang HJ:
MicroRNA-146a-5p mediates high glucose-induced endothelial
inflammation via targeting interleukin-1 receptor-associated kinase
1 expression. Front Physiol. 8:5512017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lv YN, Ou-Yang AJ and Fu LS: MicroRNA-27a
negatively modulates the inflammatory response in
lipopolysaccharide-stimulated microglia by targeting TLR4 and
IRAK4. Cell Mol Neurobiol. 37:195–210. 2017. View Article : Google Scholar
|
|
46
|
Cai Q, Wang T, Yang WJ and Fen X:
Protective mechanisms of microRNA-27a against oxygen-glucose
deprivation-induced injuries in hippocampal neurons. Neural Regen
Res. 11:1285–1292. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xue WL, Bai X and Zhang L: rhTNFR:Fc
increases Nrf2 expression via miR-27a mediation to protect
myocardium against sepsis injury. Biochem Biophys Res Commun.
464:855–861. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wu C, Zhao J, Zhu G, Huang Y and Jin L:
SiRNA directed against NF-κB inhibits mononuclear macrophage cells
releasing proinflammatory cytokines in vitro. Mol Med Rep.
16:9060–9066. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhu G, Xin X, Liu Y, Huang Y, Li K and Wu
C: Geraniin attenuates LPS-induced acute lung injury via inhibiting
NF-κB and activating Nrf2 signaling pathways. Oncotarget.
8:22835–22841. 2017.PubMed/NCBI
|
|
50
|
Kawasaki T and Kawai T: Toll-like receptor
signaling pathways. Front Immunol. 5:4612014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Steurer M, Schläpfer M, Steurer M,
Z'graggen BR, Booy C, Reyes L, Spahn DR and Beck-Schimmer B: The
volatile anaesthetic sevoflurane attenuates
lipopolysaccharide-induced injury in alveolar macrophages. Clin Exp
Immunol. 155:224–230. 2009. View Article : Google Scholar :
|
|
52
|
Voigtsberger S, Lachmann RA, Leutert AC,
Schläpfer M, Booy C, Reyes L, Urner M, Schild J, Schimmer RC and
Beck-Schimmer B: Sevoflurane ameliorates gas exchange and
attenuates lung damage in experimental lipopolysaccharide-induced
lung injury. Anesthesiology. 111:1238–1248. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tang QF, Fang ZY and Shi CH: The
protective effect and mechanism of sevoflurane on LPS-induced acute
lung injury in mice. Am J Transl Res. 9:1732–1742. 2017.PubMed/NCBI
|
|
54
|
Schläpfer M, Leutert AC, Voigtsberger S,
Lachmann RA, Booy C and Beck-Schimmer B: Sevoflurane reduces
severity of acute lung injury possibly by impairing formation of
alveolar oedema. Clin Exp Immunol. 168:125–134. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hofstetter C, Boost KA, Flondor M,
Basagan-Mogol E, Betz C, Homann M, Muhl H, Pfeilschifter J and
Zwissler B: Anti-inflammatory effects of sevoflurane and mild
hypothermia in endotoxemic rats. Acta Anaesthesiol Scand.
51:893–899. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Tanaka S, Ishikawa M, Arai M, Genda Y and
Sakamoto A: Changes in microRNA expression in rat lungs caused by
sevoflurane anesthesia: A TaqMan® low-density array
study. Biomed Res. 33:255–263. 2012. View Article : Google Scholar
|
|
57
|
Tang B, Li X, Ren Y, Wang J, Xu D, Hang Y,
Zhou T, Li F and Wang L: MicroRNA-29a regulates lipopolysaccharide
(LPS)-induced inflammatory responses in murine macrophages through
the Akt1/NF-κB pathway. Exp Cell Res. 360:74–80. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Li J, Bao L, Zha D, Zhang L, Gao P, Zhang
J and Wu X: Oridonin protects against the inflammatory response in
diabetic nephrop-athy by inhibiting the TLR4/p38-MAPK and
TLR4/NF-κB signaling pathways. Int Immunopharmacol. 55:9–19. 2018.
View Article : Google Scholar
|
|
59
|
Yan J, Li J, Zhang L, Sun Y, Jiang J,
Huang Y, Xu H, Jiang H and Hu R: Nrf2 protects against acute lung
injury and inflammation by modulating TLR4 and Akt signaling. Free
Radic Biol Med. 121:78–85. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Deng G, He H, Chen Z, OuYang L, Xiao X, Ge
J, Xiang B, Jiang S and Cheng S: Lianqinjiedu decoction attenuates
LPS-induced inflammation and acute lung injury in rats via
TLR4/NF-κB pathway. Biomed Pharmacother. 96:148–152. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Suzuki Y, Ruiz-Ortega M, Lorenzo O,
Ruperez M, Esteban V and Egido J: Inflammation and angiotensin II.
Int J Biochem Cell Biol. 35:881–900. 2003. View Article : Google Scholar : PubMed/NCBI
|