Introduction
Bladder cancer (BC) is the fourth most common
malignancy in economically developed nations (1). In Japan, there were 16,477 new cases
in 2005 and 6,625 deaths in 2009 (2). There have been significant advances
in treatment, including surgical techniques and adjuvant
chemotherapy. However, BC frequently recurs and a poor clinical
outcome is anticipated when the cancer progresses to
muscle-invasive disease (3). It is
crucial to investigate the mechanism of carcinogenesis and novel
molecular target genes that have a tumor suppressive or oncogenic
function in BC.
MicroRNAs (miRNAs) constitute a class of regulatory
RNA with a length of 20–24 nucleotides, and they
post-transcriptionally regulate gene expression in eukaryotes.
Although their precise biology is not fully understood, miRNAs are
found in diverse organisms and epigenetically function as negative
regulators of gene expression. They are associated with cell
growth, cell cycle control, invasion, proliferation, migration, and
evasion of apoptosis in oncogenesis (4). They also play an important role in
oncogenesis in BC and behave in an oncogenic or tumor suppressive
manner in various cancers (5–7). To
date, many reports have demonstrated significant roles of miRNAs in
BC (8–13). We previously demonstrated that
several down-regulated miRNAs, such as miR-1, miR-133a, miR-145,
miR-218 and miR-517a, have tumor suppressive function by
targeting oncogenes. It is noteworthy that ‘actin binding protein’
was often included in the functional annotation of these miRNAs
target genes, such as fascin homologue 1 (FSCN1),
LIM and SH3 protein 1 (LASP1), and
transgelin-2 (TAGLN2) (14–16),
and restoration of these miRNAs induced cell apoptosis (16–18).
In this study, we focused on miR-574-3p,
which was selected from the 17 down-regulated miRNAs of BC in our
previous study (16).
MiR-574-3p as well as miR-218, a tumor suppressive
miRNA (17), are located on
chromosome 4p (4p14 and 4p15), which was a typical chromosomal loss
region in BC cell lines in our previous study (18). We found that
miR-574-3p was indeed down-regulated in BC cell
lines. To find the target genes of miR-574-3p, we
performed an oligo-microarray study of miR-574-3p
transfectants. We found that MESDC1 was the most
down-regulated gene and has a putative target site for
miR-574-3p. MESDC1 is predicted to be a novel
actin-binding protein located on chromosome 15q13. Although the
gene is conserved among many species, functional studies of MESDC1
are lacking in the literature. We hypothesized that
miR-574-3p directly regulates MESDC1 and that
this gene has oncogenic activity through its anti-apoptotic
function in BC. We performed a luciferase reporter assay to
determine whether MESDC1 mRNA was actually targeted by
miR-574-3p and loss-of-function studies using BC cell lines
to investigate functional roles of MESDC1 in BC.
Materials and methods
BC cell lines and cell culture
We used two human BC cell lines: BOY, which was
established in our laboratory from an Asian male patient, age 66,
and diagnosed with stage III BC with lung metastasis (20); T24 was obtained from the American
Type Culture Collection. These cell lines were maintained in
minimum essential medium (MEM) supplemented with 10% fetal bovine
serum (FBS) in a humidified atmosphere of 5% CO2 and 95%
air at 37°C.
Tissue samples
Tissue samples were taken from 24 BC patients who
had undergone cystectomy or transurethral resection of BCs at
Kagoshima University Hospital between 2007 and 2009. The median age
of the patients was 71 years, ranging from 62 to 88 years. The BC
samples were from 14 non-muscle invasive (<T2) and 10 muscle
invasive (≥T2) cancers; 10 were low grade BC and the other 14 were
high grade BC. The samples were staged in accordance with the
tumor-node-metastasis classification system of the American Joint
Committee on Cancer-Union Internationale Contre le Cancer (UICC)
and were histologically graded (21). The study was approved by the
Bioethics Committee of Kagoshima University; written prior informed
consent and approval were given by the patients.
Tissue collection and RNA extraction
Tissue samples were immersed in RNAlater (Qiagen,
Valencia, CA, USA) and stored at −20°C until RNA was extracted.
Total RNA (including miRNA) was extracted from frozen fresh tissues
using the mirVana™ miRNA isolation kit (Ambion, Austin, TX, USA) in
accordance with the manufacturer’s protocol. The integrity of the
RNA was checked with an RNA 6000 Nano Assay Kit and a 2100
Bioanalyzer™ (Agilent Technologies, Santa Clara, CA, USA).
Quantitative real-time RT-PCR
TaqMan probes and primers for MESDC1
(TaqMan® Gene Expression Assays, P/N: Hs00739656_s1,
Applied Biosystems, Foster City, CA, USA) were assay-on-demand gene
expression products. All reactions were performed in duplicate, and
a negative control lacking cDNA was included. We followed the
manufacturer’s protocol for the PCR conditions. Stem-loop RT–PCR
for miR-574-3p (TaqMan® MicroRNA Assays,
P/N: 002349, Applied Biosystems) was used to quantitate miRNAs
according to the earlier published conditions (10). cDNA was made from 5 ng of total RNA
from each sample using the TaqMan® MicroRNA Reverse
Transcription Kit (Applied Biosystems). For quantitative analysis
of mRNA and miRNA, we used human 18s rRNA (P/N: 4319413E,
Applied Biosystems) and RNU48 (P/N: 001006, Applied
Biosystems) as an internal control, and we used the delta-delta Ct
method to calculate the fold-change. As control RNA, we used three
different lots of Premium Total RNA from normal human bladder
(AM7990, Applied Biosystems).
Mature miRNA and siRNA transfection
As described elsewhere (10), the BC cell lines were transfected
with Lipofectamine™ RNAiMAX transfection reagent (Invitrogen,
Carlsbad, CA, USA) and Opti-MEM™ (Invitrogen) with 10 nM of mature
miRNA molecules. Mature miRNA molecules, Pre-miR™
(hsa-miR-574-3p, P/N: AM17100, Applied Biosystems) and
negative control miRNA (P/N: AM17111, Applied Biosystems) were used
in the gain-of-function experiments, whereas MESDC1 siRNA
(Cat# HSS126949 and HSS126950, Invitrogen) and negative control
siRNA (D-001810-10, Thermo Fisher Scientific, Waltham, MA, USA)
were used in the loss-of-function experiments. Cells were seeded in
10-cm dishes for protein extraction (8×105 per dish), in
6-well plates for apoptosis assays (10×104 per well) and
for wound healing assays (20×104 per well), in 24-well
plates for mRNA extraction and luciferase reporter assays
(5×104 per well), and in 96-well plates for XTT assays
(3,000 per well).
Cell viability, migration, and invasion
assays
Cell viability was determined by using an XTT assay
(Roche Applied Sciences, Tokyo, Japan) performed according to the
manufacturer’s instructions. Cell migration activity was evaluated
by wound-healing assays. Cells were plated in 6-well dishes, and
the cell monolayer was scraped using a P-20 micropipette tip. The
initial gap length (0 h) and the residual gap length 24 h after
wounding were calculated from photomicrographs. A cell invasion
assay was carried out using modified Boyden Chambers consisting of
transwell-precoated matrigel membrane filter inserts with 8 μm
pores in 24-well tissue culture plates (BD Biosciences, Bedford,
MA, USA). MEM containing 10% FBS in the lower chamber served as the
chemo-attractant, as described previously (14). All experiments were performed in
triplicate.
Apoptosis analysis
BC cell lines were transiently transfected with
transfection reagent only (mock), miR-control,
miR-574-3p, si-control, or si-MESDC1 in 6-well tissue
culture plates, as described earlier. Cells were harvested 72 h
after transfection by trypsinization and washed in cold PBS. Double
staining with FITC-Annexin V and propidium iodine (PI) was carried
out using the FITC Annexin V Apoptosis Detection Kit (BD
Biosciences) according to the manufacturer’s recommendations and
analyzed within 1 h by flow cytometry (FACScan®, BD
Biosciences). Cells were discriminated into viable cells, dead
cells, early apoptotic cells and apoptotic cells by CellQuest
software (BD Biosciences), and then the percentages of early
apoptotic cells from each experiment were compared. Experiments
were done in triplicate.
Target gene search for miR-574-3p
Oligo-microarray human 44K (Agilent) was used for
expression profiling of miR-574-3p-transfected BC
cell lines (BOY and T24) in comparison with miR-negative control
transfectants, as previously described (14). Briefly, hybridization and washing
steps were performed in accordance with the manufacturer’s
instructions. The arrays were scanned using a Packard GSI Lumonics
ScanArray® 4000 (Perkin-Elmer, Boston, MA, USA). The
data obtained were analyzed with DNASIS® array software
(Hitachi Software Engineering, Tokyo, Japan), which converted the
signal intensity for each spot into text format. The log2 ratios of
the median subtracted background intensity were analyzed. Data from
each microarray study were normalized by global normalization.
The predicted target genes and their miRNA binding
site seed regions were investigated using TargetScan program
(release 5.2, http://www.targetscan.org/), MicroRNA.org (released
August 2010, http://www.microrna.org/) and Micro
Cosm Targets (version 5, http://www.ebi.ac.uk/enright-srv/microcosm/htdocs/targets/v5/).
The sequences of the predicted mature miRNAs were confirmed using
miRBase (released 16.0, Sept 2010; http://microrna.sanger.ac.uk/).
Plasmid construction and dual-luciferase
reporter assay
MiRNA target sequences were inserted between the
SgfI-PmeI restriction sites in the 3′UTR of the hRluc gene in the
psiCHECK™-2 vector (C8021, Promega, Madison, WI, USA). Primer
sequences for full-length 3′UTR of MESDC1 mRNA
(TACGCGATCGCGTCTTCGCCCAGGACTTTAC and
TAGGTTTAAACAAACTTGACGTTGGGGGAAA) were designed. Following that, BOY
and T24 cells were transfected with 15 ng of vector, 10 nM of
miRNA, and 1 μl of Lipofectamine™ 2000 (Invitrogen) in 100 μl of
Opti-MEM™ (Invitrogen). The activities of firefly and
Renilla luciferases in cell lysates were determined with a
dual-luciferase assay system (E1910; Promega). Normalized data were
calculated as the quotient of Renilla/firefly luciferase
activities.
Statistical analysis
The relationship between two variables and the
numerical values obtained by real-time RT-PCR was analyzed using
the Mann-Whitney U test. The relationship between three variables
and the numerical values was analyzed using the Bonferroni-adjusted
Mann-Whitney U test. Expert StatView® analysis software
(version 4, SAS Institute Inc., Cary, NC, USA) was used in both
cases. In the comparison of three variables, a non-adjusted
statistical level of significance of P<0.05 corresponds to a
Bonferroni-adjusted level of P<0.0167.
Results
Detection of miR-574-3p expression in
clinical BCs and BC cell lines by quantitative stem-loop
RT-PCR
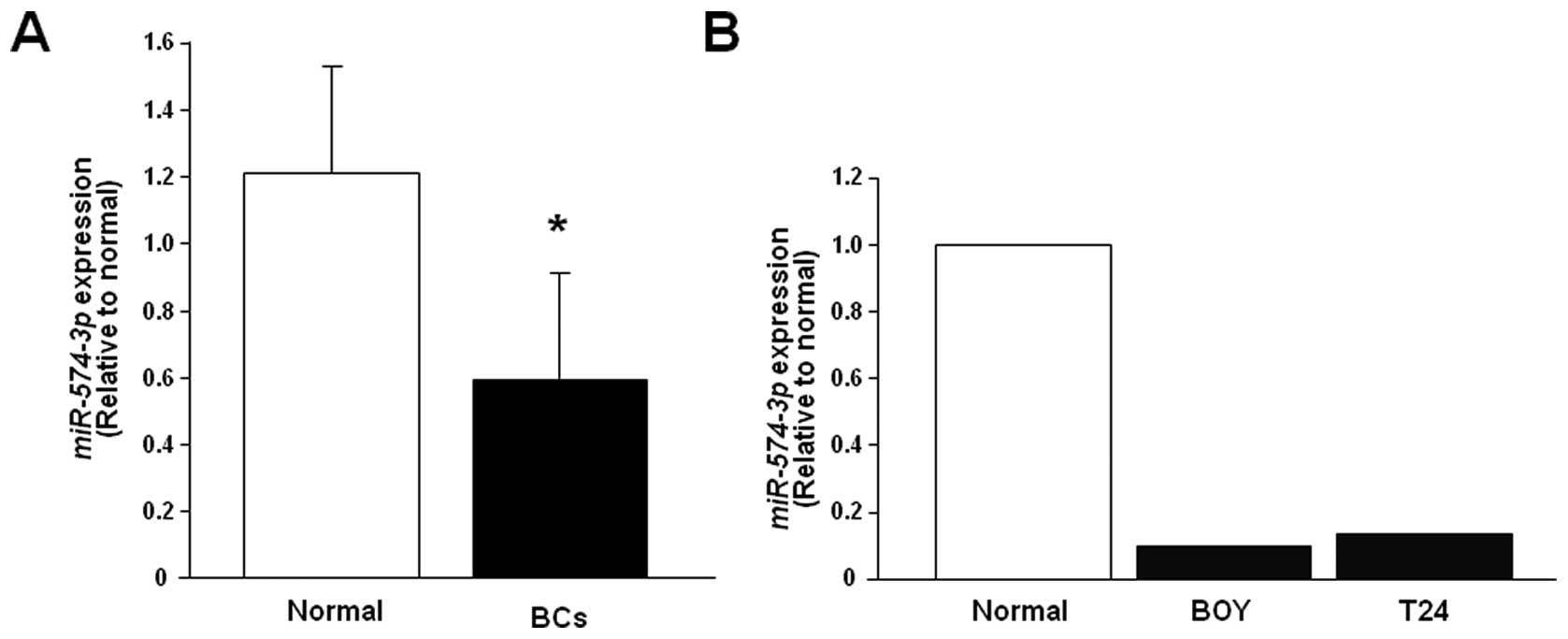
Quantitative stem-loop RT-PCR demonstrated that the
expression level of miR-574-3p was significantly lower in
BCs (n=24) than in the normal human RNAs (P=0.0128, Fig. 1A) and that it was markedly lower in
the BC cell lines (Fig. 1B). We
found no significant correlations between miR-574-3p
expressions and pathological parameters.
Effect of miR-574-3p transfection on cell
viability, migration, invasion activity, and cell apoptosis in BC
cell lines
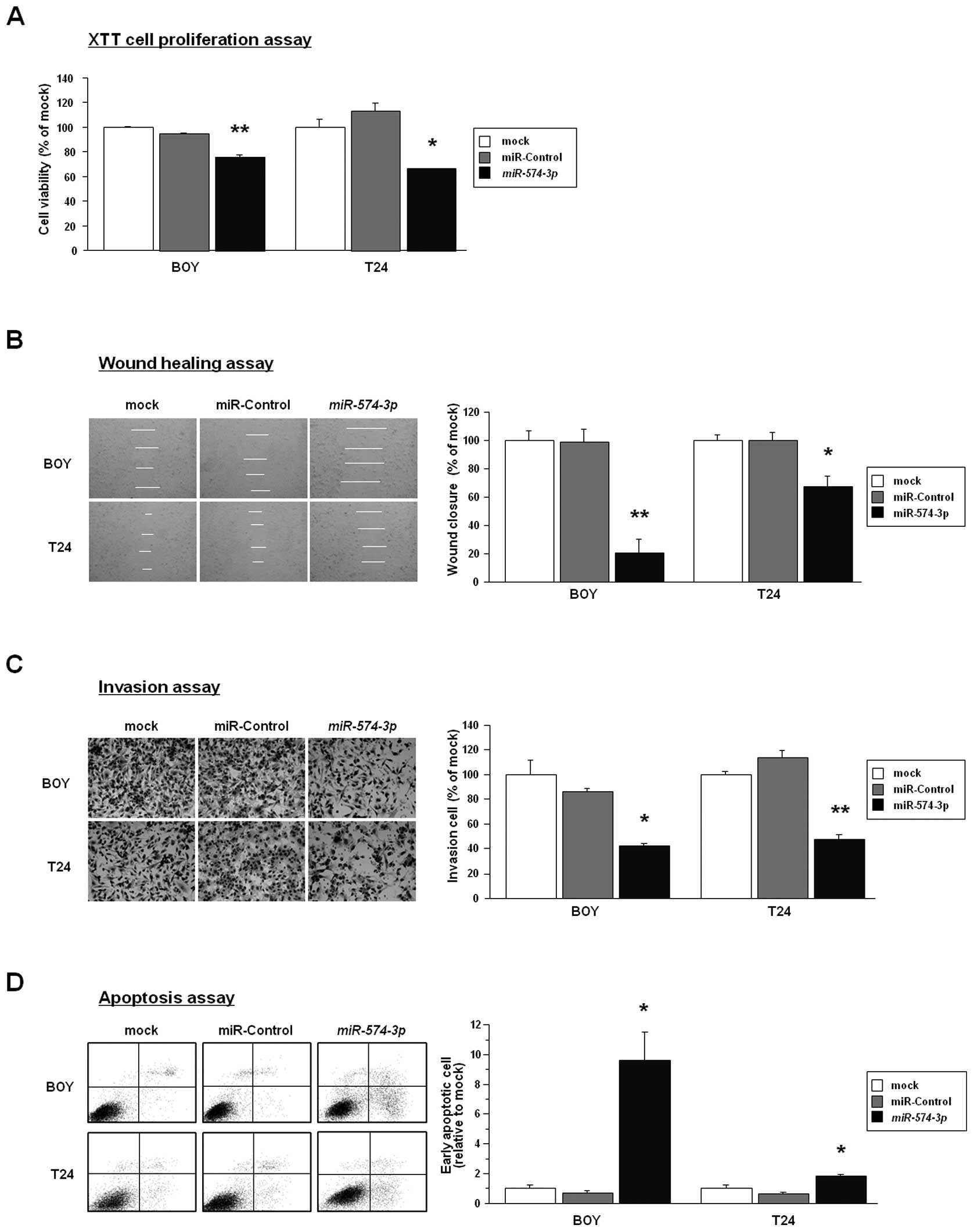
To investigate the functional role of
miR-574-3p, we performed gain-of-function
studies using miR-574-3p-transfected BC cell
lines (BOY and T24). The XTT assay demonstrated that cell
proliferation was reduced to 76% and 66% of mock in
miR-574-3p-transfected BOY and T24
(P<0.0001 and P<0.0005, Fig.
2A). The wound healing assay demonstrated that wound closure
ratio was reduced to 20% and 67% of mock in
miR-574-3p-transfected BOY and T24
(P<0.0001 and P=0.0004, Fig.
2B). The Matrigel invasion assay demonstrated that cell
invasion ratio was reduced to 42% and 48% of mock in
miR-574-3p-transfected BOY and T24 (each
P<0.0001, Fig. 2B). The early
apoptotic cell fractions (right lower quadrant) were greater in the
miR-574-3p transfectants than in the mocks and the
miR-control transfectants (relative to mock; BOY: 9.60±1.91,
1.00±0.25, and 0.71±0.15, respectively, P=0.0013; T24: 1.84±0.08,
1.00±0.22, and 0.65±0.11, respectively, P=0.0014) (Fig. 2D). These results suggested that
restoration of miR-574-3p expression might induce cell
apoptosis in BC.
Gene expression signatures of
differentially down-regulated genes in miR-574-3p
transfectants
To gain further insight into which genes were
affected by miR-574-3p transfection, we performed gene
expression analysis with miR-574-3p transfectants (BOY and
T24). A total of 11 genes were down-regulated less than −2.0-fold
in the miR-574-3p transfectants compared with the control
transfectants (Table I).
MicroRNA.org and MicroCosm Target programs showed the eight genes
had putative target sites for miR-574-3p in their 3′UTR
(Table I). The MESDC1 gene
was the most down-regulated gene that had conserved target sites
for miR-574-3p. Therefore, we focused on MESDC1 as a
promising candidate targeted by miR-574-3p. Entries from the
former and the current microarray data were approved by the Gene
Expression Omnibus (GEO) and were assigned GEO accession number,
GSE24782.
 | Table IDown-regulated genes in
miR-574-3p transfectants. |
Table I
Down-regulated genes in
miR-574-3p transfectants.
| Entrez gene ID | Fold change (log2
ratio) | Gene name | Target sites |
|---|
| Symbol | BOY | T24 | Average |
|---|
| 59274 | MESDC1 | −2.01 | −1.59 | −1.80 | Mesoderm
development candidate 1 | + |
| 10950 | BTG3 | −1.88 | −1.34 | −1.61 | BTG family,
member 3 | + |
| 27430 | MAT2B | −1.47 | −1.41 | −1.44 | Methionine
adenosyltransferase II, beta | + |
| 11160 | ERLIN2 | −1.61 | −1.15 | −1.38 | ER lipid raft
associated 2 | − |
| 2021 | ENDOG | −1.41 | −1.28 | −1.34 | Endonuclease
G | + |
| 340508 |
LOC340508 | −1.54 | −1.09 | −1.31 | Hypothetical
protein LOC340508 | − |
| 117178 | SSX2IP | −1.37 | −1.05 | −1.21 | Synovial
sarcoma, X breakpoint 2 interacting protein | + |
| 285761 | DCBLD1 | −1.40 | −1.00 | −1.20 | Discoidin, CUB
and LCCL domain containing 1 | + |
| 122416 | ANKRD9 | −1.19 | −1.16 | −1.17 | Ankyrin repeat
domain 9 | + |
| 3363 | HTR7 | −1.16 | −1.03 | −1.09 |
5-hydroxytryptamine (serotonin)
receptor 7 (adenylate cyclase-coupled) | + |
| 246329 | STAC3 | −1.01 | −1.05 | −1.03 | SH3 and cysteine
rich domain 3 | − |
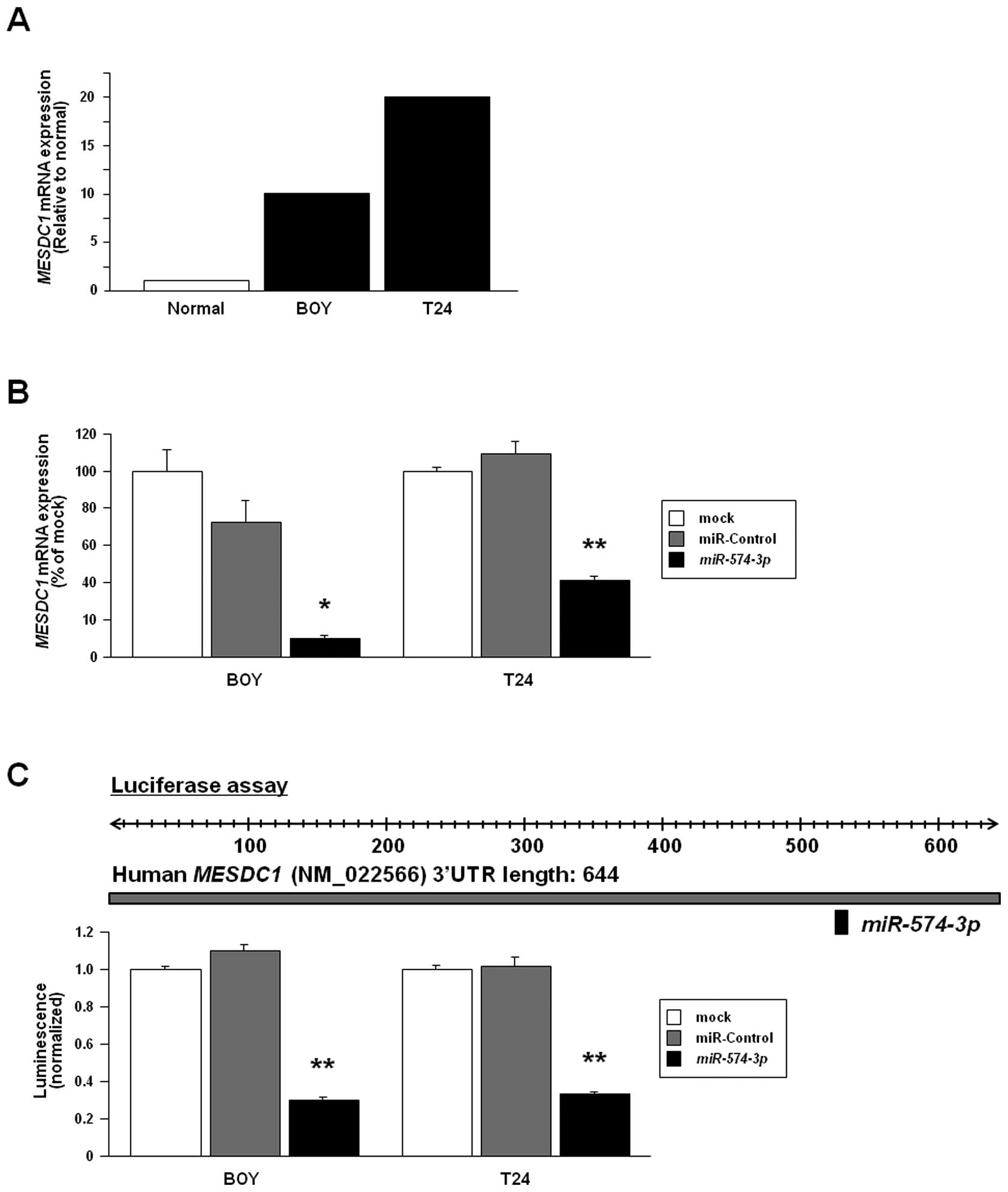
MESDC1 expression in BC cell lines and
MESDC1 silencing by miR-574-3p transfection
The quantitative real-time RT-PCR analysis showed
that mRNA expression levels of MESDC1 in BOY and T24 were
higher than that in the normal human bladder RNAs (Fig. 3A). In clinical BCs, there was no
significant difference in MESDC1 mRNA expression between
clinical BCs and the normal human bladder RNAs (data not shown). We
performed gain-of-function studies using
miR-574-3p-transfected BOY and T24. The mRNA expression
levels of MESDC1 were markedly repressed in the
miR-574-3p transfectants in comparison with the miR-control
transfectants and the mocks (Fig.
3B).
Confirmation of MESDC1 as a target of
post-transcriptional repression by miR-574-3p
We performed a luciferase reporter assay to
determine whether MESDC1 mRNA has an actual target site for
miR-574-3p. We used a vector encoding full-length 3′UTR of
MESDC1 mRNA and found that the luminescence intensity was
significantly reduced in the miR-574-3p transfectants
compared to their counterparts (Fig.
3C).
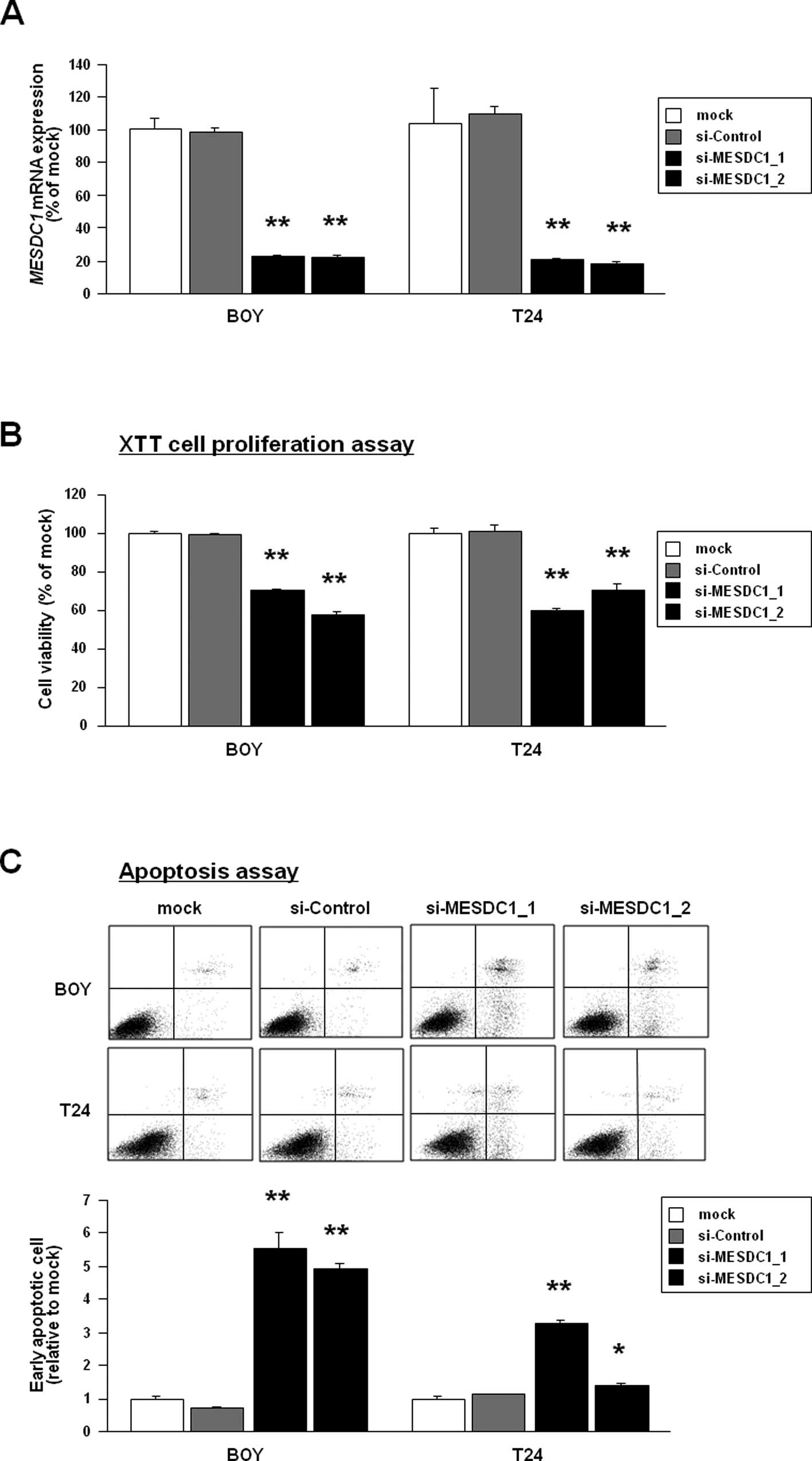
Effect of MESDC1 knockdown on cell
viability, migration, invasion activity, and cell apoptosis in BC
cell lines
To examine the functional role of MESDC1, we
performed loss-of-function studies using two different
si-MESDC1-transfected BC cell lines. The mRNA expression of
MESDC1 was markedly down-regulated by these si-MESDC1
transfections (Fig. 4A). The XTT
assay demonstrated that cell proliferation was reduced to 71% and
57% of control by both siRNAs-treatment in BOY cells (P<0.0001)
and that it was 60% and 70% of control (P<0.0001) in the T24
line (Fig. 4B). The early
apoptotic cell fractions (right lower quadrant) were greater in the
two si-MESDC1 transfectants than in the mocks and the si-control
transfectants 72 h after transfection (relative to mock; BOY:
5.53±0.48, 4.93±0.15, 1.00±0.05, and 0.73±0.01, respectively,
P<0.0001; T24: 3.30±0.08, 1.37±0.08, 1.00±0.05, and 1.11±0.03,
P<0.0001 and P=0.0024) (Fig.
4C). These results suggested that knockdown of MESDC1
expression might induce cell apoptosis in BC. The wound healing
assay also demonstrated that wound closure ratio was reduced to 21%
and 33% of mock by both siRNAs-treatment in BOY cells (each
P<0.0001) and that it was 21% and 62% of control in the T24
cells (P<0.0001 and P=0.0007) (Fig.
4D). The matrigel invasion assay demonstrated that cell
invasion ratio was reduced to 4% and 22% of mock by both
siRNAs-treatment in BOY cells (each P<0.0001) and that it was
33% and 64% of control in the T24 cells (each P<0.0001)
(Fig. 4E).
Discussion
Since Gottardo et al reported in 2007
(8), several miRNA expression
signatures of BC have been published (8–13).
Several microRNAs signatures were found to have oncogenic or tumor
suppressive function. However, it is difficult for investigators to
choose the important miRNAs to be examined. We previously
investigated chromosomal gain or loss regions in BC by using
comparative genomic hybridization (CGH) arrays (18) and found a typical loss region on
chromosome 4p which harbors tumor suppressive miR-218
(17). In this study, we focused
on miR-574-3p for two reasons: it was the
seventh down-regulated miRNA in our miRNA signature specific to BC
(16), and it was located on
chromosome 4p. The expression levels of
miR-574-3p were low in BC cell lines, and we
found that it might have tumor suppressive function. Other tumor
suppressive miRNAs in our previous study included MiR-1,
miR-133a, and miR-145. Those miRNAs are
located on chromosome 18q11.2 (miR-1 and miR-133a)
and 5q32 (miR-145) where typical chromosomal loss
regions were identified in our previous CGH-array study (17). Thus, chromosomal loss regions might
be one of the critical mechanisms regulating miRNA expression in
tumor suppression.
In previous studies, serum miR-574-3p
expression was found to be up-regulated in patients with
hepatocellular carcinoma and liver cirrhosis (22), and tissue miR-574-3p
expression was up-regulated in endometriomas (23). However, tissue
miR-574-3p expression in various cancers had yet to
be measured and previous studies did not address the functional
role of miR-574-3p. In this study, we demonstrated that
miR-574-3p significantly inhibited cell viability,
migration, and invasion through its induction of cell apoptosis. In
this study, there was no significant relationship between
miR-574-3p expression and pathological parameters,
possibly because our patient cohort was too small to evaluate the
relationship between them. A future in vivo study would be
helpful in clarifying the role of miR-574-3p in cancer
development.
Our previous study demonstrated that tumor
suppressive miRNAs, such as miR-1, miR-133a, and
miR-145 functioned through repression of LIM and
SH3 protein 1 (LASP1), transgelin-2
(TAGLN2), and Switch associated protein 70
(SWAP70) (15,16,24).
These genes are commonly classified as actin-binding proteins.
Actin-based structures are involved in cortical cell protrusions
that mediate interactions between cells and the extracellular
matrix (ECM), cell-to-cell interactions, cell migration, and
cytoplasmic microfilamentous bundles that contribute to cell
architecture and intracellular movement (25). It is plausible that the activation
of these genes through ECM substrates contributes to tumor growth,
migration, and invasion.
In this study, we focused on MESDC1, a
potential target gene of miR-574-3p, because it was listed
at the top of down-regulated genes in miRNA transfectants.
MESDC1 was actually up-regulated in BC cell lines. We
demonstrated that miR-574-3p bound to the conserved site in
the 3′ untranslated region of MESDC1 mRNA in luciferase
assays. We found several reports concerning MESDC1, but its
functional role had not been investigated (26,27).
In our functional studies using si-MESDC1 in BC cell lines, we
demonstrated that cell viability, migration and invasion were all
inhibited and apoptosis was induced. These results suggest that
MESDC1 protein has an oncogenic function in BC and is regulated by
miR-574-3p expression. In this study, there was no
significant difference in miR-574-3p expression levels
between clinical BCs and RNA obtained from the normal human
bladders. Our cohort was too small to evaluate the difference
between them. We demonstrated that oncogenic function was mediated
by the MESDC1 gene as well as other actin-binding proteins
that we investigated previously. Our series of functional analyses
of genes targeted by tumor suppressive miRNAs revealed that these
actin-binding proteins might have anti-apoptotic effects in several
cancer types (16,17,19,28–31).
Further studies are necessary to elucidate the precise mechanisms
by which apoptosis is induced by tumor suppressive miRNAs and
knockdown of their target genes.
In conclusion, our results demonstrated that
miR-574-3p was down-regulated in BC cell lines. We found
decreased cell proliferation, migration, and invasive activities,
and increased cell apoptosis in miR-574-3p transfectants,
suggesting that miR-574-3p is a candidate tumor suppressive
miRNA in human BC. Further, we found that miR-574-3p
targeting of MESDC1 plays an important role in cell viability,
migration, invasion, and apoptosis in BC cell lines.
Acknowledgements
This research was partially supported by the
Ministry of Education, Science, Sports and Culture Grants-in-Aid
for Scientific Research (C), 23501298 and 23592340, 2011. We thank
Ms. Mutsumi Miyazaki for her excellent laboratory assistance.
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar
|
|
2
|
Website of Center for Cancer Control and
Information Services. National Cancer Center; Japan: (http://ganjoho.jp/professional/statistics/statistics.html).
|
|
3
|
Herr H, Konety B, Stein J, et al:
Optimizing outcomes at every stage of bladder cancer: do we
practice it? Urol Oncol. 27:72–74. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zimmerman AL and Wu S: MicroRNAs, cancer
and cancer stem cells. Cancer Lett. 300:10–19. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fendler A, Stephan C, Yousef GM and Jung
K: MicroRNAs as regulators of signal transduction in urological
tumors. Clin Chem. 57:954–968. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar
|
|
8
|
Gottardo F, Liu CG, Ferracin M, et al:
Micro-RNA profiling in kidney and bladder cancers. Urol Oncol.
25:387–392. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang H, Dinney CP, Ye Y, et al: Evaluation
of genetic variants in microRNA-related genes and high risk of
bladder cancer. Cancer Res. 68:2530–2537. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ichimi T, Enokida H, Okuno Y, et al:
Identification of novel microRNA targets based on microRNA
signatures in bladder cancer. Int J Cancer. 125:345–352. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Catto JW, Miah S, Owen HC, et al: Distinct
microRNA alterations characterize high- and low-grade bladder
cancer. Cancer Res. 69:8472–8481. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Catto JW, Alcaraz A, Bjartell AS, et al:
MicroRNA in prostate, bladder, and kidney cancer: a systematic
review. Eur Urol. 59:671–681. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Han Y, Chen J, Zhao X, et al: MicroRNA
expression signature of bladder cancer revealed by deep sequencing.
PLoS One. 6:e182862011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chiyomaru T, Enokida H, Tatarano S, et al:
miR-145 and miR-133a function as tumor suppressors and directly
regulate FSCN1 expression in bladder cancer. Br J Cancer.
102:883–891. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chiyomaru T, Enokida H, Kawakami K, et al:
Functional role of LASP1 in cell viability and its regulation by
microRNAs in bladder cancer. Urol Oncol. 2010.[Epub ahead of
print].
|
|
16
|
Yoshino H, Chiyomaru T, Enokida H, et al:
The tumour-suppressive function of miR-1 and miR-133a targeting
TAGLN2 in bladder cancer. Br J Cancer. 104:808–818. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tatarano S, Chiyomaru T, Kawakami K, et
al: Mir-218 on the genomic loss region of chromosome 4p15.31
function as a tumor suppressor in bladder cancer. Int J Oncol.
39:13–21. 2011.PubMed/NCBI
|
|
18
|
Matsuda R, Enokida H, Chiyomaru T, et al:
LY6K is a novel molecular target in bladder cancer on basis of
integrate genome-wide profiling. Br J Cancer. 104:376–386. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yoshitomi T, Kawakami K, Enokida H, et al:
Restoration of miR-517a expression induces cell apoptosis in
bladder cancer cell lines. Oncol Rep. 24:1661–1668. 2011.PubMed/NCBI
|
|
20
|
Takemoto M, Shirahama T, Miyauchi T, et
al: Metanestin, a glycoprotein with metastasis-associated
expression in transitional cell carcinoma of the urinary bladder.
Int J Cancer. 74:7–14. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sobin LH and Wittekind C: TNM
Classification of Malignant Tumors International Union Against
Cancer (UICC). 6th edition. Wiley-Liss Publications; New York, NY:
pp. 199–202. 2002
|
|
22
|
Gui J, Tian Y, Wen X, et al: Serum
microRNA characterization identifies miR-885-5p as a potential
marker for detecting liver pathologies. Clin Sci. 120:183–193.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hawkins SM, Creighton CJ, Han DY, et al:
Functional microRNA involved in endometriosis. Mol Endocrinol.
25:821–832. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chiyomaru T, Tatarano S, Kawakami K, et
al: SWAP70, actin-binding protein, function as an oncogene
targeting tumor-suppressive miR-145 in prostate cancer. Prostate.
View Article : Google Scholar : 2011.PubMed/NCBI
|
|
25
|
Kureishy N, Sapountzi V, Prag S, Anilkumar
N and Adams JC: Fascins, and their roles in cell structure and
function. Bioessays. 24:350–361. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wines ME, Lee L, Katari MS, et al:
Identification of mesoderm development (mesd) candidate genes by
comparative mapping and genome sequence analysis. Genomics.
72:88–98. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gingras AR, Bate N, Goult BT, et al:
Central region of talin has a unique fold that binds vinculin and
actin. J Biol Chem. 285:29577–29587. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Uchida Y, Chiyomaru T, Enokida H, et al:
MiR-133a induces apoptosis through direct regulation of GSTP1 in
bladder cancer cell lines. Urol Oncol. 2011.[Epub ahead of
print].
|
|
29
|
Nohata N, Hanazawa T, Kikkawa N, et al:
Tumor suppressive microRNA-375 regulates oncogene AEG-1/MTDH in
head and neck squamous cell carcinoma (HNSCC). J Hum Genet.
56:595–601. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kawakami K, Enokida H, Chiyomaru T, et al:
The functional significance of miR-1 and miR-133a in renal cell
carcinoma. Eur J Cancer. 2011.[Epub ahead of print].
|
|
31
|
Nohata N, Hanazawa T, Kikkawa N, et al:
Identification of novel molecular targets regulated by tumor
suppressive miR-1/miR-133a in maxillary sinus squamous cell
carcinoma. Int J Oncol. 39:1099–1107. 2011.PubMed/NCBI
|