Introduction
Colorectal cancer is most commonly associated with a
mutation in adenomatous polyposis coli (APC) gene and is the second
leading cause of deaths due to cancer in the United States
(1). The
ApcMin/+ mouse exhibits a nonsense mutation at
codon 850 in the APC gene (2)
that, similar to that seen in clinical studies, exhibits intestinal
neoplasms and develops a cachectic state characterized by severe
muscle wasting with increasing tumor burden (3,4). The
etiology of cachexia is multifactorial and is most likely mediated,
at least in part, by humoral factors that are secreted from or
induced by the tumor which cause an imbalance between the rates of
protein synthesis and breakdown (5).
The factors which regulate protein turnover in
striated muscle have not been fully elucidated, however, studies
have suggested that the 5′-adenosine monophosphate-activated
protein kinase (AMPK) and PI3K/Akt/mTOR signaling pathways may play
key roles (6). AMPK is a key
cellular energy sensor that is activated by a diminished
intracellular ATP/ADP ratio. Activation of AMPK serves to increase
energy availability and suppresses energy demanding processes in
the cell. Targets of AMPK activation include the phosphorylation of
Unc-51-like kinase-1 (ULK1) which can lead to the activation of
autophagy (7). AMPK also
suppresses the mammalian target of rapamycin complex 1 (mTORC1)
formation, which functions as central regulator of cell growth that
when phosphorylated acts to stimulate increases in protein
synthesis (8).
While most published reports have focused on the
effects of cachexia on skeletal muscle, Burch and colleagues, were
amongst the first to note that cachectic cancer patients exhibited
evidence of diminished cardiac mass (9). How cancer cachexia may affect AMPK
and PI3K/Akt/mTOR signaling in the heart has, to our knowledge, not
been reported. We hypothesized that hearts from cachectic
ApcMin/+ mice would weigh less than that observed
in age-matched control animals and that this response would be
associated with decreased anabolic signaling related to the
phosphorylation (activation) of AMPK and PI3K/Akt/mTOR signaling.
Taken together, our data suggest that loss of cardiac mass during
the progression of cachexia in the ApcMin/+ mouse
is associated with a diminished rate of protein synthesis that is
characterized by suppressed mTOR activity and increases in AMPK
phosphorylation.
Materials and methods
Animals
All procedures were performed as outlined in the
Guide for the Animal Use Review Board at the University of South
Carolina. Young (12-week-old, n=6) and adult (20-week-old, n=6)
male C57BL/6 and ApcMin/+ mice were originally
purchased from Jackson Laboratories and bred at the University of
South Carolina’s Animal Resource Facility as previously described
(10). Animals were housed under a
12/12-h dark-light cycle at 22±2°C. Mice were fed with standard
rodent chow (Harlan Teklad Rodent Diet, #8604, Madison, WI) and
water was provided ad libitum. After acclimatization the
mice were sacrificed according to the regulatory guidelines of
University of South Carolina and the samples processed for further
analysis.
Materials
Anti-Akt (#9272), mTOR (#2972), S6 ribosomal protein
(#2217), AMPK-α (#2532), Beclin1 (#3738), phosphorylated Akt Thr308
(#9275), phosphorylated Akt Ser473 (#9271), phosphorylated mTOR
Ser2448 (#2971), phosphorylated S6 ribosomal protein Ser235/236
(#4858), phosphorylated AMPK-α Thr172 (#2535), phosphorylated Bad
(#9295), mouse IgG and rabbit IgG antibodies were purchased from
Cell Signaling Technology (Beverly, MA). HeLa whole cell lysate
(sc-2200) and L6 + IGF lysate (sc-24727) were purchased from Santa
Cruz Biotechnology (Santa Cruz, CA). Enhanced chemiluminescence
(ECL) western blotting detection reagent was purchased from
Amersham Biosciences (Piscataway, NJ). Restore western blot
stripping buffer was obtained from Pierce (Rockford, IL). All other
chemicals were purchased from Fisher Scientific (Hanover, IL) or
Sigma Aldrich (St. Louis, MO).
Preparation of protein samples and
immunoblotting
Hearts were pulverized using a mortar and pestle in
liquid nitrogen until a fine pellet was obtained. Samples were
lysed on ice for 15 min in T-PER buffer (2 ml/g tissue weight;
Pierce, Rockford, IL) containing protease and phosphatase
inhibitors to prevent protein degradation followed by
centrifugation for 10 min at 2000 × g to remove particulate matter.
This process was repeated twice and the supernatants were collected
for the determination of protein concentration using the 660-nm
assay (Pierce). Samples were diluted to a final concentration of
2.5 μg/μl in SDS loading buffer, boiled for 5 min at
95°C and 30 μg of protein were separated using 10 or 15%
SDS-PAGE gels. Protein detection subsequent to transfer to
nitrocellulose membranes were performed as outlined by the antibody
manufacturer and visualized using ECL (Amersham Biosciences). Time
of exposure at all times was adjusted accordingly to keep the
integrated optical densities within a linear and non-saturated
range. The intensity of band signal was quantified by imaging
software (AlphaEase, FC). GAPDH was used to determine equal loading
of protein between lanes and membranes. For direct comparisons
between the concentration levels of different signaling molecules,
membranes were stripped and re-probed using Restore western blot
stripping buffer as detailed by the manufacturer (Pierce).
Myofibrillar protein synthesis
The rate of heart myofibrillar protein synthesis in
an additional set of 20-week-old male C57BL/6 and
ApcMin/+ mice (n=6–7) was measured as previously
described for skeletal muscle with the following modifications
(11,12). Thirty minutes prior to sacrifice,
all mice received an intraperitoneal injection of 150 mM
2H5-phenylalanine (Cambridge Isotopes, MA) in
a 75-mM NaCl solution at a dose of 0.02 ml/g body weight. After
anesthesia (ketaminexylazine-acepromazine cocktail; 1.4 ml/kg body
weight s.c.), whole hearts were excised, rinsed in PBS, quickly
weighed, snap frozen in liquid nitrogen and stored at −80°C until
further analysis. Hearts were homogenized in ice-cold homogenizing
buffer (HB: 50 mM KPO4, pH 7.0, 0.25 M sucrose, 1%
Triton X-100) and myofibrillar proteins were pelleted by
centrifugation. After collection of the supernatants containing
free amino acids, myofibrillar proteins were acid hydrolysed and
the ratio of 2H5-phenylalanine to naturally
occurring phenylalanine in the myofibrillar proteins and amino acid
pool was determined by tandem mass spectrometry (ultra-pressure
liquid chromatography-MS/MS), ESI positive mode, by monitoring ions
m/z 125 and 120, respectively. Percentage protein synthesis/day was
calculated by dividing the ratio of
2H5-phenylalanine/phenylalanine in the
myofibrillar protein pellet by the ratio of
2H5-phenylalanine/phenylalanine in the free
amino acid pool taking into account that
2H5-phenylalanine was incorporated over a
period of 30 min.
Statistical analysis
Results are expressed as mean ± SEM. Data were
analyzed with SigmaStat 3.5 statistical software using a two-way
ANOVA followed by the Student-Newman-Keuls post hoc testing
where appropriate. P<0.05 was considered to be statistically
significant.
Results
Cancer cachexia and heart weight
Average body mass of the C57BL/6 mice was ∼10 and
26% greater (25.7±1.4 vs. 23.1±1.1 g and 27.3±0.3 vs. 20.1±0.8 g)
than that observed in the ApcMin/+ mice at 12 and
20 weeks, respectively. Compared to age-matched controls, heart
muscle mass was ∼8 and 6% less (120.4±3.9 vs. 111.2±3.6 mg and
115.8±3.8 vs. 108.8±5.9 mg) in the 12- and 20-week
ApcMin/+ mice.
Decreased protein synthesis rates in the
hearts of ApcMin/+ mouse is associated with
higher AMPKα and diminished mTOR phosphorylation
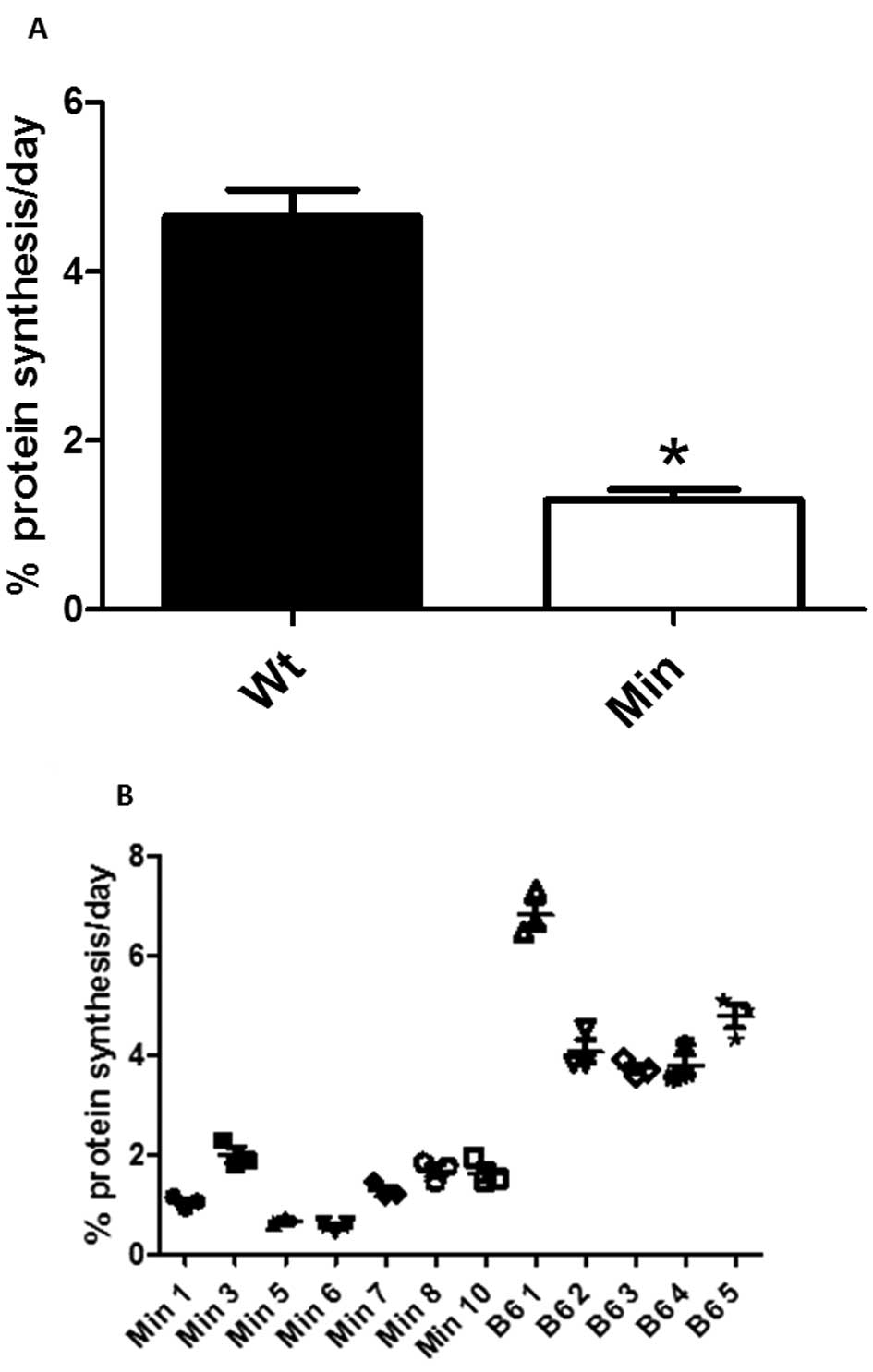
Compared to age-matched C57BL/6 mice, the rate of
myofibrillar protein synthesis as determined by tandem mass
spectrometry was less in the 20-week-old ApcMin/+ mice
(Fig. 1). To investigate the
effects of cancer cachexia on potential regulators of protein
synthesis, we next examined the cardiac lysates for changes in the
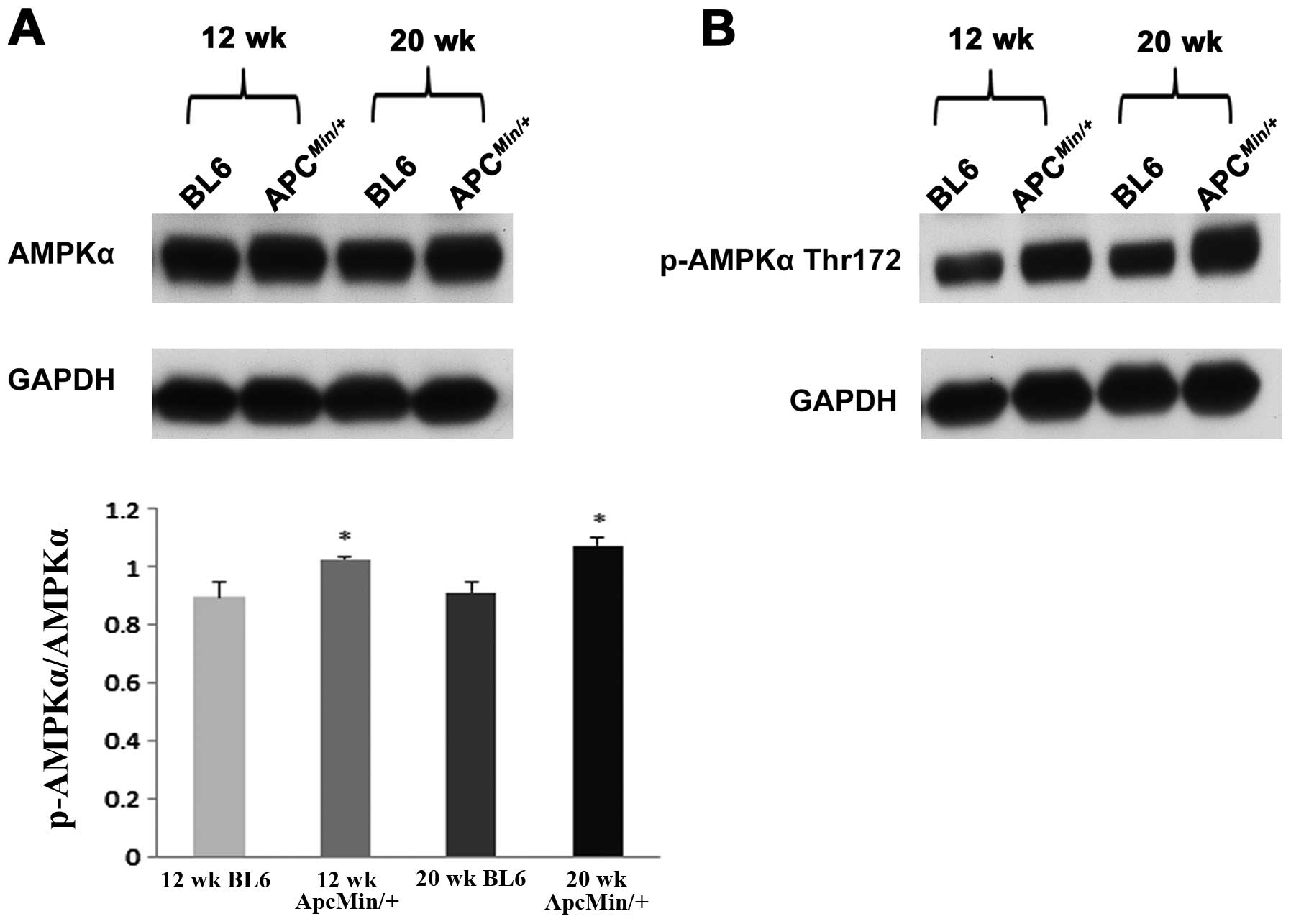
phosphorylation of AMPKα, Akt and mTOR. Compared to that observed
in the control animals, the ratio of phosphorylated to total levels
of AMPKα were ∼14 and ∼17% higher in the 12- and 20-week-old
ApcMin/+ mice, respectively (P<0.05; Fig. 2). Conversely, there was no
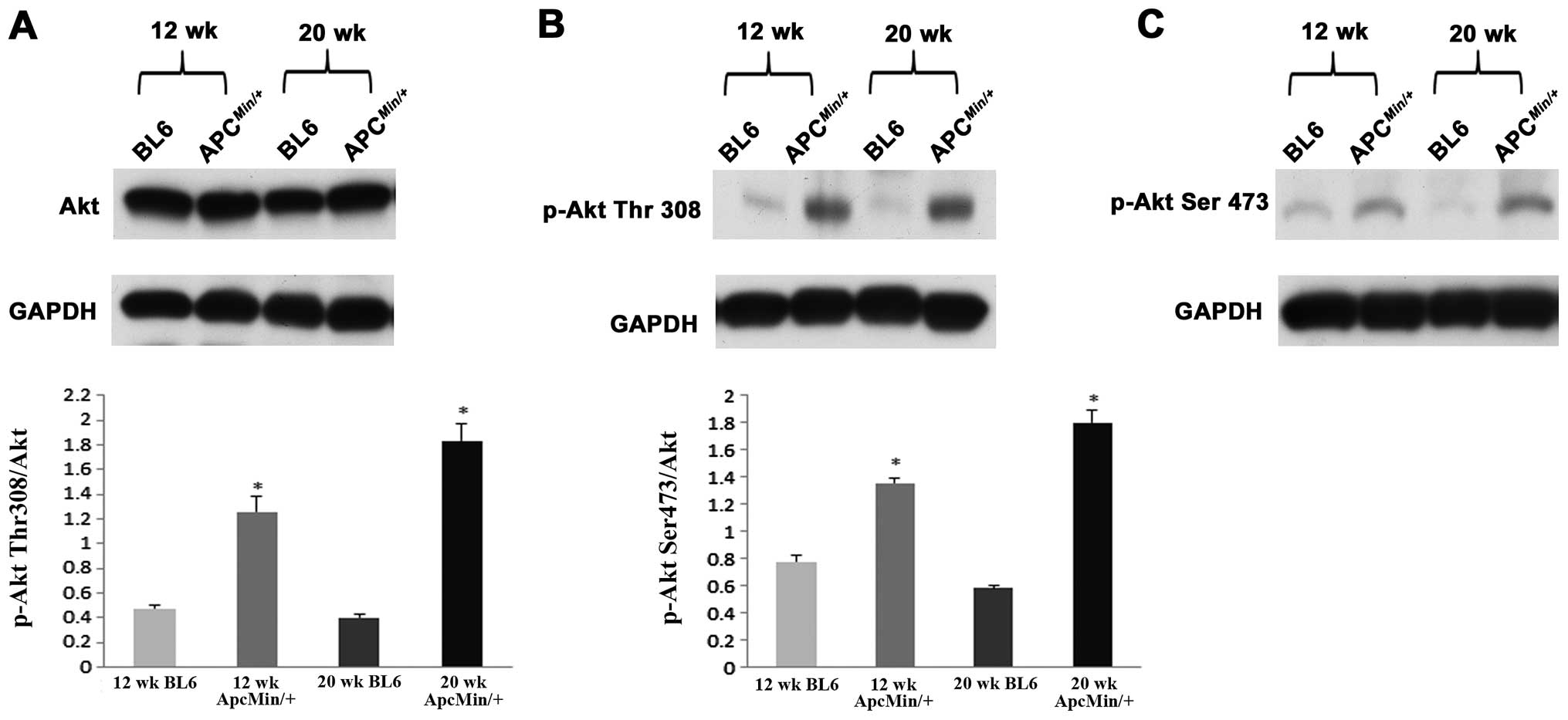
difference in Akt protein content among different groups (Fig. 3). Compared to C57BL/6 controls, the
phosphorylated to total levels of Akt (Ser308) was ∼2.6- and
∼4.5-fold higher in the 12- and 20-week-old
ApcMin/+ mice, respectively (P<0.05; Fig. 3). Similarly, with cancer the amount
of phosphorylated to total levels of Akt (Thr473) was ∼1.75- and
∼3.1-fold higher in 12- and 20-week-old ApcMin/+
mice, respectively (P<0.05; Fig.
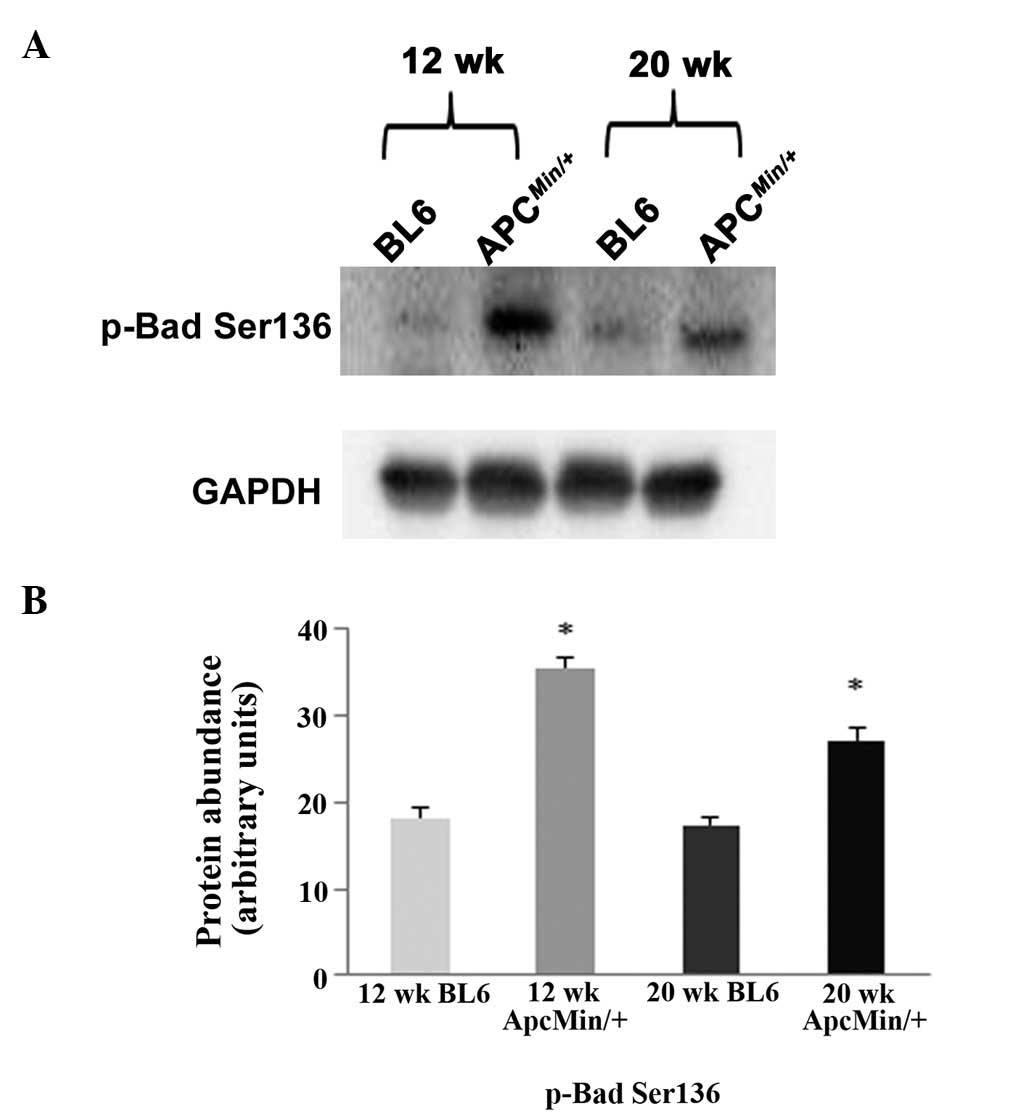
3). The phosphorylation of Bad (Ser136) was ∼100 and ∼74%
higher in the 12-and 20-week-old ApcMin/+ mice,
respectively (P<0.05; Fig. 4).
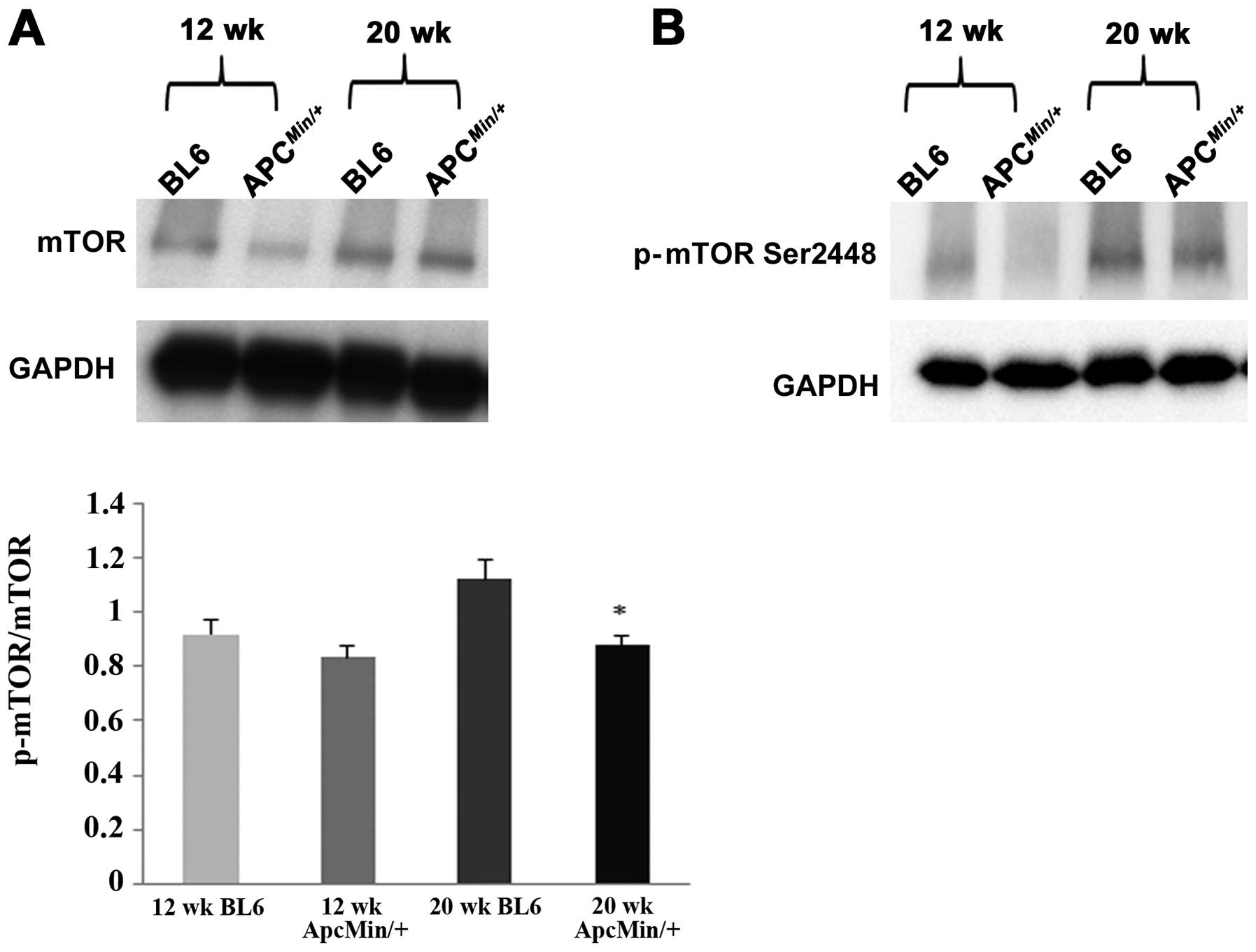
Compared to age-matched C57BL/6 mice, the phosphorylated mTOR
expression was slightly decreased by ∼9% in 12-week-old
ApcMin/+ mice. However, at 20 weeks of age
phosphorylated mTOR expression was reduced by ∼21% (P<0.05;
Fig. 5), demonstrating a
suppression of cardiac mTOR signaling during the progression of
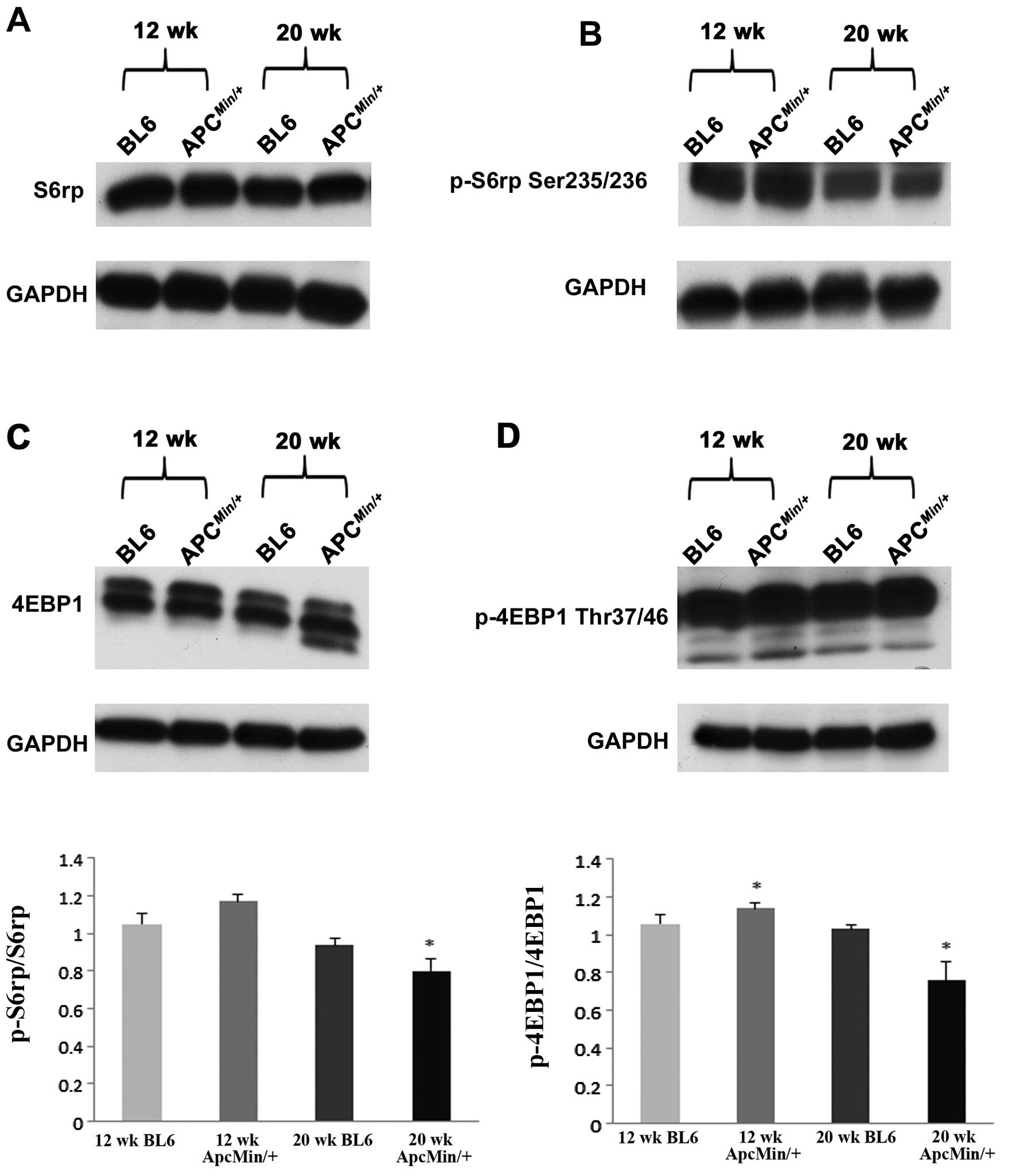
cachexia. Similarly, the ratio of phosphorylated to total level of
S6rp was ∼14% lower in the hearts of 20-week-old
ApcMin/+ mice (P<0.05; Fig. 6). Further analysis revealed that
the ratio of phosphorylated to total 4EBP1 was 8% higher in the
hearts of 12-week-old ApcMin/+ mice and ∼26%
lower in the hearts of 20-week-old ApcMin/+ mice
when compared to age-matched controls (P<0.05; Fig. 6).
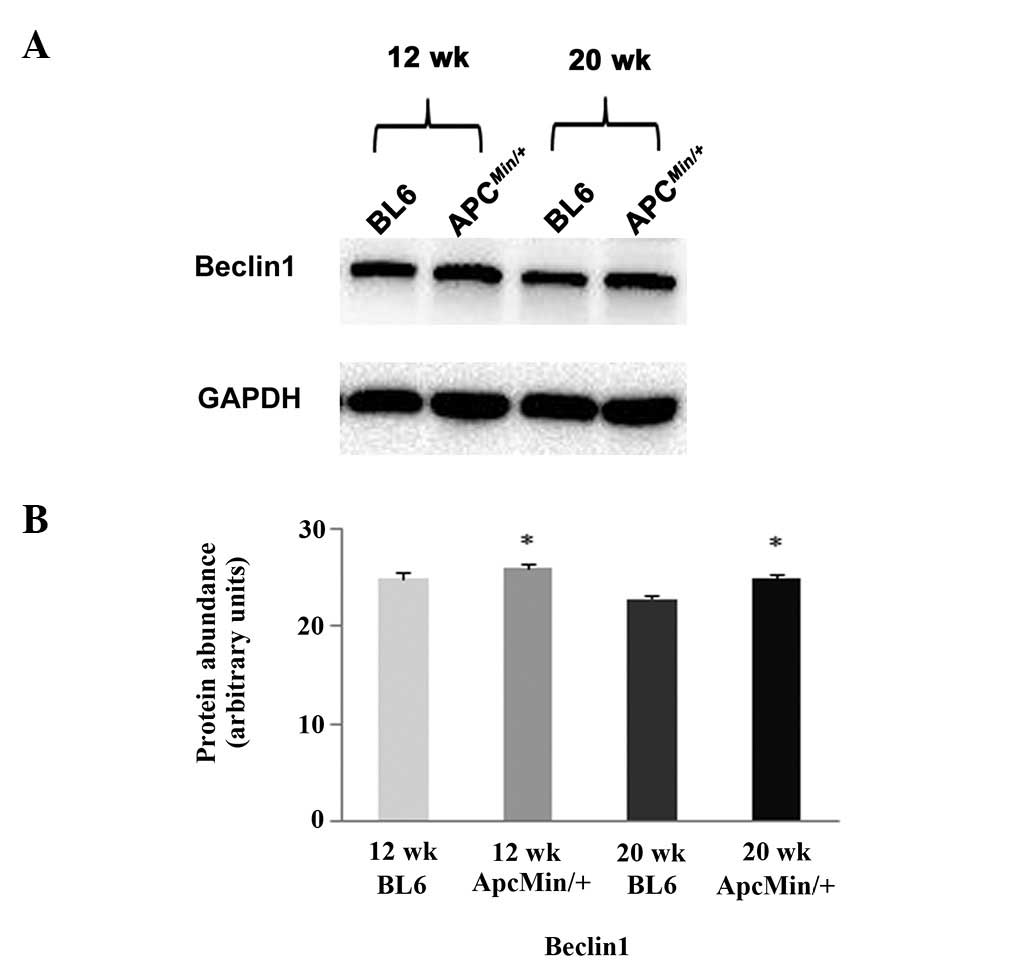
Cachexia increases the expression of
Beclin1 but does not alter protein ubiquitination or involve the
activation of caspase-3
It is thought that muscle utilizes three different
proteolytic pathways to regulate muscle protein breakdown:
calcium-dependent calpains, the ubiquitin-proteasome system and
autophagy (13). Compared to that
observed in the control animals, no evidence of increased protein
ubiquitination, increased caspase-3 activation or apoptosis with
cancer was observed (data not shown). Conversely, immunoblot
analysis revealed that the expression of the autophagy regulator
Beclin1 was ∼5 and ∼10% higher in the hearts of the 12- and
20-week-old ApcMin/+ mice, respectively
(P<0.05; Fig. 7).
Discussion
Cancer cachexia is a life endangering paraneoplastic
syndrome that can lead to the loss of muscle mass, decreased body
weight, fatigue and generalized weakness that is thought to be
responsible for nearly one-third of cancer deaths (13). How cancer may affect cardiac mass
is not well understood. Heart weight in the
ApcMin/+ mouse colorectal cancer model was
decreased by 8 and 6% at 12 and 20 weeks when compared to
age-matched control animals. This loss in muscle mass was much
smaller than that observed in the gastrocnemius (−41%) and soleus
(−26%) in the 20-week-old ApcMin/+ mouse
suggesting that muscle atrophy with cachexia may be regulated
differently in different muscle types (11). In addition, it is likely that the
degree of cardiac muscle loss may differ between cancer models. For
example, using the Yoshida tumor implant model, Costelli et
al demonstrated significant decreases in cardiac mass within
six to ten days of implant (14).
More recently, Cosper and Leinwand using colon-26 adenocarcinoma
implants, showed that cardiac mass was decreased by 8 and 21% at 15
and 27 days post-implantation, respectively (13). The reason why different cancer
models may affect the regulation of heart mass differently is not
clear but could be related to the amount or type of humoral factors
released by the tumor. For example, the muscle wasting seen in the
Yoshida hepatoma ascites model is generally considered to be the
result of elevations in TNF-α while muscle loss in the colon-26
adenocarcinoma tumor implant model has been shown to be associated
with increases in the amount of IL-1, IL-6, IL-12 and TNF-α
(15–17). Whether differences in the type or
amount of circulating mediators can, by themselves, entirely
account for the differences in cardiac muscle loss between models
remains to be determined.
In the case of the ApcMin/+ mouse
model, it is generally thought that the wasting induced in this
model can be explained, at least in part, due to an elevations in
serum IL-6 levels (3,11,18).
How IL-6 may affect cardiac structure or function is poorly
understood. Like that seen in the ApcMin/+ mouse,
recent data have suggested that burn injury and sepsis are also
characterized by increases in IL-6 levels which, at least in these
models, is associated with cardiac inflammation, increased
apoptosis, and depressed contractile function (19,20).
Others have demonstrated that increased IL-6 levels are associated
with diminished muscle protein synthesis (21) and AMPK activation (22). Our findings of diminished cardiac
protein synthesis rates (Fig. 1)
and evidence of increased AMPKα phosphorylation in both the 12- and
20-week-old ApcMin/+ mice support this contention
(Fig. 2). AMPK activation can act
as a molecular switch that turns on energy conserving pathways
while shutting down the energy consuming pathways such as protein
synthesis (23). Whether the
increase in AMPKα we observed is due to elevations in IL-6, changes
in nutrient uptake secondary to colon tumor load, or some
combination of the two is currently unclear.
In an effort to extend our understanding of how
cachexia might affect translational signaling in the
ApcMin/+ mouse, we next examined whether changes
in cardiac mass were associated with differences in the
phosphorylation of Akt and mTOR. Akt is a serine/threonine kinase
which is involved in the regulation of cell metabolism and cell
death (24). With cancer, we found
an elevation in Akt phosphorylation at both Thr308 and Ser473
residues which is consistent with notion of Akt activation
(Fig. 3). To explore whether the
observed Akt activation with cachexia might be anti-apoptotic in
nature, we examined the effect of cachexia on the phosphorylation
of Bad. Like Akt, Bad is intimately involved in regulating the
transition between cellular life and death. In its unphosphorylated
form Bad is strongly apoptotic (25,26).
Conversely, when phosphorylated at Ser136 by Akt, the apoptotic
activity of Bad is inhibited and cell death is prohibited. Compared
to that seen in the control animals, the phosphorylation of Bad
(Ser136) was significantly higher in the ApcMin/+
mouse hearts (Fig. 4). Given that
diminished Akt signaling may also be involved in increasing
proteasome activity during skeletal muscle atrophy (27), we next examined if cardiac muscle
loss was associated with increases in protein ubiquitination. As
expected from our findings of increased Akt phosphorylation and
similar to that others have seen investigating cardiac atrophy in
the C-26 adenocarcinoma tumor implant model (13) we did not find any evidence of
increased protein ubiquitination or cardiac apoptosis (data not
shown). Taken together, these data suggest that the activation of
Akt in the cachectic heart may be a compensatory response initiated
to minimize the apoptotic effects of circulating humoral factors,
e.g., IL-6 and TNF-α, to preserve cardiac mass and function.
Similar to Akt, the activation of mTOR is required
for cell survival (28). In
addition to its effects on cellular survival, the activation
(phosphorylation) of mTOR and its substrates are required for the
induction of protein synthesis (29). Unlike that observed by us for Akt,
we found significant decrease in the ratio of phosphorylated to
total mTOR with cachexia (Fig. 5).
To confirm these findings, we next examined the phosphorylation of
downstream mTOR substrates, S6rp and 4EBP1. As expected from our
assessment of mTOR phosphorylation and our findings of diminished
protein synthesis rates, we observed that the amount of
phosphorylated (active) S6rp and 4EBP1 were both diminished with
cachexia (Fig. 6). This finding is
consistent with the recent study of White and colleagues who
demonstrated diminished myofibrillar synthesis rates in the
skeletal muscles of the ApcMin/+ mouse at similar
time-points (10).
Autophagy is a mechanism for cell protection in
which intracellular substances are degraded and the subsequent
products are recycled again (30).
On the basis of recent data suggesting that cancer cachexia is
associated with increased autophagy (13) and other findings demonstrating that
increased AMPK phosphorylation can induce the expression of the
autophagy proteins (7) we next
examined if cachexia in the ApcMin/+ mice was
associated with increases in autophagy protein Beclin1. Consistent
with our findings of decreased cardiac mass, we observed that
cancer cachexia in the ApcMin/+ mice was
associated with slightly elevated levels of Beclin1 (Fig. 7). These findings suggest that
changes in cardiac mass in the ApcMin/+ mouse may
be associated with autophagy.
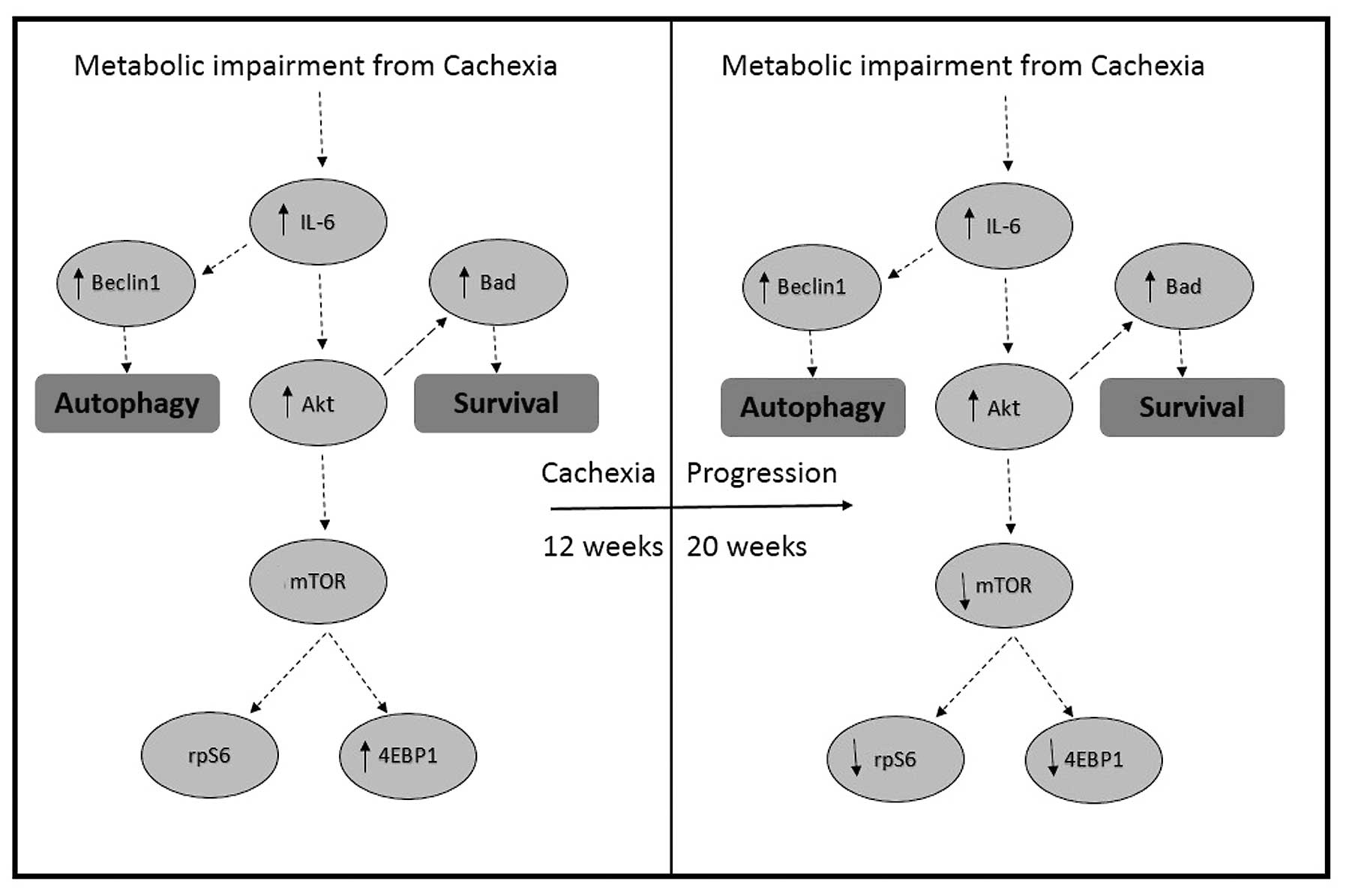
Taken together, our data demonstrate that cachexia
in the ApcMin/+ mouse model is associated with a
mild form of cardiac atrophy that may be caused by decreases in the
rate of myofibrillar protein synthesis and the suppression of
anabolic signaling through mTOR. We hypothesize that this decrease
in protein synthesis is accompanied by increases in cardiac
autophagy as suggested by our findings of increased AMPK
phosphorylation and Beclin1 protein expression. We suspect that the
activation of Akt and Bad observed in the present study are more
related to efforts directed at maintaining cell survival rather
than increasing protein synthesis (Fig. 8). Future studies examining tumor
humoral factors or mediators of systemic inflammation will no doubt
be useful in furthering our understanding of how colorectal cancer
may affect cardiac structure and function.
Acknowledgements
This study was supported from NIH
grant RO1CA121249 (to J.A.C), DOE grant (DE-PS02-09ER-01 to E.R.B).
The authors would like to thank John W. Baynes and Suichi Sato for
their technical input related to the heart protein synthesis
measurements.
References
|
1
|
Labianca R and Merelli B: Screening and
diagnosis for colorectal cancer: present and future. Tumori.
96:889–901. 2010.PubMed/NCBI
|
|
2
|
Moser AR, Luongo C, Gould KA, McNeley MK,
Shoemaker AR and Dove WF: ApcMin: a mouse model for intestinal and
mammary tumorigenesis. Eur J Cancer. 31A:1061–1064. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baltgalvis KA, Berger FG, Pena MM, Davis
JM, Muga SJ and Carson JA: Interleukin-6 and cachexia in
ApcMin/+ mice. Am J Physiol Regul Integr Comp Physiol.
294:R393–R401. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
McClellan JL, Davis JM, Steiner JL, et al:
Intestinal inflammatory cytokine response in relation to
tumorigenesis in the Apc(Min/+) mouse. Cytokine. 57:113–119.
2012.PubMed/NCBI
|
|
5
|
Argiles JM, Busquets S, Garcia-Martinez C
and Lopez-Soriano FJ: Mediators involved in the cancer
anorexia-cachexia syndrome: past, present, and future. Nutrition.
21:977–985. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schiaffino S and Mammucari C: Regulation
of skeletal muscle growth by the IGF1-Akt/PKB pathway: insights
from genetic models. Skelet Muscle. 1:42011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Egan D, Kim J, Shaw RJ and Guan KL: The
autophagy initiating kinase ULK1 is regulated via opposing
phosphorylation by AMPK and mTOR. Autophagy. 7:643–644. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hahn-Windgassen A, Nogueira V, Chen CC,
Skeen JE, Sonenberg N and Hay N: Akt activates the mammalian target
of rapamycin by regulating cellular ATP level and AMPK activity. J
Biol Chem. 280:32081–32089. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Burch GE, Phillips JH and Ansari A: The
cachetic heart. A clinico-pathologic, electrocardiographic and
roentgenographic entity. Dis Chest. 54:403–409. 1968. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
White JP, Baltgalvis KA, Puppa MJ, Sato S,
Baynes JW and Carson JA: Muscle oxidative capacity during
IL-6-dependent cancer cachexia. Am J Physiol Regul Integr Comp
Physiol. 300:R201–R211. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
White JP, Baynes JW, Welle SL, et al: The
regulation of skeletal muscle protein turnover during the
progression of cancer cachexia in the Apc(Min/+) mouse. PLoS One.
6:e246502011.
|
|
12
|
Lima M, Sato S, Enos RT, Baynes JW and
Carson JA: Development of an UPLC-mass spectrometry method for
measurement of myofibrillar protein synthesis: analysis of murine
muscles during cancer cachexia. J Appl Physiol. 114:824–828. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cosper PF and Leinwand LA: Cancer causes
cardiac atrophy and autophagy in a sexually dimorphic manner.
Cancer Res. 71:1710–1720. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Costelli P, Tullio RD, Baccino FM and
Melloni E: Activation of Ca(2+)-dependent proteolysis in skeletal
muscle and heart in cancer cachexia. Br J Cancer. 84:946–950.
2001.
|
|
15
|
Fujita J, Tsujinaka T, Yano M, et al:
Anti-interleukin-6 receptor antibody prevents muscle atrophy in
colon-26 adenocarcinoma-bearing mice with modulation of lysosomal
and ATP-ubiquitin-dependent proteolytic pathways. Int J Cancer.
68:637–643. 1996. View Article : Google Scholar
|
|
16
|
Strassmann G, Masui Y, Chizzonite R and
Fong M: Mechanisms of experimental cancer cachexia. Local
involvement of IL-1 in colon-26 tumor. J Immunol. 150:2341–2345.
1993.PubMed/NCBI
|
|
17
|
Wysong A, Couch M, Shadfar S, et al:
NF-kappaB inhibition protects against tumor-induced cardiac atrophy
in vivo. Am J Pathol. 178:1059–1068. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Carson JA and Baltgalvis KA: Interleukin 6
as a key regulator of muscle mass during cachexia. Exerc Sport Sci
Rev. 38:168–176. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mandel ID, Barr CE and Turgeon L:
Longitudinal study of parotid saliva in HIV-1 infection. J Oral
Pathol Med. 21:209–213. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang H, Wang HY, Bassel-Duby R, et al:
Role of interleukin-6 in cardiac inflammation and dysfunction after
burn complicated by sepsis. Am J Physiol Heart Circ Physiol.
292:H2408–H2416. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
van Hall G, Steensberg A, Fischer C, et
al: Interleukin-6 markedly decreases skeletal muscle protein
turnover and increases nonmuscle amino acid utilization in healthy
individuals. J Clin Endocrinol Metab. 93:2851–2858. 2008.PubMed/NCBI
|
|
22
|
Kelly M, Keller C, Avilucea PR, et al:
AMPK activity is diminished in tissues of IL-6 knockout mice: the
effect of exercise. Biochem Biophys Res Commun. 320:449–454. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chan AY and Dyck JR: Activation of
AMP-activated protein kinase (AMPK) inhibits protein synthesis: a
potential strategy to prevent the development of cardiac
hypertrophy. Can J Physiol Pharmacol. 83:24–28. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Latronico MV, Costinean S, Lavitrano ML,
Peschle C and Condorelli G: Regulation of cell size and contractile
function by AKT in cardiomyocytes. Ann NY Acad Sci. 1015:250–260.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang E, Zha J, Jockel J, Boise LH,
Thompson CB and Korsmeyer SJ: Bad, a heterodimeric partner for
Bcl-XL and Bcl-2, displaces Bax and promotes cell death. Cell.
80:285–291. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zha J, Harada H, Yang E, Jockel J and
Korsmeyer SJ: Serine phosphorylation of death agonist BAD in
response to survival factor results in binding to 14-3-3 not
BCL-X(L). Cell. 87:619–628. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kandarian SC and Jackman RW: Intracellular
signaling during skeletal muscle atrophy. Muscle Nerve. 33:155–165.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Martin DE and Hall MN: The expanding TOR
signaling network. Curr Opin Cell Biol. 17:158–166. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang X and Proud CG: The mTOR pathway in
the control of protein synthesis. Physiology. 21:362–369. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Levine B and Yuan J: Autophagy in cell
death: an innocent convict? J Clin Invest. 115:2679–2688. 2005.
View Article : Google Scholar : PubMed/NCBI
|