Introduction
Characterized by diffuse inflammatory responses,
sepsis is usually caused by a fungal, viral or bacterial infection.
According to the most recent data, sepsis is the 7th and 8th
highest contributor to mortality in children aged 1-4 years and
adults aged 65-75 years worldwide, respectively. In addition,
sepsis is the main contributor to mortality in patients with a
critical illness (1). For
example, as one of the most common and major complications in
patients with sepsis, acute kidney injury (AKI) involves the rapid
failure of the kidneys to generate sufficient urine, thus impairing
the functions of the kidneys to regulate water and ion balance by
filtering the blood. AKI substantially elevates healthcare costs
and worsens the prognosis of sepsis. For example, the rate of AKI
increases with the increasing seriousness of sepsis, and it is
estimated that AKI occurs in 64% of patients with serious
hypotension and sepsis within the first 24 h of the disease.
A decade ago, the inflammasome was considered a
major intracellular signaling node containing a receptor for the
recognition of cytosolic patterns. In particular, the inflammasome
carries absent in melanoma 2 (AIM2)-like receptors and
nucleotide-binding oligomerization domain leucine-rich repeat
containing receptors (NLRs). The NLR pyrin domain-containing 3
(NLRP3) inflammasome has been the most extensively investigated NLR
inflammasome complex, and it acts as a critical signaling platform
regulating the maturation of interleukin (IL)-1β and IL-18, two
proinflammatory cytokines in the IL-1 family (2). The stimulation of NLRP3 results in
the recruitment of the apoptosis-associated speck-like protein
containing a C-terminal caspase recruitment domain (ASC), and leads
to the transformation of pro-caspase-1 into its cleaved type
(2). Identified as an
inflammatory caspase, caspase-1 is important for the maturation of
IL-18 and IL-1β into active cytokines. In addition, caspase-1
mediates pyroptosis through its effect on activation and
autocatalysis (3). The roles of
IL-18 and IL-1 in renal diseases are better understood, although
the significance of the inflammasome in the secretion and
activation of IL-18 and IL-1β has only been noted relatively
recently. A range of primary kidney conditions are associated with
the activation of NLRP3 inflammasomes. Similarly, several systemic
conditions involving the kidneys, including hypoxia, chronic kidney
disease, sepsis, glomerulonephritis, ischemia-reperfusion (I-R)
injury, unilateral ureteral obstruction (UUO), crystal nephropathy
and glycerol-induced renal failure (4-11),
are associated with activation of the NLRP3
IL-1β/IL-18/inflammasome axis. Caspase-9, a caspase for the
initialization of mitochondrial apoptosis, is not necessary for the
secretion of IL-1β induced by apoptotic stimuli and is not involved
in the activation of canonical NLRP3. Furthermore, the absence of
NLRP3, caspase-1 or ASC (encoded by Pycard) substantially delays
the cell death triggered by nigericin and ATP, suggesting that
these NLRP3 stimuli primarily mediate cell death by pyroptosis in
this time frame but do not affect the apoptosis of mitochondria
(12). The pyroptosis-induced
cell death occurs independently of mitochondria by the ASC
recruitment of caspase-8, and the apoptosis triggered by NLRP3
stimuli is substantially dependent on the activation of its
upstream NLRP3 (12). As a potent
antioxidant and a scavenger of free radicals (13), melatonin
(N-acetyl-5-methoxytryptamine; MT) is actively taken up by the
mitochondria to maintain its homeostasis. In addition, MT has
antiseptic effects, targeting the activation of the innate immune
pathway involving nuclear factor (NF)-κB (14-17). It has been demonstrated that MT
also reduces the activation of NLRP3 inflammasomes in a number of
conditions assessed (18).
It has been shown that MT administration can reduce
the activation of inflammasomes during sepsis by suppressing the
expression of NLRP3 (19).
Furthermore, activated inflammasomes are considered to be
responsible for sepsis-induced renal injury (20). In the present study, the
therapeutic effect of MT in sepsis-induced renal injury, and its
effect on the expression of PTEN-induced putative kinase 1 (PINK1),
Parkin1, NLRP3, cleaved caspase-1 and ASC were evaluated in
rats.
Materials and methods
Animals
All experiments were performed according to the
protocol approved by the Ethical Committee of Yancheng City No. 1
People's Hospital (Yancheng, China). Obtained from the Laboratory
Animal Facility of Chinese Academy of Sciences (Yangzhou, China),
all Sprague Dawley rats were 10-12 weeks old with a body mass of
20-30 g. All animals were housed in an animal facility with a 12/12
h light-dark cycle at 25°, and had free access to rodent chow and
water.
Cecal ligation and puncture (CLP)
model
A CLP model was used to induce sepsis. In brief,
equithensin (1 ml/kg) was used to anesthetize the animals by
intraperitoneal perfusion, and the rats were fixed on an aseptic
operating table. Their cecum was exposed through a 1-cm midline
incision in a sterile environment. Subsequently, the cecum was
ligated tightly below the ileocecal valve using a 4-0 silk suture.
In the subsequent step, a 22-gauge needle was used to puncture the
cecum three times to ensure the same severity of sepsis.
Subsequently, the cecum was squeezed gently to excrete a small
quantity of stool through the puncture site. In the subsequent
step, the cecum was placed back into the peritoneal cavity and the
abdominal incision was gently stitched in two layers. Finally, 1 ml
saline solution was used to resuscitate the animals via
subcutaneous injection.
Experimental protocol
All the animals were divided into three groups: A
control group (C), a septic group (S), and a septic group
administered with MT (S + MT). The rats in the S + MT group were
treated with the following three doses of MT-containing 30% PEG
(each dose, 150 mg/kg): The first dose was administered
intraperitoneally at 30 min prior to surgery, the second dose was
administered immediately following surgery through subcutaneous
injection, and the final dose was administered at 4 h post-surgery
through subcutaneous injection. All animals were sacrificed at 8 h
post-CLP to immediately remove the heart, which was rinsed in cold
saline and treated to extract the cytosol and nuclei for
histological assessment. In order to avoid diurnal variations, all
rats underwent surgery between 8:00 a.m. and 12:00 p.m. The MT dose
was selected accordingly to inhibit the expression of inducible
nitric oxide synthase, a crucial factor of the innate immune in
septic rats undergoing CLP.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
An MirVana™ miRNA isolation kit (Roche Diagnostics
GmbH, Mannheim, Germany) was used to extract total RNA from tissue
samples following the manufacturer's protocol. A High-Capacity cDNA
reverse transcription kit (Applied Biosystems; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) was used to perform reverse
transcription to synthesize cDNA (PINK1 or NLRP3) following the
protocol of the manufacturer. An ABI 7500 Real-Time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.) was used in
conjunction with an miScript SYBR®-Green PCR kit (Qiagen
GmbH, Hilden, Germany) to measure the expression of PINK1 (forward,
5′-CCAAGCTGGCTAGCGTTTAAACG-3′; reverse,
5′-ATGGTCGACGGCGCTATTCAG-3′) and NLRP3 (forward,
5′-GTTTTCATTCCTGCACTGCCAGTG-3′; reverse,
5′-CAAAAACCCTTCTGTTTACTCACTC-3′). The reaction was performed using
10X Standard Taq Reaction Buffer (5 µl), 10 mM dNTPs (1
µl), 10 µM forward primer (1 µl), 10 µM
reverse primer (1 µl), template DNA (10 ng), Taq DNA
polymerase (0.25 µl; cat. no. 600280, Agilent Technologies,
Inc., Santa Clara, CA, USA) and nuclease-free water (50 µl)
as follows: 15 min of initial activation at 95°C and 15 sec of
denaturation at 94°C, followed by 30 sec of annealing at 55°C and a
final extension for 60 sec at 72°C. GAPDH (forward,
5′-AGAAGGCTGGGGCTCATTTG-3′; reverse, 5′-AGGGGCCATCCACAGTCTTC-3′)
was used as an internal control to normalize the expression of
PINK1 and NLRP3. Finally, the mRNA expression levels of PINK1 and
NLRP3 were calculated using the 2−ΔΔCq method (21). All reactions were run three
times.
ELISA
IL-1β, IL-18 and IL-6 ELISA detection kits (R&D
systems, Inc., Minneapolis, MN, USA) were used to determine the
levels of IL-1β, IL-18 and IL-6 following the protocol of the
supplier.
Western blot analysis
Ice-cold phosphate-buffered saline (PBS; Hyclone; GE
Healthcare Life Sciences, Logan, UT, USA) was used to wash the
homogenized tissues, followed by lysis in a RIPA lysis buffer
supplemented with 150 mM NaCl, 50 mM Tris-HCl (pH 7.4), 0.1% SDS,
1% NP-40 and protease inhibitor cocktails (Roche Diagnostics GmbH).
Subsequently, the lysate was centrifuged at 25,155 × g and 4°C for
15 min, and boiled for 10 min in 2-mercaptoethanol. The
Bicinchoninic Acid Assay kit (Thermo Fisher Scientific, Inc.) was
used to determine protein concentration, based on the concentration
of the standard protein supplied in the kit. The proteins in the
lysates were resolved using 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and subsequently
eletrotransferred onto nitrocellulose membranes (EMD Millipore,
Billerica, MA, USA) by 35 µg/lane. Subsequently, the
membranes were blocked at room temperature for 90 min in TBST
containing 20 mM Tris-HCl (pH 7.5) and 0.1% Tween-20 containing 10%
dry milk, and washed three times in TBST. Primary rabbit anti-PINK1
(cat. no. ab23707), NLRP3 (cat. no. ab214185), ASC (cat. no.
ab18193) and caspase-1 antibodies (cat. no. ab1872, 1:5,000
dilution, Abcam, Cambridge, UK) and mouse anti-β-actin antibodies
(cat. no. ab8226, 1:8,000 dilution, Abcam) were used to incubate
the membranes at room temperature for 2 h, followed by three TBST
washes. The horseradish peroxidase (HRP)-linked anti-mouse and
anti-rabbit IgG secondary antibodies at a dilution of 1:15,000
(cat. nos. AC111P and AQ132P, Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) were used to treat the membrane for 2 h at room
temperature. An ODYSSEY infrared imaging system (LI-COR
Biosciences, Lincoln, NE, USA) was used to visualize protein
signals. All experiments were performed in triplicate.
Immunohistochemistry (IHC)
Kidney samples were harvested, fixed in 4%
paraformaldehyde for 48 h, dehydrated, embedded in paraffin and
sectioned (size, 0.4 µm). Gradient ethanol was used to dewax
and hydrate the samples, followed by 25 min of antigen retrieval in
10 mM citrate (pH 6.0) under a 720 W heating condition in a
microwave. Subsequently, the sections were cooled at room
temperature for 30 min, blocked in 3% hydrogen peroxide for 15 min,
and incubated at room temperature for 2 h with primary goat
anti-PINK1 and NLRP3 (1:500 dilution, Abcam) antibodies. Following
sample washing with TBST, the HRP-linked anti-goat IgG secondary
antibodies at a dilution of 1:1,500 (Sigma; EMD Millipore) were
used to treat the samples for 2 h at room temperature. A
3,3′-diaminobenzidine substrate kit (Vector Laboratories, Inc.,
Burlingame, CA, USA) was used to visualize bound antibodies, and
hematoxylin was used to stain the cell nuclei.
Hematoxylin and eosin (H&E)
staining
At 48 h post-CLP, the kidney samples were harvested,
fixed in 4% paraformaldehyde for 48 h, dehydrated, embedded in
paraffin and sectioned. H&E was used to stain the sections,
which were then viewed under a light microscope.
TUNEL
The apoptotic cells were examined using TUNEL assays
(DeadEnd™ Fluorometric TUNEL system, Promega Corporation) in
accordance with the manufacturer's protocol. Subsequently,
TUNEL-positive cells in each tissue samples were determined using
HPFs (×400 magnification). Three independent tests were run.
Statistical analysis
All data are presented as the mean ± standard
deviation, and SPSS (version 11.5; SPSS, Inc., Chicago, IL, USA)
was used to perform statistical analysis. All results were original
and strictly verified for their correctness. Two-way analysis of
variance with a Holms-Sidak post hoc test was used to perform the
statistical comparisons between groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
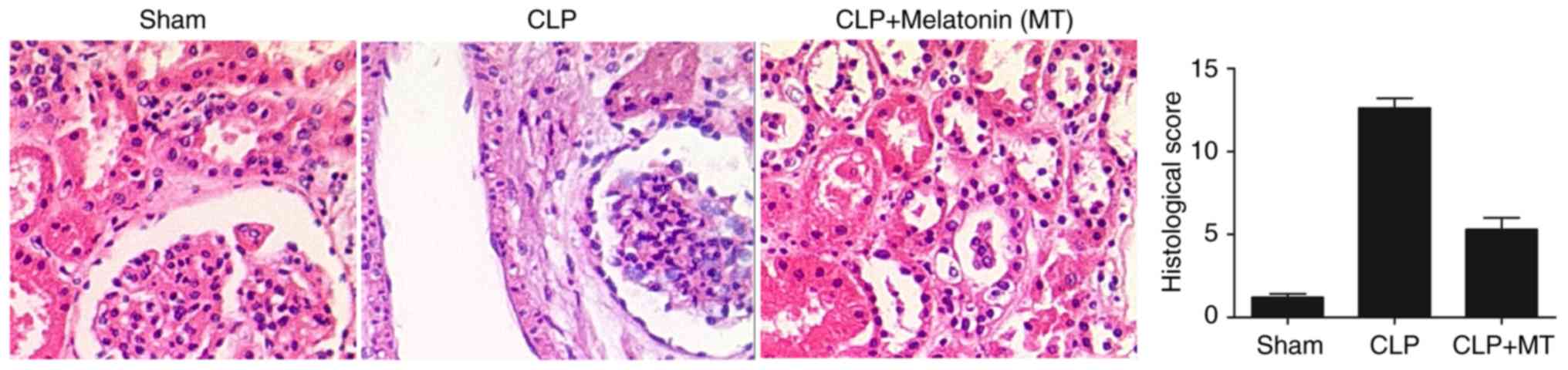
Injury of renal tissue differs among
treatment groups
The animals were divided into three groups: A sham
group, a septic group induced by CLP (CLP group) and a septic group
treated with MT (CLP + MT group). The renal tissues were collected
from all rats, and H&E staining was performed to determine the
extent of tissue damage among the sham, CLP and CLP + MT groups. As
shown in Fig. 1, the extent of
tissue damage in the CLP group was the highest and the extent of
tissue damage in the sham group was the lowest.
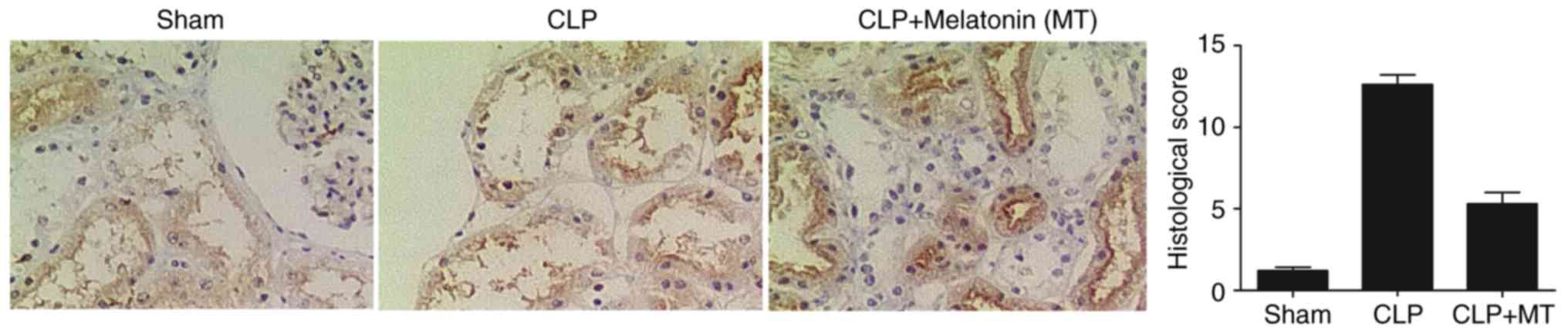
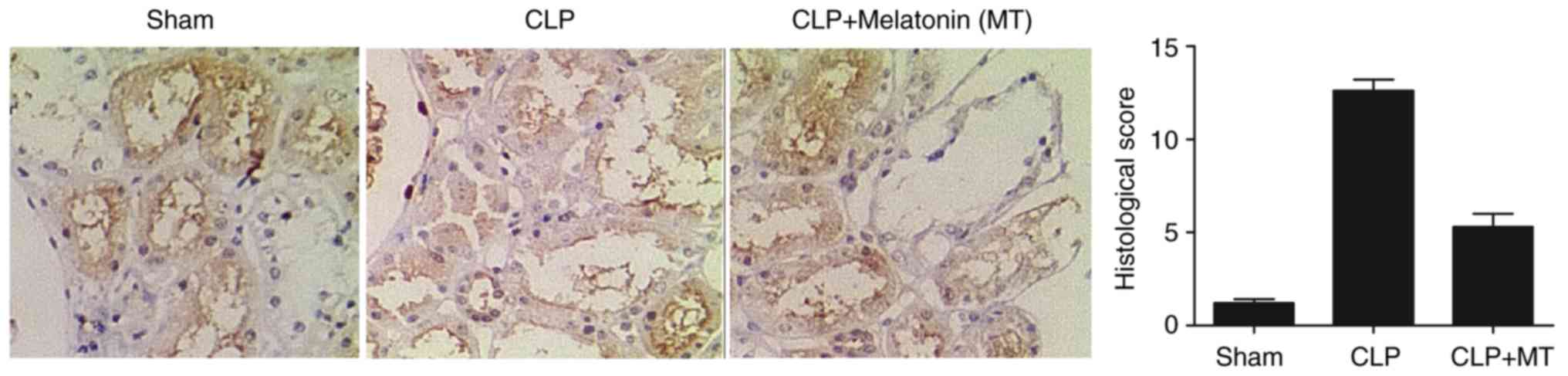
Differential expression of PINK1 and
NLRP3 among treatment groups
IHC was performed to examine the protein expression
of PINK3 and NLRP3 among the sham, CLP, and CLP + MT groups. As
shown in Fig. 2, the protein
level of PINK1 was highest in the CLP group, whereas the sham group
showed the lowest protein level of PINK1. As shown in Fig. 3, the sham group showed the lowest
protein level of NLRP3, whereas he CLP group exhibited the highest
protein level of NLRP3.
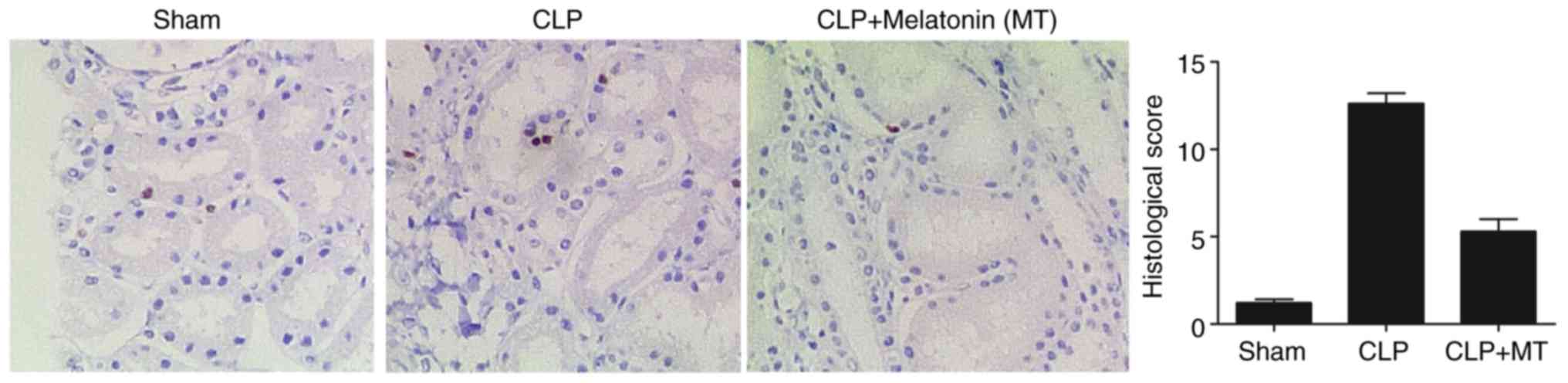
Apoptosis in the renal tissues among
treatment groups
A TUNEL assay was performed to detect the apoptosis
of renal cells in the samples from the sham, CLP, and CLP + MT
groups. As shown in Fig. 4, CLP
treatment increased cell apoptosis in the renal tissues compared
with that in the sham group, whereas the administration of MT
partially reduced the level of cell apoptosis.
Levels of PINK1, NLRP3, ASC1 and cleaved
caspase-1 are altered among treatment groups
Western blot analysis was performed to measure the
protein levels of PINK1, NLRP3, ASC1 and cleaved caspase-1 among
the sham, CLP, and CLP + MT groups. As shown in Fig. 5, the protein level of PINK1
(Fig. 5A and B) in the sham group
was the lowest, whereas the protein level of PINK1 in the CLP group
was the highest. By contrast, the levels of ASC1 (Fig. 5A and C), cleaved caspase-1
(Fig. 5A and D) and NLRP3
(Fig. 5A and E) in the sham group
were the lowest, whereas the levels of these proteins in the CLP
group were the highest. These results suggest that CLP increased
the protein levels of NLRP3, ASC1 and cleaved caspase-1, and
decreased the protein level of PINK1.
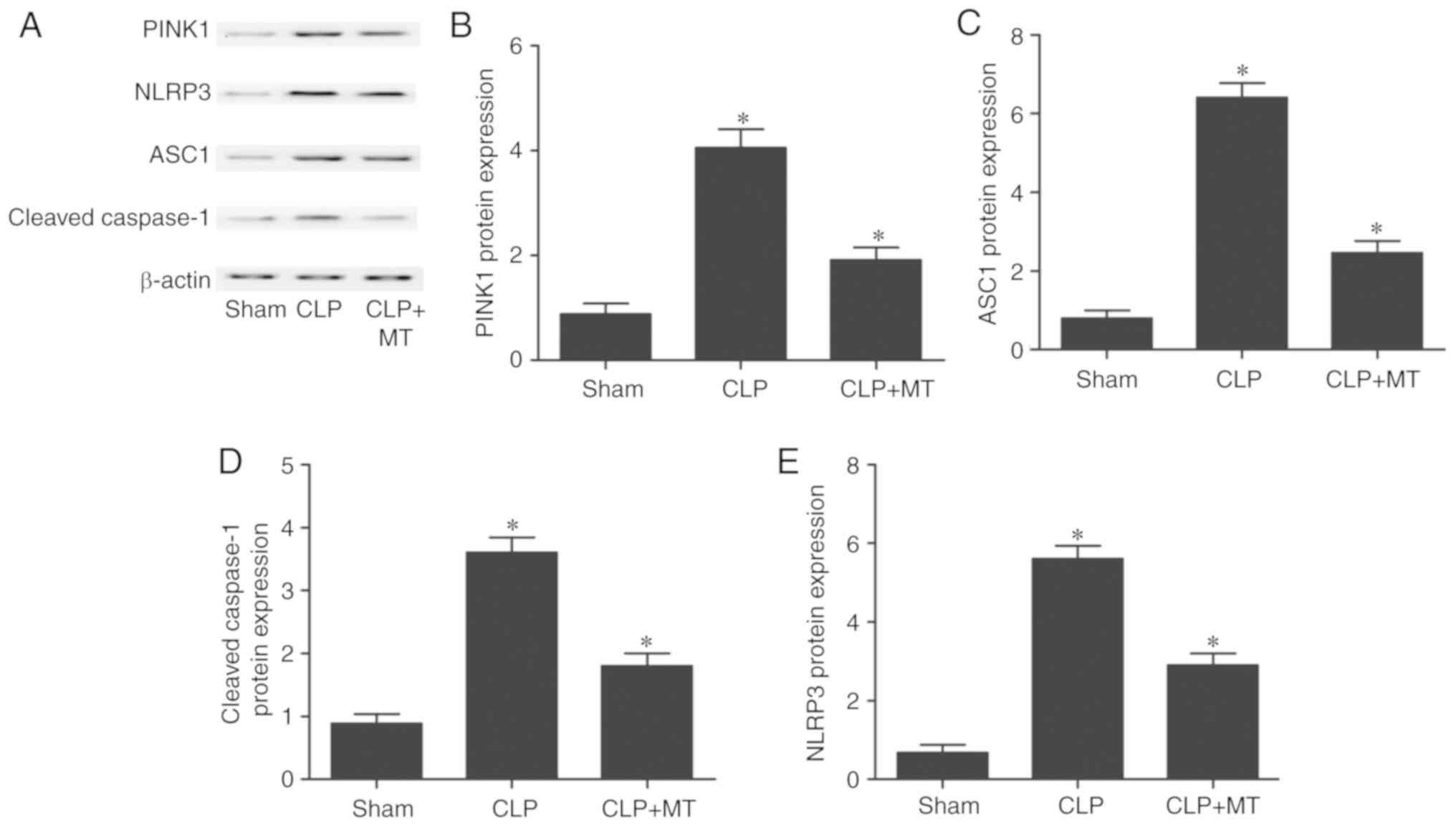 | Figure 5Comparison of protein levels of
PINK1, NLRP3, ASC1 and cleaved caspase-1 among the sham, CLP, and
CLP + MT groups. Western blot analysis was performed to compare
protein levels (one-way analysis of variance,
*P<0.05, compared with the control). (A) CLP
treatment increased the expression of NLRP3, ASC1, cleaved
caspase-1 and PINK1. MT administration showed the opposite trend.
(B) CLP treatment upregulated the expression of PINK1, which was
partially reduced by MT. (C) Protein level of ASC was highest in
the CLP group and lowest in the sham group. (D) Protein level of
cleaved caspase-1 was highest in the CLP group and lowest in the
sham group. (E) Protein level of NLRP3 was highest in the CLP group
and lowest in the sham group. CLP, cecal ligation and puncture; MT,
melatonin; PINK1, PTEN-induced putative kinase 1; NLRP3,
nucleotide-oligomerization binding domain and leucine-rich repeat
pyrin domain-containing 3; ASC1, apoptosis-associated speck-like
protein, containing a C-terminal caspase recruitment domain. |
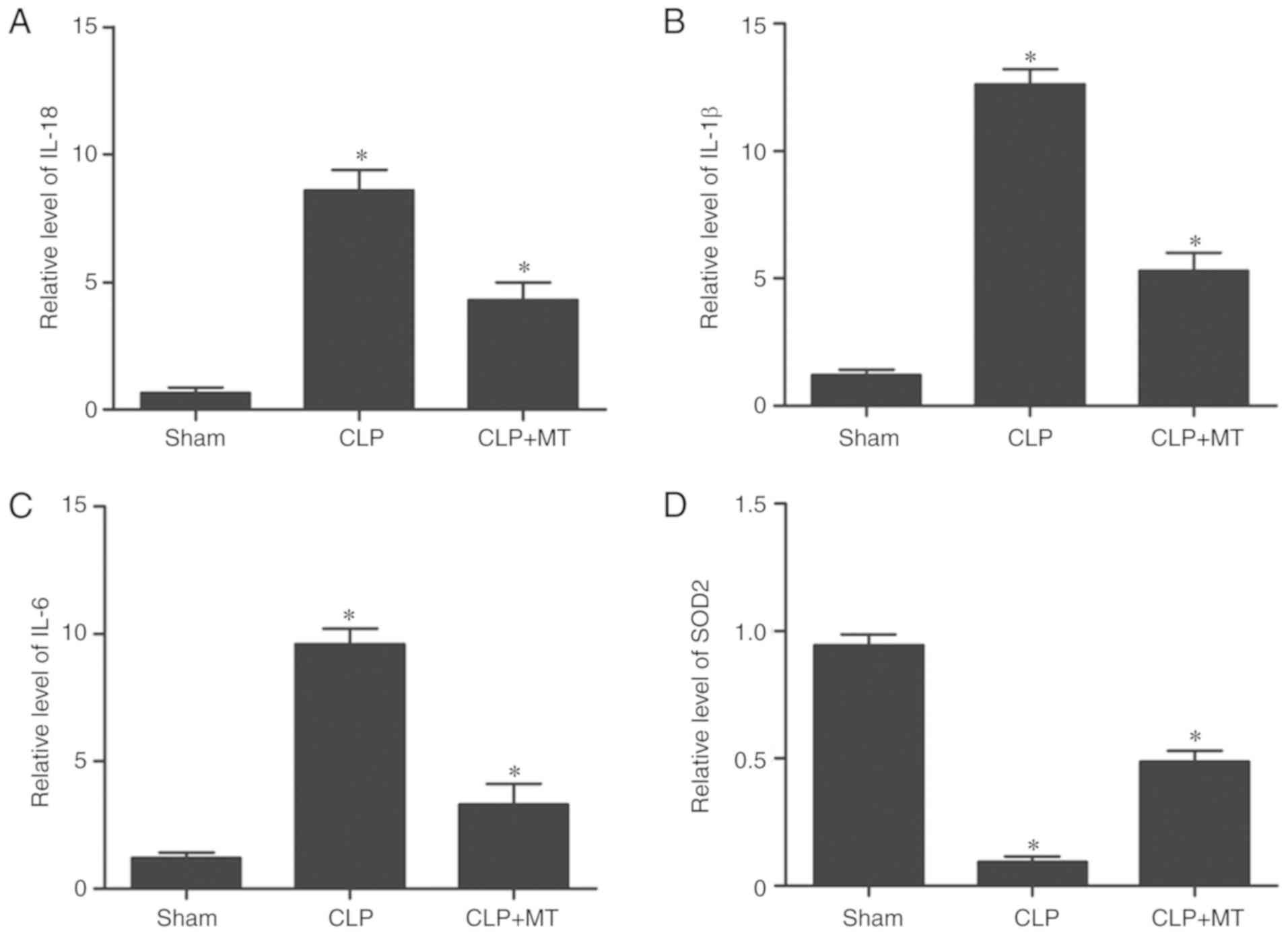
Levels of IL-18, IL-1β and IL-6 among
treatment groups
ELISA was used to examine the protein levels of
IL-18, IL-1β and IL-6 among the sham, CLP, and CLP + MT groups. As
shown in Fig. 6A-C, the sham
group showed the lowest protein levels of IL-18, IL-1β and IL-6,
whereas the CLP group showed the highest protein levels of IL-18,
IL-1β and IL-6, indicating that CLP downregulated the levels of
IL-18, IL-1β and IL-6. Superoxide dismutase 2 (SOD2) was
downregulated in the CLP group compared with that in the sham
group, whereas MT treatment increased the downregulated expression
of SOD2 in the CLP group.
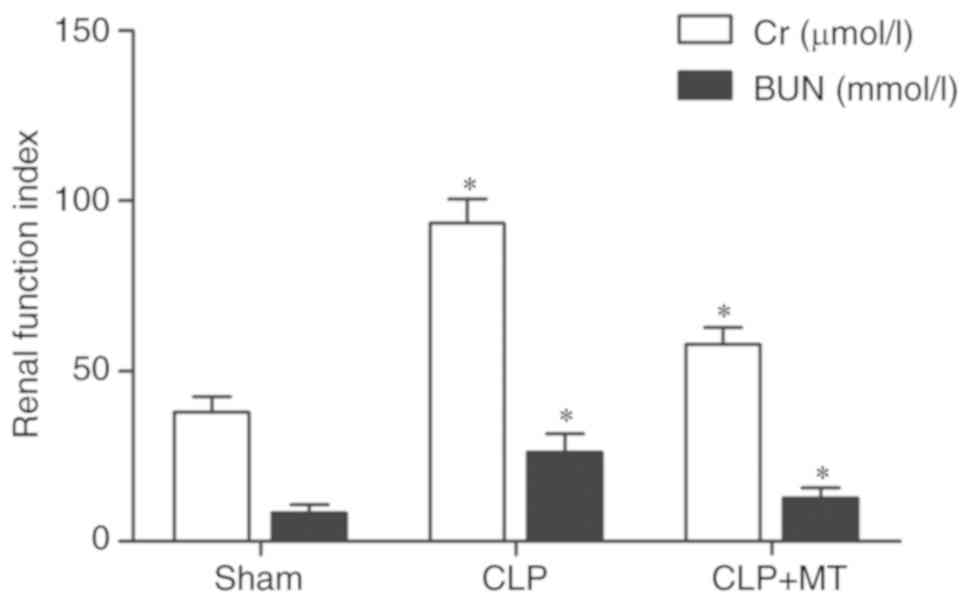
Effect of MT treatment on renal
functions
In the CLP group, the levels of blood urea nitrogen
(BUN) and creatinine (Cr) were substantially increased, whereas
treatment with MT partially restored the renal functions, as shown
in Fig. 7.
Discussion
It is well known that patients with sepsis are
particularly at risk of AKI-induced death. The pathophysiological
role of AKI in sepsis is complex and involves multiple factors,
including reactive oxygen species (ROS), chemokines, and
pro-inflammatory cytokines (22).
Several studies have demonstrated that the dysregulated activation
of NLRP3 inflammasomes is associated with the pathogenesis of AKI
and sepsis (23). Prior studies
have also shown that dysregulated apoptosis is the main
pathophysiological mechanism underlying the progression of mild
inflammatory conditions to severe inflammatory conditions (24,25).
Inflammasomes modulate the processing of pro-IL-18
and pro-IL-1β by caspase-1, and induce inflammatory cell death
(pyroptosis) (26,27). The wide range of these activating
stimuli suggests that NLRP3-dependent responses are controlled by
complex regulatory mechanisms. The generation of ROS and the
modification of thioredoxin interacting protein, TXNIP, have been
demonstrated to cooperatively 'authorize' NLRP3 inflammasomes to
process IL-1β (28). Previous
findings have also indicated that caspase 11 and caspase 8 can
affect the reaction of NLRP3 inflammasomes to certain
'non-canonical' stimuli derived from pathogens (29).
MT was first isolated in 1958 as a neuro-hormone
primarily synthesized and released from the pineal gland (30). MT is an indoleamine synthesized
from tryptophan, an essential amino acid (31). MT has been demonstrated to exert
anti-inflammatory effects in the skin, intestine, and the lungs; it
can also assist in relieving chronic pelvic pain in women with
endometriosis (32-34). It has been shown that, acting as
an anti-apoptotic and anti-oxidization agent, MT can decrease the
kidney damage induced by I-R, UUO, and severe burns (35-37). It has been shown that, by
suppressing nucleocytoplasmic translocation, pretreatment with MT
can enhance renal regeneration in folic acid-induced AKI, thus
providing further insight on the treatment of AKI (38). In the present study, a sepsis rat
model was established by CLP treatment, and H&E staining was
performed to detect the extent of renal tissue damage. The results
revealed that the extent of tissue damage in the CLP group was the
highest, whereas the extent of tissue damage in the sham group was
the lowest. Furthermore, IHC and western blot analyses were
performed to determine the protein levels of PINK1 and NLRP3 among
the sham, CLP, and CLP + MT groups. The results showed that the CLP
group was associated with the highest level of PINK1 and the
highest level of NLRP3. In addition, TUNEL assays were performed to
detect the apoptosis of renal cells in the tissue samples collected
from the sham, CLP, and CLP + MT groups. The results revealed that
the CLP group exhibited the highest apoptotic rate, whereas the
sham group exhibited the lowest apoptotic rate.
Previously, the E3 ubiquitin ligase Parkin and PINK1
were found to trigger mammalian mitophagy. Genetic studies in
Drosophila have further revealed that Parkin and PINK1 have
regulatory role in mitochondrial integrity (39). Parkin and PINK1 also facilitate
the repair of mitochondrial injury by chemical agents, including
carbonyl cyanide m-chlorophenyl-hydrazone (40). The Parkin and PINK1 pathway of
mitophagy is also involved in the modulation of mitochondrial
actions. For example, mitochondrial depolarization triggers the
binding of Parkin/PINK1 to a mitochondrial adaptor protein known as
mitochondria Rho-GTPase, which anchors a kinesin motor complex to
the surface of mitochondria (41). Increasing evidence indicates that
the interplay of Parkin-NF-κB counteracts the effect
NLRP3-inflammasomes, which can stimulate a wide range of exogenous
and endogenous stimuli to result in the caspase 1-triggered
maturation of cytokines IL-18 and IL-1β. A previous study
demonstrated the role of dopamine as an endogenous suppressor of
NLRP3 inflammasome stimulation via microglial cells and dopamine
receptor signalling (42). NF-κB
not only primed the inflammasomes by mediating the production of
pro-IL-1β and NLRP3, but also prevented the excessive activation of
NLRP3 by enhancing Parkin-dependent mitophagy. In the present
study, ELISA was performed to compare the protein levels of IL-18,
IL-1β and IL-6 among the sham, CLP, and CLP + MT groups. The
results showed that the CLP group was associated with the highest
protein levels of IL-18, IL-1β and IL-6, whereas the sham group was
associated with the lowest protein levels of IL-18, IL-1β and
IL-6.
In conclusion, MT-mediated cell apoptosis protects
against sepsis-induced renal injury by affecting the PINK1/Parkin1
signaling pathway induced by NLRP3 inflammasomes. The data of the
present study showed that MT administration mitigated the
activation of inflammasomes in sepsis by suppressing the expression
of NLRP3 via inhibiting the PINK1/Parkin1 signaling pathway. The
overexpression of NLRP3 promoted the secretion of inflammatory
cytokines (IL-1β and IL-18), which promoted cell apoptosis and
subsequently induced renal injury in sepsis.
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WD, HH and YD designed the study and prepared the
manuscript. LS and SH collected the literature, collected the data
and prepared the manuscript. LZ and LX analyzed and generated the
figures. All authors approved the final manuscript.
Ethics approval and consent to
participate
All experiments were performed according to the
protocol approved by the Ethical Committee of Sheyang Zhenyang
Hospital (Yancheng, China).
Patient consent for publication
Not applicable.
Competing interests
The authors confirm that they have no competing
interests.
References
|
1
|
Hotchkiss RS and Karl IE: The
pathophysiology and treatment of sepsis. N Engl J Med. 348:138–150.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Davis BK, Wen H and Ting JP: The
inflammasome NLRs in immunity, inflammation, and associated
diseases. Annu Rev Immunol. 29:707–735. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lamkanfi M: Emerging inflammasome effector
mechanisms. Nat Rev Immunol. 11:213–220. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Edelstein CL, Hoke TS, Somerset H, Fang W,
Klein CL, Dinarello CA and Faubel S: Proximal tubules from
caspase-1-deficient mice are protected against hypoxia-induced
membrane injury. Nephrol Dial Transplant. 22:1052–1061. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vilaysane A, Chun J, Seamone ME, Wang W,
Chin R, Hirota S, Li Y, Clark SA, Tschopp J, Trpkov K, et al: The
NLRP3 inflammasome promotes renal inflammation and contributes to
CKD. J Am Soc Nephrol. 21:1732–1744. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Granfeldt A, Ebdrup L, Tønnesen E and
Wogensen L: Renal cytokine profile in an endotoxemic porcine model.
Acta Anaesthesiol Scand. 52:614–620. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen A, Sheu LF, Chou WY, Tsai SC, Chang
DM, Liang SC, Lin FG and Lee WH: Interleukin-1 receptor antagonist
modulates the progression of a spontaneously occurring IgA
nephropathy in rats. Am J Kidney Dis. 30:693–702. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Furuichi K, Wada T, Iwata Y, Kokubo S,
Hara A, Yamahana J, Sugaya T, Iwakura Y, Matsushima K, Asano M, et
al: Interleukin-1- dependent sequential chemokine expression and
inflammatory cell infiltration in ischemia-reperfusion injury. Crit
Care Med. 34:2447–2455. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bani-Hani AH, Leslie JA, Asanuma H,
Dinarello CA, Campbell MT, Meldrum DR, Zhang H, Hile K and Meldrum
KK: IL-18 neutralization ameliorates obstruction-induced
epithelial-mesenchymal transition and renal fibrosis. Kidney Int.
76:500–511. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mulay SR, Kulkarni OP, Rupanagudi KV,
Migliorini A, Darisipudi MN, Vilaysane A, Muruve D, Shi Y, Munro F,
Liapis H and Anders HJ: Calcium oxalate crystals induce renal
inflammation by NLRP3-mediated IL-1β secretion. J Clin Invest.
123:236–246. 2013. View
Article : Google Scholar
|
|
11
|
Homsi E, Janino P and de Faria JB: Role of
caspases on cell death, inflammation, and cell cycle in
glycerol-induced acute renal failure. Kidney Int. 69:1385–1392.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Allam R, Lawlor KE, Yu EC, Mildenhall AL,
Moujalled DM, Lewis RS, Ke F, Mason KD, White MJ, Stacey KJ, et al:
Mitochondrial apoptosis is dispensable for NLRP3 inflamma-some
activation but non-apoptotic caspase-8 is required for inflammasome
priming. EMBO Rep. 15:982–990. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Galano A, Tan DX and Reiter RJ: On the
free radical scavenging activities of melatonin's metabolites, AFMK
and AMK. J Pineal Res. 54:245–257. 2013. View Article : Google Scholar
|
|
14
|
López A, García JA, Escames G, Venegas C,
Ortiz F, López LC and Acuña-Castroviejo D: Melatonin protects the
mitochondria from oxidative damage reducing oxygen consumption,
membrane potential, and superoxide anion production. J Pineal Res.
46:188–198. 2009. View Article : Google Scholar
|
|
15
|
Acuña Castroviejo D, López LC, Escames G,
López A, García JA and Reiter RJ: Melatonin-mitochondria interplay
in health and disease. Curr Top Med Chem. 11:221–240. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Crespo E, Macías M, Pozo D, Escames G,
Martin M, Vives F, Guerrero JM and Acuña Castroviejo D: Melatonin
inhibits expression of the inducible NO synthase II in liver and
lung and prevents endotoxemia in lipopolysaccharide-induced
multiple organ dysfunction syndrome in rats. FASEB J. 13:1537–1546.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Escames G, López LC, Tapias V, Utrilla P,
Reiter RJ, Hitos AB, León J, Rodríguez MI and Acuña Castroviejo D:
Melatonin counteracts inducible mitochondrial nitric oxide
synthase-dependent mitochondrial dysfunction in skeletal muscle of
septic rats. J Pineal Res. 40:71–78. 2006. View Article : Google Scholar
|
|
18
|
García JA, Volt H, Venegas C, Doerrier C,
Escames G, López LC and Acuña Castroviejo D: Disruption of the
NF-κB/NLRP3 connection by melatonin requires retinoid-related
orphan receptor-α and blocks the septic response in rats. FASEB J.
29:3863–3875. 2015. View Article : Google Scholar
|
|
19
|
Rahim I, Djerdjouri B, Sayed RK,
Fernández-Ortiz M, Fernández-Gil B, Hidalgo-Gutiérrez A, López LC,
Escames G, Reiter RJ and Acuña-Castroviejo D: Melatonin
administration to wild-type rats and nontreated NLRP3 mutant rats
share similar inhibition of the inflammatory response during
sepsis. J Pineal Res. 63:2017. View Article : Google Scholar
|
|
20
|
Patiutko IuI, Khubiev AI, Poliakov MI,
Kirsanov MI and Sorokin AI: Functional results of gastrectomy in
cancer in relation to the surgical method and creation of
esophago-intestinal anastomosis. Vestn Khir Im I I Grek. 143:23–26.
1989.In Russian. PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Schrier RW and Wang W: Acute renal failure
and sepsis. N Engl J Med. 351:159–169. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Melnikov VY, Ecder T, Fantuzzi G, Siegmund
B, Lucia MS, Dinarello CA, Schrier RW and Edelstein CL: Impaired
IL-18 processing protects caspase-1-deficient rats from ischemic
acute renal failure. J Clin Invest. 107:1145–1152. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jimenez MF, Watson RW, Parodo J, Evans D,
Foster D, Steinberg M, Rotstein OD and Marshall JC: Dysregulated
expression of neutrophil apoptosis in the systemic inflammatory
response syndrome. Arch Surg. 132:1263–1269. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mangan DF, Mergenhagen SE and Wahl SM:
Apoptosis in human monocytes: Possible role in chronic inflammatory
diseases. J Periodontol. 64:461–466. 1993.PubMed/NCBI
|
|
26
|
Schroder K and Tschopp J: The
inflammasomes. Cell. 140:821–832. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bergsbaken T, Fink SL and Cookson BT:
Pyroptosis: Host cell death and inflammation. Nat Rev Microbiol.
7:99–109. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou R, Tardivel A, Thorens B, Choi I and
Tschopp J: Thioredoxin-interacting protein links oxidative stress
to inflammasome activation. Nat Immunol. 11:136–140. 2010.
View Article : Google Scholar
|
|
29
|
Gringhuis SI, Kaptein TM, Wevers BA,
Theelen B, van der Vlist M, Boekhout T and Geijtenbeek TB: Dectin-1
is an extracellular pathogen sensor for the induction and
processing of IL-1β via a noncanonical caspase-8 inflammasome. Nat
Immunol. 13:246–254. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tamura H, Nakamura Y, Korkmaz A,
Manchester LC, Tan DX, Sugino N and Reiter RJ: Melatonin and the
ovary: Physiological and pathophysiological implications. Fertil
Steril. 92:328–343. 2009. View Article : Google Scholar
|
|
31
|
Reiter RJ, Rosales-Corral SA, Manchester
LC and Tan DX: Peripheral reproductive organ health and melatonin:
Ready for prime time. Int J Mol Sci. 14:7231–7272. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Siah KT, Wong RK and Ho KY: Melatonin for
the treatment of irritable bowel syndrome. World J Gastroenterol.
20:2492–2498. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schwertner A, Conceição Dos Santos CC,
Costa GD, Deitos A, de Souza A, de Souza IC, Torres IL, da Cunha
Filho JS and Caumo W: Efficacy of melatonin in the treatment of
endometriosis: A phase II, randomized, double-blind,
placebo-controlled trial. Pain. 154:874–881. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
de Matos Cavalcante AG, de Bruin PF, de
Bruin VM, Nunes DM, Pereira ED, Cavalcante MM and Andrade GM:
Melatonin reduces lung oxidative stress in patients with chronic
obstructive pulmonary disease: A randomized, double-blind,
placebo-controlled study. J Pineal Res. 53:238–244. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bai XZ, He T, Gao JX, Liu Y, Liu JQ, Han
SC, Li Y, Shi JH, Han JT, Tao K, et al: Melatonin prevents acute
kidney injury in severely burned rats via the activation of SIRT1.
Sci Rep. 6:321992016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yildirim ME, Badem H, Cakmak M, Yilmaz H,
Kosem B, Karatas OF, Bayrak R and Cimentepe E: Melatonin protects
kidney against apoptosis induced by acute unilateral ureteral
obstruction in rats. Cent European J Urol. 69:225–230.
2016.PubMed/NCBI
|
|
37
|
Chang YC, Hsu SY, Yang CC, Sung PH, Chen
YL, Huang TH, Kao GS, Chen SY, Chen KH, Chiang HJ, et al: Enhanced
protection against renal ischemia-reperfusion injury with combined
melatonin and exendin-4 in a rodent model. Exp Biol Med (Maywood).
241:1588–1602. 2016. View Article : Google Scholar
|
|
38
|
Zhu F, Chong Lee, Shin OL, Xu H, Zhao Z,
Pei G, Hu Z, Yang J, Guo Y, Mou J, Sun J, et al: Melatonin promoted
renal regeneration in folic acid-induced acute kidney injury via
inhibiting nucleocytoplasmic translocation of HMGB1 in tubular
epithelial cells. Am J Transl Res. 9:1694–1707. 2017.PubMed/NCBI
|
|
39
|
Greene JC, Whitworth AJ, Kuo I, Andrews
LA, Feany MB and Pallanck LJ: Mitochondrial pathology and apoptotic
muscle degeneration in Drosophila parkin mutants. Proc Natl Acad
Sci USA. 100:4078–4083. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Narendra D, Tanaka A, Suen DF and Youle
RJ: Parkin is recruited selectively to impaired mitochondria and
promotes their autophagy. J Cell Biol. 183:795–803. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang X, Winter D, Ashrafi G, Schlehe J,
Wong YL, Selkoe D, Rice S, Steen J, LaVoie MJ and Schwarz TL: PINK1
and Parkin target Miro for phosphorylation and degradation to
arrest mitochondrial motility. Cell. 147:893–906. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yan Y, Jiang W, Liu L, Wang X, Ding C,
Tian Z and Zhou R: Dopamine controls systemic inflammation through
inhibition of NLRP3 inflammasome. Cell. 160:62–73. 2015. View Article : Google Scholar : PubMed/NCBI
|