Introduction
Oral squamous cell carcinoma (OSCC) is a frequently
occurring neoplasm that is usually aggressive and has a poor
prognosis. Improvements in specificity and sensitivity of diagnosis
and prognosis depend on the elucidation of the biologic and
molecular mechanisms underlying carcinogenesis (1). The availability of biomarkers of
malignancy would be a key factor for monitoring cancer recurrence
and evaluating the efficacy of novel treatments. The accumulation
of genetic alterations during carcinogenesis is currently known but
still largely unexplored (2–5).
We previously reported gene expression profiling of
OSCCs using microarray analysis to identify genes associated with
oral carcinogenesis (6). The
analysis showed that protein O-fucosyltransferase 1 (POFUT1)
expression was significantly upregulated.
POFUT1 attaches O-fucose through
O-glycosidic linkage to conserved serine or threonine
residues in the epidermal growth factor-like (EGF-like) repeats of
a number of cellular surface and secreted proteins (7). In the present study, further analysis
of the status of POFUT1 expression showed that POFUT1 expression
increased in OSCCs compared with normal oral tissues. Based on
these data, we proposed that POFUT1 may be a key regulator of tumor
progression in OSCCs.
Materials and methods
OSCC-derived cell lines and tissue
samples
The human OSCC cell lines (Ho-1-u-1, KOSC2, HSC-4,
HSC-3 and HSC-2) were purchased from the Human Science Research
Resources Bank, Osaka, Japan. Primary cultured human normal oral
keratinocytes (HNOKs) served as normal controls (8–17).
All cells were maintained in Dulbecco’s modified Eagle’s medium
(DMEM)/F-12 Ham (Sigma, St. Louis, MO, USA) supplemented with 10%
fetal bovine serum (FBS) (Sigma) and 50 U/ml penicillin and
streptomycin (Sigma) in a humidified 5% CO2/air
atmosphere at 37°C (18–20).
One hundred and twenty-eight pairs of primary OSCC
samples and corresponding normal oral epithelial tissues were
obtained intraoperatively at Chiba University Hospital, Chiba,
Japan. All patients provided informed consent for use of the
protocol, which the institutional review board of Chiba University
reviewed and approved. The resected tissues were divided into two
parts, one of which was frozen immediately and stored at −80°C
until RNA isolation and the second was fixed in 20% buffered
formaldehyde solution for pathologic diagnosis and
immunohistochemistry (IHC). Histopathological diagnosis of each
tissue was performed according to the World Health Organization
criteria by the Department of Pathology, Chiba University Hospital.
Clinicopathological staging was determined according to the
tumor-node-metastases classification of the International Union
against Cancer. All OSCC samples were confirmed histologically and
checked to ensure the presence of tumor in >90% of
specimens.
Preparation of cDNA
Total RNA was isolated using TRIzol reagent
(Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s
instructions. cDNA was generated from 5 μg of total RNA
using Ready-To-Go You-Prime First-Strand Beads (GE Healthcare,
Buckinghamshire, UK) and oligo(dT) primer (Hokkaido System Science,
Sapporo, Japan) according to the manufacturer’s instructions.
mRNA expression analysis
Real-time quantitative reverse
transcriptase-polymerase chain reaction (qRT-PCR) was performed to
evaluate the expression level of POFUT1 mRNA in the OSCC-derived
cell lines and HNOKs. Primary OSCCs and paired specimens of normal
oral tissues obtained from 128 patients also were evaluated. The
expression levels were determined using primers and probes that
were designed using the Universal Probe Library (Roche Diagnostics,
Mannheim, Germany) following the manufacturer’s recommendations.
The primer sequences used for qRT-PCR were: POFUT1, forward
5′-CTGATGACCCGATGGTAAGC-3′; reverse 5′-AAG CCTCCTTTCACCAACCT-3′;
and universal probe no. 78. All qRT-PCR analyses were performed
using a LightCycler® 480 PCR system (Roche Diagnostics).
Amplifications were initiated by a 10-min pre-incubation at 95°C,
followed by 45 cycles of 10 sec at 95°C for template denaturation,
30 sec at 55°C for primer annealing/extension and cooling for 30
sec at 40°C. The transcript amount for POFUT1 was estimated from
the respective standard curves and normalized to the
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (forward,
5′-CAT CTCTGCCCCCTCTGCTGA-3′; reverse, 5′-GGATGACCTT
GCCCACAGCCT-3′; and universal probe no. 60) transcript amount
determined in corresponding samples.
Protein extraction
The cells were washed twice with cold phosphate
buffered saline (PBS) and centrifuged at 500 × g. The cell pellets
were incubated at 4°C for 30 min in a lysis buffer [7 M urea, 2 M
thiourea, 4% w/v CHAPS and 10 mM Tris (pH 7.4)] with the proteinase
inhibitor cocktail (Roche Diagnostics). The protein concentration
was measured with a protein assay (Bio-Rad Laboratories, Hercules,
CA, USA).
Western blot analysis
Cytoplasmic and nuclear proteins (50 μg) were
separated by sodium dodecyl sulfate polyacrylamide gel
electrophoresis in 4–12% gel, transferred to nitrocellulose
membranes and blocked for 1 h at room temperature in Blocking One
(Nacalai Tesque, Tokyo, Japan). The membranes were incubated with
anti-POFUT1 polyclonal antibody (OriGene Technologies, Rockville,
MD, USA) and GAPDH (Santa Cruz Biotechnology, Santa Cruz, CA, USA)
antibodies for 4 h at room temperature. The membranes were washed
with 0.1% Tween-20 in Tris-buffered saline, incubated with
secondary antibodies and coupled to horseradish
peroxidase-conjugated anti-rabbit IgG (Promega, Madison, WI, USA)
in Blocking One for 1 h at room temperature. The proteins were
detected by SuperSignal Chemiluminescent substrate (Thermo,
Waltham, MA, USA). Finally, immunoblotting results were visualized
by exposing the membrane to a cooled CCD camera system (Light
Capture®, ATTO, Tokyo, Japan). Signal intensities were
quantitated using the CS Analyzer® version 3.0
(ATTO).
IHC
IHC was performed on 4-μm sections of
paraffin-embedded specimens using rabbit anti-POFUT1 polyclonal
antibody (OriGene Technologies). Briefly, after deparaffinization
and hydration, the endogenous peroxidase activity was quenched by
30-min incubation in a mixture of 0.3% hydrogen peroxide solution
in 100% methanol. The sections were blocked for 2 h at room
temperature with 1.5% blocking serum (Santa Cruz Biotechnology) in
PBS before reacting with anti-POFUT1 antibody (1:1,000 dilution) at
4°C in a moist chamber overnight. Upon incubation with the primary
antibody, the specimens were washed three times in PBS and treated
with Envision reagent (Dako, Carpinteria, CA, USA) followed by
color development in 3,3′-diaminobenzidine tetrahydrochloride
(Dako). Finally, the slides were counterstained lightly with
hematoxylin, dehydrated with ethanol, cleaned with xylene and
mounted. Non-specific binding of the antibody to proteins other
than the antigen sometimes occurred. As a negative control,
triplicate sections were immunostained without exposure to primary
antibody, which confirmed the staining specificity. To quantify the
state of POFUT1 protein expression in those components, we used IHC
scoring systems described previously (6,8–17).
The intensity of the POFUT1 immunoreaction was scored as follows:
1+, weak; 2+, moderate; and 3+, intense. The cell numbers and the
staining intensity then were multiplied to produce the POFUT1 IHC
score. Cases with a POFUT1 IHC score exceeding 71.5 [the maximal
score within +3 standard deviations (SD) of the mean of normal
tissues] were defined as POFUT1-positive. The ±3 SD cutoff, which
statistically is just 0.2% of the measurement and expected to fall
outside this range, was used because it was unlikely to be affected
by a random experimental error produced by sample manipulation
(21). Two independent
pathologists, neither of whom had knowledge of the patients’
clinical status, made these judgments.
Transfection with shRNA plasmid
The OSCC cell lines, KOSC2 and HSC-2, in which
POFUT1 protein expression was higher than in the other cell lines,
were stably transfected with POFUT1 shRNA (shPOFUT1) or control
shRNA (mock) (Santa Cruz Biotechnology) using Lipofectamine LTX and
Plus Reagents (Invitrogen). The stable transfectants were isolated
by the culture medium containing 1 mg/ml puromycin (Invitrogen).
Two to 3 weeks after transfection, viable colonies were picked up
and transferred to new dishes. shPOFUT1- and mock-transfected cells
were used for further experiments.
Proliferation assays
To investigate the effect of POFUT1 knockdown on
cellular proliferation, shPOFUT1- and mock-transfected cells were
seeded in 6-well plates at a density of 1×104 viable
cells/well. At the indicated time-points, the cells were
trypsinized and counted in triplicate using a hemocytometer and
Cell Counting Kit-8 (Dojindo Molecular Technologies, Kumamoto,
Japan) (22). Images were obtained
every 12 h using a Leica LCD microscope (Leica Microsystems,
Wetzlar, Germany) (original magnification, ×200) and were
representative of three independent experiments.
Migration assay
To evaluate the effect of POFUT1 knockdown on
migration, we performed the wound-healing assay described
previously (17). Briefly, after
uniform wounds were made in confluent culture of the shPOFUT1 and
mock cells, the extent of closure was monitored visually every 4 h
for 24 h. The results were visualized by measuring the wound spaces
using Lenalaf220 software (available at http://www.vector.co.jp/download/file/win95/art/fh442375.html).
The mean value was calculated from data obtained from three
separate chambers.
Invasiveness assay
To evaluate the effect of POFUT1 knockdown on
invasiveness, we performed an invasiveness assay (23). A total of 2.5×105 cells
were seeded on a polyethylene terephthalate membrane insert (pore
size, 3 μm) in a transwell apparatus (Becton-Dickinson,
Franklin Lakes, NJ, USA). In the lower chamber, 1 ml of DMEM with
10% FBS and puromycin (Invitrogen) were added as chemoattractants.
After the cells were incubated for 22 h at 37°C, the insert was
washed with PBS and the cells on the top of the insert surface were
removed with a cotton swab. Cells adhering to the lower surface of
the membrane were fixed with methanol and stained with crystal
violet; the numbers of cells invading the pores in five random
fields were counted using a light microscope at ×100
magnification.
Statistical analysis
The significance of the POFUT1 expression levels was
evaluated using Fisher’s exact test or the Mann-Whitney U test.
P<0.05 was considered statistically significant. The data are
expressed as the mean ± standard error of the mean (SEM).
Results
Evaluation of POFUT1 expression in
OSCC-derived cell lines
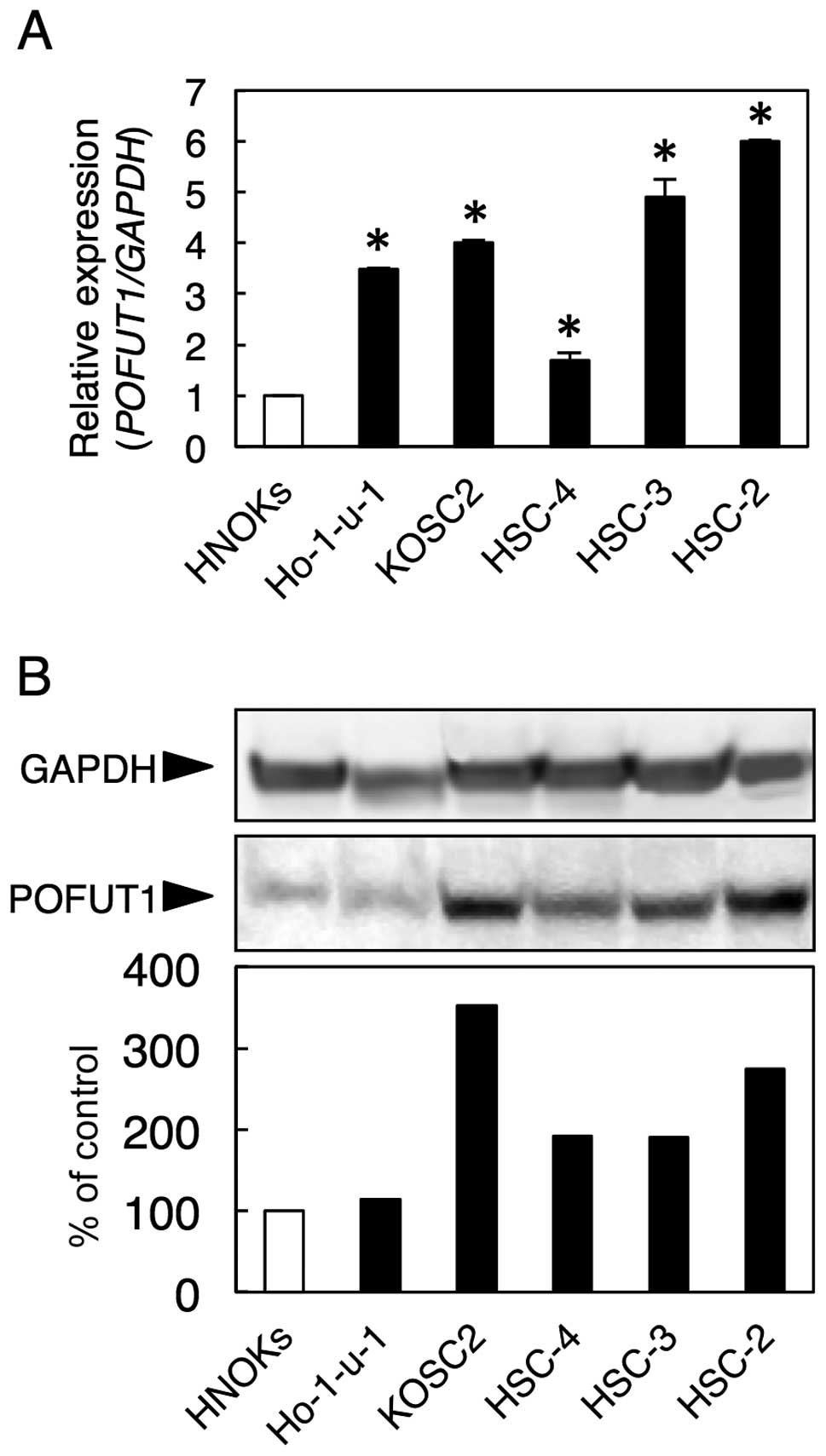
We examined POFUT1 mRNA and protein expression in
five OSCC-derived cell lines (Ho-1-u-1, KOSC2, HSC-4, HSC-3 and
HSC-2) and HNOKs using qRT-PCR and western blot analysis.
Significant upregulation of POFUT1 mRNA occurred in all OSCC cell
lines compared with the HNOKs (Fig.
1A, P<0.05).
Fig. 1B shows
representative results of the western blot analyses. The values
obtained from densitometric analysis of POFUT1 protein were
normalized to the GAPDH levels and then expressed as a percentage
of the HNOK values. A significant increase in POFUT1 protein
expression was seen in all OSCC cell lines compared with the HNOKs
(Fig. 1B).
Evaluation of POFUT1 expression in
primary OSCCs
We evaluated POFUT1 mRNA expression in the normal
oral tissues and paired primary OSCCs from 128 patients. Similar to
the data obtained from the OSCC-derived cell lines, qRT-PCR showed
that POFUT1 mRNA expression was upregulated in 104 (81.2%) of 128
primary OSCCs compared with the matched normal oral tissues (data
not shown).
Representative IHC results for POFUT1 protein in
normal oral tissue and primary OSCC are shown in Fig. 2A. The POFUT1 IHC scores of the
cytoplasm in primary OSCCs were significantly (P<0.05) higher
than in normal tissues. The IHC scores in the normal oral tissues
and OSCCs ranged from 76.2 to 180.0 (median, 165.8) and 112.5 to
225.5 (median, 192.4), respectively (Fig. 2B). The POFUT1 IHC scores of T3/T4
were significantly (P<0.05) higher than that of T1/T2. The IHC
scores for T1/T2 and T3/T4 ranged from 77.8 to 215 (median, 189.5)
and 149.8 to 240 (median, 204.5), respectively (Fig. 2C). Table I shows the correlations between the
clinicopathologic characteristics of the patients with OSCC and the
status of POFUT1 protein expression using the IHC scoring system.
Among the clinical classifications, POFUT1-positive OSCCs were
correlated with primary tumor size (P=0.048) (Table I).
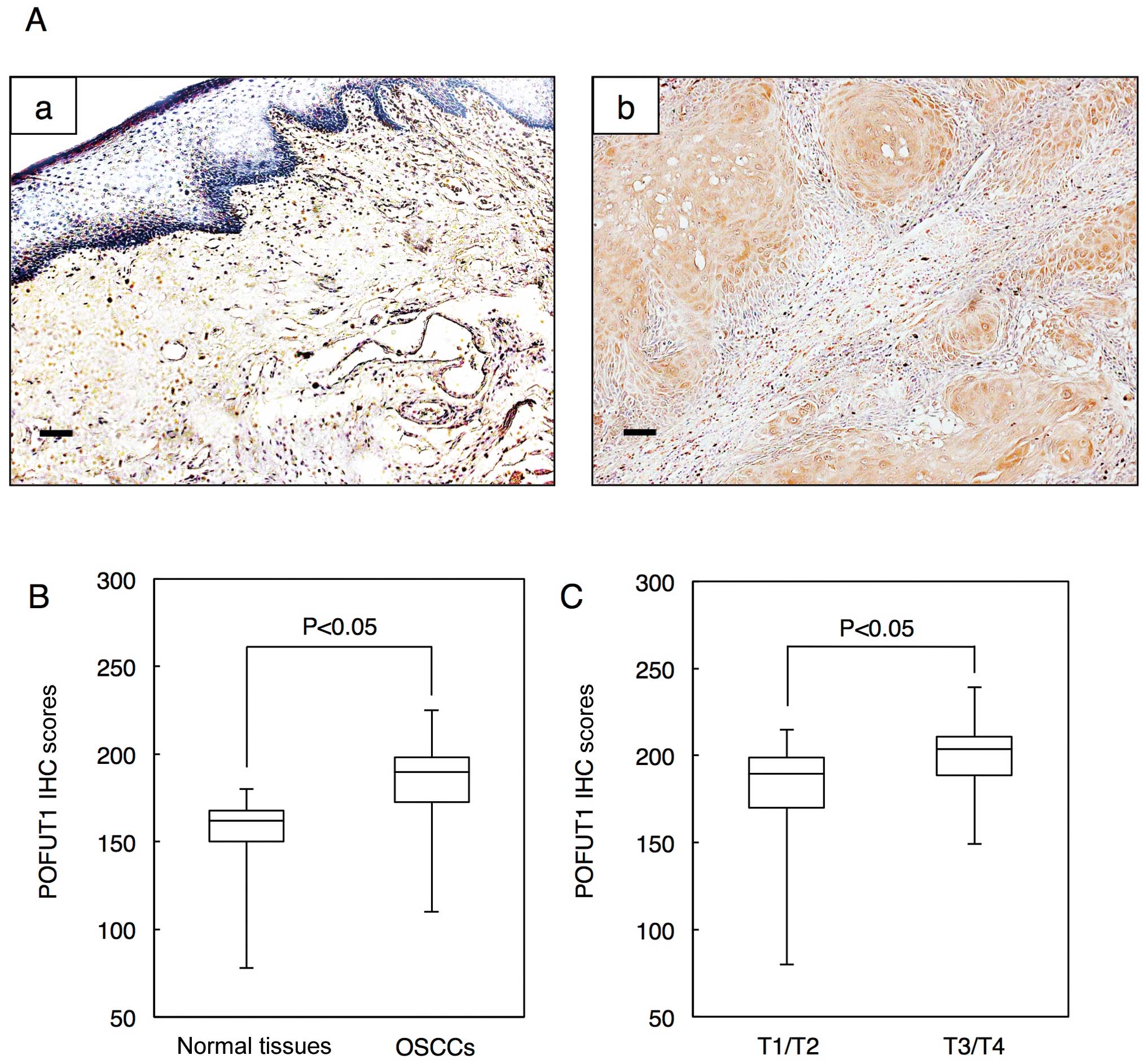 | Figure 2.Evaluation of POFUT1 protein
expression in primary OSCCs. (A) Representative IHC results of
POFUT1 in normal oral tissue and primary OSCC. (a) Normal oral
tissues exhibit negative POFUT1 protein expression. Skeletal muscle
tissues are immunostained for POFUT1 (positive control). Original
magnification, ×100. Scale bars, 50 μm. (b) Positive
immunoreactivity for POFUT1 in OSCCs is detected in the cytoplasm.
Original magnification, ×100. Scale bars, 50 μm. (B) State
of POFUT1 protein expression in normal oral tissues (n=128) and
primary OSCCs (n=128). The POFUT1 IHC scores are calculated as
follows: IHC score = 1 × (number of weakly stained cells in the
field) + 2 × (number of moderately stained cells in the field) + 3
× (number of intensely stained cells in the field). The POFUT1 IHC
scores for normal oral tissues and OSCCs range from 76.2 to 180.0
(median, 165.8) and 112.5 to 225.5 (median, 192.4), respectively.
The POFUT1 protein expression level in OSCCs is significantly
(P<0.05, Mann-Whitney U test) higher than that in normal oral
tissues. (C) The POFUT1 IHC scores for T3/T4 (149.8 to 240; median,
204.5) are significantly (P<0.05, Mann-Whitney U test) higher
than those of T1/T2 (77.8 to 215; median, 189.5). |
 | Table I.Correlation between expression of
POFUT1 and clinical parameters in OSCCs. |
Table I.
Correlation between expression of
POFUT1 and clinical parameters in OSCCs.
| No. of patients
|
|---|
| Results of
immunostaining
| |
|---|
| Clinical
parameters | Total | POFUT1-negative | POFUT1-positive | P-value |
|---|
| Age at surgery
(years) | | | | |
| <60 | 52 | 14 | 38 | 0.767 |
| 60–70 | 50 | 11 | 39 | |
| >70 | 26 | 5 | 21 | |
| Gender | | | | |
| Male | 70 | 15 | 55 | 0.676 |
| Female | 58 | 15 | 43 | |
| T-primary tumor
size | | | | |
| T1 | 27 | 7 | 20 | 0.048a |
| T2 | 49 | 17 | 32 | |
| T3 | 28 | 4 | 24 | |
| T4 | 24 | 2 | 22 | |
| T1/T2 | 76 | 24 | 52 | 0.010a |
| T3/T4 | 52 | 6 | 46 | |
| N-regional lymph
node metastasis | | | | |
| N(+) | 68 | 15 | 53 | 0.835 |
| N(−) | 60 | 15 | 45 | |
| Stage | | | | |
| I | 29 | 5 | 24 | 0.128 |
| II | 42 | 14 | 28 | |
| III | 29 | 8 | 21 | |
| IV | 28 | 3 | 25 | |
| I/II | 71 | 19 | 52 | 0.403 |
| III/IV | 57 | 11 | 46 | |
| Histopathological
type | | | | |
| Well
differentiated | 68 | 19 | 49 | 0.400 |
| Moderately
differentiated | 55 | 10 | 45 | |
| Poorly
differentiated | 5 | 1 | 4 | |
| Tumor site | | | | |
| Tongue | 46 | 10 | 36 | 0.831 |
| Gingiva | 38 | 11 | 27 | |
| Buccal
mucosa | 23 | 6 | 17 | |
| Palate | 14 | 2 | 12 | |
| Oral floor | 7 | 1 | 6 | |
| Total | 128 | 30 | 98 | |
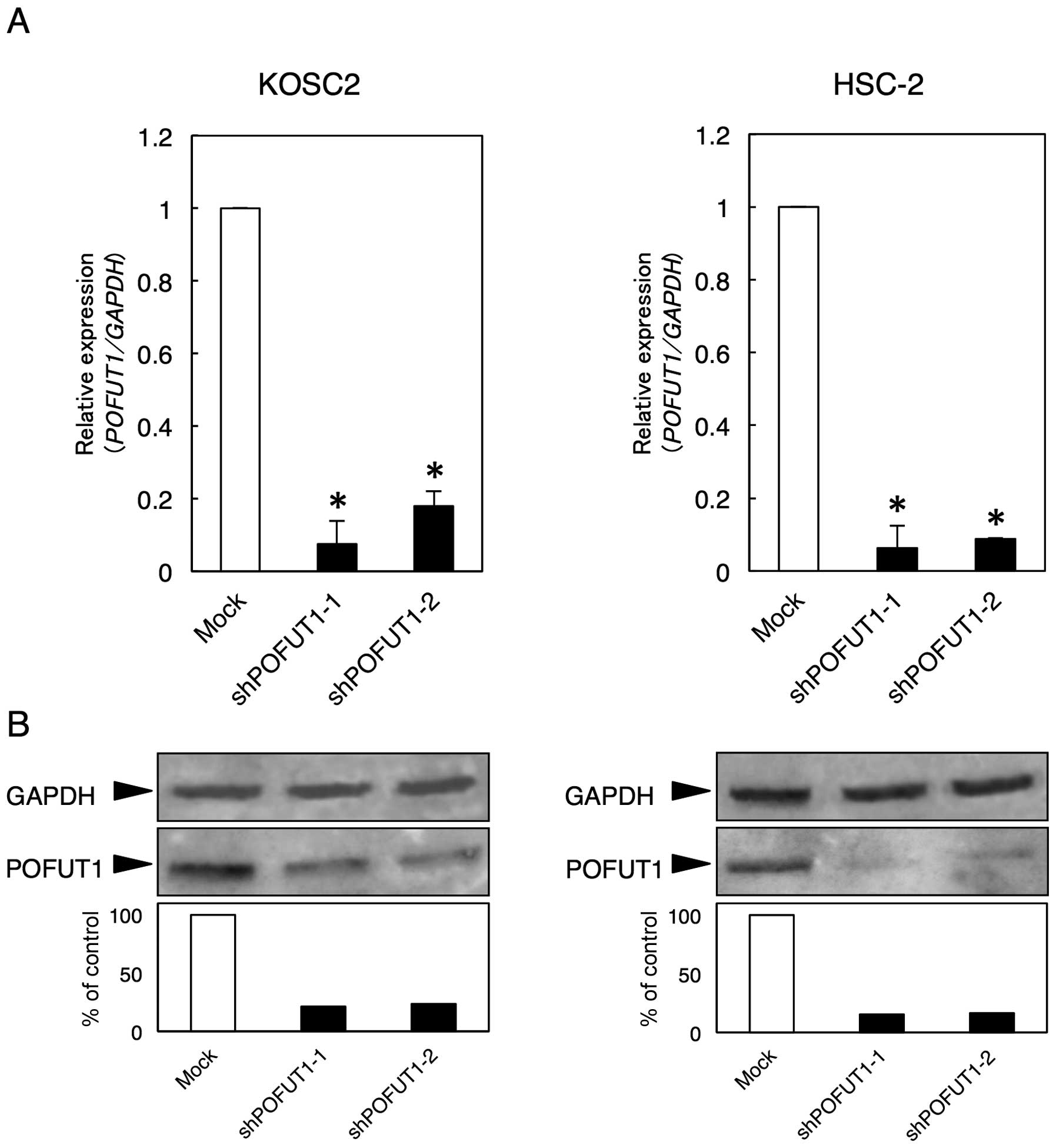
Establishment of POFUT1 knockdown
cells
Since POFUT1 expression was upregulated in the OSCC
cell lines, we assumed that POFUT1 might play an important role in
OSCCs. To assess the POFUT1 functions in vitro, an shRNA
experiment was performed using the KOSC2 and HSC-2 cells.
Expressions of POFUT1 mRNA and protein in the shPOFUT1-transfected
cells were significantly (P<0.05) lower than those in the
mock-transfected cells (KOSC2 and HSC-2 derived transfectant cells)
(Fig. 3).
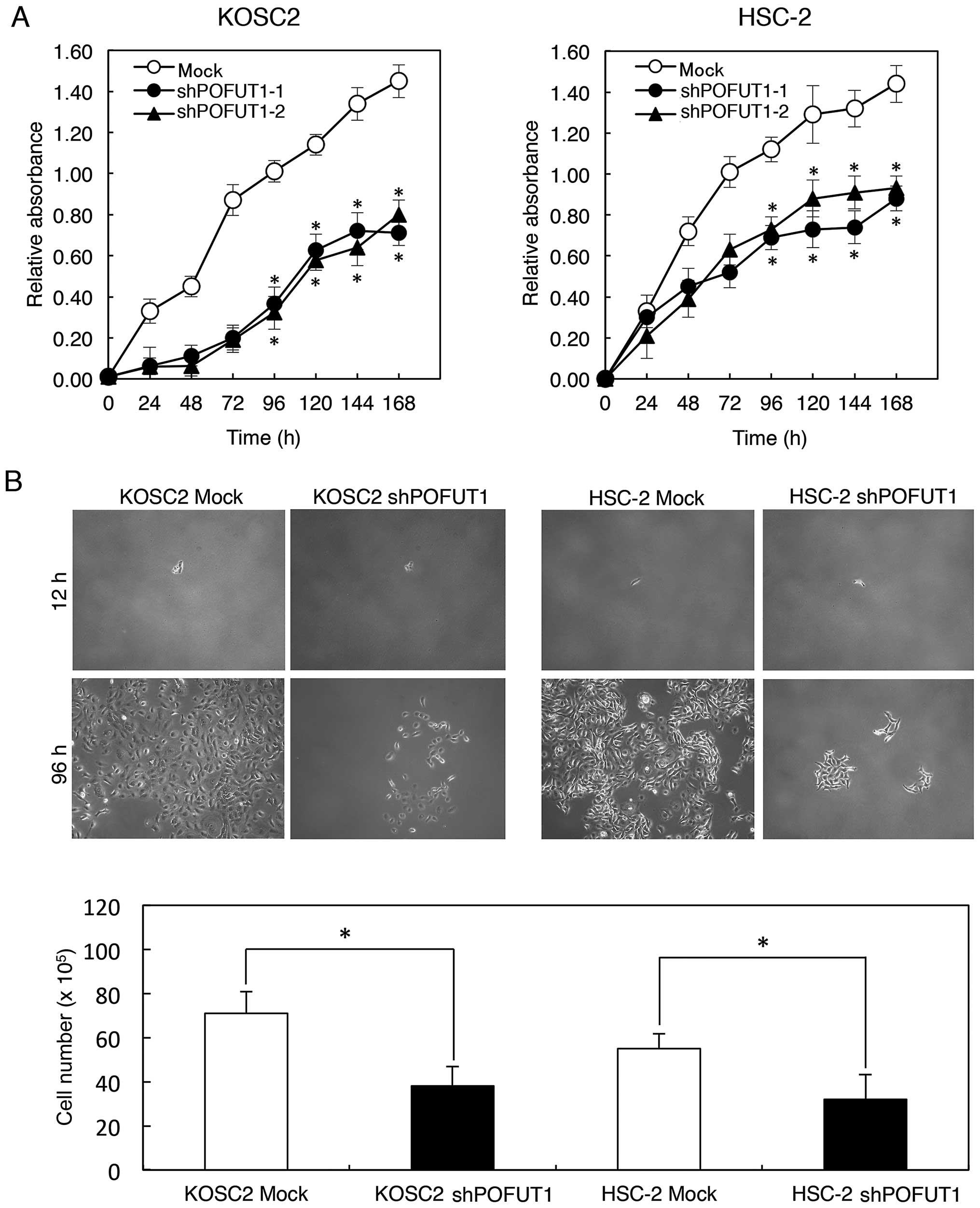
Functional analysis of POFUT1 knockdown
cells
To evaluate the effect of POFUT1 knockdown on
cellular growth, we performed a cellular proliferation assay.
shPOFUT1- and mock-transfected cells were seeded in 6-well plates
at a density of 1×104 viable cells/well counted on 7
consecutive days. There was a significant (P<0.05) decrease in
cellular growth of the shPOFUT1-transfected cells compared with the
mock-transfected cells (KOSC2 and HSC-2 derived transfectant cells)
(Fig. 4A). Photomicrographs
(Fig. 4B) showed the cytostatic
effect (P<0.05) of cell line-transfected shPOFUT1 and mock
cells. In the migration assay, when we visually monitored the
uniform wound area in confluent cell culture, the wound areas in
POFUT1 knockdown cells were the same as that in the mock cells. In
the invasiveness assay, the number of penetrating POFUT1 knockdown
cells was similar to that of the mock cells (data not shown).
Discussion
We previously reported the gene expression profiling
of OSCC to identify genes associated with oral carcinogenesis
(6). Using microarray analysis,
POFUT1 was one of the most markedly upregulated genes in
OSCC-derived cell lines.
POFUT1, located on chromosome 20q11.21, produces an
O-fucose modification on EGF-like repeats of a number of
cellular surface and secreted proteins (7). Recent studies have reported that
POFUT1 affected anterior-posterior somite patterning in mammalian
embryos (24) and regulated T-,
myeloid- and B-lineage differentiation in mammals (25). Several laboratories have reported
the relationship between POFUT1 expression and malignancies, such
as acute myeloid leukemia, myeloid dysplastic syndrome,
glioblastoma and colon cancer (26–29).
Taken together, we anticipated that POFUT1 may play an important
role in various tumoral behavior.
The present study showed significant upregulation of
POFUT1 levels in OSCCs and five OSCC-derived cell lines compared
with the matched normal counterparts. We investigated the
relationship between POFUT1 expression and clinical behavior of
patients with OSCC. The POFUT1 protein expression levels in primary
OSCCs were associated significantly with tumoral size. The POFUT1
IHC scores of T3/T4 were much higher than those of T1/T2. These
results suggested a strong association between POFUT1 and
progression of OSCC. We also examined the effect of POFUT1 on
cellular proliferation. POFUT1 knockdown in OSCC-derived cells
resulted in obvious reduction of growth. A migration assay showed
that the uniform wound area in POFUT1 knockdown cell was the same
as that in the mock cells. An invasiveness assay indicated that the
number of penetrating POFUT1 knockdown cells was similar to that of
the mock cells. These results are consistent with the association
between POFUT1 level and lymph node metastasis in OSCC (Table I).
In conclusion, POFUT1 is overexpressed frequently in
OSCCs and could play an important role in the course of cellular
proliferation. POFUT1 is likely to be not only a molecular marker
for tumoral growth but also an efficacious treatment strategy for
preventing progression in OSCCs.
Acknowledgements
We thank Ms. Lynda C. Charters for
editing this manuscript.
References
|
1.
|
Severino P, Alvares AM, Michaluart P Jr,
Okamoto OK, Nunes FD, Moreira-Filho CA and Tajara EH: Global gene
expression profiling of oral cavity cancers suggests molecular
heterogeneity within anatomic subsites. BMC Res Notes. 1:113–121.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Scully C, Field JK and Tanzawa H: Genetic
aberrations in oral or head and neck squamous cell carcinoma
(SCCHN): 1. Carcinogen metabolism, DNA repair and cell cycle
control. Oral Oncol. 36:256–263. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Scully C, Field JK and Tanzawa H: Genetic
aberrations in oral or head and neck squamous cell carcinoma 3:
clinicopathological applications. Oral Oncol. 36:404–413. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Sticht C, Freier K, Knöpfle K,
Flechtenmacher C, Pungs S, Hofele C, Hahn M, Joos S and Lichter P:
Activation of MAP kinase signaling through ERK5 but not ERK1
expression is associated with lymph node metastases in oral
squamous cell carcinoma (OSCC). Neoplasia. 10:462–470.
2008.PubMed/NCBI
|
|
5.
|
Macfarlane GJ, Zheng T, Marshall JR,
Boffetta P, Niu S, Brasure J, Merletti F and Boyle P: Alcohol,
tobacco, diet and the risk of oral cancer: a pooled analysis of
three case-control studies. Eur J Cancer B Oral Oncol. 31B:181–187.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Yamano Y, Uzawa K, Shinozuka K, Fushimi K,
Ishigami T, Nomura H, Ogawara K, Shiiba M, Yokoe H and Tanzawa H:
Hyaluronan-mediated motility: a target in oral squamous cell
carcinoma. Int J Oncol. 32:1001–1009. 2008.PubMed/NCBI
|
|
7.
|
Wang Y, Lee GF, Kelley RF and Spellman MW:
Identification of a GDP-L-fucose: polypeptide fucosyltransferase
and enzymatic addition of O-linked fucose to EGF domains.
Glycobiology. 6:837–842. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Koike H, Uzawa K, Grzesik WJ, Seki N, Endo
Y, Kasamatsu A, Yamauchi M and Tanzawa H: GLUT1 is highly expressed
in cementoblasts but not in osteoblasts. Connect Tissue Res.
46:117–124. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Kasamatsu A, Uzawa K, Nakashima D, Koike
H, Shiiba M, Bukawa H, Yokoe H and Tanzawa H: Galectin-9 as a
regulator of cellular adhesion in human oral squamous cell
carcinoma cell lines. Int J Mol Med. 16:269–273. 2005.PubMed/NCBI
|
|
10.
|
Endo Y, Uzawa K, Mochida Y, Shiiba M,
Bukawa H, Yokoe H and Tanzawa H: Sarcoendoplasmic reticulum Ca(2+)
ATPase type 2 downregulated in human oral squamous cell carcinoma.
Int J Cancer. 110:225–231. 2004.
|
|
11.
|
Sakuma K, Kasamatsu A, Yamatoji M, Yamano
Y, Fushimi K, Iyoda M, Ogoshi K, Shinozuka K, Ogawara K, Shiiba M,
Tanzawa H and Uzawa K: Expression status of Zic family member 2 as
a prognostic marker for oral squamous cell carcinoma. J Cancer Res
Clin Oncol. 136:553–559. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Yamatoji M, Kasamatsu A, Kouzu Y, Koike H,
Sakamoto Y, Ogawara K, Shiiba M, Tanzawa H and Uzawa K:
Dermatopontin: a potential predictor for metastasis of human oral
cancer. Int J Cancer. 130:2903–2911. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Yamatoji M, Kasamatsu A, Yamano Y, Sakuma
K, Ogoshi K, Iyoda M, Shinozuka K, Ogawara K, Takiguchi Y, Shiiba
M, Tanzawa H and Uzawa K: State of homeobox A10 expression as a
putative prognostic marker for oral squamous cell carcinoma. Oncol
Rep. 23:61–67. 2010.PubMed/NCBI
|
|
14.
|
Kato Y, Uzawa K, Yamamoto N, Kouzu Y,
Koike H, Shiiba M, Bukawa H, Yokoe H, Shibahara T and Tanzawa H:
Overexpression of Septin1: possible contribution to the development
of oral cancer. Int J Oncol. 31:1021–1028. 2007.PubMed/NCBI
|
|
15.
|
Kouzu Y, Uzawa K, Koike H, Saito K,
Nakashima D, Higo M, Endo Y, Kasamatsu A, Shiiba M, Bukawa H, Yokoe
H and Tanzawa H: Overexpression of stathmin in oral squamous-cell
carcinoma: correlation with tumour progression and poor prognosis.
Br J Cancer. 94:717–723. 2006.PubMed/NCBI
|
|
16.
|
Iyoda M, Kasamatsu A, Ishigami T,
Nakashima D, Endo-Sakamoto Y, Ogawara K, Shiiba M, Tanzawa H and
Uzawa K: Epithelial cell transforming sequence 2 in human oral
cancer. PLoS One. 5:e140822010. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Onda T, Uzawa K, Nakashima D, Saito K,
Iwadate Y, Seki N, Shibahara T and Tanzawa H: Lin-7C/VELI3/MALS-3:
an essential component in metastasis of human squamous cell
carcinoma. Cancer Res. 67:9643–9648. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Saito Y, Kasamatsu A, Yamamoto A, Shimizu
T, Yokoe H, Sakamoto Y, Ogawara K, Shiiba M, Tanzawa H and Uzawa K:
ALY as a potential contributor to metastasis in human oral squamous
cell carcinoma. J Cancer Res Clin Oncol. 139:585–594. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Koike K, Kasamatsu A, Iyoda M, Saito Y,
Kouzu Y, Koike H, Sakamoto Y, Ogawara K, Tanzawa H and Uzawa K:
High prevalence of epigenetic inactivation of the human four and a
half LIM domains 1 gene in human oral cancer. Int J Oncol.
42:141–150. 2013.PubMed/NCBI
|
|
20.
|
Usukura K, Kasamatsu A, Okamoto A, Kouzu
Y, Higo M, Koike H, Sakamoto Y, Ogawara K, Shiiba M, Tanzawa H and
Uzawa K: Tripeptidyl peptidase II in human oral squamous cell
carcinoma. J Cancer Res Clin Oncol. 139:123–130. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Verburg FA, Wäschle K, Reiners C,
Giovanella L and Lentjes EG: Heterophile antibodies rarely
influence the measurement of thyroglobulin and thyroglobulin
antibodies in differentiated thyroid cancer patients. Horm Metab
Res. 42:736–739. 2010. View Article : Google Scholar
|
|
22.
|
Imam JS, Buddavarapu K, Lee-Chang JS,
Ganapathy S, Camosy C, Chen Y and Rao MK: MicroRNA-185 suppresses
tumor growth and progression by targeting the Six1 oncogene in
human cancers. Oncogene. 29:4971–4979. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Ogoshi K, Kasamatsu A, Iyoda M, Sakuma K,
Yamatoji M, Sakamoto Y, Ogawara K, Shiiba M, Tanzawa H and Uzawa K:
Dickkopf-1 in human oral cancer. Int J Oncol. 39:329–336.
2011.PubMed/NCBI
|
|
24.
|
Schuster-Gossler K, Harris B, Johnson KR,
Serth J and Gossler A: Notch signalling in the paraxial mesoderm is
most sensitive to reduced POFUT1 levels during early mouse
development. BMC Dev Biol. 9:62009. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Stanley P and Guidos CJ: Regulation of
Notch signaling during T- and B-cell development by O-fucose
glycans. Immunol Rev. 230:201–215. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Gurvich N, Perna F, Farina A, Voza F,
Menendez S, Hurwitz J and Nimer SD: L3MBTL1 polycomb protein, a
candidate tumor suppressor in del(20q12) myeloid disorders, is
essential for genome stability. Proc Natl Acad Sci USA.
107:22552–22557. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Mackinnon RN, Selan C, Wall M, Baker E,
Nandurkar H and Campbell LJ: The paradox of 20q11.21 amplification
in a subset of cases of myeloid malignancy with chromosome 20
deletion. Genes Chromosomes Cancer. 49:998–1013. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Kroes RA, Dawson G and Moskal JR: Focused
microarray analysis of glycogene expression in human glioblastomas.
J Neurochem. 103(Suppl 1): 14–24. 2007. View Article : Google Scholar
|
|
29.
|
Loo LW, Tiirikainen M, Cheng I, Lum-Jones
A, Seifried A, Church JM, Gryfe R, Weisenberger DJ, Lindor NM,
Gallinger S, Haile RW, Duggan DJ, Thibodeau SN, Casey G and Le
Marchand L: Integrated analysis of genome-wide copy number
alterations and gene expression in microsatellite stable, CpG
island methylator phenotype-negative colon cancer. Genes
Chromosomes Cancer. 52:450–466. 2013. View Article : Google Scholar : PubMed/NCBI
|