Introduction
Clinical cancer therapy includes the use of most
common treatments such as surgery, radiation and chemotherapy for
cancer patients. Chemotherapeutic agents can regulate uncontrolled
proliferation of abnormal cancer cells and are often combined with
surgery and/or radiation therapy. The majority of chemotherapeutic
drugs can be classified as alkylating agents, antimetabolites,
anthracyclines, plant alkaloids, topoisomerase inhibitors,
monoclonal antibodies, and other antitumor agents (1–4).
Numerous studies are progressing to develop antitumor drugs such as
molecular-targeting drugs although few drug therapies lead to
complete recovery in cancer patients (5,6).
Therefore, the development of more effective chemotherapeutic drugs
is essential for the treatment of cancer.
Liquid crystal is the state of matter existing
between liquid and crystalline phase and is characterized by the
partial or complete loss of positional order of the constituent
molecules (7). Liquid crystal
compounds (LCCs) are widely used in display media such as in
television screens. In general, liquid crystals are synthesized
from various precursors such as a flexible chain, a rigid aromatic
core, and hydrophobic and hydrophilic units. In contrast, various
biological cell structures such as cell membranes consist of
amphiphilic phospholipids. Therefore, LCCs or their precursors,
namely liquid crystal-related compounds (LCRCs), possibly interact
with biological cell structures. In fact, there has been an
interest in the biological and pharmacological effects of these
compounds (8–10).
We previously demonstrated that LCCs suppressed the
cell growth of the non-small cell lung cancer (NSCLC) cell line
A549 through G1-phase arrest, although it did not induce cell death
(11). Recently, we also reported
that some LCRCs dramatically suppressed cell growth and induced
apoptosis in human chronic myelogenous leukemia K562 cells
(12). Although leukemic cells are
more sensitive to chemotherapy and ionizing radiation compared with
solid malignancies (13), there is
a possibility that these LCRCs potentially suppress cell growth and
induce cell death in solid malignancies including NSCLC. Therefore,
in the present study, we investigated the antitumor effect of LCRCs
in the NSCLC cell line A549. Furthermore, the radiosensitization
effect of LCRC was also examined because combined treatment with
radiotherapy and chemotherapy has been extensively used in the
management of many types of solid malignancies including NSCLC.
Materials and methods
Reagents
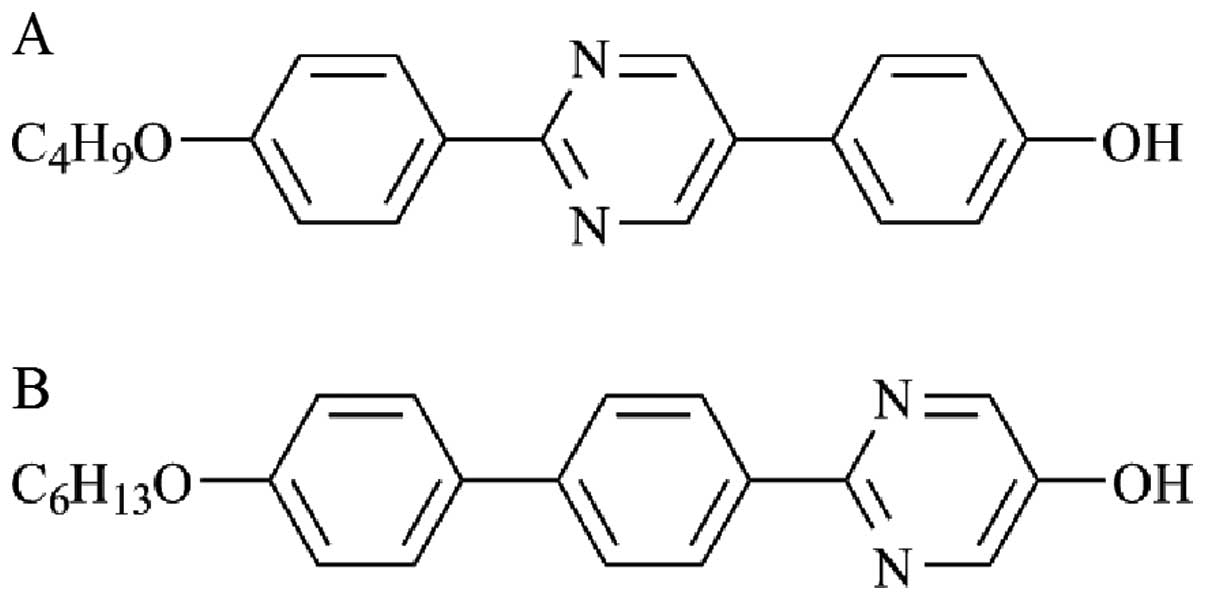
The structures of LCRCs used in the present study
are shown in Fig. 1. The compounds
2-(4-butoxyphenyl)-5-(4-hydroxyphenyl)pyrimidine (LCRC-1) and
2-{4-(4-hexyloxyphenyl) phenyl}-5-hydroxypyrimidine (LCRC-2) were
purchased from Midori Kagaku Co Ltd. (Tokyo, Japan). Each compound
was dissolved in pure dimethyl sulfoxide (DMSO) (Sigma-Aldrich, St.
Louis, MO, USA). A pan-caspase inhibitor,
benzyloxycarbonyl-Val-Ala-Asp fluoromethyl ketone (Z-VAD-FMK), was
purchased from Promega Co Ltd. (Madison, WI, USA). Propidium iodide
(PI) was purchased from Sigma-Aldrich.
Cell culture
Human lung cancer cell A549 and normal fibro-blasts
called human embryonic fibroblast, lung-derived cell line (WI-38)
were purchased from the RIKEN Bio-Resource Center (Tsukuba, Japan).
The A549 cells were maintained in Dulbecco’s modified Eagle’s
medium (DMEM, Sigma-Aldrich) supplemented with 10% heat-inactivated
fetal bovine serum (FBS, Japan Bioserum Co. Ltd., Japan) at 37°C in
a humidified atmosphere containing 5% CO2. The WI-38
cells were maintained in Minimum Essential Medium Eagle (MEM,
Sigma-Aldrich) supplemented with 10% heat-inactivated FBS at 37°C
in a humidified atmosphere containing 5% CO2.
Liquid culture
The A549 cells (6.0×104) were seeded onto
a 35-mm culture dish (Iwaki, Chiba, Japan) and cultured overnight
to allow adherence to the dish. Next day, each compound (12
μM) or vehicle (DMSO) was added in the culture medium and
cultured for 3 days. After 3-day culture, the cells were harvested
with 0.1% trypsin-EDTA (Gibco® Invitrogen, CA, USA) and
viable cells were counted by trypan blue exclusion assay. To
investigate the effect of LCRC-1 in detail, the A549 cells were
treated with 1.5–12 μM LCRC-1 for 72 h and then the viable
cells were counted as described above.
Cell cycle analysis by flow
cytometry
The A549 cells were seeded onto a 60-mm culture dish
(Iwaki) and incubated overnight to adhere to the dish. The cells
treated with the compound were harvested and fixed with 70% ethanol
overnight at 4°C. The fixed cells were washed with PBS(-) and then
treated with RNase (200 μg/ml) at 37°C for 30 min to
hydrolyze RNA. After treatment, the cells were washed with PBS(-)
and stained with PI (25 μg/ml) for 30 min in the dark. A
flow cytometer (Cytomics FC500, Beckman-Coulter, Fullerton, CA,
USA) was used to analyze the cell cycle distribution.
Detection of apoptosis
The extent of apoptosis was determined by Annexin
V-FITC (BioLegend, San Diego, CA, USA) and PI staining according to
the manufacturer’s instructions. Briefly, the cells treated with
compound were harvested, washed, and suspended in the binding
buffer (10 mM HEPES/NaOH, 140 mM NaCl, 2.5 mM CaCl2, pH
7.4). The Annexin V-FITC (2.5 μg/ml) and PI solution (50
μg/ml) were added in the cell suspension and incubated for
15 min at room temperature in the dark. Apoptosis cells were
determined by flow cytometry. In the Annexin V/PI quadrant gating,
Annexin V(−)/PI(−), Annexin V(+)/PI(−), and Annexin V(+)/PI(+) were
used to identify the fraction of viable cells, early apoptotic
cells, and late apoptotic/necrotic cells, respectively.
Detection of activated caspase-3
An FITC-conjugated monoclonal active caspase-3
antibody apoptosis kit (BD Biosciences, San Diego, CA, USA) was
used to detect active caspase-3 according to the manufacturer’s
instructions. Briefly, the cells treated with compound were
harvested and washed with PBS(-). The cells were suspended in
Cytofix/Cytoperm™ and incubated for 20 min on ice. After
incubation, the cells were washed with Perm/Wash™ buffer and
resuspended in Perm/Wash™ buffer containing 5% FITC-conjugated
anti-active caspase-3 antibody. After 30 min incubation at room
temperature in the dark, the cells were washed and then analyzed by
flow cytometry.
Inhibition of caspase by Z-VZD-FMK
The A549 cells were preincubated with 100 μM
Z-VAD-FMK, a pan-caspase inhibitor, for 1 h before adding to the
compound. After 48 h culturing in the presence of compound, the
cells were harvested and the viable cells were counted.
Furthermore, the analysis of apoptosis (Annexin V-FITC and PI
staining) and detection of active caspase-3 were performed as
described above.
Immunofluorescence detection of
γ-H2AX
The A549 cells (3.0×104) were grown on
chamber slides II (Iwaki) overnight to allow adherence to the
slides. After treatment with compound for the indicated periods,
the cells were fixed with cold methanol for 20 min and acetone for
7 sec, dried, treated with 0.5% Triton X-100 (Wako, Osaka, Japan)
for 10 min, and washed in PBS(-). The cells were then incubated
with anti-phospho-histone H2AX monoclonal antibody (JBW301, Upstate
Biotechnology, Lake Placid, NY, USA) at a 300-fold dilution with
TBST (20 mM Tris-HCl, pH 7.4, 137 mM NaCl, 0.1% Tween-20)
containing 5% skim milk for 60 min at room temperature. After the
cells were washed with PBS(-), they were then incubated with an
AlexaFluor 488® conjugated anti-mouse immunoglobulin G
second antibody (Molecular Probes, Eugene, OR, USA) at a 400-fold
dilution with TBST containing 5% skim milk for 60 min at room
temperature, and washed in PBS(-). The slides were stained and
mounted with Vectashield® Mounting Medium with DAPI
(Vector Laboratories Inc., Burlingame, CA, USA). Photographs of the
cells were taken with a Laser Scanning Microscope 710 (Carl Zeiss
Microscopy Co Ltd., Tokyo, Japan).
Clonogenic survival assay
The A549 cells were seeded onto a 60-mm culture dish
(Iwaki) and incubated overnight to allow adherence to the dish. The
compounds were added in culture medium 1 h before X-ray
irradiation. Aluminum (0.5 mm) and copper (0.3 mm) filters at a
distance of 45 cm from the focus were used to expose the cells to
X-rays (150 kVp, 20 mA) at a dose rate of 1.0 Gy/min (MBR-1520R-3
Hitachi Medical Co., Tokyo, Japan) in the range of 1–8 Gy. The
cells were cultured at 37°C in a humidified atmosphere containing
5% CO2. After 7-day culture, the cells were fixed in
methanol for 10 min at room temperature, air dried, and stained by
Gimsa stain solution. The colonies consisting of more than 50 cells
were counted by inversion microscopy.
Statistical analysis
The significance of differences between the control
and experimental groups was determined by two-sided Student’s
t-test and two-sided Mann-Whitney U test depending on the data
distribution. A one-way ANOVA model and the Tukey-Kramer test were
used to analyze the data from multiple groups (i.e., Fig. 4D and E). The significance level was
set to p<0.05. Excel 2010 software (Microsoft, USA) with the
add-in software Statcel 2 was used for statistical analysis
(14).
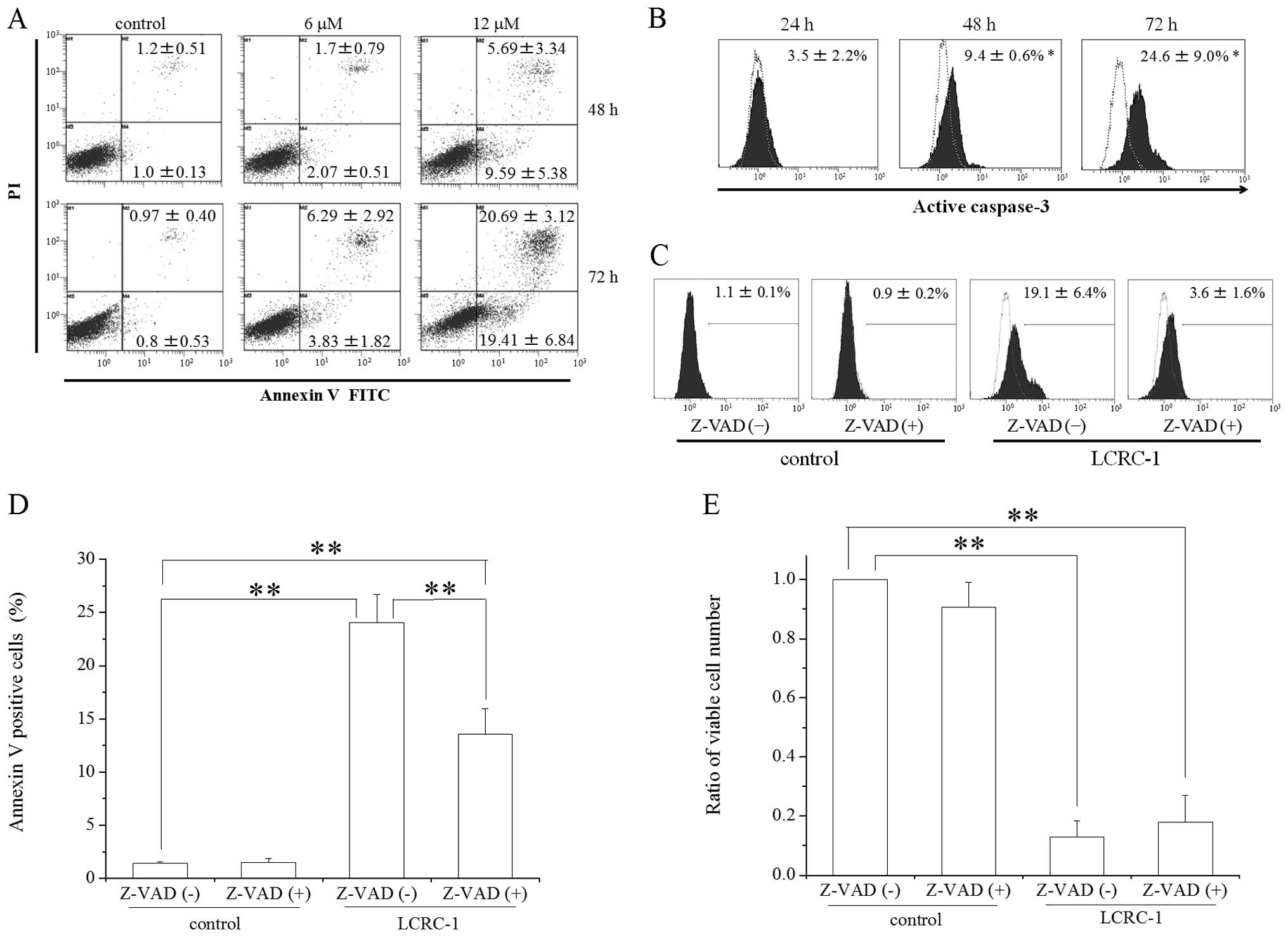 | Figure 4Effect of LCRC-1 on the induction of
apoptosis in the A549 cells. The A549 cells cultured in the
presence of 6 and 12 μM LCRC-1 for 48 and 72 h were
harvested, and the apoptotic cells were analyzed. (A) Annexin/PI
staining was performed by flow cytometry, as described in the
Materials and methods section. Representative histograms are shown,
and the inset numbers are the percentage of Annexin(+)/PI(-) and
Annexin(+)/PI(+) cells. The data are presented as the means ± SD of
four independent experiments. *p<0.05 and
**p<0.01 by two-sided Student’s t-test, respectively.
(B) The expression of active caspase-3 was analyzed by flow
cytometry, as described in the Materials and methods section.
Representative histograms are shown, and the inset numbers are the
percentage of active caspase-3 positive cells. The expression of
active caspase-3 in the A549 cells treated with vehicle is shown as
a dotted line. The data are presented as the mean ± SD of four
independent experiments. *p<0.05 and
**p<0.01 by two-sided Student’s t-test, respectively.
(C-E) The A549 cells preincubated with Z-VAD-FMK were cultured in
the presence of 12 μM LCRC-1 for 48 h. The cells were
harvested, followed by analysis of (C) active caspase-3, (D)
annexin/PI staining and determination of the (E) viable cell number
count. The data are presented as the mean ± SD of three independent
experiments. Z-VAD, Z-VAD-FMK; *p<0.05 by
Tukey-Kramer test. |
Results
LCRCs suppresses the growth of A549
cells
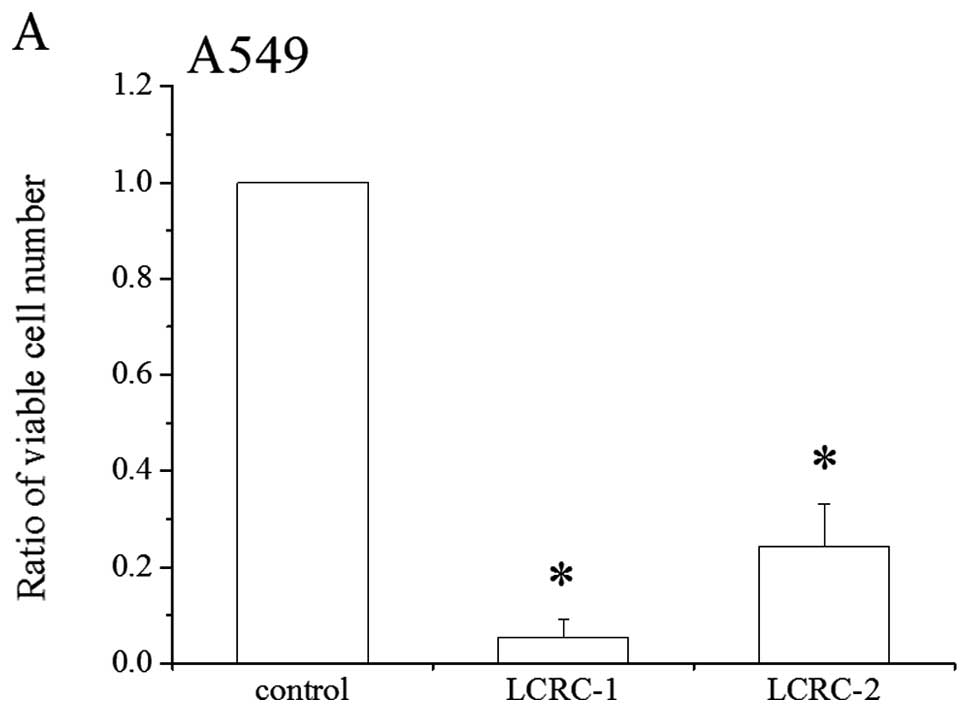
The suppressive effects of LCRCs, whose cell
suppressive effect had been previously demonstrated in the K562
cells, were investigated in the NSCLC cell line A549 and in WI-38
normal fibroblasts. As shown in Fig.
2A, both LCRCs suppressed the growth of the A549 cells. The
suppressive effects of LCRC-1 and 2 caused 95 and 78% inhibition,
respectively. The effects of these LCRCs on WI-38 proliferation
were also examined. Although LCRCs slightly suppressed WI-38
proliferation, there was no statistically significant difference in
suppression between the treatments with vehicle and LCRCs (Fig. 2B). These results indicate that the
suppressive effect in the A549 cells was more dramatic for LCRC-1
than that for LCRC-2, whereas the effect of either compound was
minimal to non-existent on growth of the WI-38 cells. Because the
50% inhibitory concentration (IC50) of LCRC-1 for 72 h
was 5.52±1.88 μM (data not shown), we selected 6 μM
(approximately IC50) and 12 μM (2 ×
IC50) LCRC-1 concentrations for following
experiments.
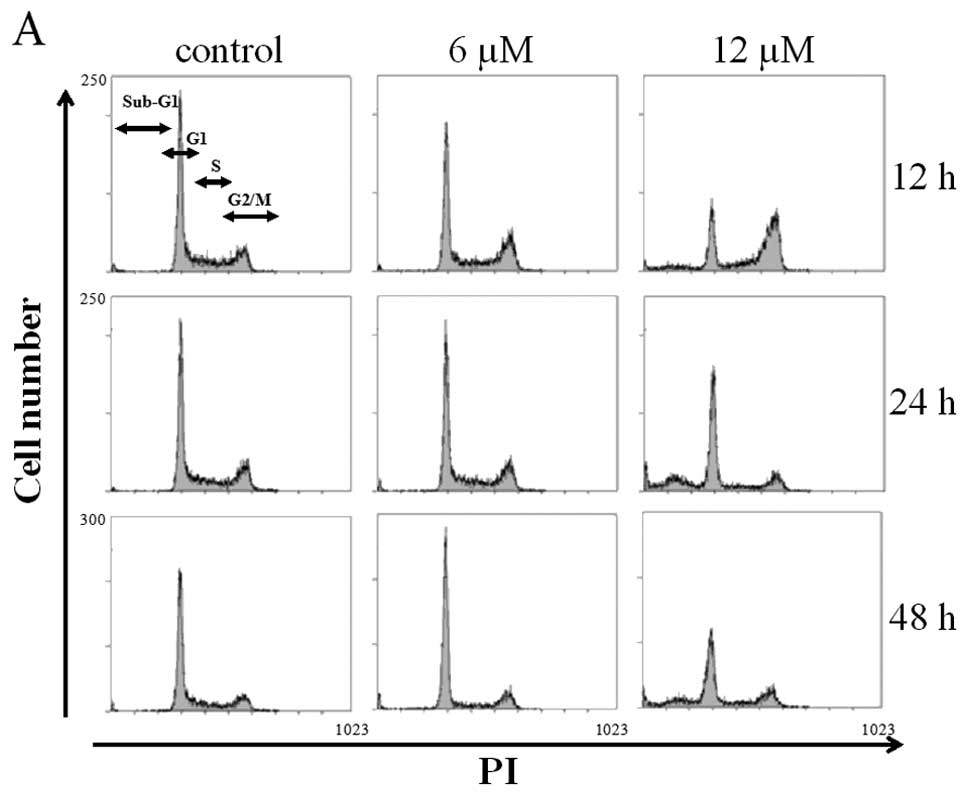
LCRC-1 induces G2/M arrest in the A549
cells
The effect of LCRC-1 on the cell cycle profile in
the A549 cells was analyzed. The treatment with 12 μM LCRC-1
for 12 h significantly increased the G2/M fraction compared with
treatment with the control (Fig.
3A). The increase in the G2/M fraction caused by treatment with
12 μM LCRC-1 disappeared at 24 h, whereas the sub-G1
fraction, a hallmark of apoptosis, was increased (Fig. 3B). In contrast, no statistically
significant difference in the cell cycle profiles was observed
between the treatments with the control and 6 μM LCRC-1.
LCRC-1 induces apoptosis in the A549
cells
Because the sub-G1 fraction was induced in the cells
treated with 12 μM LCRC-1, the analysis of apoptosis was
performed in detail. First, Annexin V/PI staining methods were used
to analyze the apoptosis induction. The percentages of the Annexin
V(+)/PI(−) cells with early apoptotic cells and Annexin V(+)/PI(+)
cells with late apoptotic cells were significantly higher in the
cells treated with LCRC-1 for 72 h compared with those of the
control (Fig. 4A). Although the
treatment with LCRC-1 for 48 h also increased those fractions, no
statistically significant difference was observed. To investigate
in detail the induction of apoptosis by LCRC-1, the expression of
active caspase-3, an executioner of apoptosis, was analyzed.
Caspase-3 activation in the cells treated with 12 μM LCRC-1
for 48 and 72 h was higher than that in the vehicle control
(Fig. 4B). To clarify the
involvement of caspase-3 activation in the induction of apoptosis
by LCRC-1, Z-VAD-FMK, a pan-caspase inhibitor, was used.
Pretreatment with Z-VAD-FMK moderately inhibited caspase-3
activation by LCRC-1 (Fig. 4C) and
reduced the percentage of Annexin positive cells in the cells
treated with LCRC-1 (Fig. 4D).
However, the reduction of cell number was not recovered by
treatment with Z-VAD-FMK (Fig.
4E).
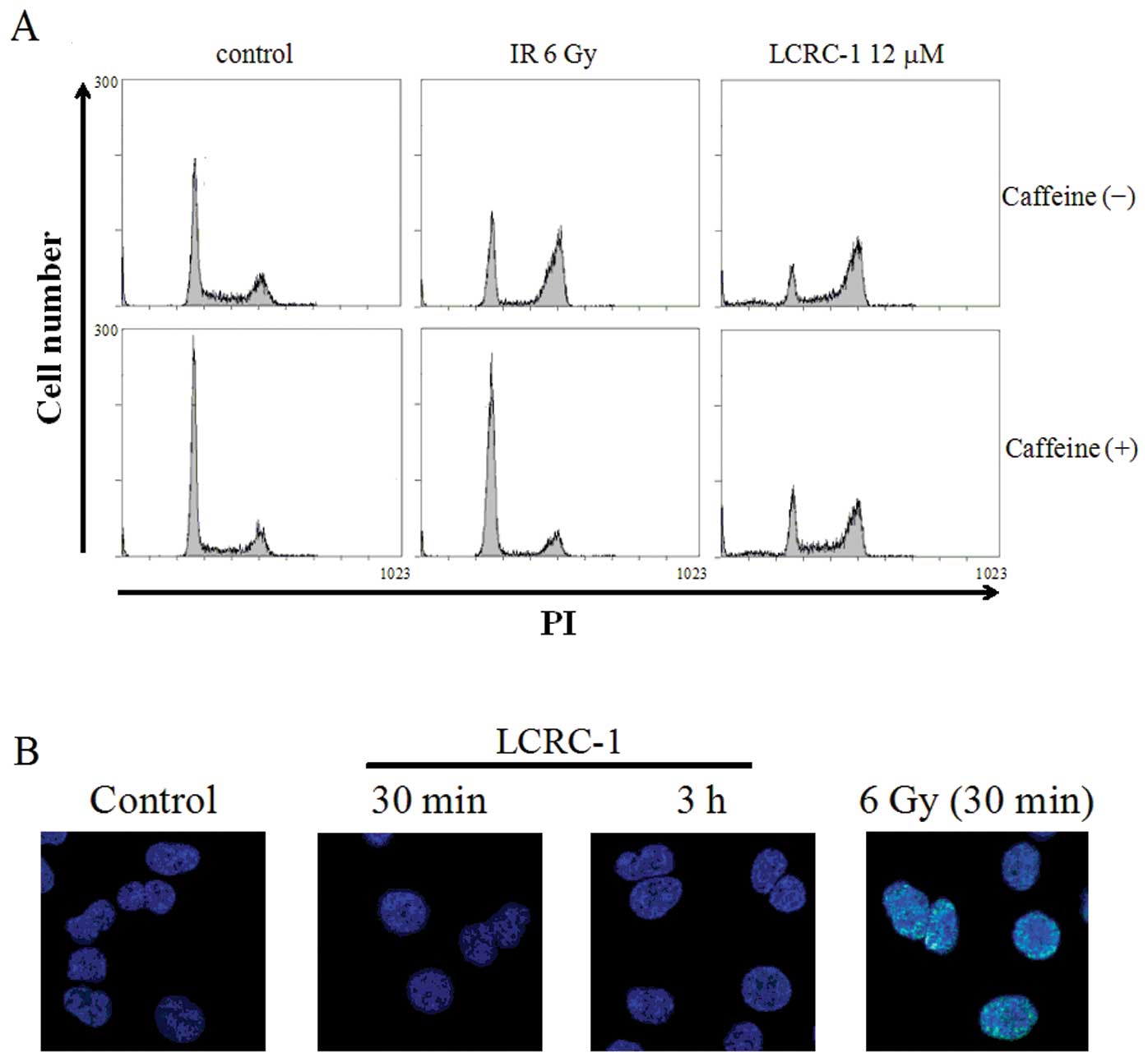
The effects of LCRC-1 in causing DNA
damage
To investigate the mechanisms of G2/M arrest by
LCRC-1, we focused on DNA damage. Because ataxia telangiectasia
mutated (ATM), which is one of the DNA damage-responsive kinases,
activates G2/M checkpoint arrest following DNA damage such as
exposure to ionizing radiation (15), caffeine, as an ATM inhibitor, was
used to investigate the involvement of ATM in G2/M arrest by LCRC-1
(16). Pretreatment with caffeine
completely inhibited G2/M arrest by 6-Gy irradiation, whereas it
hardly affected G2/M arrest by LCRC-1 (Fig. 5A). Furthermore, although the
expression of γ-H2AX, a marker of DNA damage (17), was observed in 6-Gy irradiated
cells prepared as positive controls, it was not observed in the
cells treated with LCRC-1 at any time points (0.5, 3, 24 h)
(Fig. 5B and data not shown).
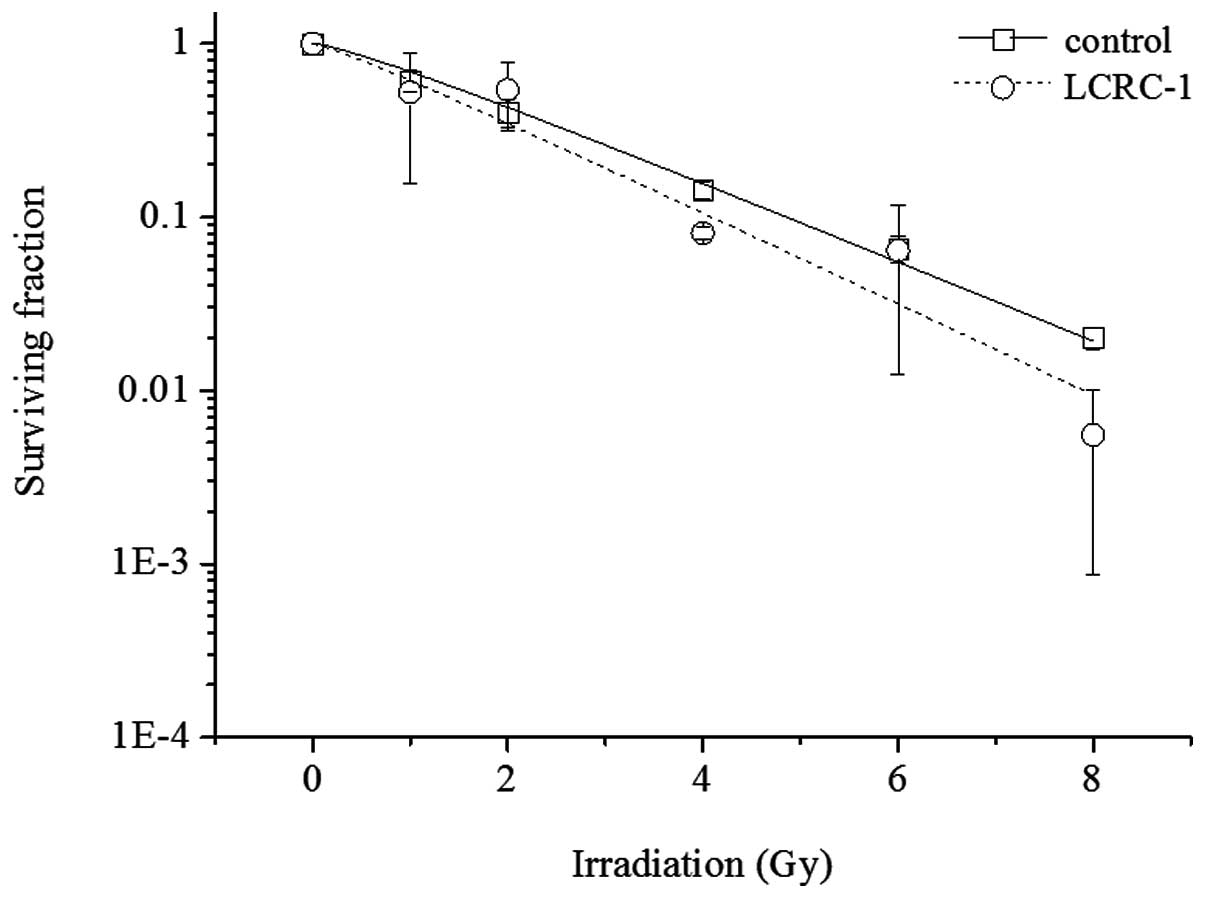
Radiosensitization effect of LCRC-1
The radiosensitization of LCRC-1 was examined. The
survival curves of the A549 cells exposed to X-rays with or without
6 μM LCRC-1 are shown in Fig.
6. Although LCRC-1 slightly enhanced radiosensitivity in the
A549 cells, no statistically significant difference was
observed.
Discussion
In the present study, we demonstrated that LCRCs
possessing three aromatic rings, whose potential to induce
apoptosis had been previously demonstrated in human chronic
myelogenous leukemia K562 cells (12), suppressed cell growth in human
non-small cell lung cancer A549 cells (Fig. 2A). Takahashi et al
previously reported that some amphiphilic LCCs such as
cyanobiphenyl derivatives with a terminal hydroxyl and
phenylpyrimidine derivatives possessing D-glucamine inhibited cell
growth through G1-phase arrest in the A549 cells, although they did
not induce cell death (11).
However, LCRCs used in this study (particularly LCRC-1) increased
the sub-G1 fraction and Annexin V-positive cells and activated
caspase-3 in the A549 cells (Figs.
3, 4A and B), thus showing
that LCRC-1 can induce apoptosis in the A549 cells. Furthermore,
the increase of Annexin V-positive cells caused by LCRC-1 was
partly inhibited by caspase inhibitor (Fig. 4D). This result indicates that
LCRC-1 induced apoptosis through a caspase-dependent pathway. In
contrast, the suppression of cell numbers by LCRC-1 did not recover
after treatment with a caspase inhibitor (Fig. 3E); the probable reason for this is
that the specific caspase inhibitor used may not prevent cell cycle
arrest by LCRC-1.
ATM is a DNA damage sensor and activates the DNA
repair process (18). Following
exposure to DNA damage, including that caused by ionizing
radiation, ATM activates G2/M-checkpoint arrest. However, caffeine,
an ATM inhibitor, hardly affected G2/M arrest by LCRC-1 (Fig. 5A). Furthermore, treatment with
LCRC-1 did not induce the expression of γ-H2AX, a marker of
DNA damage (Fig. 5B). Considering
these results, it is not likely that LCRC-1 induces DNA damage and
then results in G2/M arrest. On the other hand, c-Jun N terminal
kinase (JNK) is involved in not only apoptosis but also cell cycle
arrest (19). Furthermore, since
Fukushi et al reported that LCRC-1 activated JNK in the K562
cells (12), there is a
possibility that JNK is associated with suppression of LCRC-1 in
the A549 cells. However, a JNK inhibitor VIII
(Calbiochem®) did not affect cell suppression by LCRC-1
in the A549 cells (data not shown). Therefore, other mechanisms
such as endoplasmic reticulum stress may be involved in the
suppressive effects of LCRC-1 (20).
Combination treatment of radiotherapy with
chemotherapy such as cisplatin has been extensively used in the
management of many types of solid malignancies including NSCLC
(21,22). Although, the radiosensitization
effect of 6 μM LCRC-1 was expected in the A549 cells, it
hardly enhanced the radiosensitization (Fig. 6). However, LCRC-1 did not attenuate
the effect of ionizing radiation at other concentrations (1.5 and 3
μM, data not shown). Therefore, the combination treatment of
LCRC-1 with radiotherapy may be possible.
In conclusion, we demonstrated that an LCRC
possessing three aromatic rings (LCRC-1) can potentially suppress
cell growth through cell cycle arrest at the G2/M phase and
induction of apoptosis in the A549 cells. Interestingly, LCRC-1 did
not inhibit the proliferation of WI-38 normal fibroblasts (Fig. 1B). Although the mechanisms of cell
growth suppression by LCRC-1 were not elucidated in the present
study, the tumor-specific suppressive effect of LCRC-1 is
attractive for development of chemotherapeutic drugs. Therefore,
further studies regarding the molecular mechanisms and
structure-activity relationship of LCRCs may lead to the
development of tumor-specific and more powerful chemotherapeutic
drugs.
Acknowledgements
This study received support from a
grant for Hirosaki University Institutional Research (2011). This
study was partly supported by KAKENHI, Grant-in-Aid for Young
Scientists (B, No. 23791383 HY).
References
|
1
|
Tang N, Du G, Wang N, Liu C, Hang H and
Liang W: Improving penetration in tumors with nanoassemblies of
phospholipids and doxorubicin. J Natl Cancer Inst. 99:1004–1015.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cheng JH, Hung CF, Yang SC, Wang JP, Won
SJ and Lin CN: Synthesis and cytotoxic, anti-inflammatory, and
anti-oxidant activities of 2′,5′-dialkoxylchalcones as cancer
chemopreventive agents. Bioorg Med Chem. 16:7270–7276. 2008.
|
|
3
|
Goel A, Prasad AK, Parmar VS, Ghosh B and
Saini N: 7,8-Dihydroxy-4-methylcoumarin induces apoptosis of human
lung adenocarcinoma cells by ROS-independent mitochondrial pathway
through partial inhibition of FNK/MAPK signaling. FEBS Lett.
581:2447–2454. 2007. View Article : Google Scholar
|
|
4
|
Chou TC, Zhang X, Zhong ZY, Li Y, Feng L,
Eng S, Myles DR, Johnson R, Wu N, Yin YI, Wilson RM and Danishefsky
SJ: Therapeutic effect against human xenograft tumors in nude mice
by the third generation microtubule stabilizing epothilones. Proc
Natl Acad Sci USA. 35:13157–13162. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Park SY, Kim YM and Pyo H: Gefitinib
radiosensitizes non-small cell lung cancer cells through inhibition
of ataxia telangiectasia mutated. Mol Cancer. 9:2222010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Santin AD, Hermonat PL, Ravaggi A, Bellone
S, Roman J, Pecorelli S, Cannon M and Parham GP: Effects of
concurrent cisplatinum administration during radiotherapy vs.
radiotherapy alone on the immune function of patients with cancer
of the uterine cervix. Int J Radiat Oncol Biol Phys. 48:997–1006.
2000. View Article : Google Scholar
|
|
7
|
Stevenson CL, Bennett DB and
Lechuga-Ballesteros D: Pharmaceutical liquid crystals: the
relevance of partially ordered systems. J Pharm Sci. 94:1861–1880.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Goodby JW, Görtz V, Cowling SJ, Mackenzie
G, Martin P, Plusquellec D, Benvegnu T, Boullanger P, Lafont D,
Queneau Y, Chambert S and Fitremann J: Thermotropic liquid
crystalline glycolipids. Chem Soc Rev. 36:1971–2032. 2007.
View Article : Google Scholar
|
|
9
|
Luk YY, Campbell SF, Abbott NL and Murphy
CJ: Non-toxic thermotropic liquid crystals for use with mammalian
cells. Liq Cryst. 31:611–621. 2004. View Article : Google Scholar
|
|
10
|
Cervin C, Vandoolaeghe P, Nistor C, Tiberg
F and Johnsson M: A combined in vitro and in vivo study on the
interactions between somatostatin and lipid-based liquid
crystalline drug carriers and bilayers. Eur J Pharm Sci.
36:377–385. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Takahashi Y, Hazawa M, Takahashi K,
Nishizawa A, Yoshizawa A and Kashiwakura I: Suppressive effects of
liquid crystal compounds on the growth of the A549 human lung
cancer cell line. Invest New Drugs. 29:659–665. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fukushi Y, Hazawa M, Takahashi K,
Yoshizawa A and Kashiwakura I: Liquid crystal-related
compound-induced cell growth suppression and apoptosis in the
chronic myelogenous leukemia K562 cell line. Invest New Drugs.
29:827–832. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hannun YA: Apoptosis and the dilemma of
cancer chemotherapy. Blood. 89:1845–1853. 1997.PubMed/NCBI
|
|
14
|
Yanai H: Statcel - The useful add-in
software forms on Excel. 2nd edition. OMS: Tokyo; 2006
|
|
15
|
Shibata A, Barton O, Noon AT, Dahm K,
Deckbar D, Goodarzi AA, Löbrich M and Jeggo PA: Role of ATM and the
damage response mediator proteins 53BP1 and MDC1 in the maintenance
of G(2)/M checkpoint arrest. Mol Cell Biol. 30:3371–3383. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Blasina A, Price BD, Turenne GA and
McGowan CH: Caffeine inhibits the checkpoint kinase ATM. Curr Biol.
9:1135–1138. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kuo LJ and Yang LX: Gamma-H2AX - a novel
biomarker for DNA double-strand breaks. In Vivo. 22:305–309.
2008.PubMed/NCBI
|
|
18
|
Tomita M: Involvement of DNA-PK and ATM in
radiation-and heat-induced DNA damage recognition and apoptotic
cell death. J Radiat Res. 51:493–501. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cho SD, Li G, Hu H, Jiang C, Kang KS, Lee
YS, Kim SH and Lu J: Involvement of c-Jun N-terminal kinase in G2/M
arrest and caspase-mediated apoptosis induced by sulforaphane in
DU145 prostate cancer cells. Nutr Cancer. 52:213–224. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bourougaa K, Naski N, Boularan C,
Mlynarczyk C, Candeias MM, Marullo S and Fåhraeus R: Endoplasmic
reticulum stress induces G2 cell-cycle arrest via mRNA translation
of the p53 isoform p53/47. Mol Cell. 38:78–88. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Brattström D, Bergqvist M, Hesselius P,
Wagenius G and Brodin O: Different fraction schedules and
combinations with chemotherapy in radiation treatment of non-small
cell lung cancer. Anticancer Res. 20:2087–2090. 2000.PubMed/NCBI
|
|
22
|
Gupta S, Koru-Sengul T, Arnold SM, Devi
GR, Mohiuddin M and Ahmed MM: Low-dose fractionated radiation
potentiates the effects of cisplatin independent of the
hyper-radiation sensitivity in human lung cancer cells. Mol Cancer
Ther. 10:292–302. 2011. View Article : Google Scholar : PubMed/NCBI
|