Introduction
TRAIL, also known as Apo2L and TNFSF10, belongs to
the tumor necrosis factor (TNF) superfamily that induces cell death
in various types of cancer cell lines in vivo and in
vitro but has little or no effect on normal cell lines. The
cytotoxic effects of TRAIL are carried out by its functional
receptors, death receptor 4 (DR4) and death receptor 5 (DR5) (also
known as TRAIL-R1 and TRAIL-R2), and death inducing signaling
complex (DISC). DISC formation involves recruitment of an adaptor
molecule, Fas-associated death domain (FADD), as well as apical
caspase-8 and caspase-10. DISC assembly results in the
auto-catalytic activation of caspase-8 and caspase-10, which
initiates the caspase cascade, leading to apoptosis (1,2).
Upon binding to DR4 and DR5, TRAIL activates the nuclear factor
(NF)-κB and JNK signaling pathways (3). NF-κB plays an important role
regulating anti- and pro-apoptotic events, depending on the
physiological environment (4).
NF-κB mediates the expression of anti-apoptotic genes, such as
members of the inhibitor of apoptosis (IAP) family (cIAP-1 and -2
and XIAP), cellular Flice-like inhibitory proteins (c-FLIP) and
Bcl-2, all of which determine susceptibility to TRAIL-derived
apoptosis (5,6).
TRAIL is regarded as a promising anticancer agent
because of its selective cytotoxicity against transformed cell
lines; however, the potential clinical use of TRAIL has been
blocked because some tumor cell lines become resistant to
TRAIL-derived apoptosis (7,8).
Resistance is generally induced by low expression of DR4/DR5 or
high expression of IAP family members (8,9);
thus, the discovery of agents that alleviate TRAIL resistance may
allow the use of combined TRAIL-based therapeutic regimens to treat
resistant cancers (10).
Recombinant human TRAIL and several agonistic monoclonal antibodies
are currently in phase II clinical trials. The agonistic antibodies
include mapatumumab, which targets DR4, and lexatumumab, apomab,
AMG655, CS-1008, and LBY-135, which target DR5 (11). However, increasing evidence
indicates that death receptor agonists alone may not be sufficient
to effectively activate apoptosis in many cancers.
Chemotherapy is not a standard treatment for
hepatocellular carcinoma (HCC), and HCC is highly resistant to
other conventional anti-neoplastic agents (12,13).
Recent studies have shown that many cancer cell lines are not
susceptible to the cytotoxic effects of TRAIL (14); thus, the discovery of agents that
alleviate TRAIL resistance may be useful for the establishment of
TRAIL-based combined regimens for improved HCC treatment.
A formulated red ginseng extract (RGE) derived from
heat-processed Panax ginseng C.A. Meyer, is a functional food and
dietary supplement consumed worldwide and is used in traditional
oriental medicine to increase energy and life expectancy. RGE has
been reported to reduce fatigue and to possess antioxidant and
antitumor activities (15–17). Ginseng has been reported to possess
antioxidant, antistress and immune-stimulating activities. These
activities require the scavenging of hydroxyl,
1-1-diphenyl-2-picryhydrazyl (DPPH) and superoxide radicals to
decrease lipid peroxidation through chelation of transition metal
ions and to reduce oxidative DNA damage caused by the Fenton
reaction or UV light exposure (18). Ginseng saponins also protect human
low-density lipoproteins from oxidative damage in vitro
(19) and induce Cu/Zn-superoxide
dismutase expression at the transcriptional level (20).
Findings indicate that the RGE could be useful as a
dietary supplement in a combined cancer treatment, as it sensitizes
human HCC cell lines to TRAIL-derived cell death via a mechanism
involving upregulation of DR5, which then enhances activation of
the extrinsic apoptosis pathway. In this case, upregulation of DR5
is induced by C/EBP homologous (CHOP), an endoplasmic reticulum
(ER) stress responsive element. shRNA-mediated downregulation of
CHOP expression effectively suppresses cell death induced by RGE
plus TRAIL in HepG2 cells, indicating that CHOP is essential for
RGE-triggered enhancement of TRAIL-derived apoptosis. To summarize,
data from the present study suggest that RGE may sensitize cells to
TRAIL, thereby diminishing their resistance. RGE could be combined
with TRAIL in a cocktail for use as a novel therapeutic strategy to
more effectively treat HCC.
Materials and methods
Reagents
The following reagents were purchased and used
according to the manufacturer’s instructions: glutathione
S-transferase (GST)-TRAIL and anti-DR5 antibodies were from Koma
Biotechnologies (Seoul, Korea); anti-caspase 3, anti-PARP and
anti-CHOP antibodies were from Cell Signaling Technology; anti-DR4
antibody was from Rockland; anti-tubulin antibody was from Abcam;
anti-actin antibody, thapsigargin (Tg), necrostatin-1, NAC and BHA
were from Sigma; and zVAD was from R&D Systems. A formulated
RGE was provided by the Korea Ginseng Corporation (Seoul,
Korea).
Cell culture
The SK-Hep1, HepG2 and Hep3B cell lines were grown
on culture plates in Dulbecco’s modified Eagle’s medium
supplemented with 10% fetal bovine serum (FBS), 2 mM glutamine, 100
U/ml penicillin and 100 μg/ml streptomycin. Huh-7 cells were
grown on culture plates in RPMI-1640 medium supplemented with 10%
FBS, 2 mM glutamine, 100 U/ml penicillin and 100 μg/ml
streptomycin. A commercially available normal liver cell line
(HL-7702) was grown on culture plates in RPMI-1640 medium
supplemented with 20% FBS, 2 mM glutamine, 100 U/ml penicillin and
100 μg/ml streptomycin.
Western blot analysis
Upon treatment, cell lines were lysed in 20 mM Tris
(pH 7.0), 250 mM NaCl, 3 mM EDTA, 3 mM EGTA, 2 mM DTT, 0.5% NP-40,
0.5 mM PMSF, 20 mM β-glycerol phosphate, 1 mM sodium vanadate and 1
μg/ml leupeptin. Lysates were loaded onto 10 or 12% SDS-PAGE
gels. Following transfer and blotting, the proteins of interest
were visualized by enhanced chemiluminescence (ECL, Pierce) and
analyzed.
Cytotoxicity assays
Viable cells were measured by the MTT assay (dual
labeling method with 2 μM calcein-AM and 4 μM
EthD-1), and MTT absorbance was read at 570 nm. Lactate
dehydrogenase (LDH) leakage was quantified using a cytotoxicity
detection kit (Promega). Images were recorded by microscope and the
data presented are from at least three independent experiments.
Lentiviral shRNA experiments
shRNA-encoding plasmids were purchased from
Sigma-Aldrich, and the targets were coding region sequences or the
3′ untranslated region of CHOP mRNA (NM: 004083). The 293T cell
line (LV900A-1) was transfected with lentiviral plasmids using
Lipofectamine 2000 transfection reagent (Invitrogen, 11668019).
HepG2 cell lines were infected with pseudoviral particles collected
from the 293T cell line 2 days after transfection in the presence
of Polybrene (8 μg/ml). The lentivirus-infected HepG2 cell
line was then isolated by puromycin (1 μg/ml) resistance 2
days after infection, and knockdown of CHOP was confirmed by
western blot analysis. CHOP knockdown cell lines were seeded on
6-well plates and treated with the RGE or Tg for the times
indicated for each experiment and were analyzed by western blot
analysis.
Results and Discussion
Analysis of the formulated RGE
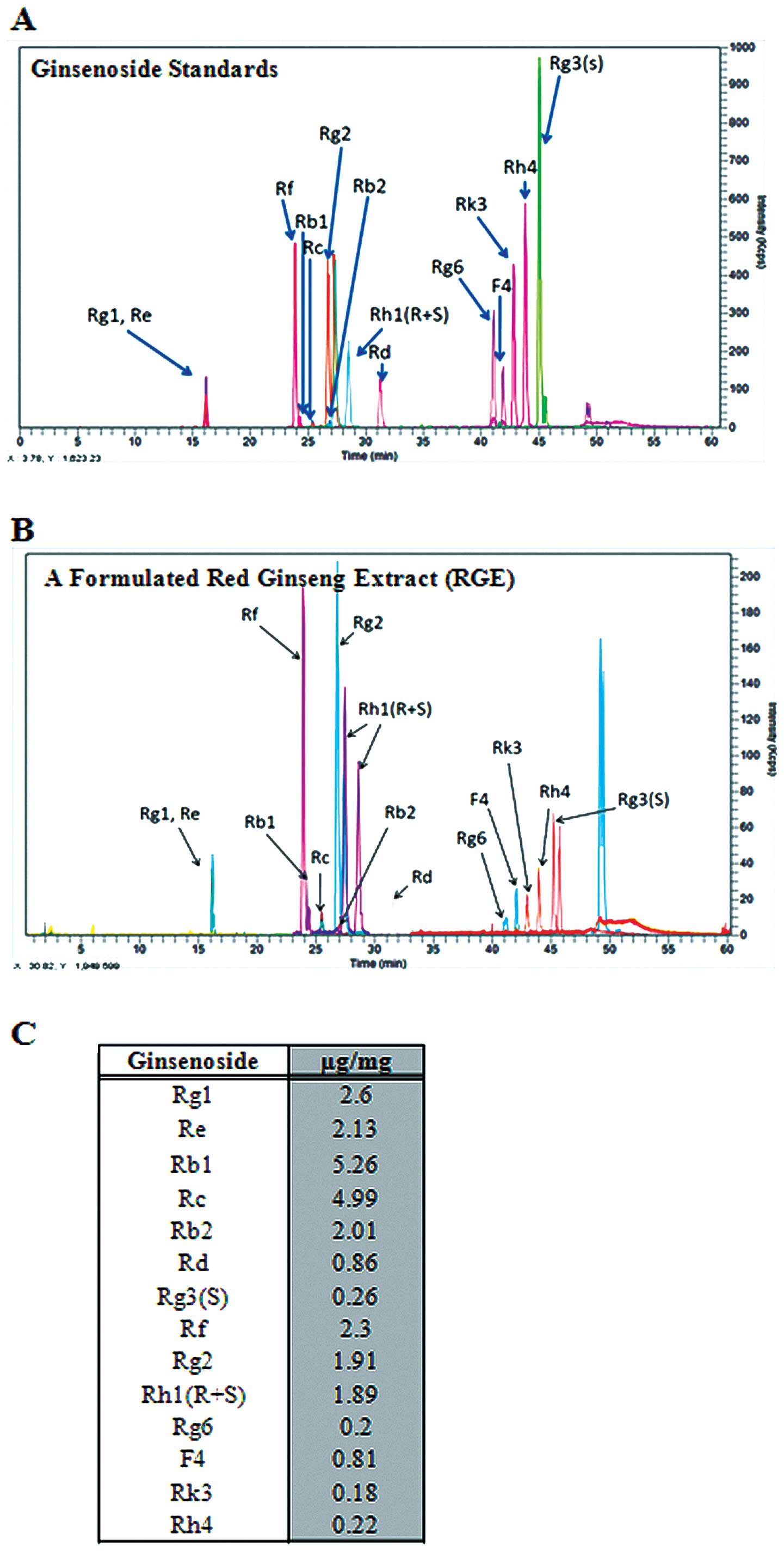
A high-performance liquid chromatography/mass
spectrometry (LC/MS) method was used for the targeted 14
ginsenosides to determine the ginsenoside content in the formulated
RGE. Butanol extracts of white, red and sun ginseng were employed
to isolate the standard ginsenosides, and each ginsenoside was
isolated and purified by silica gel chromatography or preparative
LC (21,22). Calibration curves for each
ginsenoside standard were plotted (data not shown), and the
ginsenoside contents of the formulated RGE were calculated
(Fig. 1, Table I). Korean red ginseng has been
reported to induce antitumor and immune-stimulating, antistress and
antioxidant activities. The cytoprotective and chemoprotective
properties of ginseng have been attributed to its ability to reduce
oxidative or nitrosative stress (23–25).
Previous studies have suggested that RGE, a major component of
ginseng, inhibits cancer cell propagation and metastasis (24). Based on these observations, we
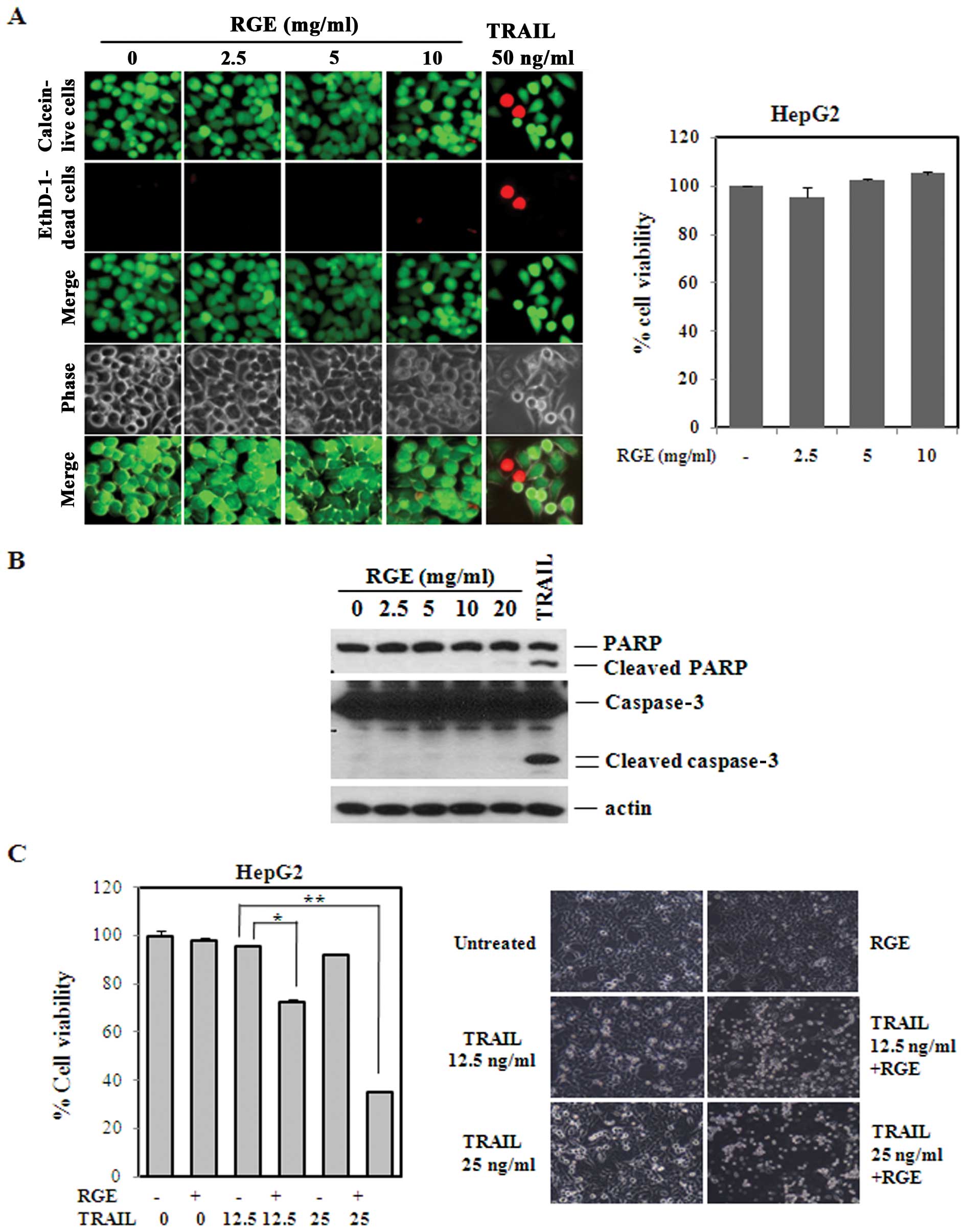
tested the cytotoxicity of the RGE in the HepG2 cell lines. As
shown in Fig. 2A, RGE
concentrations up to 10 mg/ml showed no significant effects on
cells (left panel) and no decrease in viability, as measured by LDH
release (right panel). As a positive control, we treated the HepG2
cell lines with TRAIL (50 ng/ml) and observed the induction of cell
death, as evidenced by the EthD-1-positive cell line. We found that
TRAIL (25 ng/ml) induced poly (ADP-ribose) polymerase (PARP)
cleavage at 12 h, but that the RGE alone had little effect on PARP
and caspase-3 cleavage (Fig.
2B).
 | Table I.LC/MS conditions for the
quantification of ginsenoside in RGE. |
Table I.
LC/MS conditions for the
quantification of ginsenoside in RGE.
| Instrument | Perkin Elmer FX-10
Ultra High Performance Liquid Chromatography/SQ 300 |
| Column | Phenomenex Luna 5
μm C18 (250×4.60 mm, ID 5 μm) |
| Mobile phase
condition | Eluent A, water;
eluent B, acetonitrile; gradient, 0–20 min (15–34.5% B), 20–30 min
(34.5–36% B), 30–40 min (36–47.5% B), 40–45.2 min (47.5–55% B),
45.2–46.2 min (55–100% B), 46.2–60 min (100% B) |
| Flow rate
(ml/min) | 1 ml/min → 0.4
ml/min (split mode) |
| Column
temperature | 20°C |
| Detector | SQ 300 MS detector
(single quadrupole) |
| Ionization
source | ESI (electrospray
ionization) |
| Drying gas
temperature | 300°C |
| Drying gas flow
rate | 10 l/min |
| Nebulizer gas
pressure | 80 psi |
TRAIL-derived cell death was augmented in
the HCC cell lines by pretreatment with the RGE
Because our data suggested that the RGE alone has no
obvious effect on cancer cell cytotoxicity, we then measured the
sensitization effect of the RGE on TRAIL-derived cell death. When
the RGE and TRAIL were added to cells consecutively, dramatic cell
death occurred according to the results of the MTT colorimetric
assay (Fig. 2C). Concentrations of
TRAIL >50 ng/ml were required to induce significant
cytotoxicity; however, concentrations of TRAIL <25 ng/ml showed
minimal cytotoxicity. Viability measurements from concentration
curves obtained from 1:2 serial dilutions demonstrated that 10
mg/ml of the RGE was necessary to achieve robust TRAIL-derived cell
death (data not shown), but RGE alone showed a minimal cytotoxic
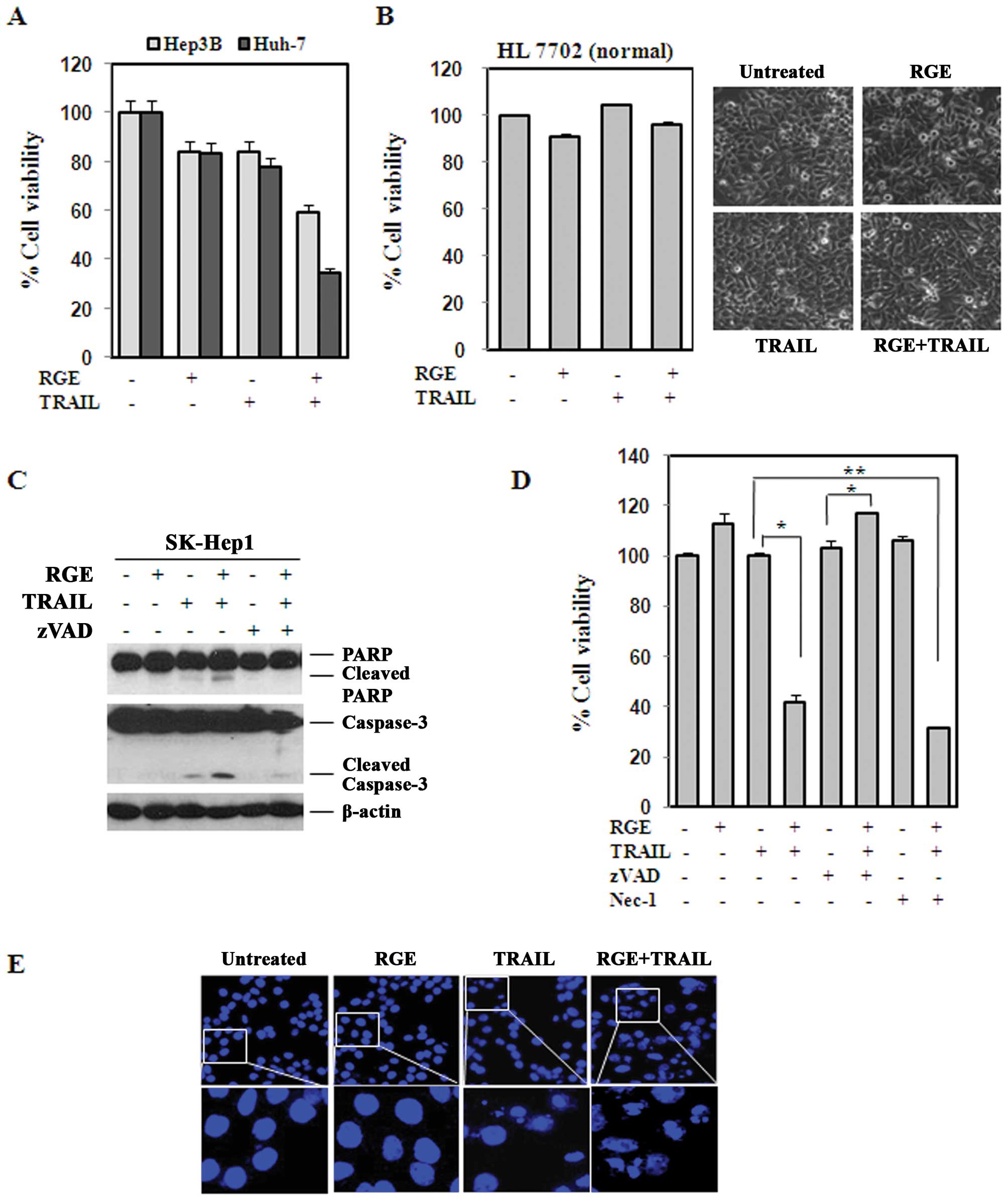
effect. We further tested the sensitization effect in the Hep3B and
Huh-7 cell lines. A similar degree of TRAIL (25 ng/ml)
sensitization was observed, indicating that this effect was
universal in HCC cell lines (Fig.
3A). In contrast, a combination of the RGE and TRAIL sensitize
normal human liver cell lines, confirming that this sensitization
was specific to the tumor cell lines (Fig. 3B).
One of the primary problems when clinically applying
TRAIL as a cancer therapeutic agent is resistance. One effective
strategy to surmount this issue would be to discover anticancer
agents for use in combination therapy (26–29).
Several natural products and other chemical compounds have been
identified that sensitize cancer cell lines to TRAIL-derived
apoptosis. In this study, we tested a formulated RGE that has been
suggested to possess anticancer properties and the results provide
substantial evidence that the RGE was capable of sensitizing HCC to
TRAIL-derived apoptosis by upregulating the CHOP, which then
induced DR5 expression.
Sensitization effect of the RGE in
TRAIL-derived cell death is caspase-dependent
We next examined the effects of the RGE on TRAIL
activation of the caspase cascade in the SK-Hep1 cell line. As
shown in Fig. 3C, treatment with a
combination of RGE and TRAIL induced cleavage of caspase-3 and
PARP, a well-established caspase substrate, whereas treatment with
the RGE alone did not. In addition, cell death was apoptotic
because necrostatin-1 failed to inhibit cell death, as shown in
Fig. 3D. To further confirm RGE
and TRAIL-derived apoptosis, cells were cultured on coverslips and
treated with RGE, TRAIL or RGE plus TRAIL. The cells were fixed and
stained with DAPI after 16–18 h, and examined for morphologic
changes such as nuclear DNA fragmentation and/or condensation,
which are indicative of apoptotic death. As shown in Fig. 3E, the RGE alone had no clear effect
on nuclear DNA fragmentation and/or condensation, but RGE
pretreatment significantly augmented TRAIL-derived nuclear DNA
fragmentation and/or condensation in HepG2 cells.
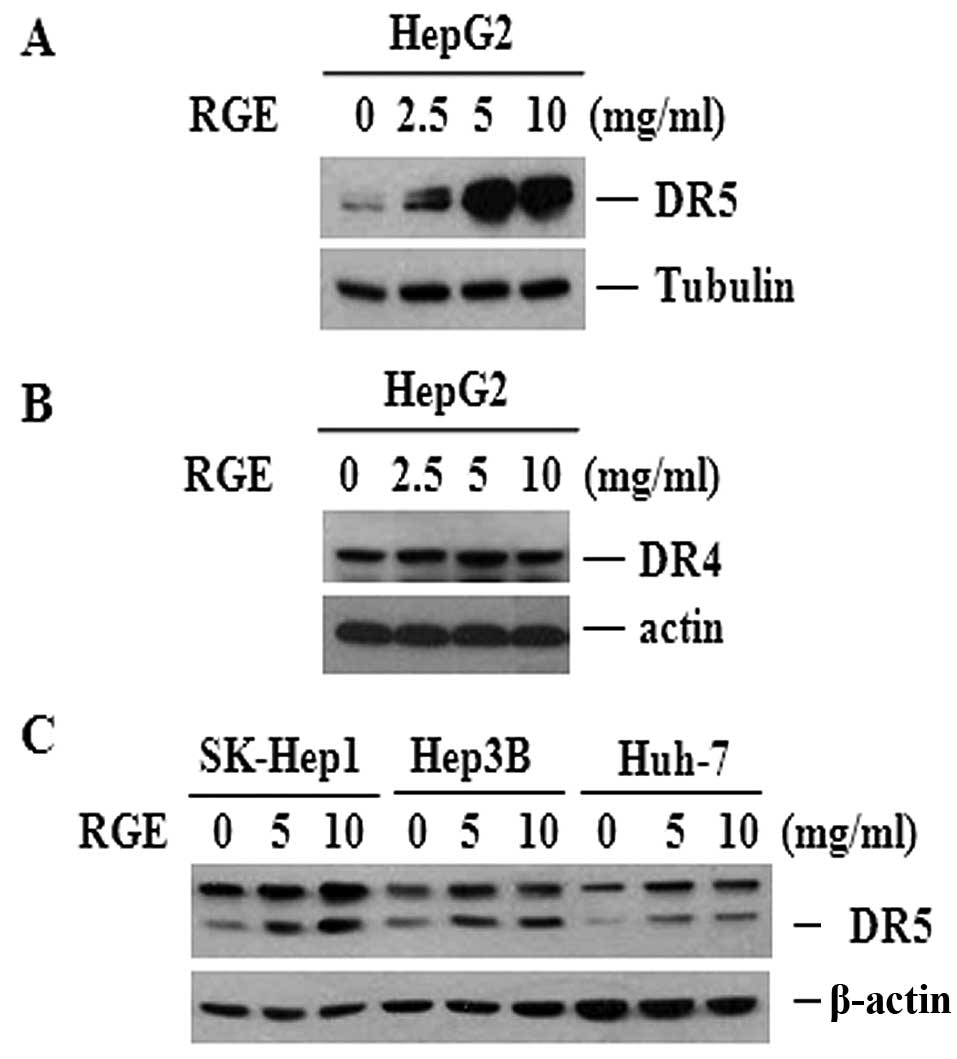
RGE induces upregulation of DR5
expression
TRAIL resistance is facilitated by the
downregulation of DR4 and DR5 and upregulation of DcR1 and DcR2
(8,30–32).
We examined TRAIL receptor protein levels in cells treated with the
RGE to explore the molecular mechanisms underlying the
sensitization effect of the RGE to TRAIL-derived cell death.
Treatment with the RGE significantly increased the expression of
DR5 in a dose-dependent manner in HepG2 cells (Fig. 4A), although no clear changes were
found in DR4 expression (Fig. 4B).
We compared the effects of the RGE in three different HCC cell
lines to determine whether this effect was cell-type specific
(Fig. 4C). All cell lines tested
showed upregulated DR5 expression following the RGE treatment,
suggesting that the effects of RGE on the upregulation of DR5 are
not cell-type specific.
Many mechanisms have been elucidated for inducing
DR5, including ER stress, p53 induction, reactive oxygen species
(ROS) generation and NF-κB/MAPK activation (33–35).
It has been suggested that C/EBP homologous protein/GADD153, a
transcription factor of the C/EBP family that has a role in ER
stress, is involved in activating DR5 expression and thus
contributes to the effectiveness of the RGE and TRAIL combined
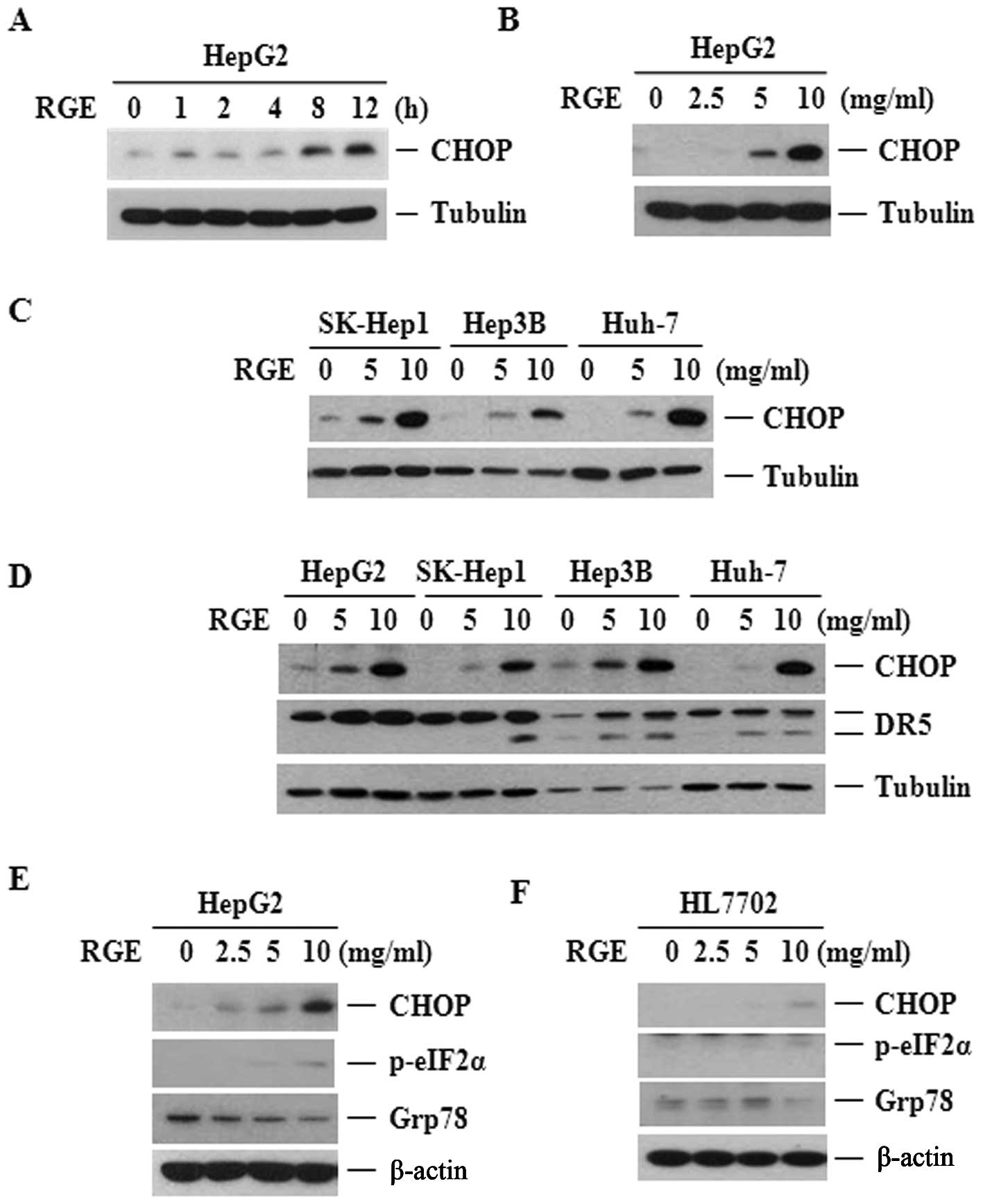
therapy (36–38). The RGE treatment increased CHOP
levels in the HepG2 cell lines corresponding to its sensitization
effect on TRAIL-derived cell death (Fig. 5A–C). As shown in Fig. 5D, RGE-induced CHOP upregulation
preceded the increase in DR5 levels. Such a temporal pattern
further supports the notion that CHOP plays a role in the
activation of DR5 expression by the RGE. As CHOP is a major
transcription factor induced by ER stress, we examined whether RGE
treatment would induce ER stress in HCC. Thus, we probed for two
markers of ER stress: Ser-51 phosphorylation of eIF2α (P-eIF2α) and
induction of the ER chaperone protein GRP78. As shown in Fig. 5E, the RGE upregulated C/EBP
homologous protein but did not induce GRP78 or eIF2α
phosphorylation in the HepG2 cell lines, indicating that the RGE
was capable of over-expressed the CHOP without substantively
affecting ER stress. However, the RGE did not influence CHOP
expression in the normal liver cell lines, which strongly agrees
with the observation that the RGE did not boost the effect of TRAIL
in the normal cell lines (Fig.
3B). Additionally, the RGE was unable to induce expression of
the C/EBP homologous protein or other ER stress markers in the
normal cell lines (Fig. 5F).
When investigating the induction of the TRAIL
receptors DR4 and DR5 in tumors it is very important to assess
susceptibility of the tumor to TRAIL treatment (39). Several studies have shown that high
expression of the TRAIL receptors DR4 and DR5 facilitates
sensitization of cancer cells to TRAIL-derived cell death (1,40).
One therapeutic approach being tested is to induce the expression
of death receptors, including DR4 and particularly DR5, by small
molecules, which results in TRAIL-derived tumor cell death or
sensitization of TRAIL-resistant cell lines to cell death (33). Furthermore, DR5 expression levels
are highly correlated with TRAIL, which binds to its receptors, DR4
and DR5, and activates the extrinsic apoptosis pathway (34,41,42).
In the present study, we found that the improved effects of the
combination therapy were primarily dependent upon control of DR5
induction. We also found that the combination therapy induced
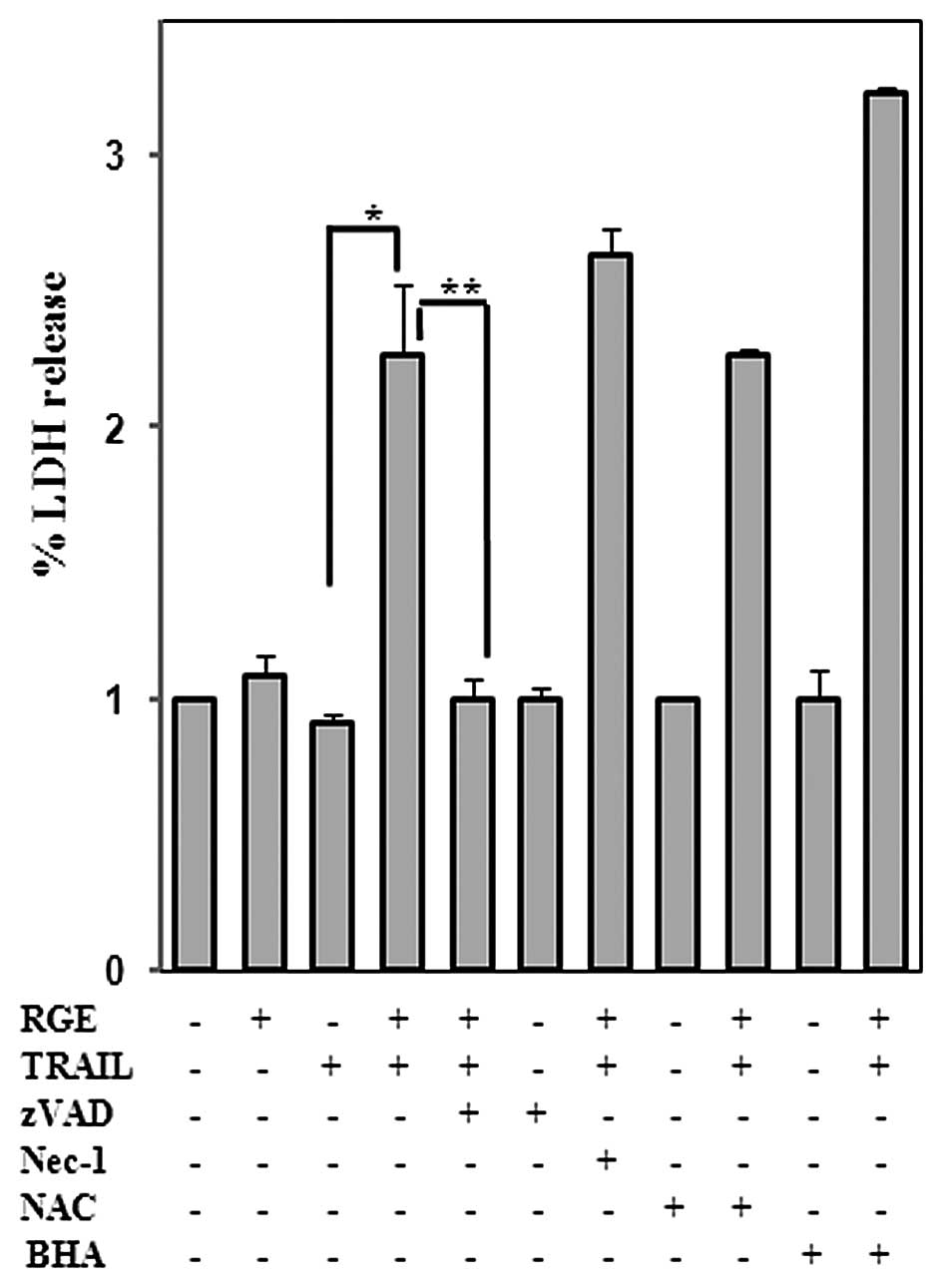
apoptosis in the presence of ROS (Fig.
6), suggesting that ROS do not contribute to RGE-facilitated
sensitization of TRAIL-derived cell death.
Increased expression of CHOP is related
to DR5 upregulation
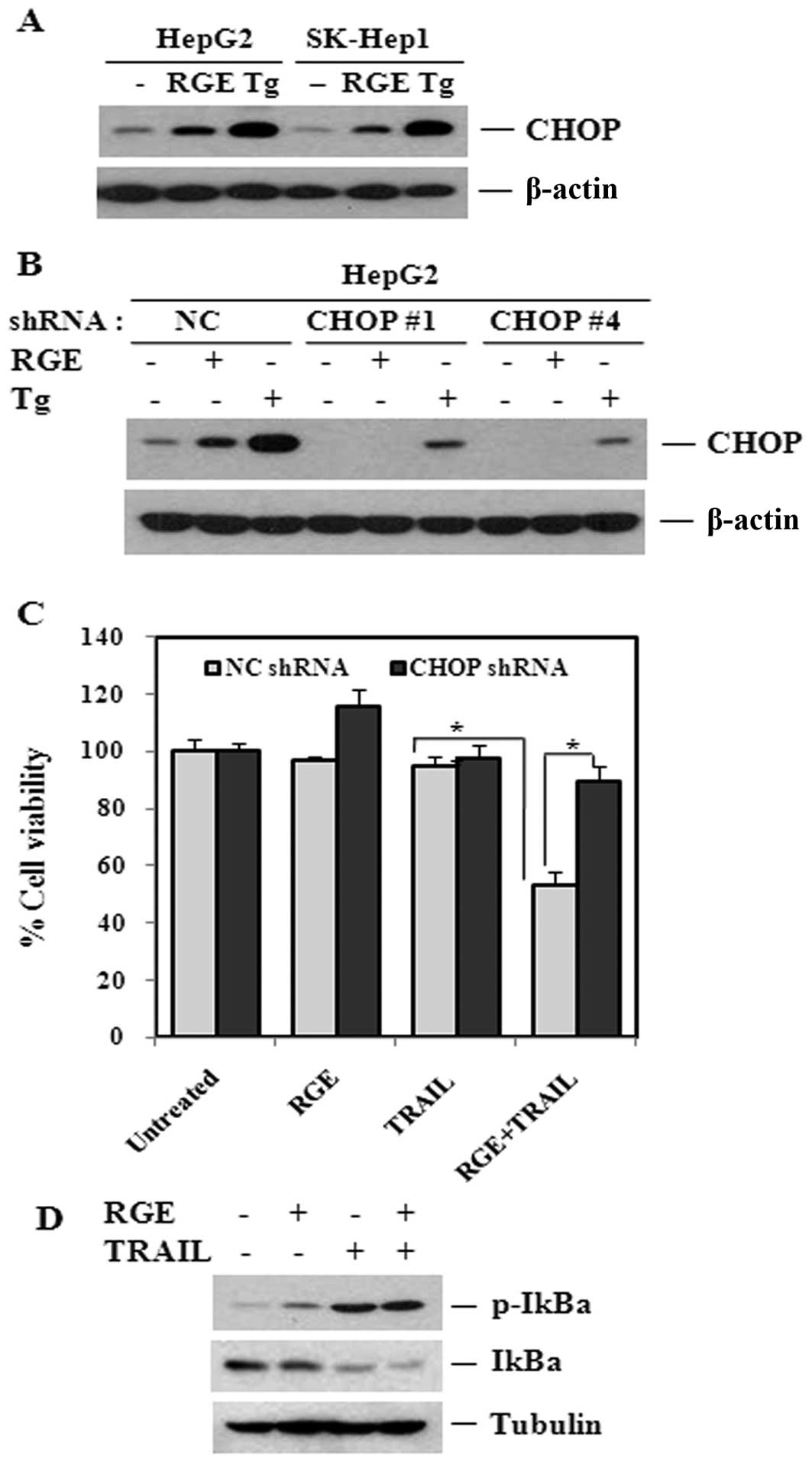
Tg treatment was compared with RGE treatment to
confirm that upregulation of CHOP plays a key role in the RGE
sensitization effect, as shown in Fig.
7A(43,44). Next, we established stable cell
lines for knockdown of the C/EBP homologous protein using a
lentiviral shRNA system. As shown in Fig. 7B, we confirmed knockdown of CHOP.
In addition, knockdown of CHOP inhibited the robust upregulation of
CHOP induced by Tg. We then examined the effect of CHOP knockdown
on RGE-induced sensitization to TRAIL-derived cell death. CHOP
knockdown inhibited the effect of the RGE plus TRAIL treatment in
HepG2 cell lines, suggesting that augmentation of cell death by the
RGE combined treatment in HCC was mediated by CHOP expression
(Fig. 7C).
Activation of NF-κB in cancer cell lines contributes
to the induction of DR5 as well as resistance to TRAIL-derived
apoptosis. Therefore, we examined whether the RGE might regulate
NF-κB to facilitate DR5 upregulation and, consequently, sensitize
cell lines to TRAIL-derived apoptosis. As shown in Fig. 7D, degradation and phosphorylation
of IκBα did not occur in response to the RGE. In addition, the RGE
did not affect TRAIL-derived NF-κB activation, suggesting that
RGE-induced sensitization to cell death is not mediated by NF-κB.
Although TRAIL is capable of activating NF-κB in some cancer cell
lines and the induction of DR5 is linked to NF-κB activation,
activated NF-κB also upregulates the anti-apoptotic gene Bcl-xL,
effectively blocking TRAIL-derived apoptosis (10,27,34).
We failed to detect upregulation of Bcl-xL in response to RGE
treatment (data not shown), which further indicates that NF-κB was
activated by the RGE. Additionally, RGE-induced CHOP upregulation
was not caused by ER stress, which can lead to apoptosis in HCC
cell lines. Inducing CHOP expression or TRAIL sensitization was
unsuccessful when RGE was applied to the normal cell line HL7702,
suggesting that CHOP protein upregulation by the RGE is specific to
the tumor cell lines.
Although we have not tested the antitumor activity
of the RGE in vivo, the formulated RGE examined could
potentially be further developed as an important chemosensitizer to
be utilized as a dietary supplement with anticancer drugs.
Acknowledgements
This study was supported by the 2011
grant from the Korean Society of Ginseng funded by Korea Ginseng
Corporation and the Yujeonja-Donguibogam project based on
Traditional herbs (Grant No. 2012M3A9C4048796), Republic of Korea.
This study was also supported by a National Research Foundation of
Korea (NRF) grant funded by the Korean government (2011-0030834) to
Y.-S. Kim and by the 2010 grant from Department of Medical
Sciences, the Graduate School, Ajou University.
References
|
1.
|
Kischkel FC, Lawrence DA, Chuntharapai A,
Schow P, Kim KJ and Ashkenazi A: Apo2L/TRAIL-dependent recruitment
of endogenous FADD and caspase-8 to death receptors 4 and 5.
Immunity. 12:611–620. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Deveraux QL, Roy N, Stennicke HR, et al:
IAPs block apoptotic events induced by caspase-8 and cytochrome c
by direct inhibition of distinct caspases. EMBO J. 17:2215–2223.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Hu W, Johnson H and Shu H: Tumor necrosis
factor-related apoptosis-inducing ligand receptors signal NF-κB and
JNK activation and apoptosis through distinct pathways. J Biol
Chem. 274:306031999.
|
|
4.
|
Lin B, Williams-Skipp C, Tao Y, et al:
NF-κB functions as both a proapoptotic and antiapoptotic regulatory
factor within a single cell type. Cell Death Differ. 6:570–582.
1999.
|
|
5.
|
Son YG, Kim EH, Kim JY, et al: Silibinin
sensitizes human glioma cells to TRAIL-mediated apoptosis via DR5
up-regulation and down-regulation of c-FLIP and survivin. Cancer
Res. 67:8274–8284. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Ding L, Yuan C, Wei F, et al: Cisplatin
restores TRAIL apoptotic pathway in glioblastoma-derived stem cells
through up-regulation of DR5 and down-regulation of c-FLIP. Cancer
Invest. 29:511–520. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Park SJ, Bijangi-Vishehsaraei K and Safa
AR: Selective TRAIL-triggered apoptosis due to overexpression of
TRAIL death receptor 5 (DR5) in P-glycoprotein-bearing multidrug
resistant CEM/VBL1000 human leukemia cells. Int J Biochem Mol Biol.
1:90–100. 2010.
|
|
8.
|
Fossati S, Ghiso J and Rostagno A: TRAIL
death receptors DR4 and DR5 mediate cerebral microvascular
endothelial cell apoptosis induced by oligomeric Alzheimer’s Aβ.
Cell Death Dis. 3:e3212012.PubMed/NCBI
|
|
9.
|
Griffith TS, Fialkov JM, Scott DL, et al:
Induction and regulation of tumor necrosis factor-related
apoptosis–inducing ligand/Apo-2 ligand-mediated apoptosis in renal
cell carcinoma. Cancer Res. 62:3093–3099. 2002.
|
|
10.
|
Kim JY, Lee JY, Kim DG, Koo GB, Yu JW and
Kim YS: TRADD is critical for resistance to TRAIL-induced cell
death through NF-κB activation. FEBS Lett. 585:2144–2150. 2011.
|
|
11.
|
Tian X, Ye J, Alonso-Basanta M, Hahn SM,
Koumenis C and Dorsey JF: Modulation of CCAAT/enhancer binding
protein homologous protein (CHOP)-dependent DR5 expression by
nelfinavir sensitizes glioblastoma multiforme cells to tumor
necrosis factor-related apoptosis-inducing ligand (TRAIL). J Biol
Chem. 286:29408–29416. 2011. View Article : Google Scholar
|
|
12.
|
Carr BI: Hepatocellular carcinoma: current
management and future trends. Gastroenterology. 127:S218–S224.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Yang JF, Cao JG, Tian L and Liu F:
5,7-Dimethoxyflavone sensitizes TRAIL-induced apoptosis through DR5
upregulation in hepatocellular carcinoma cells. Cancer Chemother
Pharmacol. 69:195–206. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Zhang B, Shan H, Li D, et al: Cisplatin
sensitizes human hepatocellular carcinoma cells, but not
hepatocytes and mesenchymal stem cells, to TRAIL within a
therapeutic window partially depending on the upregulation of DR5.
Oncol Rep. 25:461–468. 2011.
|
|
15.
|
Lee SA, Jo HK, Im BO, Kim SU, Whang WK and
Ko SK: Changes in the Contents of Prosapogenin in the Red Ginseng
(Panax ginseng) Depending on Steaming Batches. J Ginseng Res.
36:102–106. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
In G, Ahn NG, Bae BS, Han ST, Noh KB and
Kim CS: New method for simultaneous quantification of 12
ginsenosides in red ginseng powder and extract: in-house method
validation. J Ginseng Res. 36:205–210. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Sun S, Qi LW, Du GJ, Mehendale SR, Wang CZ
and Yuan CS: Red notoginseng: higher ginsenoside content and
stronger anti-cancer potential than Asian and American ginseng.
Food Chem. 125:1299–1305. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Keum YS, Park KK, Lee JM, et al:
Antioxidant and anti-tumor promoting activities of the methanol
extract of heat-processed ginseng. Cancer Lett. 150:41–48. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Li J, Huang M, Teoh H and Man RY: Panax
quinquefolium saponins protects low density lipoproteins from
oxidation. Life Sci. 64:53–62. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Chang MS, Lee SG and Rho HM:
Transcriptional activation of Cu/Zn superoxide dismutase and
catalase genes by panaxadiol ginsenosides extracted from Panax
ginseng. Phytother Res. 13:641–644. 1999. View Article : Google Scholar
|
|
21.
|
Ji HY, Lee HW, Kim HK, et al: Simultaneous
determination of ginsenoside Rb(1) and Rg(1) in human plasma by
liquid chromatography-mass spectrometry. J Pharm Biomed Anal.
35:207–212. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Cai F, Sun L, Gao S, Yang Y, Yang Q and
Chen W: A rapid and sensitive liquid chromatography-tandem mass
spectrometric method for the determination of timosaponin B-II in
blood plasma and a study of the pharmacokinetics of saponin in the
rat. J Pharm Biomed Anal. 48:1411–1416. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Woo SS, Song JS, Lee JY, et al: Selection
of high ginsenoside producing ginseng hairy root lines using
targeted metabolic analysis. Phytochemistry. 65:2751–2761. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Park JD, Rhee DK and Lee YH: Biological
activities and chemistry of saponins from Panax ginseng CA Meyer.
Phytochem Rev. 4:159–175. 2005. View Article : Google Scholar
|
|
25.
|
Lee DCW and Lau ASY: Effects of Panax
ginseng on tumor necrosis factor-α-mediated inflammation: a
mini-review. Molecules. 16:2802–2816. 2011.
|
|
26.
|
Sayers TJ: Targeting the extrinsic
apoptosis signaling pathway for cancer therapy. Cancer Immunology,
Immunotherapy. 1–8. 2011.
|
|
27.
|
Ravi R, Bedi GC, Engstrom LW, et al:
Regulation of death receptor expression and TRAIL/Apo2L-induced
apoptosis by NF-κB. Nat Cell Biol. 3:409–416. 2001.
|
|
28.
|
Ganten TM, Koschny R, Sykora J, et al:
Preclinical differentiation between apparently safe and potentially
hepatotoxic applications of TRAIL either alone or in combination
with chemotherapeutic drugs. Clin Cancer Res. 12:2640–2646. 2006.
View Article : Google Scholar
|
|
29.
|
Baritaki S, Huerta-Yepez S, Sakai T,
Spandidos DA and Bonavida B: Chemotherapeutic drugs sensitize
cancer cells to TRAIL-mediated apoptosis: up-regulation of DR5 and
inhibition of Yin Yang 1. Mol Cancer Ther. 6:1387–1399. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Mellier G, Huang S, Shenoy K and Pervaiz
S: TRAILing death in cancer. Mol Aspects Med. 31:93–112. 2010.
View Article : Google Scholar
|
|
31.
|
Kouhara J, Yoshida T, Nakata S, et al:
Fenretinide upregulates DR5/TRAIL-R2 expression via the induction
of the transcription factor CHOP and combined treatment with
fenretinide and TRAIL induces synergistic apoptosis in colon cancer
cell lines. Int J Oncol. 30:679–687. 2007.
|
|
32.
|
Bertram H, Nerlich A, Omlor G, Geiger F,
Zimmermann G and Fellenberg J: Expression of TRAIL and the death
receptors DR4 and DR5 correlates with progression of degeneration
in human intervertebral disks. Mod Pathol. 22:895–905. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Xiaowen H and Yi S: Triptolide sensitizes
TRAIL-induced apoptosis in prostate cancer cells via p53-mediated
DR5 up-regulation. Mol Biol Rep. 39:8763–8770. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Lin FL, Hsu JL, Chou CH, Wu WJ, Chang CI
and Liu HJ: Activation of p38 MAPK by damnacanthal mediates
apoptosis in SKHep 1 cells through the DR5/TRAIL and TNFR1/TNF-α
and p53 pathways. Eur J Pharmacol. 650:120–129. 2011.PubMed/NCBI
|
|
35.
|
Deng Z, Yan H, Hu J, et al: Hepatitis C
virus sensitizes host cells to TRAIL-induced apoptosis by
up-regulating DR4 and DR5 via a MEK1-dependent pathway. PloS One.
7:e377002012. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Shiraishi T, Yoshida T, Nakata S, et al:
Tunicamycin enhances tumor necrosis factor-related
apoptosis-inducing ligand-induced apoptosis in human prostate
cancer cells. Cancer Res. 65:6364–6370. 2005. View Article : Google Scholar
|
|
37.
|
Chen CY, Yiin SJ, Hsu JL, Wang WC, Lin SC
and Chern CL: Isoobtusilactone A sensitizes human hepatoma Hep G2
cells to TRAIL-induced apoptosis via ROS and CHOP-mediated
up-regulation of DR5. J Agric Food Chem. 60:3533–3539. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Oyadomari S and Mori M: Roles of
CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ.
11:381–389. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Bin L, Thorburn J, Thomas LR, Clark PE,
Humphreys R and Thorburn A: Tumor-derived mutations in the TRAIL
receptor DR5 inhibit TRAIL signaling through the DR4 receptor by
competing for ligand binding. J Biol Chem. 282:28189–28194. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Sheridan JP, Marsters SA, Pitti RM, et al:
Control of TRAIL-induced apoptosis by a family of signaling and
decoy receptors. Science. 277:818–821. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Seol DW: p53-Independent up-regulation of
a TRAIL receptor DR5 by proteasome inhibitors: a mechanism for
proteasome inhibitor-enhanced TRAIL-induced apoptosis. Biochem
Biophys Res Commun. 416:222–225. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42.
|
Sung B, Ravindran J, Prasad S, Pandey MK
and Aggarwal BB: Gossypol induces death receptor-5 through
activation of the ROS-ERK-CHOP pathway and sensitizes colon cancer
cells to TRAIL. J Biol Chem. 285:35418–35427. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43.
|
Zinszner H, Kuroda M, Wang X, et al: CHOP
is implicated in programmed cell death in response to impaired
function of the endoplasmic reticulum. Genes Dev. 12:982–995. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
44.
|
Zou W, Yue P, Khuri F and Sun S: Coupling
of endoplasmic reticulum stress to CDDO-Me-induced up-regulation of
death receptor 5 via a CHOP-dependent mechanism involving JNK
activation. Cancer Res. 68:7484–7492. 2008. View Article : Google Scholar : PubMed/NCBI
|