Introduction
According to the American Cancer Society, about
11,280 new soft tissue sarcomas would be diagnosed in 2012 (6,110
cases in males and 5,170 cases in females) and 3,900 Americans
(2,050 males and 1,850 females) were expected to die of soft tissue
sarcomas (1). The most common
types of sarcoma in adults are malignant fibrous
histiocytoma/fibrosarcoma, liposarcoma (a malignancy of fat cells),
and uterine leiomyosarcoma (1).
Chondrosarcoma, a malignancy of cartilaginous origin, primarily
affects the cartilage cells of femur, arm, pelvis, knee and spine.
Forty percent of primary bone cancers are chondrosarcomas (2). Fibrosarcoma, a rare form of cancer
(4% of bone cancers), is an aggressive and highly metastatic cancer
of the connective tissue that primarily develops in metaphases of
long tubular bones and affects both children and adults (2,3).
Primary cancers of bones account for less than 0.2% of all cancers.
Synovial sarcoma, also a rare soft tissue cancer (accounting for
less than 5–10% of all soft tissue sarcomas) develops in the
synovial membrane of the joints, most frequently occurs in the
lower limbs, but can also occur in the trunk and head/neck
(4). Poor prognosis is attributed
to both the aggressive metastatic spread characteristic of these
cancers and the lack of efficacy in current treatment modalities to
prevent or counteract tumor progression. The overall relative
5-year survival rate of people with soft tissue sarcomas is around
50% according to statistics from the National Cancer Institute
(1). The corresponding 5-year
relative survival rates were: 83% for localized sarcomas (56% of
soft tissue sarcomas were localized when they were diagnosed); 54%
for regional stage sarcomas (19% were in this stage); 16% for
sarcomas with distant spread (16% were in this stage) (1).
Metastasis occurs secondary to cancer cell
detachment from the primary tumor, invasion through degraded
basement membrane into the surrounding stroma, and entry into and
transport through the vascular or lymphatic system to distal sites
such as the liver, lungs and brain, and extravasation, tumor cell
proliferation and angiogenesis at distal sites (5–9).
Tumor cell invasion depends upon degradation of the extracellular
matrix (ECM), which, when intact, acts as a barrier to block cancer
cell invasion. The ECM is composed of collagen, proteoglycans,
fibronectin, laminin and other glycoproteins (10–12).
Two families of proteases, the matrix metalloproteinases (MMPs) and
urokinase plasminogen activators (u-PA) are involved in tumor
invasion and metastasis. Numerous clinical and experimental studies
have demonstrated that elevated levels of u-PA and MMPs are
associated with tumor growth, cancer progression, metastasis and
shortened survival in patients (4,13,14).
MMPs, especially MMP-2 and MMP-9 play key roles in
tumor cell invasion and metastasis due to their ability to degrade
type IV collagen, a major component of the ECM (12,15,16).
MMP-2 and -9 are secreted as inactive pro-enzymes in their latent
zymogenic form, and activated by other MMPs or proteases.
Proteolytic activities of MMP-2 and MMP-9 are inhibited by specific
inhibitors, tissue inhibitors of metalloproteinases (TIMPs). Thus,
a critical determinant of net proteolytic degradation is the
balance between MMP and TIMP levels. Clinical studies note the
association of MMP expression with progression of sarcomas
(4). For example, when Benassi
et al examined biopsied tissue immunohistochemically from
patients with soft tissue sarcomas, he noted that poor prognosis
was significantly correlated with elevated MMP-2 and lack of TIMP-2
expression in all sarcomas studied and that elevated MMP-2 and
MMP-9 levels significantly correlated with metastasis in
liposarcoma (17).
The serine protease u-PA converts plasminogen to
plasmin, which is capable of promoting tumor growth and
angiogenesis, degrading the ECM and basement membrane and
activating pro-MMPs (18).
Components of the u-PA system such as u-PA, plasminogen activator
inhibitor-1 (PAI-1), and urokinase-type plasminogen activator
receptor (u-PAR) are overexpressed in a variety of cancer types,
most notably in breast cancer (19), but also in sarcomas (14), and correlate with cancer
progression, metastasis and poor prognosis. Thus the u-PA system
represents a potential target for anticancer strategies.
Rath and Pauling (20) proposed using nutrients such as
lysine and ascorbic acid to target plasmin-mediated connective
tissue degradation as a universal approach to tumor growth and
expansion. Binding to plasminogen active sites, lysine blocks
plasminogen activation into plasmin by tissue plasminogen activator
(t-PA). Thus it modulates the plasmin-induced MMP activation
cascade (21). Subsequent studies
confirmed this approach and lead to identifying a novel formulation
composed of lysine, ascorbic acid, proline and green tea extract
and other micronutrients (NM), which has shown significant
anticancer activity against a large number (∼40) of cancer cell
lines, blocking cancer growth, tissue invasion and MMP expression
both in vitro and in vivo (22). In this study, we focused on the
modulating effect of NM on the activities of MMP-2 and -9, TIMPs
and u-PA in adult human sarcomas: fibrosarcoma, chondrosarcoma,
liposarcoma, synovial sarcoma and uterine leimyosarcoma cell
lines.
Materials and methods
Materials
Human sarcoma cell lines fibrosarcoma HT-1080 (FS),
chondrosarcoma SW-1353 (CS), liposarcoma SW-872 (LPS), synovial
sarcoma SW-982 (SS) and uterine leiomyosarcoma SK-UT-1 (LS), along
with their culture media were obtained from ATCC (Manassas, VA).
Antibiotics, penicillin and fetal bovine serum (FBS), were obtained
from Gibco-BRL (Long Island, NY). Twenty-four-well tissue culture
plates were obtained from Costar (Cambridge, MA). Gelatinase
zymography was performed in 10% Novex Precast SDS polyacrylamide
gel (Invitrogen) with 0.1% gelatin in non-reducing conditions. The
nutrient mixture (NM), prepared by VitaTech (Hayward, CA) was
composed of the following ingredients in the relative amounts
indicated: vitamin C (as ascorbic acid and as Mg, Ca and palmitate
ascorbate) 700 mg; L-lysine 1,000 mg; L-proline 750 mg; L-arginine
500 mg; N-acetyl cysteine 200 mg; standardized green tea extract
(80% polyphenol) 1,000 mg; selenium 30 μg; copper 2 mg;
manganese 1 mg. All other reagents used were of high quality and
were obtained from Sigma, unless otherwise indicated.
Cell cultures
The sarcoma cell lines were grown in their
respective media: fibrosarcoma in MEM, chondrosarcoma in DEM,
liposarcoma in MEM, synovial sarcoma in DME and uterine
leiomyosarcoma in DEME, supplemented with 10% FBS, penicillin (100
U/ml) and streptomycin (100 μg/ml) in 24-well tissue culture
plates. The cells were plated at a density of 1×105
cells/ml and grown to confluency in a humidified atmosphere at 5%
CO2 at 37°C. Serum-supplemented media were removed and
the cell monolayer was washed once with PBS with the recommended
serum-free media. The cells were treated with the nutrient mixture,
dissolved in media and tested at 0, 50, 100, 250, 500 and 1,000
μg/ml in triplicate at each dose for u-PA and TIMP-2
studies. For MMP analysis, cells were treated with NM at 0, 10, 50,
100, 500 and 1,000 μg/ml. Parallel sets of cultures were
treated with PMA (100 ng/ml) for induction of MMP-9. Control and
PMA treatments were done in triplicates. The plates were then
returned to the incubator. The conditioned media were collected
separately, pooled, and centrifuged at 4°C for 10 min at 3,000 rpm
to remove cells and cell debris. The supernatant was collected and
used to assess for u-PA activity (by fibrin zymography on 10%
SDS-PAGE gels containing fibrinogen and plasminogen), MMP-2 and -9
(by gelatinase zymography), and TIMPs (by reverse zymography).
Fibrin zymography
Fibrin zymography was used to analyze u-PA activity
on 10% SDS-PAGE gels containing fibrinogen (5.5 mg/ml) and
plasminogen (50 μg/ml). After electrophoresis, the gels were
washed twice with 2.5% Triton X-100 for 30 min. The gels were then
incubated overnight at 37°C with 0.1% glycine buffer pH 7.5 and
then stained with 0.5% Coomassie brilliant Blue R250 and destained.
Electrophoresis of u-PA and t-PA were conducted for comparison.
Fibrin zymograms were scanned using CanoScan 9950F Canon
Scanner.
Gelatinase zymography
Gelatinase zymography was performed in 10% Novex
Precast SDS Polyacrylamide Gel (Invitrogen) in the presence of 0.1%
gelatin under non-reducing conditions. Culture media (20 μl)
were mixed with sample buffer and loaded for SDS-PAGE with
Tris-glycine SDS buffer as suggested by the manufacturer (Novex).
Samples were not boiled before electrophoresis. Following
electrophoresis the gels were washed twice in 2.5% Triton X-100 for
30 min at room temperature to remove SDS. The gels were then
incubated at 37°C overnight in substrate buffer containing 50 mM
Tris-HCl and 10 mM CaCl2 at pH 8.0 and stained with 0.5%
Coomassie Blue R250 in 50% methanol and 10% glacial acetic acid for
30 min and destained. Upon renaturation of the enzyme, the
gelatinases digest the gelatin in the gel and give clear bands
against an intensely stained background. Protein standards were run
concurrently and approximate molecular weights were determined by
plotting the relative mobilities of known proteins.
Reverse zymography
TIMPs were analyzed by reverse zymography on 15% SDS
gels containing serum-free conditioned medium from cells. After
electrophoresis the gels were washed twice with 2.5% Triton X-100
for 30 min at room temperature to remove SDS. The gels were then
incubated at 37°C overnight in 50 mM Tris-HCl and 10 mM
CaCl2 at pH 7.6 and stained with 0.5% Coomassie Blue
R25, destained and scanned.
Scanning of gelatinase, reverse and
fibrin zymograms
Gelatinase, reverse and fibrin zymograms were
scanned using CanoScan 9950F Canon scanner at 300 dpi. The
intensity of the bands was evaluated using the pixel-based
densitometer program Un-Scan-It, Version 5.1, 32-bit, by Silk
Scientific Corporation (Orem, UT), at a resolution of 1 Scanner
Unit (1/100 of an inch for an image that was scanned at 100 dpi).
The pixel densitometer calculates the optical density of each pixel
(values 0 to 255) using the darkly stained background of the gel as
a pixel value of 0. A logarithmic optical density scale was used
since the optical density of film and gels is logarithmically
proportional to the concentration. The pixel densitometer sums the
optical density of each pixel to give the band density.
Statistical analysis
Pearson's correlation coefficient was determined
between NM effect on mean MMP-2 or MMP-9, u-PA and TIMP-2
expressions of sarcoma cell lines using MedCalc Software
(Mariakerke, Belgium).
Results
Table I provides an
overview of the u-PA, MMP and TIMP-2 activity in the tested sarcoma
cell line.
 | Table I.Overview of MMP-2 and -9, u-PA and
TIMP-2 expression of adult sarcoma cell lines. |
Table I.
Overview of MMP-2 and -9, u-PA and
TIMP-2 expression of adult sarcoma cell lines.
| Cancer cell
line | MMP-2 | MMP-9 | u-PA | TIMP-2 |
|---|
| Fibrosarcoma
HT-1080 | + | + | + | + |
| Chondrosarcoma
SW-1353 | + | + | + | + |
| Liposarcoma
SW-872 | + | + | + | + |
| Synovial sarcoma
SW-982 | + | + | − | + |
| Uterine
leiomyosarcoma SK-UT-1 | − | + | + | + |
Effect of NM on u-PA activity in human
adult sarcoma cell lines
Activity of u-PA was detected in fibrosarcoma,
chondrosarcoma, liposarcoma and uterine leiomyosarcoma cell lines.
CS and LS showed 2 bands corresponding to subunits 1 and 2 at 55
and 33 kD, while FS and LPS showed one band corresponding to
subunit 1 (55 kD). However, no bands corresponding to u-PA were
detected for synovial sarcoma cell line. NM exerted dose response
inhibition with virtual block of u-PA activity at 50 μg/ml
in LPS (linear trend R2=0.429), 100 μg/ml in CS
(linear trends R2=0.690 and 0.698 for subunits 1 and 2,
respectively) and LS (linear trends R2=0.461 and 0.429
for subunits 1 and 2, respectively) and 250 μg/ml in FS
(linear trend R2=0.659). See Fig. 1 for respective fibrin zymograms and
densitometry analyses.
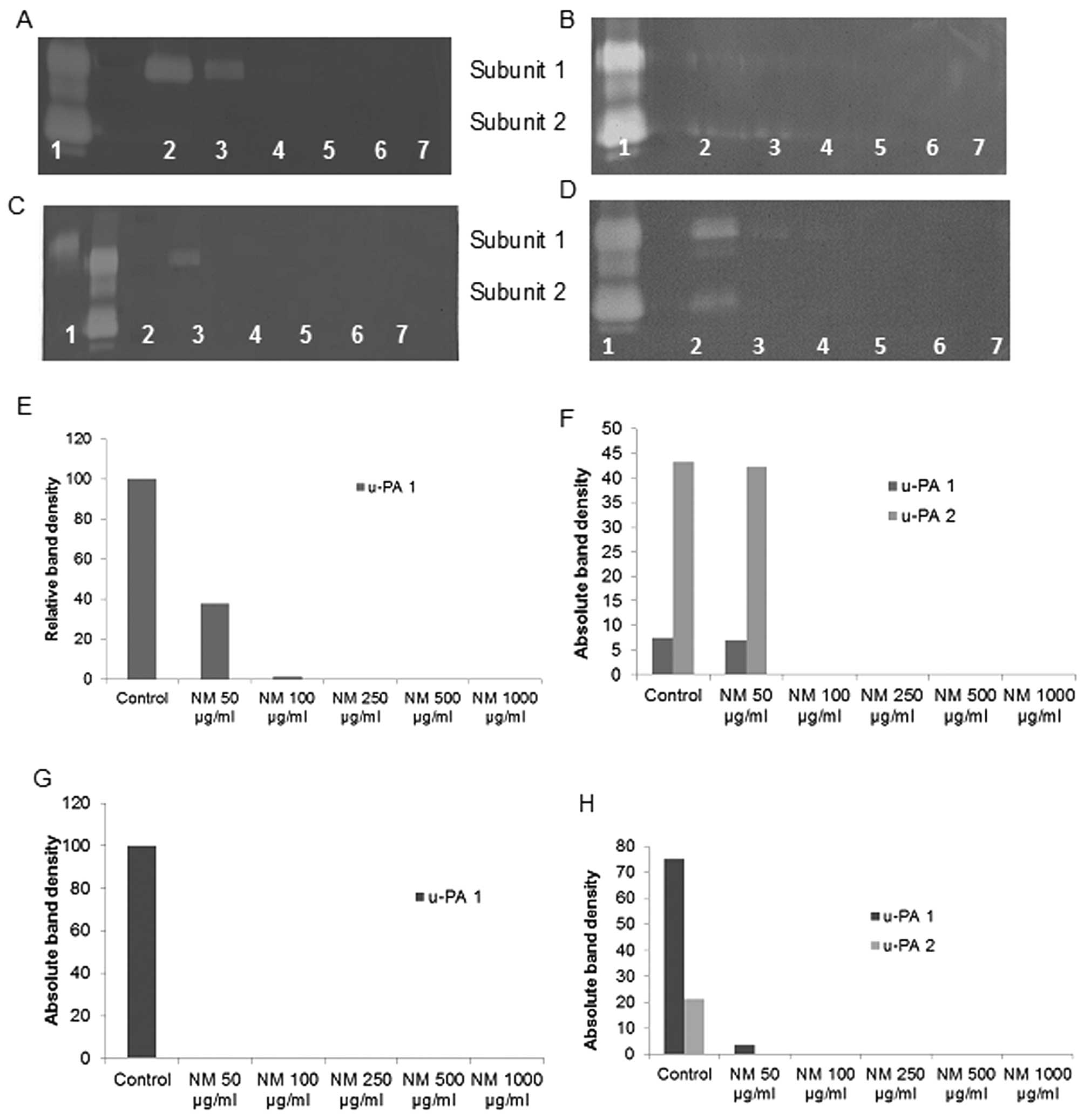 | Figure 1.Effect of NM on fibrosarcoma HT-1080,
chondrosarcoma SW-1353, liposarcoma SW-872 and leiomyosarcoma
SK-UT-1 u-PA expression. Fibrin zymo-grams of (A) HT-1080, (B)
SW-1353, (C) SW-872 and (D) SK-UT-1 u-PA expression. Lane 1, u-PA;
lane 2, markers; lane 3, control; lanes 4–8, NM 50, 100, 250, 500
and 1,000 μg/ml. Densitometric analyses of (E) HT-1080, (F)
SW-1353, (G) SW-872 and (H) SK-UT-1 u-PA expression. |
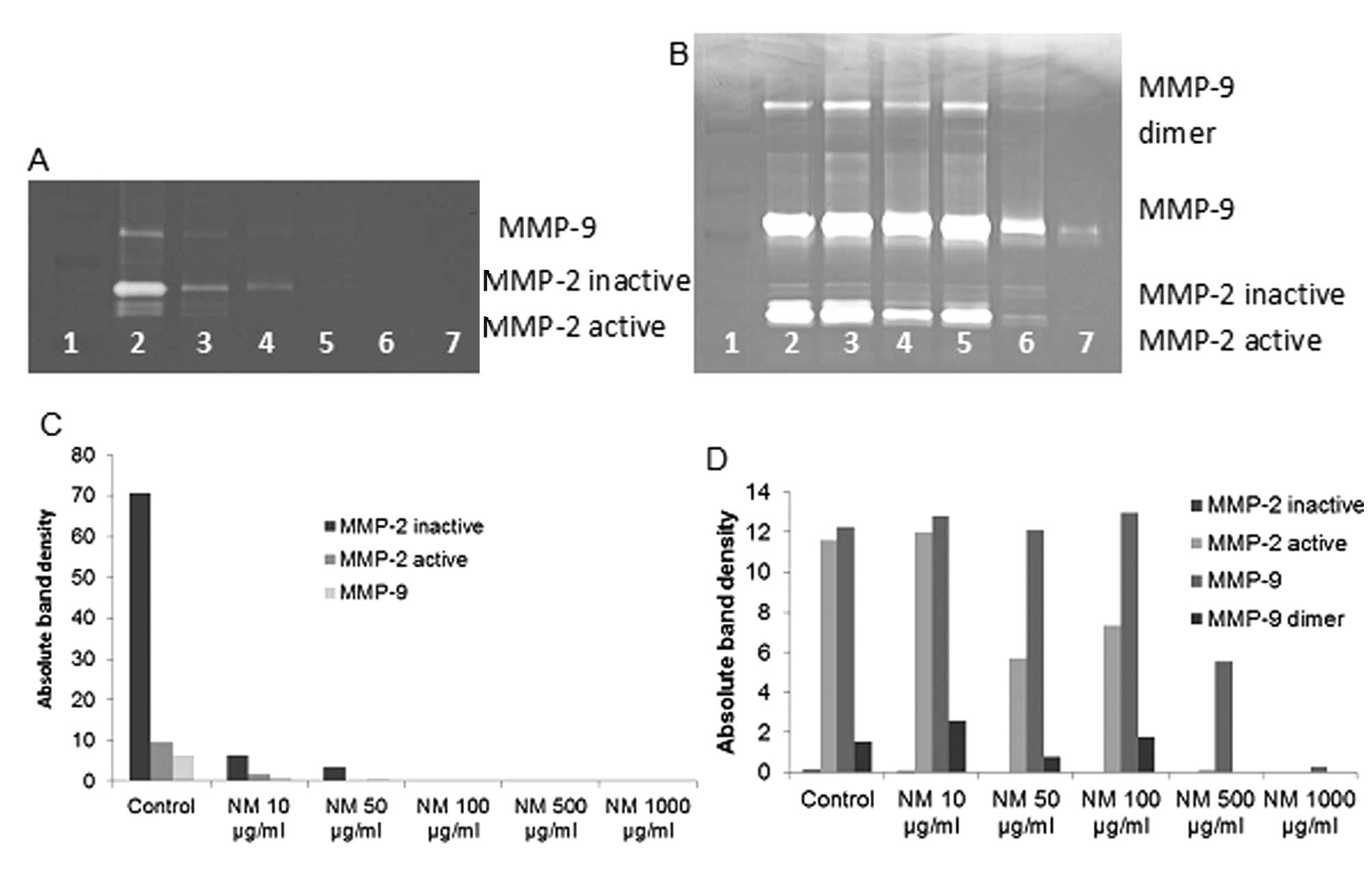
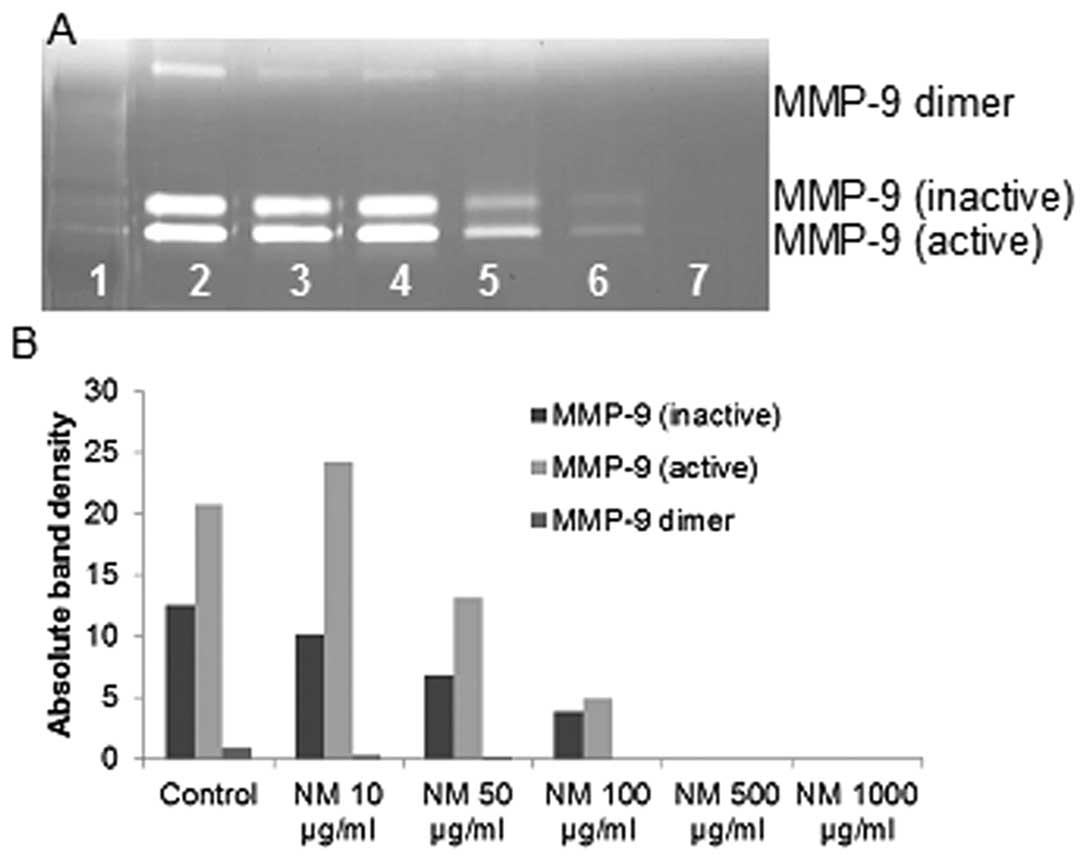
Effect of NM on MMP-2 and MMP-9
expression by fibrosarcoma cell line HT-1080
On gelatinase zymography, FS cells demonstrated
strong expression of MMP-2 inactive and faint MMP-2 active, both
greater than MMP-9, which were inhibited by NM in a dose-dependent
fashion with virtual total inhibition of MMP-9 at 100 μg/ml
(linear trend R2=0.546) and MMP-2 at 250 μg/ml
(linear trend R2=0.510). PMA (100 ng/ml) treatment
enhanced MMP-2 active and MMP-9 expression by HT-108-cells; NM
inhibited MMP-2 and MMP-9 in a dose-dependent manner with total
block of MMP-2 active at 500 μg/ml (linear trend
R2=0.865) and near virtual block of MMP-9 at 1,000
μg/ml (linear trend R2=0.675). See Fig. 2 for gelatinase zymograms and
densitometry analyses.
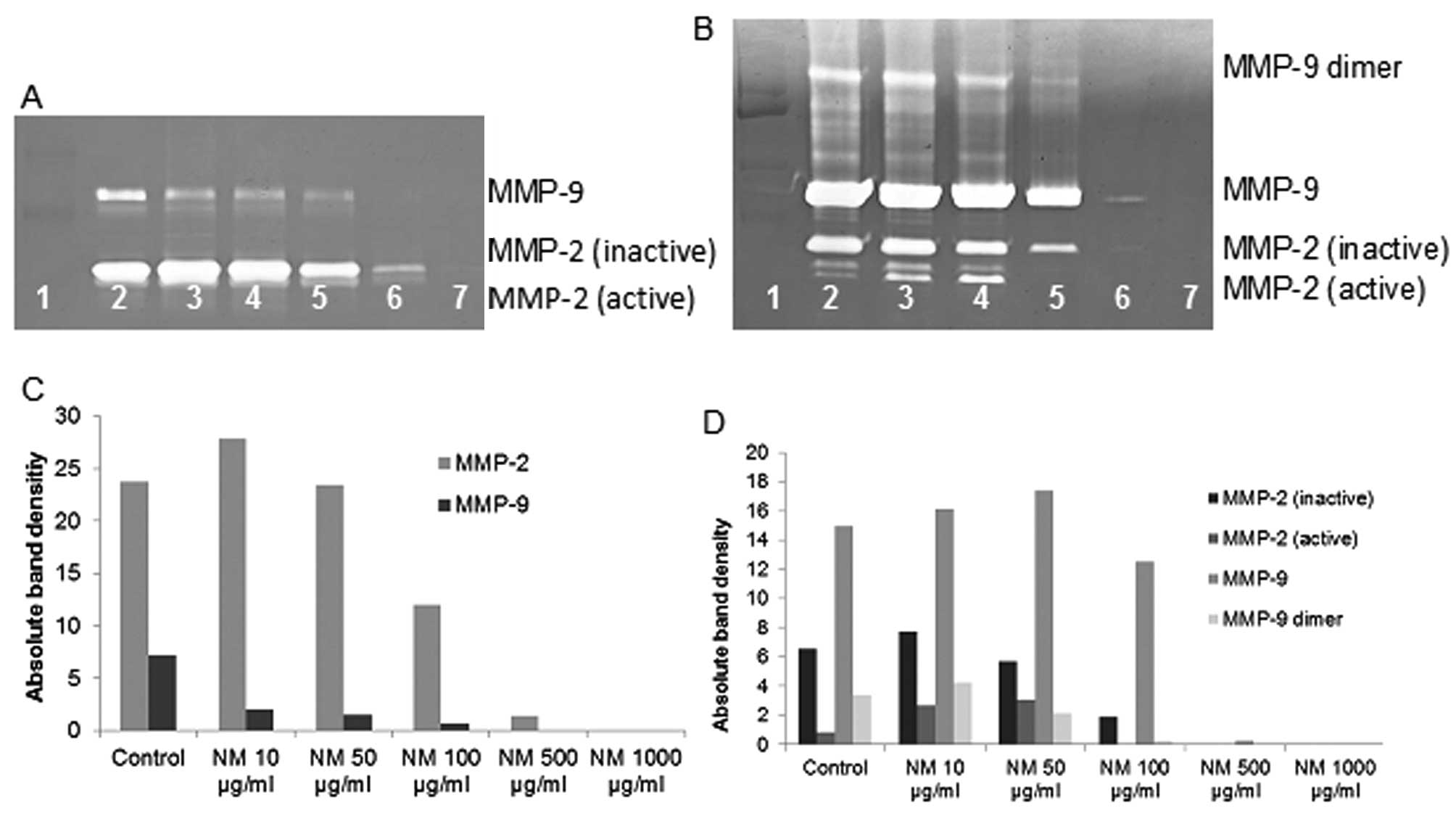
Effect of NM on MMP-2 and MMP-9
expression by chondrosarcoma cell line SW-1353
On gelatinase zymography, CS cells demonstrated
strong expression of MMP-2 and slight expression of MMP-9, which
were inhibited by NM in a dose-dependent fashion with virtual total
inhibition of MMP-9 at 500 μg/ml (linear trend
R2=0.713) and MMP-2 at 1,000 μg/ml (linear trend
R2=0.860). PMA (100 ng/ml) treatment profoundly enhanced
MMP-9 expression by SW-1353 cells; NM inhibited MMP-2 and MMP-9 in
a dose-dependent manner with total block of MMP-2 at 500
μg/ml (linear trend R2=0.866) and MMP-9 at 500
μg/ml (linear trend R2=0.729). See Fig. 3 for gelatinase zymograms and
densitometry analyses.
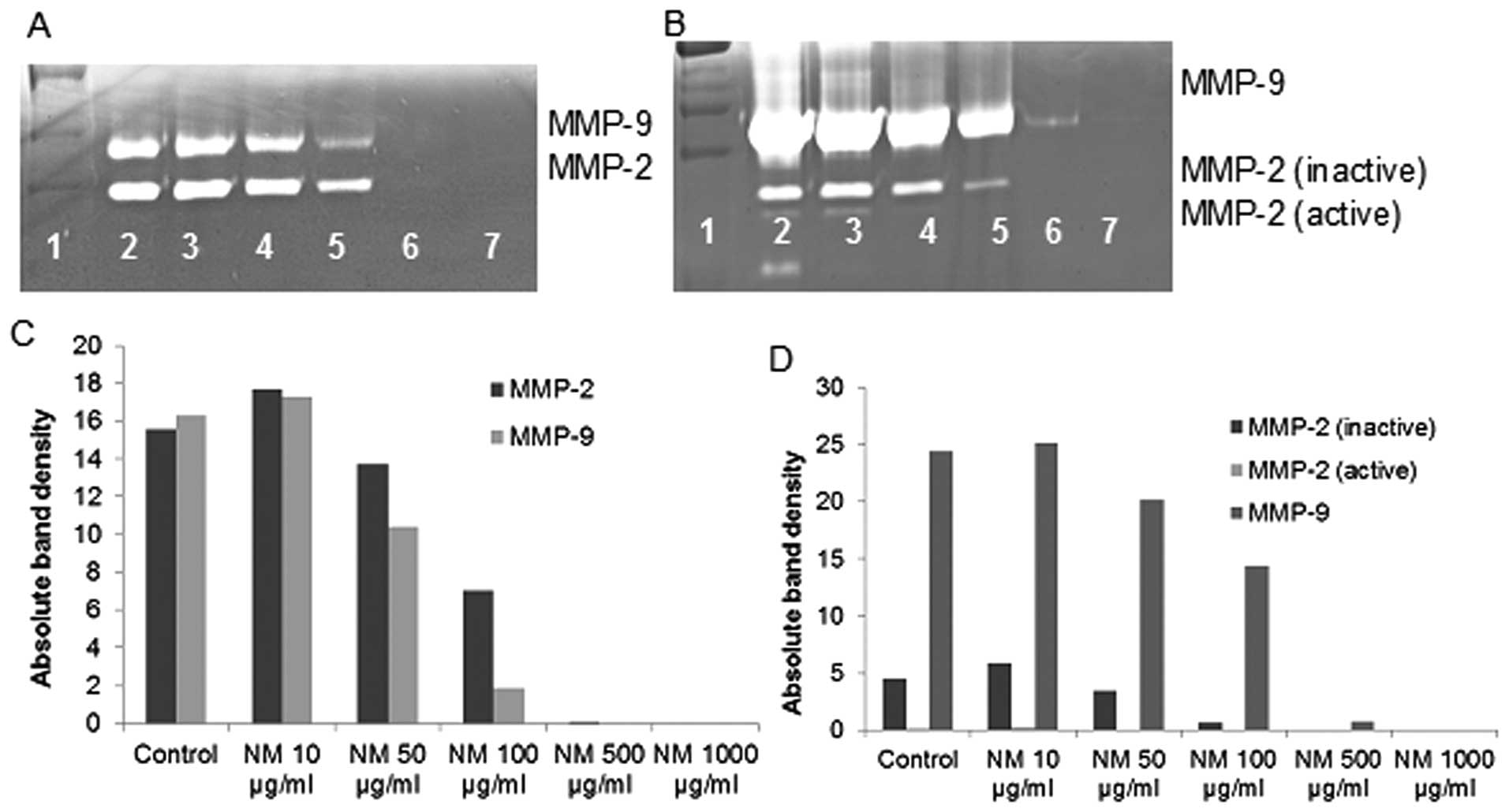
Effect of NM on MMP-2 and MMP-9
expression by liposarcoma cell line SW-872
Zymography demonstrated strong expression of MMP-2
and MMP-9 by LPS cells that were inhibited by NM in a
dose-dependent fashion with virtual total inhibition of MMP-2 and
MMP-9 at 500 μg/ml (linear trend for both
R2=0.885). PMA (100 ng/ml) treatment profoundly enhanced
MMP-9 expression by SW-872 cells; NM inhibited MMP-2 and MMP-9 in a
dose-dependent manner with total block of MMP-2 at 500 μg/ml
(linear trend R2=0.821) and MMP-9 at 1,000 μg/ml
(linear trend R2=0.898). See Fig. 4 for gelatinase zymograms and
densitometry analyses.
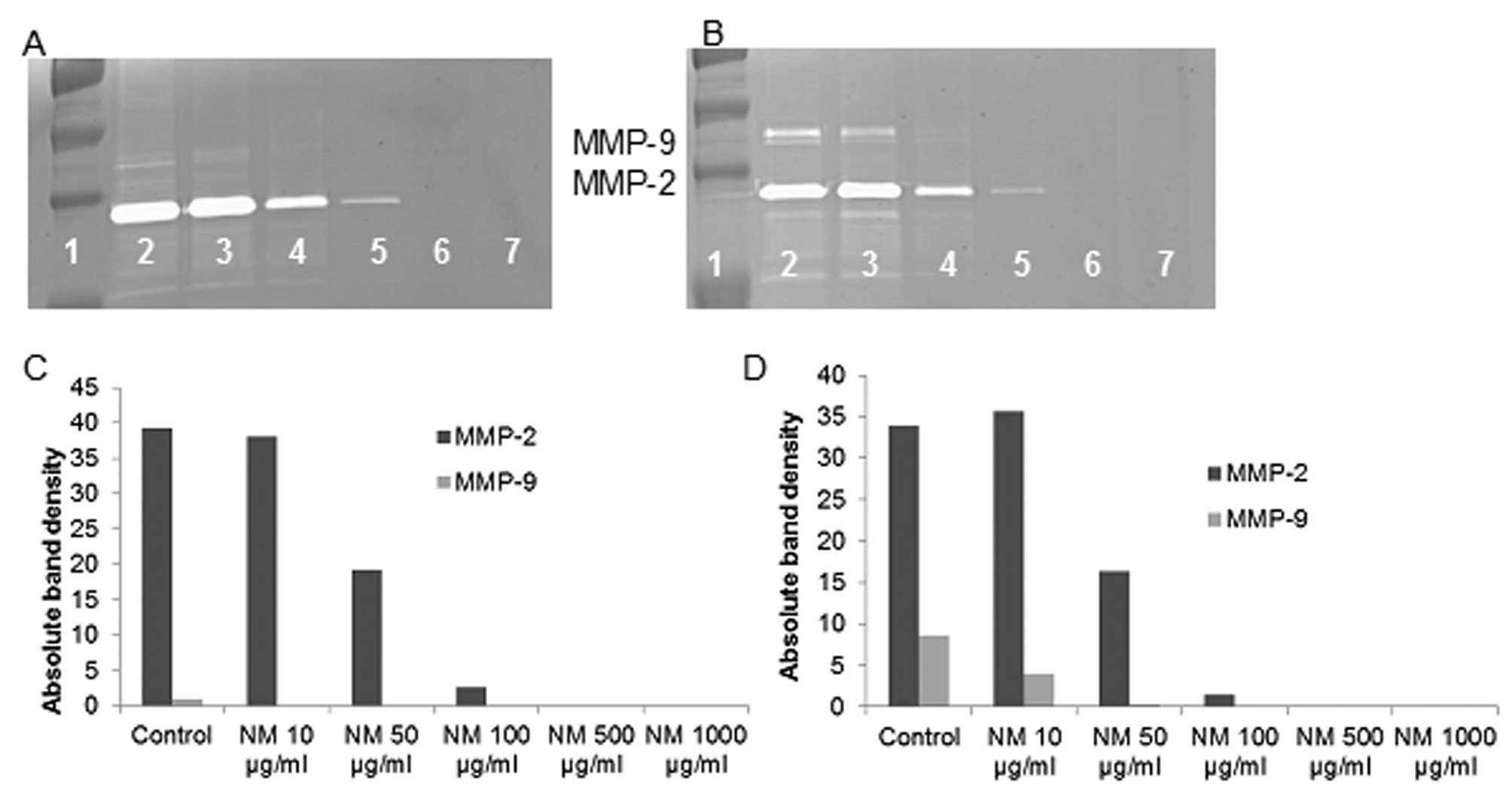
Effect of NM on MMP-2 and MMP-9
expression by synovial sarcoma cell line SW-982
Zymography demonstrated secretion of MMP-2 and a
faint band corresponding to MMP-9 by uninduced SS cells that were
inhibited by NM in a dose-dependent fashion with virtual total
inhibition of MMP-2 at 500 μg/ml (linear trend
R2=0.886) and MMP-9 at 50 μg/ml (linear trend
R2=0.429). PMA (100 ng/ml) treatment enhanced MMP-9
expression by SW-982 cells, but to a lower degree than the other
cell lines; NM inhibited MMP-2 and MMP-9 in a dose-dependent manner
with total block of MMP-2 at 500 μg/ml (linear trend
R2=0.855) and MMP-9 at 50 μg/ml (linear trend
R2=0.694). See Fig. 5
for gelatinase zymograms and densitometry analyses.
Effect of NM on MMP-2 and MMP-9
expression by uterine leimyosarcoma cell line SK-UT-1 uninduced and
PMA-treated
Normal LS cells did not secrete MMP-2 or MMP-9, but
PMA (100 ng/ml)-treated cells demonstrated induced MMP-9 (active)
greater than MMP-9 (inactive) secretions that was inhibited by NM
in a dose-dependent fashion with virtual total inhibition of both
at 500 μg/ml (linear trends R2=0.884 and 0.970,
respectively) See Fig. 6 for
gelatinase zymogram and densitometry analysis.
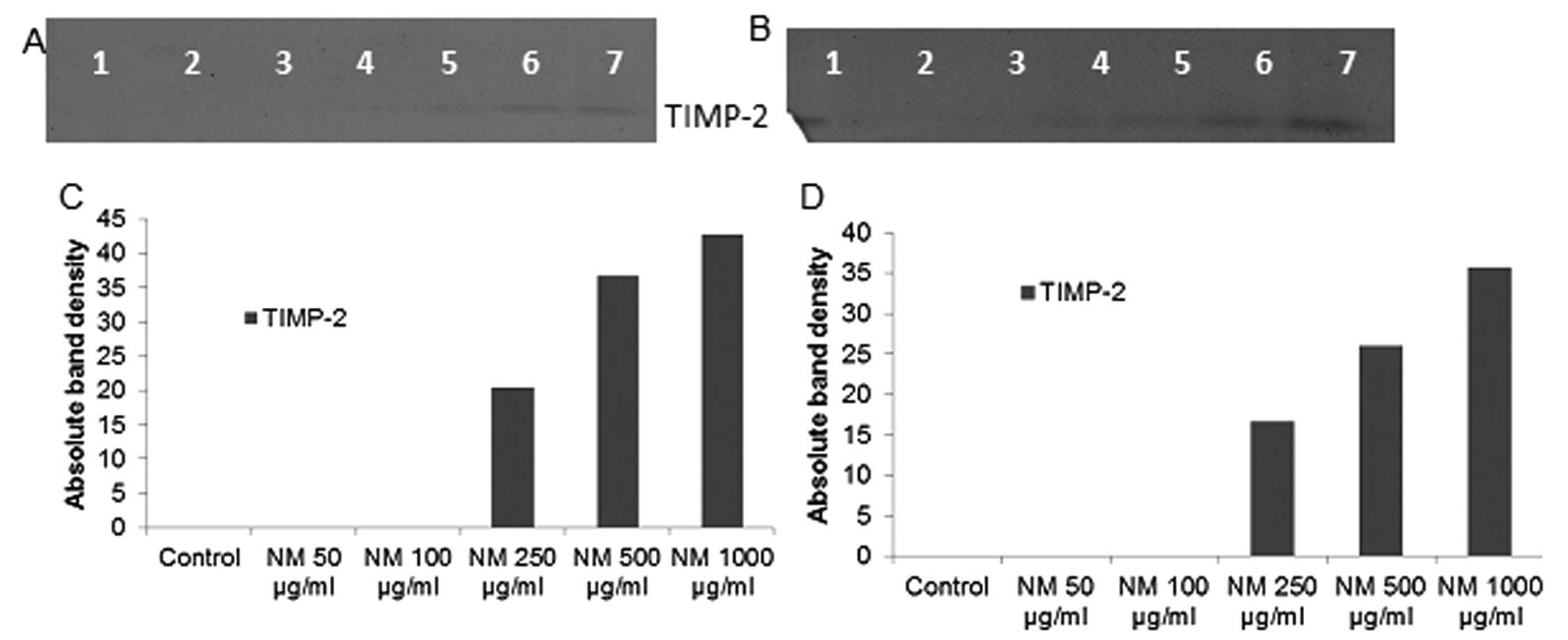
Effect of NM on TIMPs activity in
fibrosarcoma, chondrosarcoma and liposarcoma
Reverse zymography revealed upregulation of TIMP-2
activity with NM treatment in all cancer cell lines in a
dose-dependent manner. Minimum activity was expressed at 50 and
maximum at 1,000 μg/ml NM. See Fig. 7 for respective reverse zymograms
and densitometry analyses.
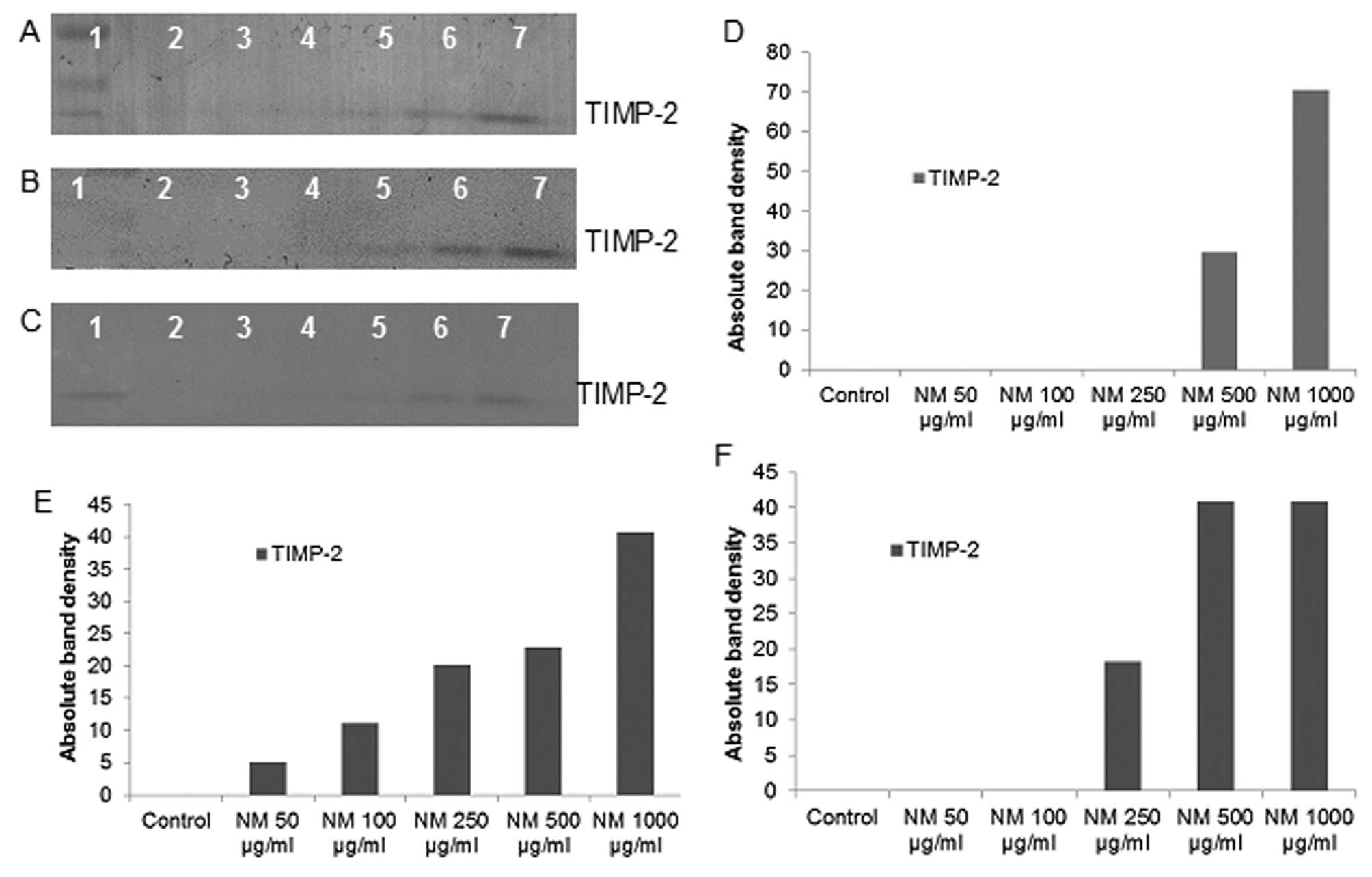 | Figure 7.Effect of NM on fibrosarcoma HT-1080,
chondrosarcoma SW-1353 and liposarcoma SW-872 TIMP-2 expression.
Reverse zymograms of (A) HT-1080, (B) SW-1353 and (C) SW-872 TIMP-2
expression. Lane 1, markers; lane 2, control; lanes 3–7, NM 50,
100, 250, 500 and 1,000 μg/ml. Densitometric analyses of (D)
1080, (E) SW-1353 and (F) SW-872 TIMP-2 expression. |
Effect of NM on TIMP activity in synovial
sarcoma and uterine leiomyosarcoma
Reverse zymography revealed upregulation of TIMP-2
activity with NM treatment in both sarcoma cell lines in a
dose-dependent manner. Minimum activity was expressed at 50 and
maximum at 1,000 μg/ml NM. See Fig. 8 for respective reverse zymograms
and densitometry analyses.
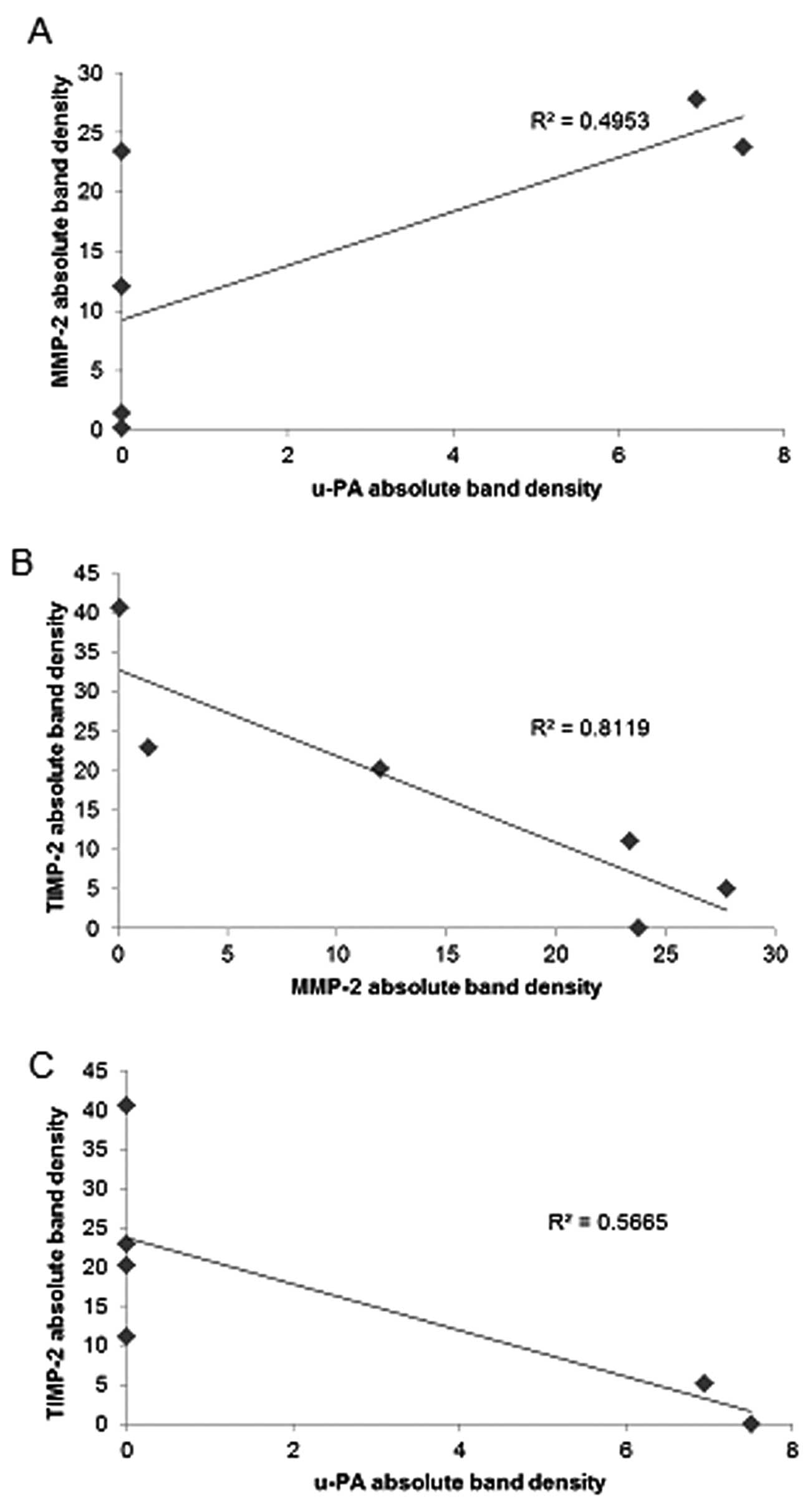
Correlation between adult sarcoma u-PA,
TIMP-2 and MMP expressions
Analysis revealed a positive correlation between
u-PA and MMP-2 expressions of NM-treated adult sarcoma cell lines,
the fibrosarcoma HT-1080, chondrosarcoma SW1353, and liposarcoma
SW-872, as shown in Table II.
Uterine leiomyosarcoma SK-UT-1 demonstrated a positive correlation
between u-PA and MMP-9. Fig. 9A
shows the correlation for chondrosarcoma u-PA and MMP-2 with a
correlation coefficient r=0.703. Negative correlations were found
between the expressions of MMP-2 or MMP-9 and TIMP-2 in all adult
sarcoma cell lines treated with NM as shown in Table II. The correlation (r=−0.901)
between MMP-2 and TIMP-2 is shown for chondrosarcoma in Fig. 9B. Negative correlations were found
between expressions of TIMP-2 and u-PA in all NM-treated adult
sarcoma cell lines studied, except synovial sarcoma, which did not
express u-PA. The correlation (r=−0.753) between u-PA and TIMP-2 is
shown for chondrosarcoma in Fig.
9C.
| Table II.Correlation between effects of NM on
sarcoma cell u-PA, MMPs and TIMPs. |
Table II.
Correlation between effects of NM on
sarcoma cell u-PA, MMPs and TIMPs.
| Cell line | u-PA and MMPs | MMPs and TIMPs | u-PA and TIMPs |
|---|
| Fibrosarcoma
HT-1080 | r=0.974 | r=−0.360 | r=−0.397 |
| Chondrosarcoma
SW-1353 | r=0.703 | r=−0.901 | r=−0.753 |
| Liposarcoma
SW-872 | r=0.412 | r=−0.983 | r=−0.407 |
| Synovial sarcoma
SW-982 | N/A | r=−0.861 | N/A |
| Uterine
leiomyosarcoma SK-UT-1 | r=0.683 | r=−0.917 | r=−0.436 |
Discussion
Tumor cell invasion requires the critical steps of
cell attachment, degradation of the ECM and migration through the
disrupted matrix. The two families of proteases, matrix
metalloproteinases and urokinase plasminogen activators play key
roles in tumor cell invasion. Experimental studies have
demonstrated the role of urokinase plasminogen, especially cell
surface u-PA, as an initiator of ECM proteolysis and associated
tumor cell invasion (21). The
protease u-PA converts plasminogen to plasmin, which is capable of
promoting tumor growth and angiogenesis, degrading the ECM and
basement membrane and activating pro-MMPs (18). Overexpression of u-PA in sarcoma
patients has been correlated with cancer progression, metastasis
and poor prognosis (14). Matrix
metalloproteinases, especially MMP-2 and MMP-9, play pivotal roles
in tumor cell invasion and metastasis due to their ability to
degrade type IV collagen, a major component of the ECM.
Overproduction of MMPs, especially MMP-2 and -9 and low levels of
TIMPs have been shown to be associated with a more aggressive
behavior of sarcomas (4,17,23).
Our study demonstrated that the specific mixture of
nutrients tested significantly inhibited u-PA secretion in
fibrosarcoma HT-1080, chondrosarcoma SW-1353, liposarcoma SW-872
and leiomyosarcoma SK-UT-1 cell lines (synovial sarcoma cell line
SW-982 was not found to secrete u-PA in this study). Furthermore,
the NM demonstrated dose-dependent decrease in MMP secretion and
increase in TIMP-2 secretion by all sarcoma cell lines. As
expected, a significant positive correlation was found between the
secretion of u-PA and MMP-2 and a significant negative correlation
between u-PA and TIMP-2 and between MMP-2 and TIMP-2 secretion by
NM treatment of fibrosarcoma, chondrosarcoma and liposarcoma cells.
As anticipated, a significant positive correlation was found
between the secretion of u-PA and MMP-9 and a significant negative
correlation was found between MMP-9 and TIMP-2 and between u-PA and
TIMP-2 secretion by leiomyosarcoma SK-UT-1 cell line. Furthermore,
a previous study demonstrated significant correlation between NM
inhibition of Matrigel invasion and NM modulation of the MMP-2 and
-9 activities of the sarcoma cell lines studied (24). A significant negative correlation
was found between NM modulation of Matrigel invasion inhibition and
MMP-2 secretion with fibrosarcoma HT-1080 (r=−0.911),
chondrosarcoma SW-1353 (r=−0.942), liposarcoma SW-872 (r=−0.957)
and synovial sarcoma SW-982 (r=−0.878). For uterine leiomyosarcoma
SK-UT-1 cells, a significant negative correlation (r=−0.956) was
found between NM modulation of Matrigel invasion inhibition and
MMP-9 secretion. Previous in vivo studies of the effects of
NM 0.5% dietary effect on xenograft tumor growth of fibrosarcoma
and synovial sarcoma cells in nude mice support these results in
that they demonstrated significant inhibition of xenograft tumor
growth: 59%, p=0.0005 in fibrosarcoma HT-1080 xenografts (25) and 44%, p=0.01 in synovial sarcoma
Hs 701.T xenografts (26).
In contrast to the associated toxicity and limited
efficacy of standard cancer chemotherapy and radiation therapy, the
efficacy and safety of dietary and botanical natural compounds in
cancer prevention has been extensively documented (27). The nutrient mixture was formulated
by selecting nutrients that act on critical physiological targets
in cancer progression and metastasis, as documented in both
clinical and experimental studies. Combining these micronutrients
expands metabolic targets, maximizing biological impact with lower
doses of components. A previous study of the comparative effects of
NM, green tea extract and EGCG on inhibition of MMP-2 and MMP-9
secretion of different cancer cell lines with varying MMP secretion
patterns, revealed the superior potency of NM over GTE and EGCG at
equivalent doses (28). These
results can be understood from the more comprehensive treatment
offered by the combination of nutrients in NM over individual
components of NM since MMP-2 and MMP-9 are mediated by differential
pathways.
Optimal ECM structure depends upon adequate supplies
of ascorbic acid and the amino acids lysine and proline to ensure
proper synthesis and hydroxylation of collagen fibers. In addition,
lysine contributes to ECM stability as a natural inhibitor of
plasmin-induced proteolysis (20,29).
Manganese and copper are also essential for collagen formation.
There is considerable documentation of the potency of green tea
extract in modulating cancer cell growth, metastasis, angiogenesis
and other aspects of cancer progression (30–36).
N-acetyl cysteine and selenium have demonstrated inhibition of
tumor cell MMP-9 and invasive activities, as well as migration of
endothelial cells through ECM (37–39).
Ascorbic acid demonstrates cytotoxic and antimeta-static actions on
malignant cell lines (40–44) and cancer patients have been found
to have low levels of ascorbic acid (45,46).
Low levels of arginine, a precursor of nitric oxide (NO), can limit
the production of NO, which has been shown to predominantly act as
an inducer of apoptosis (47).
In conclusion, the NM demonstrated potent anticancer
activity by targeting primary mechanisms responsible for the
aggressive spread of adult sarcomas. In this in vitro study,
the NM significantly inhibited secretion of u-PA and MMP-2 and/or
-9 and increased secretion of TIMP-2 in fibrosarcoma,
chondrosarcoma, liposarcoma and uterine leiomyosarcoma cells,
suggesting its potential in modulating cancer invasion and
metastasis. Malignant synovial sarcoma cells did not secrete u-PA;
however, MMP-2 and -9 secretions by SW-982 were inhibited by NM and
secretion of TIMP-2 was enhanced by NM. NM inhibition of MMP
secretion was found to be correlated significantly with Matrigel
invasion of all the sarcoma cell lines studied. Furthermore, use of
the nutrient mixture would not pose any toxic effect clinically,
especially in the relevant doses, as in vivo safety studies
demonstrate. An in vivo toxicology study showed that NM had
no adverse effects on vital organs (heart, liver and kidney), or on
the associated functional serum enzymes (48).
Acknowledgements
Mr. Monterrey provided assistance in
scanning the gels. The research study was funded by Dr Rath Health
Foundation (Santa Clara, CA, USA) a non-profit organization.
References
|
1.
|
American Cancer Society: Adult soft tissue
cancer. What are key statistics of soft tissue cancers? http://www.cancer.org/cancer/sarcoma-adultsofttissuecancer/detailedguide/sarcoma-adult-soft-tissue-cancer-key-statistics.
Accessed January 21, 2013.
|
|
2.
|
American Cancer Society: Bone cancer. What
are the key statistics about bone cancer? http://www.cancer.org/cancer/bonecancer/detailedguide/bone-cancer-key-statistics.
Accessed January 21, 2013.
|
|
3.
|
Papagelopoulos PJ, Galanis E, Frassica FJ,
Sim FH, Larson DR and Wold LE: Primary fibrosarcoma of bone.
Outcome after primary surgical treatment. Clin Orthop Relat Res.
373:88–103. 2000.PubMed/NCBI
|
|
4.
|
Benassi MS, Gamberi G, Magagnoli G,
Molendini L, Ragazzini P, Merli M, Chiesa F, Balladelli A, Manfrini
M, Bertoni F, Mercri M and Picci P: Metalloproteinase expression
and prognosis in soft tissue sarcomas. Ann Oncol. 12:75–80. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Fidler IJ: Molecular biology of cancer:
invasion and metastasis. Cancer Principles and Practice of
Oncology. De Vita VT, Hellman S and Rosenberg SA: 5th edition.
Lippincott-Raven; Philadelphia, PA: pp. 135–152. 1997
|
|
6.
|
Egeblad M and Werb Z: New functions for
the matrix metalloproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Folkman J: Role of angiogenesis in tumor
growth and metastasis. Semin Oncol. 29(Suppl 16): 15–18. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Chambers AF and Matrisian LM: Changing
views on the role of matrix metalloproteinases in metastasis. J
Natl Cancer Inst. 89:1260–1270. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Kleiner DL and Stetler-Stevenson WG:
Matrix metalloproteinases and metastasis. Cancer Chemother
Pharmacol. 43(Suppl): 42s–51s. 1999. View Article : Google Scholar
|
|
10.
|
Yurchenko PD and Schitny JC: Molecular
architecture of basement membranes. FASEB J. 4:1577–1590.
1990.PubMed/NCBI
|
|
11.
|
Barsky SH, Siegel GP, Jannotta F and
Liotta LA: Loss of basement membrane components by invasive tumors
but not by their benign counterparts. Lab Investig. 49:140–147.
1983.PubMed/NCBI
|
|
12.
|
Liotta LA, Tryggvason K, Garbisa A, Hart
I, Foltz CM and Shafie S: Metastatic potential correlates with
enzymatic degradation of basement membrane collagen. Nature.
284:67–68. 1980. View
Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Nelson AR, Fingleton B, Rothenberg ML and
Matrisian LM: Matrix metalloproteinases: biologic activity and
clinical implications. J Clin Oncol. 18:1135–1149. 2000.PubMed/NCBI
|
|
14.
|
Taubert H, Würl P, Greither T, Kappler M,
Bache M, Lautenschläger C, Füssel S, Meye A, Eckert AW, Holzhausen
HJ, Magdolen V and Kotzsch M: Co-detection of members of the
urokinase plasminogen activator system in tumour tissue and serum
correlates with a poor prognosis for soft-tissue sarcoma patients.
Br J Cancer. 102:731–737. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Stetler-Stevenson WG: The role of matrix
metalloproteinases in tumor invasion, metastasis and angiogenesis.
Surg Oncol Clin N Am. 10:383–392. 2001.PubMed/NCBI
|
|
16.
|
Stetler-Stevenson WG: Type IV collagenases
in tumor invasion and metastasis. Cancer Metastasis Rev. 9:289–303.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Benassi MS, Magagnoli G, Ponticelli F,
Pazzaglia L, Zanella L, Gamberti G, Ragazzini P, Ferrari C, Mercuri
M and Picci P: Tissue and serum loss of metalloproteinases
inhibitors in high-grade soft tissue sarcomas. Histol Histopathol.
18:1035–1040. 2003.PubMed/NCBI
|
|
18.
|
Dano K, Andreasen PA, Grondahl-Hansen J,
Kristensen P, Nielsen LS and Skriver L: Plasminogen activators,
tissue degradation and cancer. Adv Cancer Res. 44:139–266. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Duffy MJ, Duggan C, Mulcahy HE, McDermott
EW and O'Higgins NJ: Urokinase plasminogen activator: a prognostic
marker in breast cancer including patients with axillary
node-negative disease. Clin Chem. 44:1177–1183. 1998.PubMed/NCBI
|
|
20.
|
Rath M and Pauling L: Plasmin-induced
proteolysis and the role of apoprotein(a), lysine and synthetic
analogs. Orthomolecular Med. 7:17–23. 1992.
|
|
21.
|
Andreasen PA, Kjøller L, Christensen L and
Duffy MJ: The urokinase-type plasminogen activator system in cancer
metastasis: a review. Int J Cancer. 72:1–22. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Niedzwiecki A, Roomi MW, Kalinovsky T and
Rath M: Micronutrient synergy - a new tool in effective control of
metastasis and other key mechanisms of cancer. Cancer Metastasis
Rev. 29:529–543. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Roebuck MM, Helliwell TR, Chaudhry IH,
Kalogrianitis S, Carter S, Kemp G, Ritchie DA, Jane MJ and Frostick
SP: Matrix metalloproteinase expression is related to angiogenesis
and histologic grade in spindle cell soft tissue neoplasms of the
extremities. Am J Clin Path. 123:405–414. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Roomi MW, Monterrey JC, Kalinovsky T,
Niedzwiecki A and Rath M: Inhibition of invasion and MMPs by a
nutrient mixture in human cancer cell lines: a correlation study.
Exp Oncol. 32:243–248. 2010.PubMed/NCBI
|
|
25.
|
Roomi MW, Ivanov V, Kalinovsky T, Rath M
and Niedzwiecki A: In vivo and in vitro antitumor effect of
ascorbic acid, lysine, proline, arginine and green tea extract on
human fibrosarcoma cells HT-1080. Med Oncol. 23:105–112. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Roomi MW, Ivanov V, Kalinovsky T,
Niedzwiecki A and Rath M: In vitro and in vivo anti-tumor effect of
a nutrient mixture containing ascorbic acid, lysine, proline and
green tea extract on human synovial sarcoma cancer cells. JAMA.
9:30–34. 2006.
|
|
27.
|
Amin ARMR, Kucek O, Khuri FR and Shin DM:
Perspectives for cancer prevention with natural compounds. J Clin
Oncol. 27:2712–2725. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Roomi MW, Monterrey JC, Kalinovsky T, Rath
M and Niedzwiecki A: Comparative effects of EGCG, green tea and a
nutrient mixture on the patterns of MMP-2 and MMP-9 expression in
cancer cell lines. Oncol Rep. 24:747–757. 2010.PubMed/NCBI
|
|
29.
|
Sun Z, Chen YH, Wang P, Zhang J, Gurewich
V, Zhang P and Liu JN: The blockage of high-affinity lysine binding
sites of plasminogen by EACA significantly inhibits
prourokinase-induced plasminogen activation. Biochem Biophys Acta.
1596:182–192. 2002.
|
|
30.
|
Kemberling JK, Hampton JA, Keck RW, Gomez
MA and Selman SH: Inhibition of bladder tumor growth by the green
tea derivative epigallocatechin-3-gallate. J Urol. 170:773–776.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Sato D and Matsushima M: Preventive
effects of urinary bladder tumors induced by
N-butyl-N-(4-hydroxybutyl)-nitrosamine in rat by green tea leaves.
Int J Urol. 10:160–166. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Valcic S, Timmermann BN, Alberts DS,
Wachter GA, Krutzsch M, Wymer J and Guillen JM: Inhibitory effect
of six green tea catechins and caffeine on the growth of four
selected human tumor cell lines. Anticancer Drugs. 7:461–468. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Mukhtar H and Ahmed N: Tea polyphenols:
prevention of cancer and optimizing health. Am J Clin Nutr.
71:1698s–1702s. 2000.PubMed/NCBI
|
|
34.
|
Yang GY, Liao J, Kim K, Yurtow EJ and Yang
CS: Inhibition of growth and induction of apoptosis in human cancer
cell lines by tea polyphenols. Carcinogenesis. 19:611–616. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Taniguchi S, Fujiki H, Kobayashi H, Go H,
Miyado K, Sadano H and Shimikawa R: Effect of (-) epigallocatechin
gallate, the main constituent of green tea, on lung metastasis with
mouse B16 melanoma cell lines. Cancer Lett. 65:51–54. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Hara Y: Green tea: Health Benefits and
Applications. Marcel Dekker; New York: 2001, View Article : Google Scholar
|
|
37.
|
Kawakami S, Kageyama Y, Fujii Y, Kihara K
and Oshima H: Inhibitory effects of N-acetyl cysteine on invasion
and MMP 9 production of T24 human bladder cancer cells. Anticancer
Res. 21:213–219. 2001.PubMed/NCBI
|
|
38.
|
Morini M, Cai T, Aluigi MG, Noonan DM,
Masiello L, De Floro S, D'Agostinin F, Albini A and Fassima G: The
role of the thiol N-acetyl cysteine in the prevention of tumor
invasion and angiogenesis. Int J Biol Markers. 14:268–271.
1999.PubMed/NCBI
|
|
39.
|
Yoon SO, Kim MM and Chung AS: Inhibitory
effects of selenite on invasion of HT 1080 tumor cells. J Biol
Chem. 276:20085–20092. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Naidu KA, Karl RC and Coppola D:
Antiproliferative and proapoptotic effect of ascorbyl stearate in
human pancreatic cancer cells: association with decreased
expression of insulin-like growth factor 1 receptor. Dig Dis Sci.
48:230–237. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Anthony HM and Schorah CJ: Severe
hypovitaminosis C in lung-cancer patients: The utilization of
vitamin C in surgical repair and lymphocyte-related host
resistance. Br J Cancer. 46:354–367. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
42.
|
Maramag C, Menon M, Balaji KC, Reddy PG
and Laxmanan S: Effect of vitamin C on prostate cancer cells in
vitro: effect on cell number, viability and DNA synthesis.
Prostate. 32:188–195. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43.
|
Koh WS, Lee SJ, Lee H, Park C, Park MH,
Kim WS, Yoon SS, Park K, Hong SI, Chung MH and Park CH:
Differential effects and transport kinetics of ascorbate
derivatives in leukemic cell lines. Anticancer Res. 8:2487–2493.
1998.PubMed/NCBI
|
|
44.
|
Chen Q, Espey MG, Krishna MC, Mitchell JB,
Corpe CP, Buettner GR, Shacter E and Levine M: Pharmacologic
ascorbic acid concentrations selectively kill cancer cells: action
as a pro-drug to deliver hydrogen peroxide to tissues. Proc Natl
Acad Sci USA. 102:13604–13609. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45.
|
Nunez C, Ortiz de Apodaca Y and Ruiz A:
Ascorbic acid in the plasma and blood cells of women with breast
cancer. The effect of consumption of food with an elevated content
of this vitamin. Nutr Hosp. 10:368–372. 1995.(In Spanish).
|
|
46.
|
Kurbacher CM, Wagner U, Kolster B,
Andreotti PE, Krebs D and Bruckner HW: Ascorbic acid (vitamin C)
improves the antineoplastic activity of doxorubicin, cisplatin and
paclitaxel in human breast carcinoma cells in vitro. Cancer Lett.
103:183–189. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
47.
|
Cooke JP and Dzau VJ: Nitric oxide
synthase: role in the genesis of vascular disease. Annu Rev Med.
48:489–509. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
48.
|
Roomi MW, Ivanov V, Netke SP, Niedzwiecki
A and Rath M: Serum markers of the liver, heart, and kidney and
lipid profile and histopathology in ODS rats treated with nutrient
synergy. J Am Coll Nutr. 22:477abst 86,. 2003.
|