Introduction
Gastric cancer is the second leading cause of
cancer-related deaths worldwide. In general, high degree of
mortality and short median survival time in gastric cancer is due
to the fact that by the time diagnosis is arrived at it is too
late, and that, in part, due to the advanced stage at which
patients seek medical attention, which can be ascribed to its
asymptomatic nature in early stages (1–3).
Previously, it was reported that selenium-binding protein 1 (SBP1)
may serve as an independent prognostic marker of human gastric
cancer. It is known that SBP1 levels may correlate with
differentiation, TNM stage, and lymph node metastasis in several
types of cancer (3,4). For instance, decreased expression of
SBP1 had been reported in prostate, stomach, colon, esophagus,
uterus, ovary and lung cancers. Due to its wide expression in a
variety of cancers, SBP1 could be an attractive target for cancer
therapy (5–10). Low levels of expression of SBP1
indicates poor prognosis in gastric, lung adenocarcinoma and
colorectal cancers (10,11). In contrast, high level of SBP1
expression is found in normal colonic epithelia (10) while its decreased expression is a
late event in colorectal cancer (12) that may indicate the rapid
progression of colorectal carcinoma (12). Furthermore, SBP1 is overexpressed
in the chemosensitive tissues (13) suggesting that its expression could
be considered as an important predictor of response of cancer to
chemotherapeutic drugs (6). It has
been suggested that SBP1 could form an important independent risk
factor to predict overall survival and disease recurrence (14).
SBP1 is an α-β protein present throughout the cell
and localizes at several places in the cell depending on the cell
types (15,16). SBP1 is predominantly present in the
cytosol and participates in the late stages of intra-Golgi protein
transport and participates in the intracellular transport of
selenium (17). SBP1 binds to
selenium via cysteine (Cys57) (16) and is thus, involved in
ubiquitination/deubiquitination pathways. Selenium by virtue of its
incorporation into selenoproteins shows a wide range of pleiotropic
effects, including its ability to prevent cancer and its
progression (6,10,18).
Diminished expression of SBP1 in human cancers is due to
methylation of the SBP1 promoter and by alternative splicing of
SBP1 mRNA (6). However, the exact
relationship between selenium levels and SBP1 expression in cancer
progression is yet to be clarified. Previous studies suggested that
selenium may exert its growth inhibitory action by modifying the
function of pre-existing proteins. SBP1 is a target of the
hypoxia-inducible factor-1 α (HIF1α) through which selenium may
modify SBP1 expression (19),
enhances glutathione peroxidase 1 (GPX1) activity without altering
its expression, and directly interact with von Hippel-Lindau
protein (pVHL) that plays a role in the proteasomal degradation
pathway in a selenium-dependent manner (6,15,19).
However, the exact mechanisms of SBP1 regulation and its anticancer
effects is not clear and needs to be investigated.
In the present study, the differences in protein
expression among normal gastric mucosa, early gastric carcinoma and
the corresponding gastric carcinoma tissues were investigated in a
cohort of patients by two-dimensional fluorescence difference in
gel electrophoresis (2D-DIGE) coupled with mass spectrometry (MS).
These evaluations have showed that the gradual SBP1 loss was
associated with an increased malignant grade. Then we evaluated the
role of SELENBP1 in gastric cancer by studying the expression of
SBP1 and its function employing gene knockout and overexpression
techniques especially its role in the proliferation, migration,
senescence and chemoresistance of gastric cancer cells to
cisplatin.
Materials and methods
Cell culture
SGC7901 and BGC823 cancer cells, used in the present
study, were cultured in RPMI-1640 media supplemented with 10% fetal
bovine serum (FBS) and 1% antibiotic antimyocytic (Gibco) and
maintained at 37°C in a humidified atmosphere containing 5%
CO2. Exponentially growing cells were removed from the
culture flasks using trypsin/EDTA, centrifuged at 800 rpm for 5
min, resuspended, and counted for use in subsequent experiments.
Stock cultures of each cell line were routinely sub-cultured at
least once a week and the media were changed every 2–3 days.
Construction of SELENBP1 stable cell
line
The pEGFP-SELENBP1, SELENBP1 small interfering RNA
(siRNA) and pEGFP vector (control) were purchased from Genepharma
Co. Ltd (Shanghai, China) and were stably transfected into SGC7901
and BGC823 cells using Lipofectamine 2000 (Invitrogen) according to
the manufacturer’s instruction. Stably transfected cells were
selected with G418. Selected clones were maintained in growth
medium containing 400 μg/ml geneticin.
Cell proliferation assay
Approximately 3,000 cells in 100 μl of medium
were plated in 96-well plates and allowed to attach for 24 h.
Subsequently, cells were treated with the cell proliferation
reagent MTS (Promega, Madison, WI, USA) to spectrophotometrically
evaluate cell proliferation, viability and chemosensitivity in
accordance with the manufacturer’s directions. Cultures were
collected at different time-points (24, 48, 72, 96, 120 h).
Relative proliferation rates were calculated as a percentage of the
initial T0 reading within each cell line.
Flat plate colony formation assay
The cells transfected steadily with
pEGFP-siRNA/pEGFP-SELENBP1 or empty vector were harvested and then
500 cells were plated in RPMI-1640/10% FBS on 6-well plates per
well, and were cultured for 2 weeks in the incubator at 37°C. The
clones were stained by 1 ml 0.1% crystal violet for 20 min. The
number of the clones in 10 randomly chosen fields was assessed by a
microscope.
Cell migration assay
Cells were trypsinized and resuspended in RPMI-1640
medium supplemented with 0.5% FBS. The cell suspension was adjusted
to a concentration of 1×106 cells/ml, and 200 μl
of cells were pipetted into the upper chamber of a Transwell plate
(Costar, Cambriged, MA, USA). The cells at 37°C in 5%
CO2 were allowed to attach for 1 h and the lower chamber
was filled with 600 μl of RPMI-1640 medium containing 20%
FBS. After 24 h of incubation at 37°C, the filter side of the upper
chamber was scraped with a cotton tip to eliminate cells that had
not migrated, the filter was removed and stained with 600 μl
0.1% crystal violet for 20 min. The cell number in 10 randomly
chosen fields was determined using a light microscope.
RNA preparation, complementary DNA
synthesis and quantitative real-time PCR
Total RNA was isolated from SGC7901 and BGC823 cell
lysates using an RNeasy mini kit according to the manufacturer’s
instructions. Total RNA was then treated with DNase I in the
presence of anti-RNase to remove DNA contamination before
complementary DNA synthesis. The complementary DNA was synthesized
with random primers and avian myeloblastosis virus reverse
transcriptase (Promega). Real-time PCR (power SYBR-Green, ABI,
Warrington, UK) analysis was performed using an ABI Prism 7500
sequence detector according to the manufacturer’s protocol. Primer
sequences were as follows: for SELENBP1, 5′-ATCACCGACATCCTGCTCT-3′
(forward), 5′-GACTTTAGTTCCTCGTCCTCC-3′ (reverse); and for β-actin,
5′-ATCATGTTTGAGACCTTCAA-3′ (forward), 5′-CATCTCTTGCTCGAAGTCCA-3′
(reverse). Fold changes in the genes of interest were calculated
after normalisation with the endogenous control β-actin and using
the comparative threshold cycle (Ct) method.
Protein extraction and immunoblot
analysis
Cells were scraped from culture plates, and the
final protein concentration of the cell lysates was determined
using the bicinchoninic acid (BCA) method with bovine serum albumin
as the standard. Equivalent cell extracts (20–40 μg of
protein) were boiled in 5X sodium dodecyl sulfate (SDS) at 95°C for
5 min, cooled on ice, and then total protein extracts were
separated by 10–12% SDS-PAGE (20–50 per lane), and
electro-transferred to polyvinylidene fluoride membranes.
Anti-SELENBP1 (1:1,000, Abcam, Cambridge, UK) and anti-β-actin
(1:1,000, Abcam) antibodies were diluted in TBST (TBST/tween; 5%
BSA powder) and incubated with the membranes at 4°C overnight. The
appropriate secondary antibody was applied (1:1,000, anti-rabbit)
at room temperature for 1 h. Immunoreactive proteins were
visualized by enhanced chemiluminescence.
Cell cycle analysis by flow
cytometry
SGC7901 and BGC823 cells (2.5×105/well)
were seeded onto 6-well cell culture plates and incubated with 20
μm of CDDP for 24 h. The cells were harvested and washed by
centrifugation. For apoptosis determination, cells were fixed by
70% ethanol in −20°C overnight and then re-suspended in PBS
containing 40 μg/ml PI in 37°C for 30 min and then added 100
μg/ml RNase A in 4°C dark room for 30 min. The cell
apoptosis was determined by flow cytometry (FACSCalibur™, Becton
Dickinson, Franklin Lakes, NJ, USA).
Senescence-associated β-galactosidase
(SA-b-Gal) activity
Cells for this assay were seeded in the dishes,
washed twice with PBS and fixed in PBS that contained 2%
formaldehyde/0.2% glutaraldehyde for 7 min at room temperature.
SA-b-Gal staining was performed in fresh senescence-associated
X-Gal staining solution at 37°C (no CO2). Incubation
typically lasted for 12 h. Cells were rinsed in PBS and stored in
PBS with 70% glycerol. Cells were then examined under a microscope
for blue-green staining of the cytoplasm that was indicative of
senescence. Digital imagines were taken at ×20 optical
magnification, and stained cells were counted and expressed as a
percentage of total cell number in three independent fields per
well per treatment group to obtain an average value for
β-galactosidase staining activity.
Xenograft assay
Cells were collected by trypsinization and washed
twice before injection. Cells (2×106 cells in 150
μl PBS) were injected subcutaneously into nude mice. All
injected mice formed tumours. Tumour volumes were measured twice
every week from week 2 to week 6 and calculated using the following
formula: 0.5236 × L1 × (L2)2, where L1 is the long axis
and L2 is the short axis of the tumour. Seven weeks after the
inoculation of the cancer cells, tumors were isolated and tumor
volume and weight were determined. Tissues were fixed in 10%
buffered formaldehyde solution and SBP1 expression level was
determined by western blot analysis and immunohistochemistry.
Experimental statistical analysis
Graphpad Prism5 and SPSS were used for statistical
analysis. Statistical comparison between two groups was performed
using the Student’s t-test. For comparison of more than three
groups, we used one-way analysis of variance, followed by Tukey’s
multiple comparison. P-values <0.05 were considered
statistically significant.
Results
Construction of SELENBP1 and siRNA stable
cell line and its confirmation
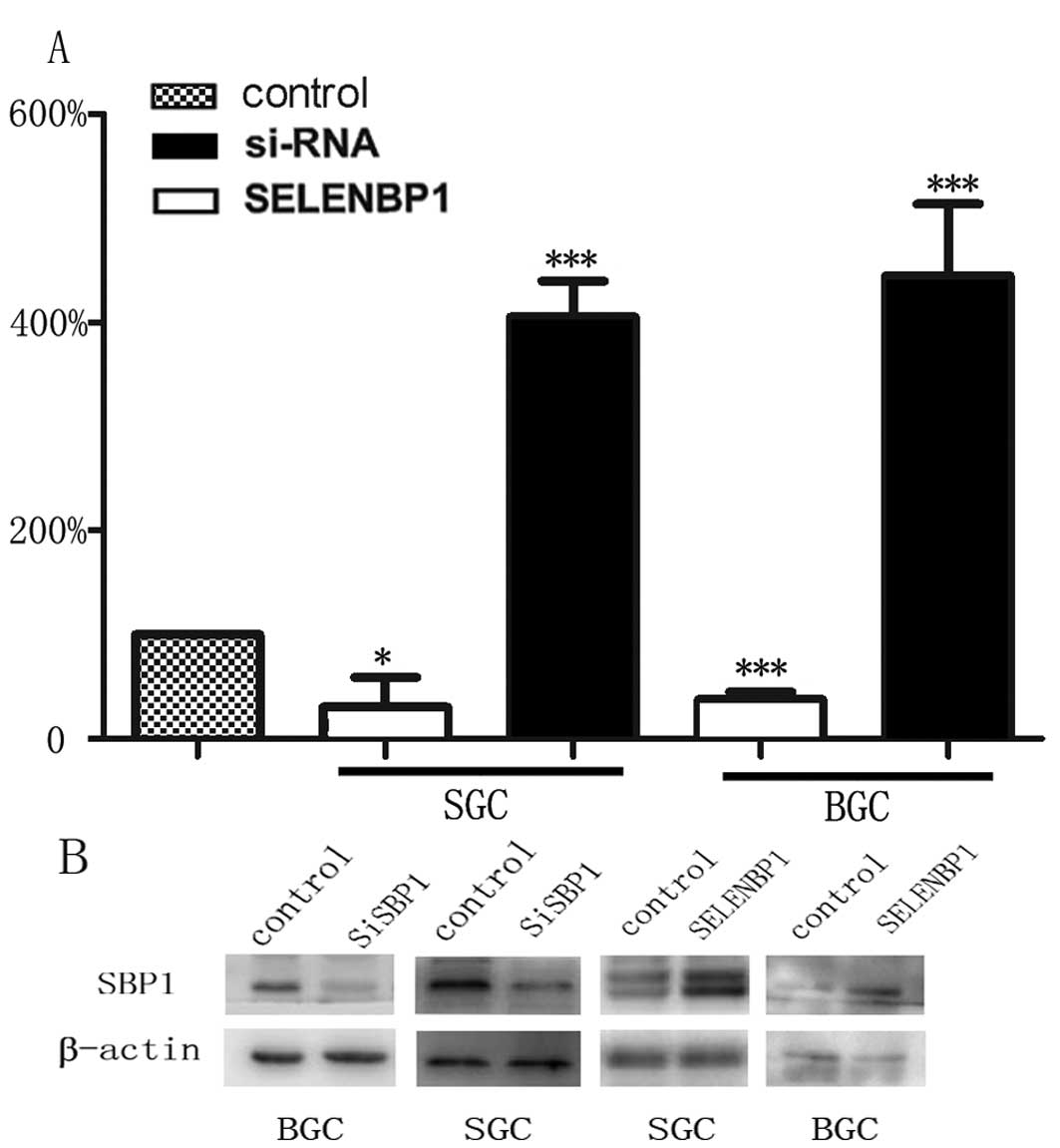
As reported previously, we observed that a subset of
tumor samples had higher expression of SBP1. To examine the
biological significance of SBP1 in gastric cancer, BGC823 and
SGC7901 cells were stably transfected with either
pEGFP-SELENBP1/pEGFP-siRNA or corresponding empty vector plasmid,
and the levels of SBP1 expression were determined by immunoblot
analysis and real-time PCR. Clone SELENBP1 expressed high levels,
whereas siRNA expressed low levels of SBP1 compared with empty
vector and untransfected BGC823 and SGC7901 cells (Fig. 1). When BGC823 and SGC7901 cells
were transfected with pEGFP-SELENBP1, the expression of SBP1 was
enhanced by 4-fold in SGC7901 cells and 5-fold in BGC823 cells,
while when transfected with pEGFP-siRNA, the expression of SBP1 was
suppressed by at least 70% in both SGC7901 and BGC823 cells.
Roles of SBP1 on cell growth in vivo and
in vitro
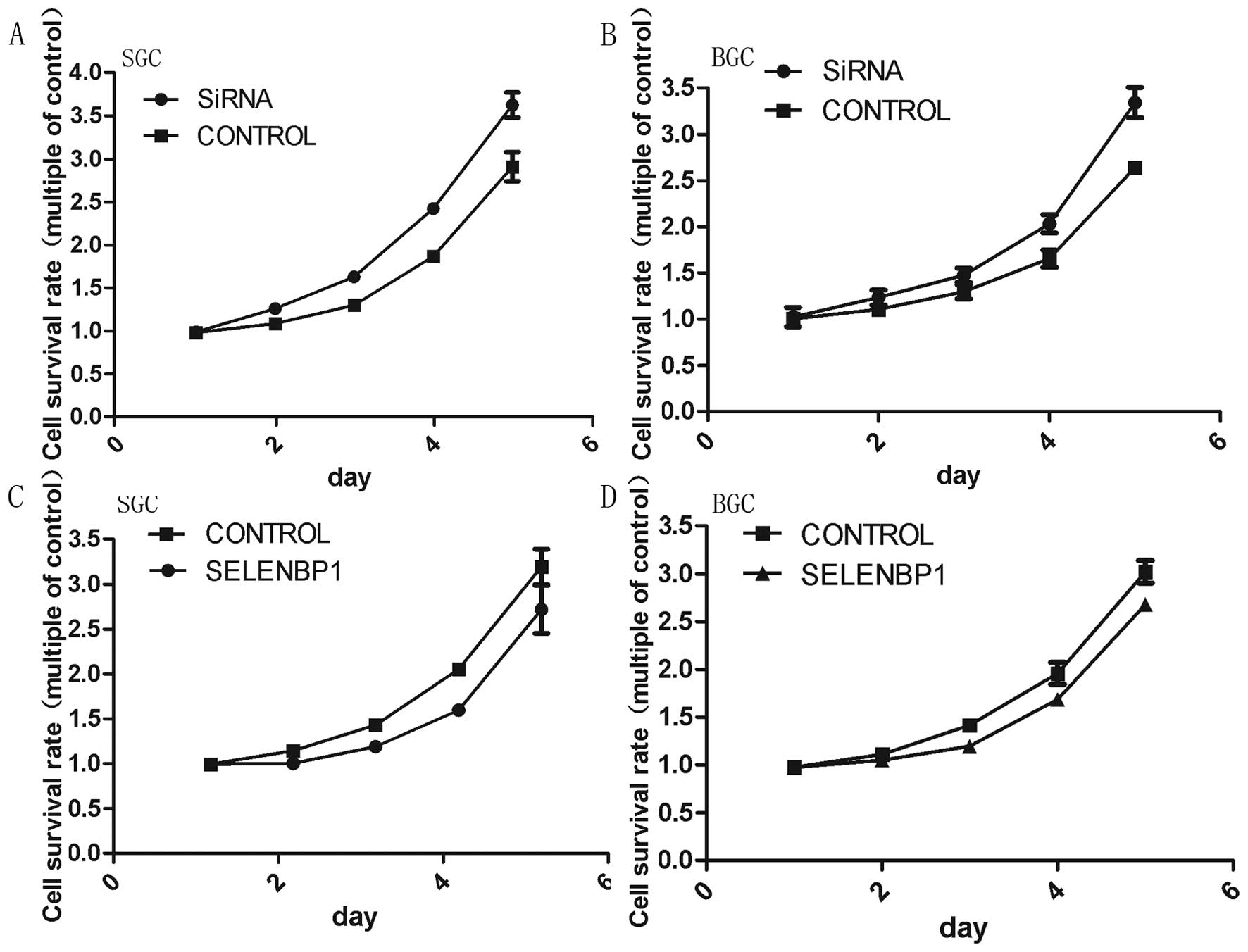
To determine the role of SBP1 on the proliferation
of cells, we conducted MTS assays after the transfection of
pEGFP-SELENBP1/ pEGFP-siRNA and the empty vector. Cultures were
collected at different time points for analysis of cell
proliferation. SBP1 downregulation significantly enhanced the
proliferation of SGC7901 and BGC823 cells, while SBP1 upregulation
remarkably reduced the proliferation of both cell lines (Fig. 3). The cell proliferation curve
showed that the proliferation of pEGFP-siRNA transfected cells
decreased at 24 h in both cell lines. Cell growth was much reduced
at 48, 72, 96 and 120 h, compared with the control groups
(P<0.01) (Fig. 3A and B). The
stable transfectants expressing SELENBP1 had incomplete inhibition
but moderate proliferation retardation (P<0.01) (Fig. 3C and D).
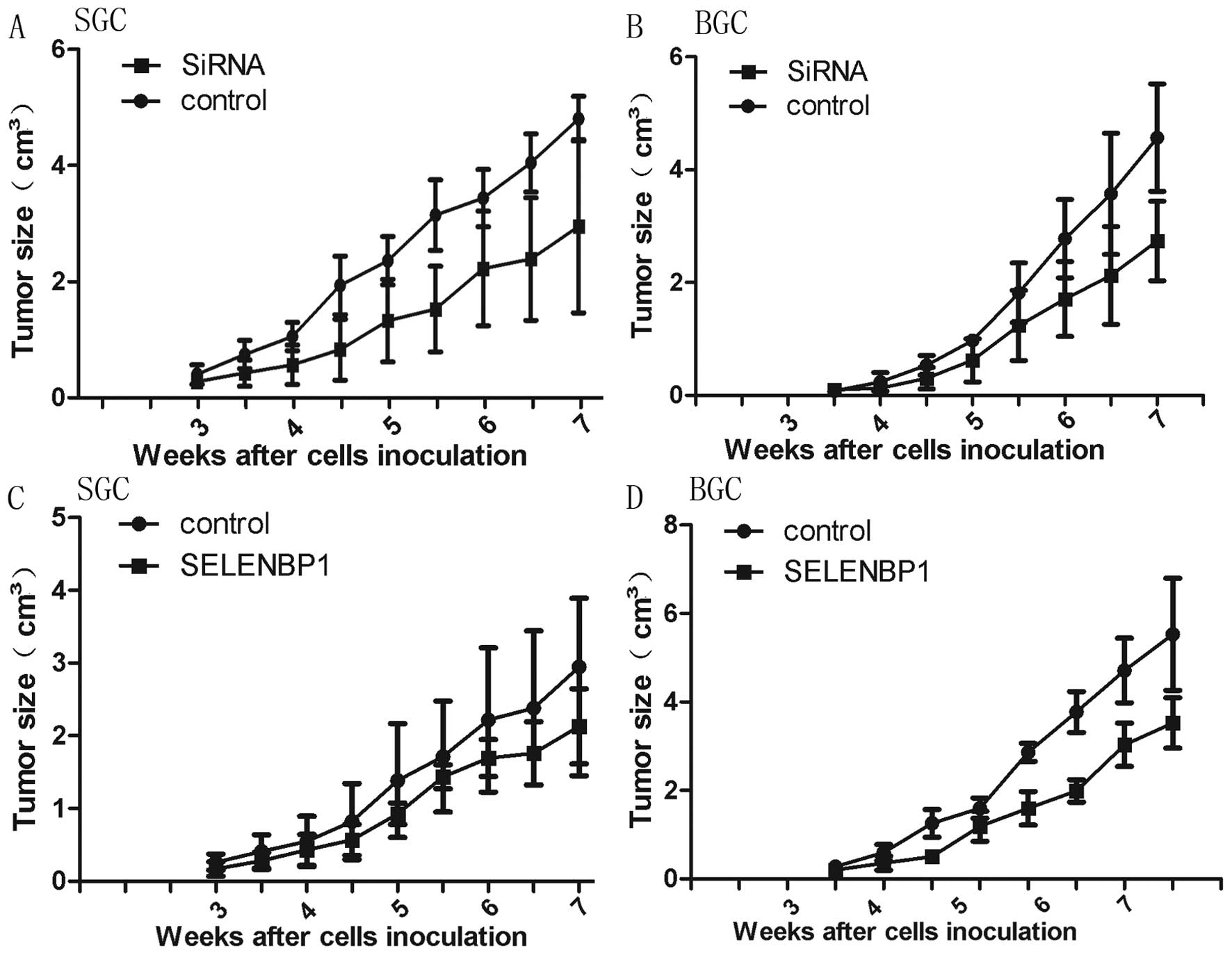
To confirm the effect of pEGFP-siRNA and
pEGFP-SELENBP1 in vivo, we subcutaneously inoculated nude
mice with BGC823 and SGC7901 which were either stably transfected
with pEGFP-siRNA/pEGFP-SELENBP1 or empty vector that formed the
control. We observed significant rate of enhancement of growth of
cancer cells that were stably transfected with pEGFP-siRNA and
decreased growth of cancer cells transfected with pEGFP-SELENBP1
(Fig. 4). Consistent with in
vitro data, mice injected with SBP1 overexpressing SGC7901 and
BGC823 cells showed significantly smaller tumor volume than mice
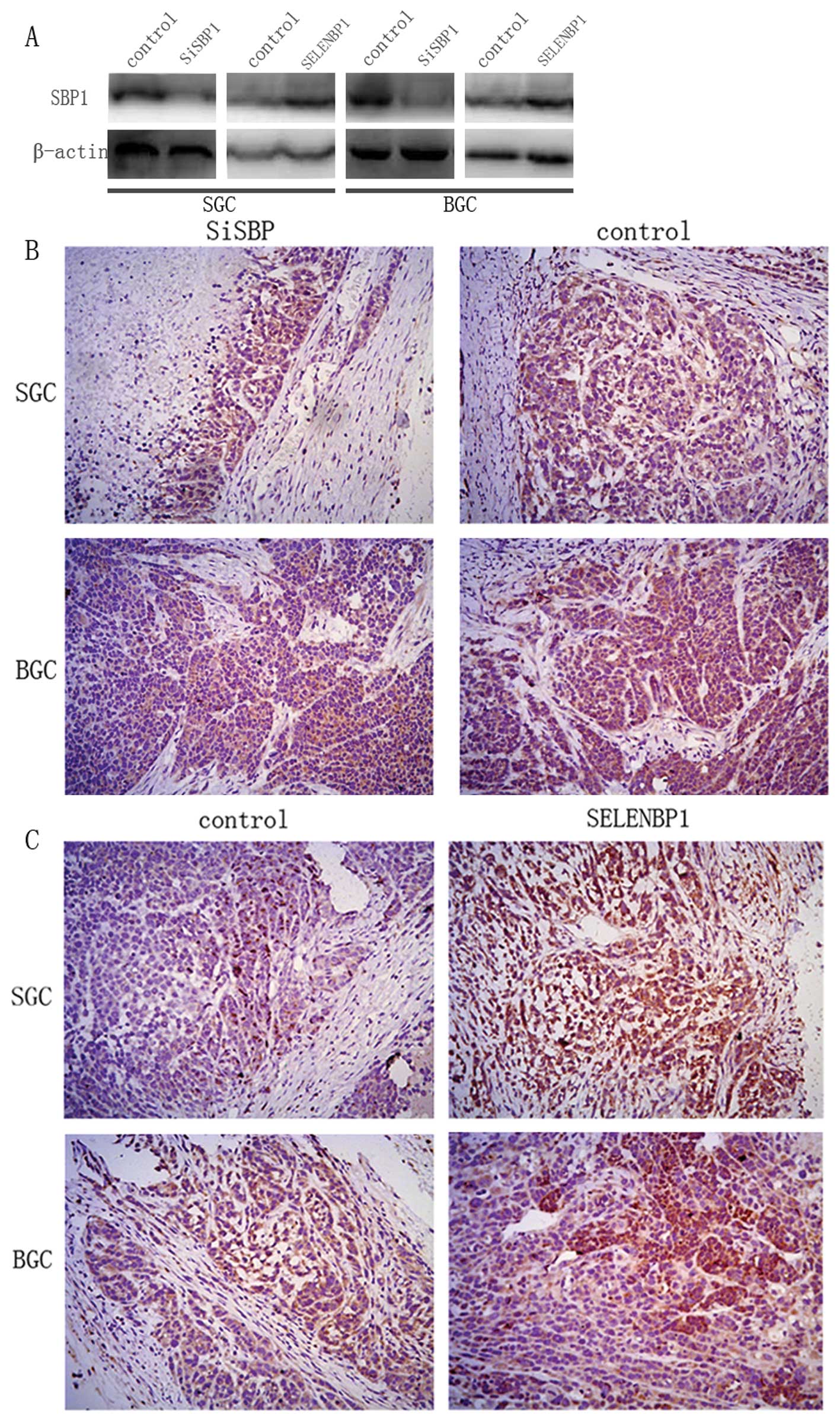
injected with control. Examination of tumour tissues of xenografts
showed significantly reduced expression of SBP1 in SiRNA group
while SELENBP1 group showed enhanced expression compared with the
control group (Fig. 5).
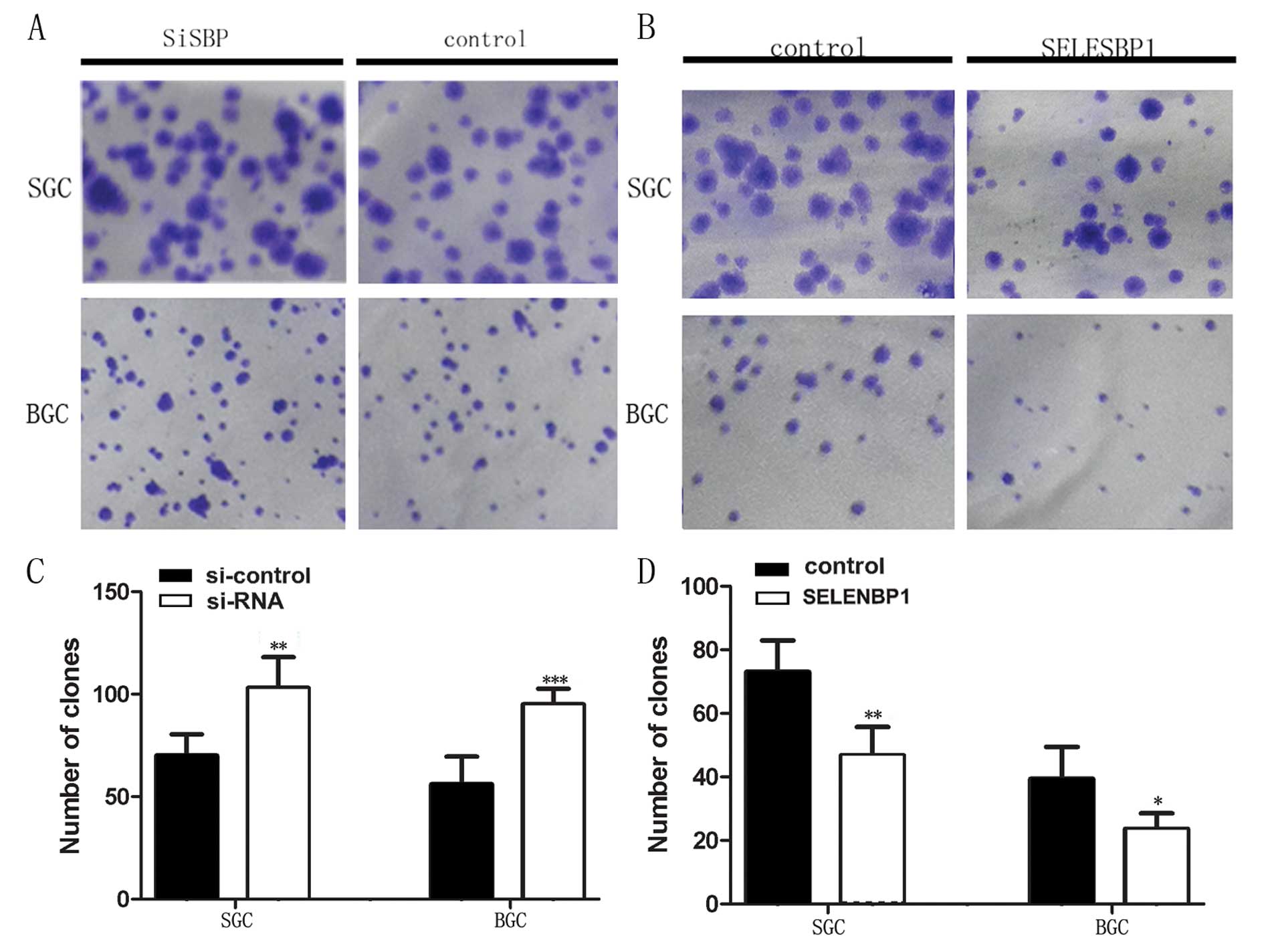
To examine the effect of downregulation and
overexpression of SBP1 on the colony formation of cells, stably
transfected SGC7901 and BGC823 cells were inoculated into 6-well
plates (500 cells). Number of colonies formed by SGC7901 and BGC823
cells transfected with pEGFP-siRNA were enhanced by 147%
(P<0.01) and 170% (P<0.001), respectively, compared to
coresponding cells transfected with empty vector (Fig. 2A and C). The colonies formed by the
pEGFP-SELENBP1 transfected SGC7901 and BGC823 cells were suppressed
to 64% (P<0.01) and 60% (P<0.05), respectively, compared to
control (Fig. 2B and D).
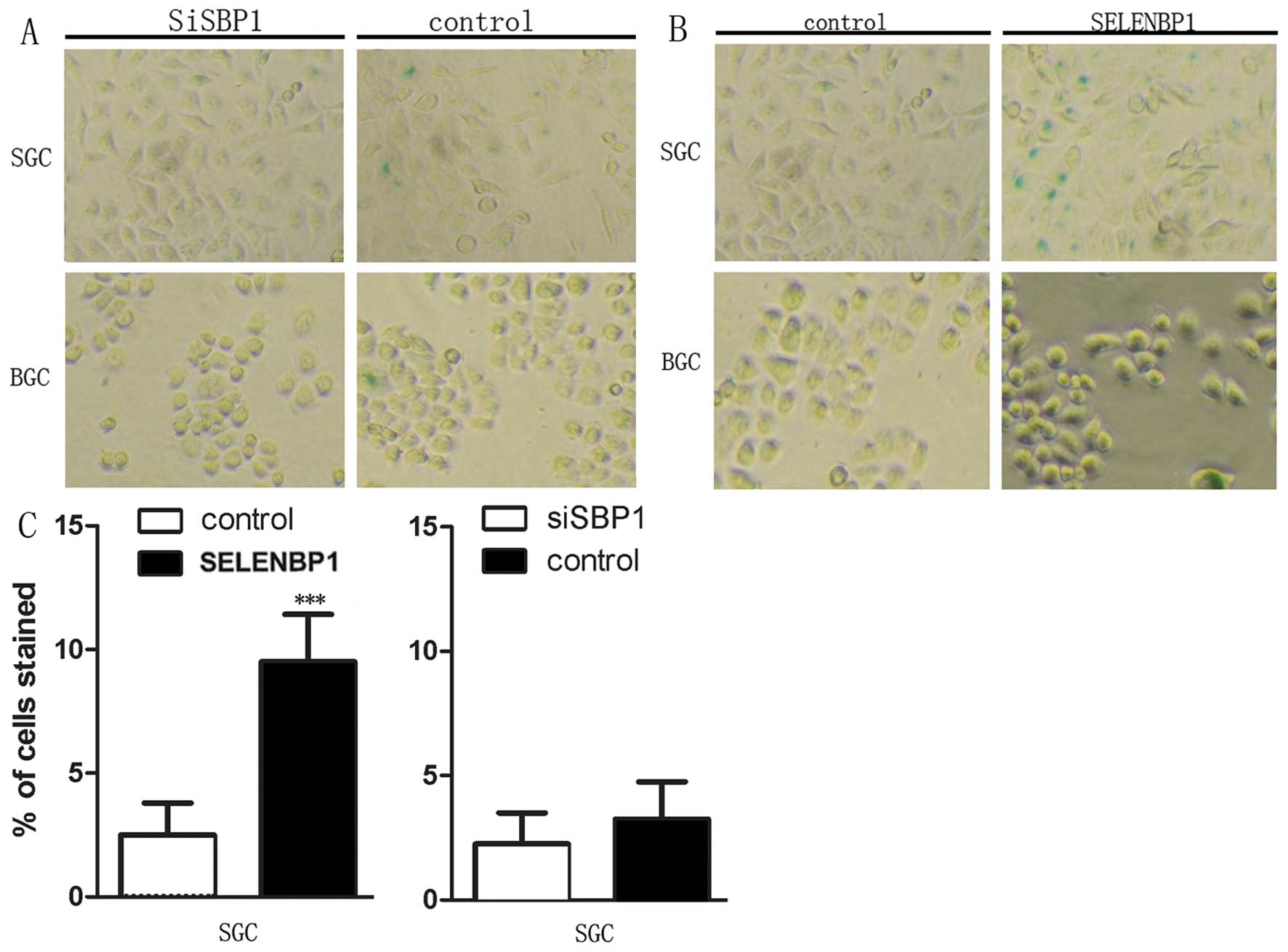
In addition, we also studied the role of SBP1 in
cancer cell senescence. It was noted that a significant difference
exists between cells transfected with empty vector and
pEGFP-siRNA/pEGFP-SELENBP1 in gastric cancer cell line SGC7901.
SBP1 significantly enhanced the senescence of SGC7901 cells as
assayed by β-galactosidase staining. Downregulation of SBP1
decreased senescence of SGC7901 gastric carcinoma cells (Fig. 6).
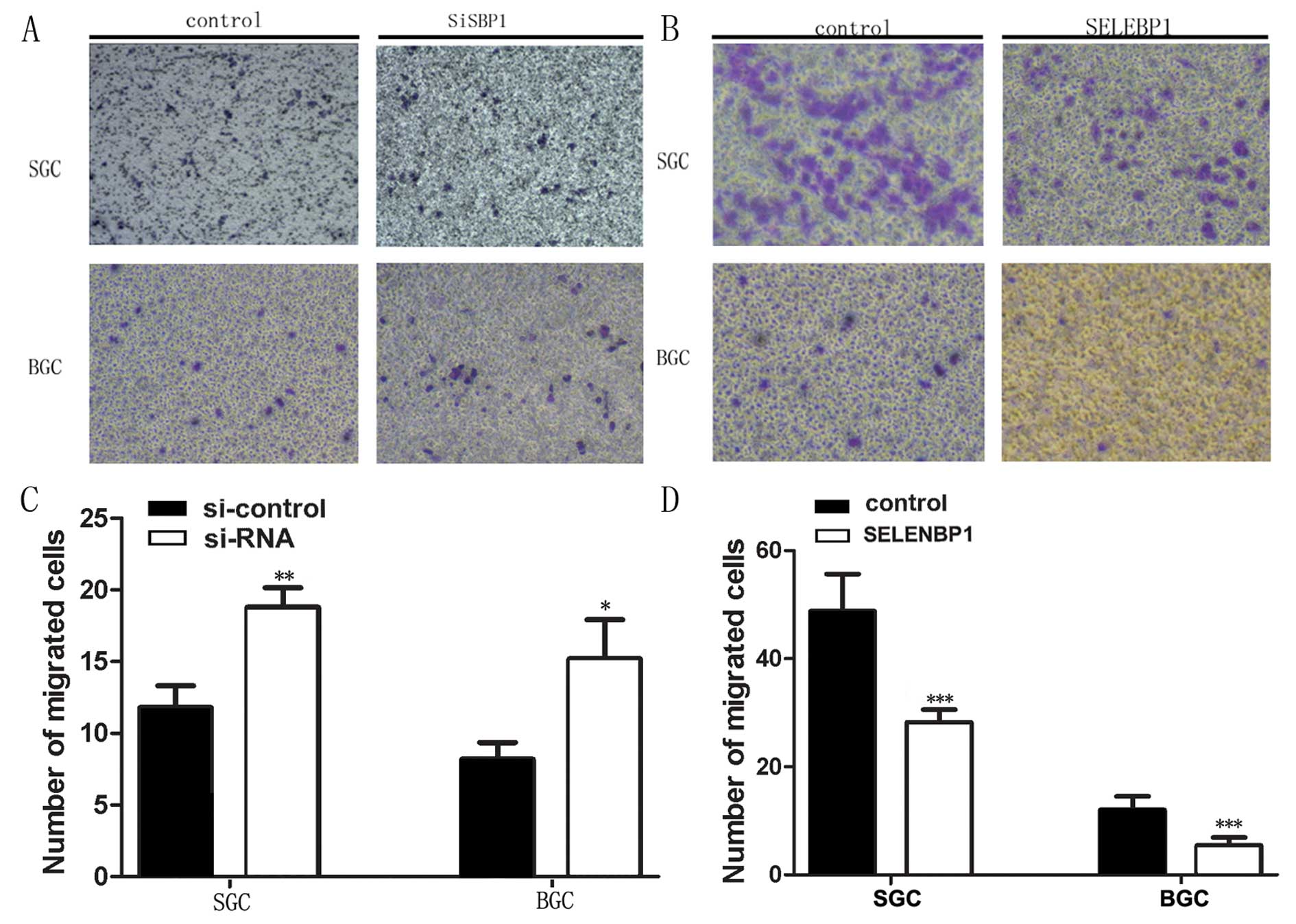
Effects of SELENBP1 on cell
migration
We next examined the role of SBP1 in cancer cell
migration. We observed a significant difference between cells
transfected with empty vector and pEGFP-siRNA/pEGFP-SELENBP1. SBP1
downregulation by pEGFP-siRNA (SiRNA) enhanced SGC7901 and BGC823
cell migration to 159% (P<0.01) and 185% (P<0.05),
respectively (Fig. 7A and C),
while SGC7901 and BGC823 stably transfected with pEGFP-SELENBP1
(SELENBP1) cells showed reduced migration by almost half the normal
[57% (P<0.001) and 45% (P<0.001), respectively] (Fig. 7B and D).
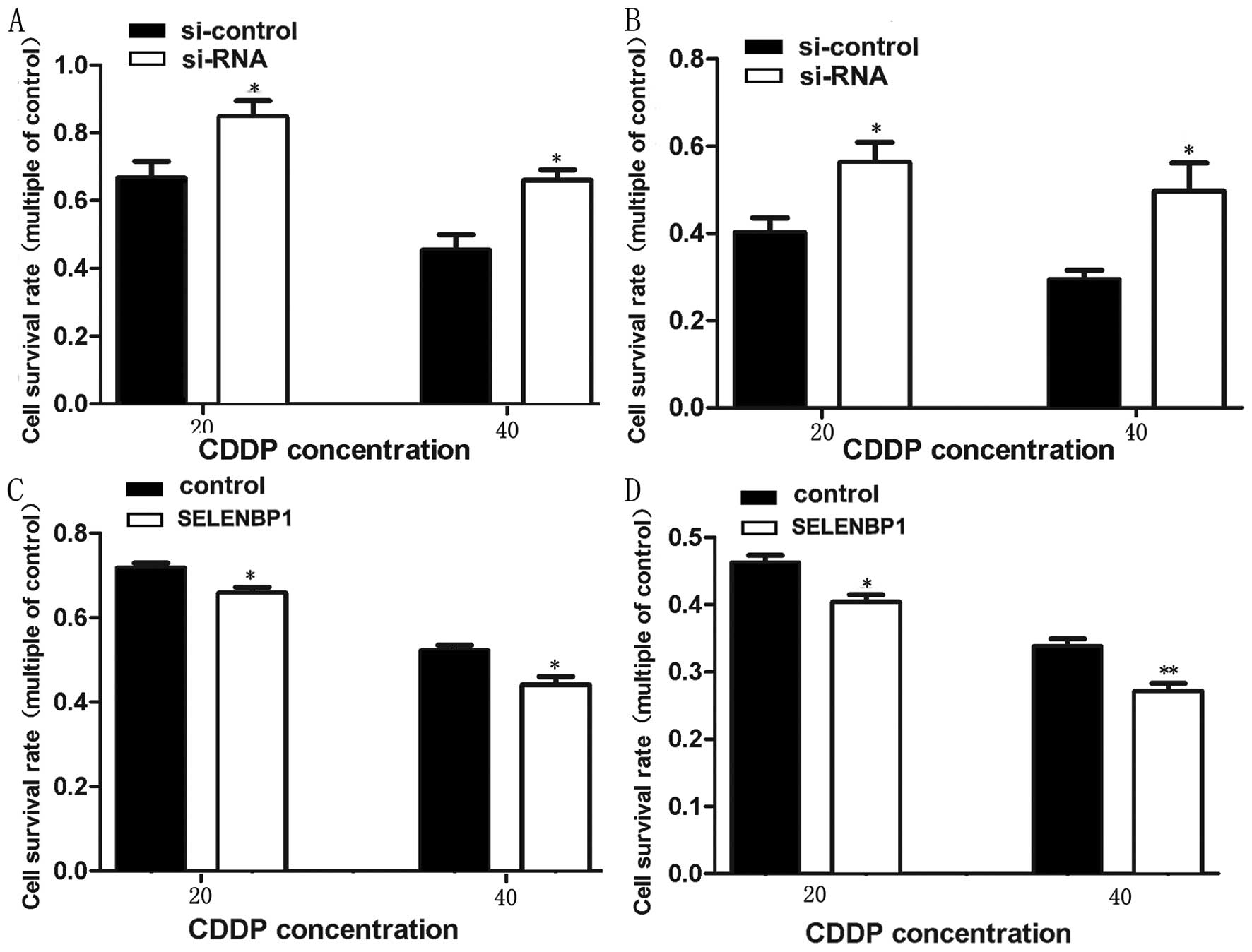
Roles of SBP1 on chemosensitivity to
CDDP
To determine if the pEGFP-siRNA and pEGFP-SELENBP1
transfected SGC823 and BGC7901 cells could change the cytotoxicity
of CDDP, stably transfected SGC7901 and BGC823 cells with
pEGFP-siRNA and pEGFP-SELENBP1 were exposed to different
concentrations of cisplatin and cell viability was analyzed at the
end of 24 h of treatment by MTS assay. It was observed that
pEGFP-siRNA transfected gastric cancer cells treated with 20 and 40
μmol/l cisplatin showed significantly enhanced viability
compared with control, while the viability pEGFP-SELENBP1
transfected cells was reduced (Fig.
8).
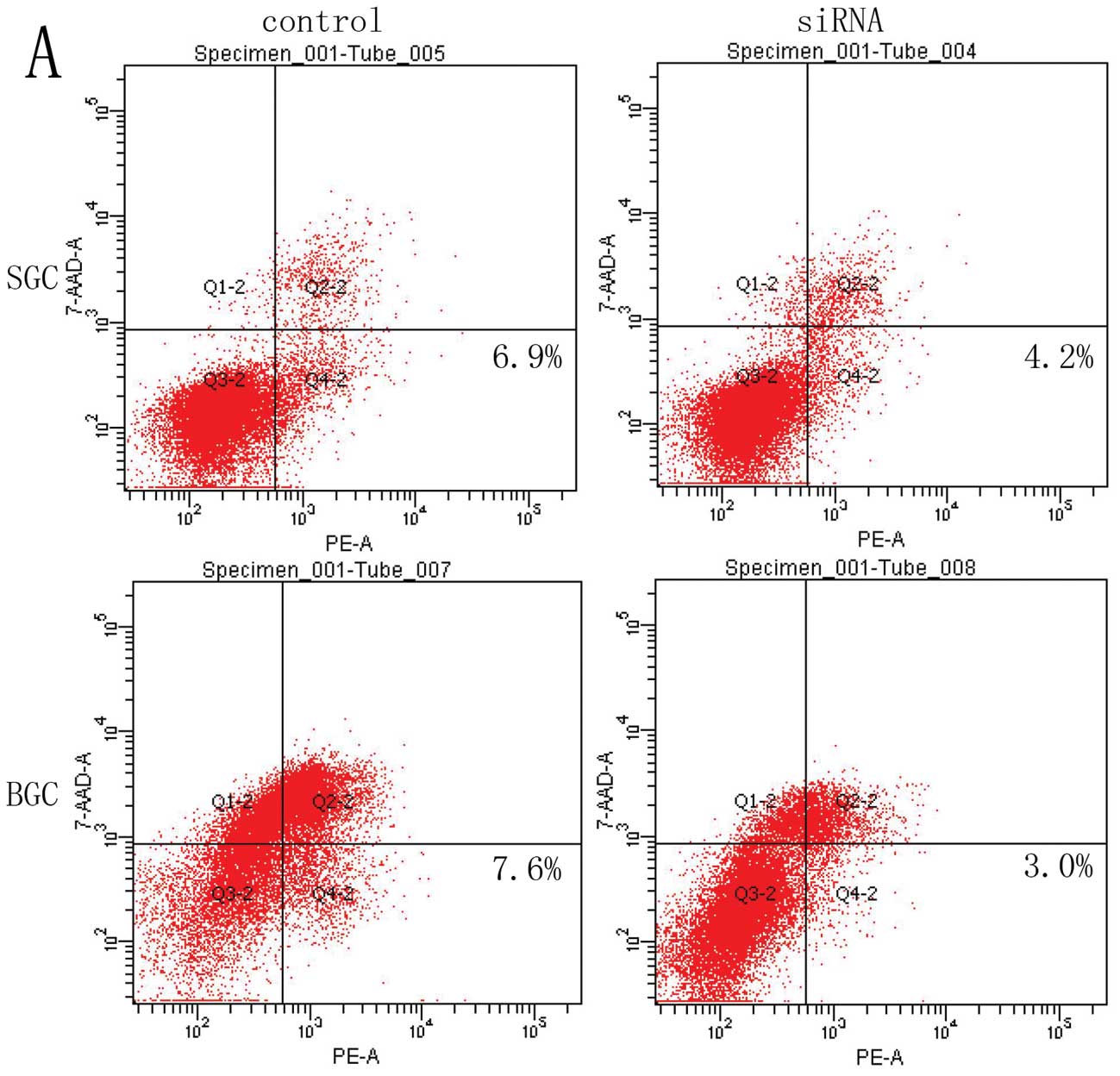
Apoptosis was measured by flow cytometry, but no
significant difference in the index of apoptosis was detected
between pEGFP siRNA/SELENBP1 transfected SGC823 and BGC7901 cells
and control (Fig. 9A and B).
However, when pEGFP-siRNA and pEGFP-SELENBP1 transfected SGC7901
and BGC823 cells were treated with 20 μmol/l cisplatin for
24 h, downregulation of SBP1 decreased the apoptosis of SGC7901 and
BGC823 cells in comparison with the empty vector-transfected
control. On the other hand, overexpression of SBP1 enhanced
cisplain-mediated apoptosis of both cell lines (Fig. 9C and D).
Discussion
Gastric cancer is common in Asian countries. Five
year survival rate of the advanced gastric cancer is only 30–40%,
whereas when gastric cancer is detected early, it is about 90%. The
mechanism of the development of gastric cancer is still not clear.
Previous studies suggested that SBP1 has the potential to be a
novel biomarker in predicting prognosis of gastric cancer. SBP1
level can be correlated with differentiation, TNM stage, and lymph
node metastasis of gastric cancer. The 3-year survival rate of
patients with high expression of SBP1 was significantly higher than
that of those with low expression. Suppression of SBP1 may be a
crucial event that may play a significant role in the progression
of gastric cancer (3,20) that implies that it has the
potential as a reliable diagnostic marker of gastric cancer
(21). In the present study, we
attempted a systematic study to understand the biological
significance of selenium-binding protein 1 in gastric cancer in
vitro and in vivo.
The observation that decreased expression of SBP1
occurs in human cancers led to the suggestion that it (SBP1) may
have tumor suppressive action. In the present study, we explored
that the probable tumor suppressor action of SBP1 by silencing and
overexpressing endogenous SBP1 expression in SGC7901 and BGC823
cells and analyzed phenotypic changes of stably transfected cells.
We could achieve almost complete downregulation and significant
upregulation of SBP1 expression using the transfection of SGC7901
and BGC823 cells by pEGFP-siRNA/pEGFP-SELENBP1 plasmids.
Uncontrolled cell proliferation and colony formation
are important hallmarks of cancer. Previously, it was reported that
overexpression of SBP1 in HCT116 cells suppressed cell
proliferation and inhibited tumor growth in vivo(6). In esophageal adenocarcinoma model, it
was noted that overexpression of SBP1 potentiated the
antiproliferative response of selenium supplementation,
particularly those cells that showed higher SBP1 expression
(9).
Inhibition of SBP1 effectively increased cell
motility, promoted cell proliferation in hepatocellular carcinoma
(14). In the present study, we
observed for the first time that SBP1 regulates proliferation of
gastric cancer cells SGC7901 and BGC823. SBP1 downregulation in
SGC7901 and BGC823 cells led to a significant inhibition of
proliferation and colony formation, while SBP1 upregulation
promoted proliferation and colony formation of both cell lines.
SBP1 downregulation and upregulation seems to be a determining
factor that alters SGC7901 and BGC823 cell growth in vitro
that, in turn, is supported by the consistent results obtained
in vivo. Mice injected with SBP1 overexpressing SGC7901 and
BGC823 cells had a significantly smaller tumor volume than the mice
injected with control cells. Besides, SBP1 expression was
significantly increased in tumor tissues injected with
pEGFPSELENBP1 transfected cells and decreased in tumor tissues
injected with pEGFP-siRNA transfected cells. These results are in
tune with the suggestion that SBP1 has the potential to suppress
gastric tumor formation. We propose that one mechanism by which
SBP1 regulates cell proliferation is by virtue of its ability to
covalently bind to selenium. Population-based studies suggested
higher serum selenium concentration is inversely related to the
incidence of a variety of epithelial malignancies (22). But the exact relationship between
selenium levels and SBP1 expression in cancer progression is not
known.
Overexpression of SBP1 accelerated senescence in
mice (9,23). Selenium supplements were suggested
to work in conjunction with SBP1 to activate senescence in human
esophageal adenocarcinoma and thus, retard their growth. To
determine if SBP1 overexpression activated cellular senescence in
human gastric cancer cells, we studied stably transfected SGC7901
and BGC823 cells for the expression of senescence-associated
β-galactosidase which revealed that that enhanced expression of
SBP1 in SGC7901 cells, but not BGC823, increased senescence as
determined by β-galactosidase staining. It was also noted that
diminished expression of SBP1 decreased senescence of SGC7901
gastric carcinoma cells. However, we noted that only few BGC cells
showed staining for β-galactosidase. We suspected that this
negative results could have resulted from either BGC823 cells not
being sensitive to β-galactosidase staining or β-galactosidase
expression was not high enough to be detected in BGC823 cell lines.
Nonetheless, SBP1-transfected cells showed increased senescence.
These data suggest that SBP1 activates the senescence pathway in
some gastric cancer cells.
Downregulation of SBP1 increased the migration of
SGC7901 and BGC823 gastric cancer cells, while pEGFP-SELENBP1
transfected SGC7901 and BGC823 cells showed decreased migration.
These results emphasize the important role of SBP1 in its ability
to suppress tumor growth and metastasis. SBP1 is known to suppress
colorectal cancer cell migration (6). Decreased expression of SBP1 promoted
tumor invasiveness by increasing GPX1 activity and diminishing
HIF-1α expression in hepatocellular carcinoma (14). Regulation of cancer cell migration
by SBP1 may explain greater lymph node metastasis and poor
prognosis observed in patients with gastric cancers with low SBP1
expression.
The significance of SBP1 in response of cancer cells
to anticancer drugs observed in the present study is rather
interesting. SGC7901 and BGC823 cells showed decreased vitality in
response to cis-platinum treatment. Overexpression of SELENBP1
enhanced cisplatin-mediated apoptosis. We suggest that enhanced
SBP1 expression in gastric cancer may increase the chemosensitivity
of gastric cancer cells via apoptotic signaling pathways. This is
supported by the observation that SBP1 overexpression in colorectal
cancer cells sensitized them to H2O2-induced
growth inhibition (9). Decreased
SBP1 expression effectively inhibited apoptosis under oxidative
stress and greatly enhanced glutathione peroxidase 1 (GPX1)
activity without altering GPX1 expression and downregulated
hypoxia-inducible factor-1α (HIF-1α) expression. SBP1 and GPX1
formed nuclear bodies and are co-localized under oxidative stress
(14). Similar results have been
reported in esophageal adenocarcinoma where it was noted that
diminished SBP1 expression blunted the cellular response to
selenium supplementation (6).
Based on these results, we propose that SBP1 could serve as a
reliable target for gastric carcinoma therapy. It has been
suggested that enhanced oxidative stress in cancer cells would lead
to cellular apoptosis due to the inhibition of GPX1 activity by the
upregulated SBP1 (14). We suspect
that similar mechanism may be occurring in gastric cancer
cells.
It is evident from the results of previous studies
and the present investigation that diminished SBP1 expression may
indicate poor prognosis in the gastric cancer. SBP1 overexpression
in gastric cancer may increase the chemosensitivity of gastric
cancer cells via apoptotic signaling pathways. However, further
studies are needed to explore other possible mechanisms of the
anticancer action of SBP1.
Acknowledgements
This research was supported by grants
from the National Health Key Special Fund (no. 200802112), the
Health Department Fund (no. 2007A093), the Traditional Chinese
Medicine Bureau Fund (no. 2007ZA019), the Natural Science Fund of
Zhejiang Province (nos. Y2080001, Y12H160121 and Z2080514), and the
Key Project of Zhejiang Province (no. 2009C03012-5). U.N.D. is a
recipiant of Ramalingaswami Fellowship of the Department of
Biotechnology, India during the tenure of this study.
References
|
1.
|
Wagner AD, Grothe W, Haerting J, Kleber G,
Grothey A and Fleig WE: Chemotherapy in advanced gastric cancer: a
systematic review and meta-analysis based on aggregate data. J Clin
Oncol. 24:2903–2909. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Lee JH, Kim KM, Cheong JH and Noh SH:
Current management and future strategies of gastric cancer. Yonsei
Med J. 53:248–257. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Xia YJ, Ma YY, He XJ, Wang HJ, Ye ZY and
Tao HQ: Suppression of selenium-binding protein 1 in gastric cancer
is associated with poor survival. Hum Pathol. 42:1620–1628. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Zhang J, Dong WG and Lin J: Reduced
selenium-binding protein 1 is associated with poor survival rate in
gastric carcinoma. Med Oncol. 28:481–487. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Zhang P, Zhang C, Wang X, et al: The
expression of selenium-binding protein 1 is decreased in uterine
leiomyoma. Diagn Pathol. 5:802010. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Silvers AL, Lin L, Bass AJ, et al:
Decreased selenium-binding protein 1 in esophageal adenocarcinoma
results from posttranscriptional and epigenetic regulation and
affects chemosensitivity. Clin Cancer Res. 16:2009–2021. 2010.
View Article : Google Scholar
|
|
7.
|
Zhang C, Wang YE, Zhang P, et al:
Progressive loss of selenium-binding protein 1 expression
correlates with increasing epithelial proliferation and papillary
complexity in ovarian serous borderline tumor and low-grade serous
carcinoma. Hum Pathol. 41:255–261. 2010. View Article : Google Scholar
|
|
8.
|
Wu C, Luo Z, Chen X, et al:
Two-dimensional differential in-gel electrophoresis for
identification of gastric cancer-specific protein markers. Oncol
Rep. 21:1429–1437. 2009.PubMed/NCBI
|
|
9.
|
Pohl NM, Tong C, Fang W, Bi X, Li T and
Yang W: Transcriptional regulation and biological functions of
selenium-binding protein 1 in colorectal cancer in vitro and in
nude mouse xenografts. PLoS One. 4:e77742009. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Li T, Yang W, Li M, et al: Expression of
selenium-binding protein 1 characterizes intestinal cell maturation
and predicts survival for patients with colorectal cancer. Mol Nutr
Food Res. 52:1289–1299. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Chen G, Wang H, Miller CT, et al: Reduced
selenium-binding protein 1 expression is associated with poor
outcome in lung adenocarcinomas. J Pathol. 202:321–329. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Kim H, Kang HJ, You KT, et al: Suppression
of human selenium-binding protein 1 is a late event in colorectal
carcinogenesis and is associated with poor survival. Proteomics.
6:3466–3476. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Pan S, Cheng L, White JT, et al:
Quantitative proteomics analysis integrated with microarray data
reveals that extracellular matrix proteins, catenins, and p53
binding protein 1 are important for chemotherapy response in
ovarian cancers. OMICS. 13:345–354. 2009. View Article : Google Scholar
|
|
14.
|
Huang C, Ding G, Gu C, et al: Decreased
selenium-binding protein 1 enhances glutathione peroxidase 1
activity and down-regulates HIF-1alpha to promote hepatocellular
carcinoma invasiveness. Clin Cancer Res. 18:3042–3053. 2012.
View Article : Google Scholar
|
|
15.
|
Jeong JY, Wang Y and Sytkowski AJ: Human
selenium binding protein-1 (hSP56) interacts with VDU1 in a
selenium-dependent manner. Biochem Biophys Res Commun. 379:583–588.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Raucci R, Colonna G, Guerriero E, et al:
Structural and functional studies of the human selenium binding
protein-1 and its involvement in hepatocellular carcinoma. Biochim
Biophys Acta. 1814:513–522. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Porat A, Sagiv Y and Elazar Z: A 56-kDa
selenium-binding protein participates in intra-Golgi protein
transport. J Biol Chem. 275:14457–14465. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Rayman MP: Selenium and human health.
Lancet. 379:1256–1268. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Scortegagna M, Martin RJ, Kladney RD,
Neumann RG and Arbeit JM: Hypoxia-inducible factor-1alpha
suppresses squamous carcinogenic progression and
epithelial-mesenchymal transition. Cancer Res. 69:2638–2646. 2009.
View Article : Google Scholar
|
|
20.
|
He QY, Cheung YH, Leung SY, Yuen ST, Chu
KM and Chiu JF: Diverse proteomic alterations in gastric
adenocarcinoma. Proteomics. 4:3276–3287. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Zhang J, Zhan N and Dong WG: Altered
expression of selenium-binding protein 1 in gastric carcinoma and
precursor lesions. Med Oncol. 28:951–957. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Blot WJ, Li JY, Taylor PR, et al:
Nutrition intervention trials in Linxian, China: supplementation
with specific vitamin/mineral combinations, cancer incidence, and
disease-specific mortality in the general population. J Natl Cancer
Inst. 85:1483–1492. 1993. View Article : Google Scholar
|
|
23.
|
Cho YM, Bae SH, Choi BK, et al:
Differential expression of the liver proteome in senescence
accelerated mice. Proteomics. 3:1883–1894. 2003. View Article : Google Scholar : PubMed/NCBI
|