Introduction
Breast cancer is one of the leading causes of
malignancy-related death among females worldwide (1). Most breast cancer-related deaths are
caused by distant metastasis from the primary tumor site. Despite
successful treatment of the primary malignancy, relapse and
subsequent metastatic spread can still occur at other areas of the
body through the bloodstream or lymphatic channels. This leads to
local, regional or distant metastasis in such tissues including
bone, lung, liver, kidney, thyroid and brain (2). Accordingly, the major focus of breast
cancer treatment is to identify chemopreventive drugs for the
treatment of breast cancer metastasis (3).
Invasion and metastasis are fundamental processes
and major causes of morbidity and mortality in breast cancer
patients. Molecular mechanisms of cancer cell invasion and
metastasis involve a complex series of events. One such early event
involves proteolytic degradation of extracellular matrix (ECM)
components (4). The ECM provides
biochemical and mechanical barriers to cell movement of cancer
cells (5). ECM consists of type IV
collagen, laminin, heparan sulfate proteoglycans, nidogen and
fibronectin (6). Therefore, ECM
degradation requires extracellular proteinases, of which the matrix
metalloproteinases (MMPs) have been shown to play a crucial role in
breast cancer.
MMPs, a major group of enzymes that regulate
cellular matrix composition, are zinc- and calcium-dependent
endopeptidases consisting of four subclasses based on substrate,
including collagenases, gelatinases and stromelysins. In
particular, MMP-9 is considered to be an important MMP involved in
cancer invasion and has been found to be directly associated with
invasion, metastasis and poor prognosis of breast cancer (7,8). A
variety of stimuli exist, including growth factors (e.g.
fibro-blast growth factor-2, epidermal growth factor and hepatocyte
growth factor), cytokines (e.g. tumor necrosis factor-α), oncogenes
(e.g. Ras), and 12-O-tetradecanoylphorbol-13-acetate
(TPA) (9–11). Among these stimulators, TPA is a
well-known selective activator of protein kinase C (PKC) (12) and can stimulate MMP-9 synthesis and
secretion during breast cancer cell invasion (13). Additionally, TPA treatments can
induce MMP-9 expression via activation of transcription factors
such as nuclear factor-κ B (NF-κB) and activator protein-1 (AP-1)
(14,15). Consequently, inhibition of MMP-9
expression and/or its upstream regulatory pathways may be an
important approach to treat malignant tumors, including breast
carcinoma. NF-κB and AP-1 are transcription factors important for
regulating MMP-9, as the MMP-9 gene promoter contains binding sites
for both factors (16).
Mitogen-activated protein kinase (MAPK) signaling pathways are
important for AP-1 activation and NF-κB activation and requires
I-κB kinase, ERK, JNK or p38 MAPK, depending on the cell type
(11,17).
Decursin is a coumarin compound found in the roots
of Angelica gigas Nakai, which has been traditionally used
in Korean folk medicine as a tonic and for the treatment of anemia
and other diseases (18). Decursin
induces cell cycle arrest and apoptosis in human prostate cancer
cells, human breast cancer cells, human bladder cancer cells and
colon cancer cells (19–21). Recent studies demonstrated that
decursin blocks MMP-9 expression through inhibition of NF-κB
activation in macrophages and cancer cells (21–23).
However, the mechanism by which decursin mediates anti-invasiveness
is not well understood. Therefore, it has been hypothesized that
decursin may have anticancer properties through the inhibition of
cell invasion. In this study, decursin was examined for its
potential effects on TPA-induced cell invasion and MMP-9 expression
in MCF-7 cells. Furthermore, the related molecular mechanisms were
also investigated.
Materials and methods
Cells and materials
MCF-7 cells were obtained from the American Type
Culture Collection (Manassas, VA, USA). Cells were cultured in DMEM
supplemented with 10% fetal bovine serum (FBS) and 1% antibiotics
at 37°C in a 5% CO2 incubator. Decursin, TPA,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
and anti-β-actin antibody were obtained from Sigma-Aldrich (St.
Louis, MO, USA). Antibodies against p38, phosphorylated p38
(p-p38), c-Jun N-terminal kinase (JNK), p-JNK, extracellular
signal-regulated kinase (ERK), p-ERK, p-c-Jun, p-IκBα, and p-IKKα
were purchased from Cell Signaling Technology (Beverly, MA, USA).
Antibodies against MMP-9, p50, p65, IκBα, IKKα, IKKβ, PKCα, PKCδ,
proliferating cell nuclear antigen (PCNA), and horseradish
peroxidase (HRP)-conjugated IgG were purchased from Santa Cruz
Biotechnology (Santa Cruz, CA, USA). [α-32P]dCTP was
obtained from Amersham (Buckinghamshire, UK). DMEM containing a
high concentration of glucose, FBS, and phosphate-buffered saline
(PBS) were obtained from Gibco-BRL (Gaithersburg, ME, USA).
Determination of cell viability
The effect of decursin on MCF-7 cell viability was
determined using an established MTT assay. In brief,
3×l04 cells were seeded in wells and incubated at 37°C
for 24 h to allow attachment. The attached cells were untreated or
treated with 1, 5, 10, 25 or 50 μM decursin for 24 h at
37°C. The cells were washed with PBS prior to adding MTT (0.5 mg/ml
PBS) and incubated at 37°C for 30 min. Formazan crystals were
dissolved with dimethyl sulfoxide (100 μl/well) and detected
at 570 nm using a model 3550 microplate reader (Bio-Rad, Richmond,
CA, USA).
Western blot analysis
MCF-7 cells (5×105) were pre-treated with
25 μM decursin for 1 h and then incubated with TPA for 24 h
at 37°C. Cells were lysed with ice-cold M-PER® Mammalian
Protein Extraction Reagent (Pierce Biotechnology, Rockford, IL,
USA), and protein concentration was determined using the Bradford
method. Samples (20 μg) were separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis with 10% acrylamide and
transferred to Hybond™ polyvinylidene fluoride membranes (GE
Healthcare Life Sciences, Buckinghamshire, UK) using a western blot
apparatus. Each membrane was blocked for 2 h with 2% bovine serum
albumin or 5% skim milk and then incubated overnight at 4°C with 1
μg/ml of a 1:2000 dilution of primary antibody.
HRP-conjugated IgG (1:2000 dilution) was used as the secondary
antibody. Protein expression levels were determined by signal
analysis using an image analyzer (Fuji-Film, Tokyo, Japan).
Gelatin zymography assay
Conditioned media was collected after 24 h
stimulation, mixed with non-reducing sample buffer, and
electrophoresed in a polyacrylamide gel containing 0.1% (w/v)
gelatin. The gel was washed at room temperature for 30 min with
2.5% Triton X-100 solution, and subsequently incubated at 37°C for
16 h in 5 mM CaCl2, 0.02% Brij and 50 mM Tris-HCl (pH
7.5). The gel was stained for 30 min with 0.25% (w/v) Coomassie
brilliant blue in 40% (v/v) methanol/7% (v/v) acetic acid and
photographed on an image analyzer (Fuji-Film). Proteolysis was
visualized as a white zone in a dark blue field. Densitometric
analysis was performed using Multi Gauge Image Analysis software
(Fuji-Film).
Quantitative real-time polymerase chain
reaction
Total RNA was extracted from cells using a FastPure™
RNA kit (Takara, Shiga, Japan). RNA concentration and purity were
determined by absorbance at 260/280 nm. cDNA was synthesized from 1
μg total RNA using a PrimeScript™ RT reagent kit (Takara).
MMP-9 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA
expression were determined by real-time PCR using the ABI PRISM
7900 sequence detection system and SYBR® Green (Applied
Biosystems, Foster City, CA, USA). The primers were: MMP-9 (NM
004994) sense, CCTGGAGACCTGAGAACCAATCT, antisense, CCACCC
GAGTGTAACCATAGC; and GAPDH (NM 002046) sense,
ATGGAAATCCCATCACCATCTT, antisense, CGCCC CACTTGATTTTGG. To control
for variation in mRNA concentration, all results were normalized to
the GAPDH housekeeping gene. Relative quantitation was performed
using the comparative ΔΔCt method according to the
manufacturer’s instructions.
Preparation of nuclear extract
MCF-7 cells (2×106) were treated with
decursin in the presence or absence of TPA for 4 h. Cells were
immediately washed twice, scraped into 1.5 ml of ice-cold PBS (pH
7.5), and pelleted at 1,500 × g for 3 min. Cytoplasmic and nuclear
extracts were prepared from cells using the NE-PER®
Nuclear and Cytoplasmic Extraction Reagents (Pierce
Biotechnology).
Membrane fractionation
MCF-7 cells (5×107) were pre-treated with
25 μM decursin for 1 h and then incubated with TPA for 30
min at 37°C. Cells were immediately washed twice, scraped into 1.5
ml of ice-cold PBS (pH 7.5), and pelleted at 4,000 rpm for 3 min.
Cell lysis was carried out in homogenization buffer (20 mM
Tris-HCl, 5 mM DTT, 2 mM EDTA, 5 mM EGTA, protease inhibitor,
phosphatase inhibitors, pH 7.5) with brief sonication (5 times, 10
sec and 10% amplitude) after incubate on ice for 15–30 min. Cell
debris was removed by centrifuging the sample at 3,000 rpm for 10
min at 4°C. Cell lysate was centrifuged at 13,000 rpm for 30 min at
4°C to separate out soluble (cytosolic) and pellet (membrane)
fraction. Pellet fraction was incubated in homogenization buffer
containing 1% Triton X-100 for 30 min in ice, centrifuged at 50,000
rpm for 1 h and the supernatant was collected as the membrane
fraction.
Electrophoretic mobility shift assay
(EMSA)
Activation of NF-κB and AP-1 was assessed with a gel
mobility shift assay using nuclear extracts. An oligonucleotide
containing the κ-chain (κB, 5′-CCGGTTAACAGAGGGGGCTTTCCGAG-3′) or
AP-1 (5′-CGCTTGATGAGTCAGCCGGAA-3′) binding site was synthesized and
used as a probe for the gel retardation assay. The two
complementary strands were annealed and labeled with
[α-32P]dCTP. Labeled oligonucleotides (10,000 cpm), 10
μg of nuclear extracts, and binding buffer (10 mM Tris-HCl,
pH 7.6, 500 mM KCl, 10 mM EDTA, 50% glycerol, 100 ng poly (dI·dC),
1 mM dithiothreitol) were then incubated for 30 min at room
temperature in a final volume of 20 μl. The reaction
mixtures were analyzed by electrophoresis on 4% polyacrylamide gels
in 0.5X Tris-borate buffer. The gels were dried and examined by
autoradiography. Specific binding was controlled by competition
with a 50-fold excess of cold κB oligonucleotide.
Invasion assay
The invasion assay was carried out in 24-well
chambers (8-μm pore size) coated with 20 μl matrigel
diluted DMEM. The matrigel coating was re-hydrated in 0.5 ml DMEM
for 30 min immediately before the experiments. Cells
(2×105) were added to the upper chamber with
chemoattractant in the bottom well. Conditioned medium (0.5 ml) was
added to the lower compartment of the invasion chamber. The
chambers were incubated for 24 h. Following incubation, cells on
the upper side of the chamber were removed using cotton swabs, and
cells that had migrated were fixed and stained with Toluidine blue
solution. Invading cells were counted in five random areas of the
membrane using a light microscope. Analyzed data are the means ± SE
from three individual experiments performed in triplicate.
Statistical analysis
Statistical data analysis was performed using ANOVA
and Duncan’s test. Differences with p<0.05 were considered
statistically significant.
Results
Decursin does not affect MCF-7 cell
viability
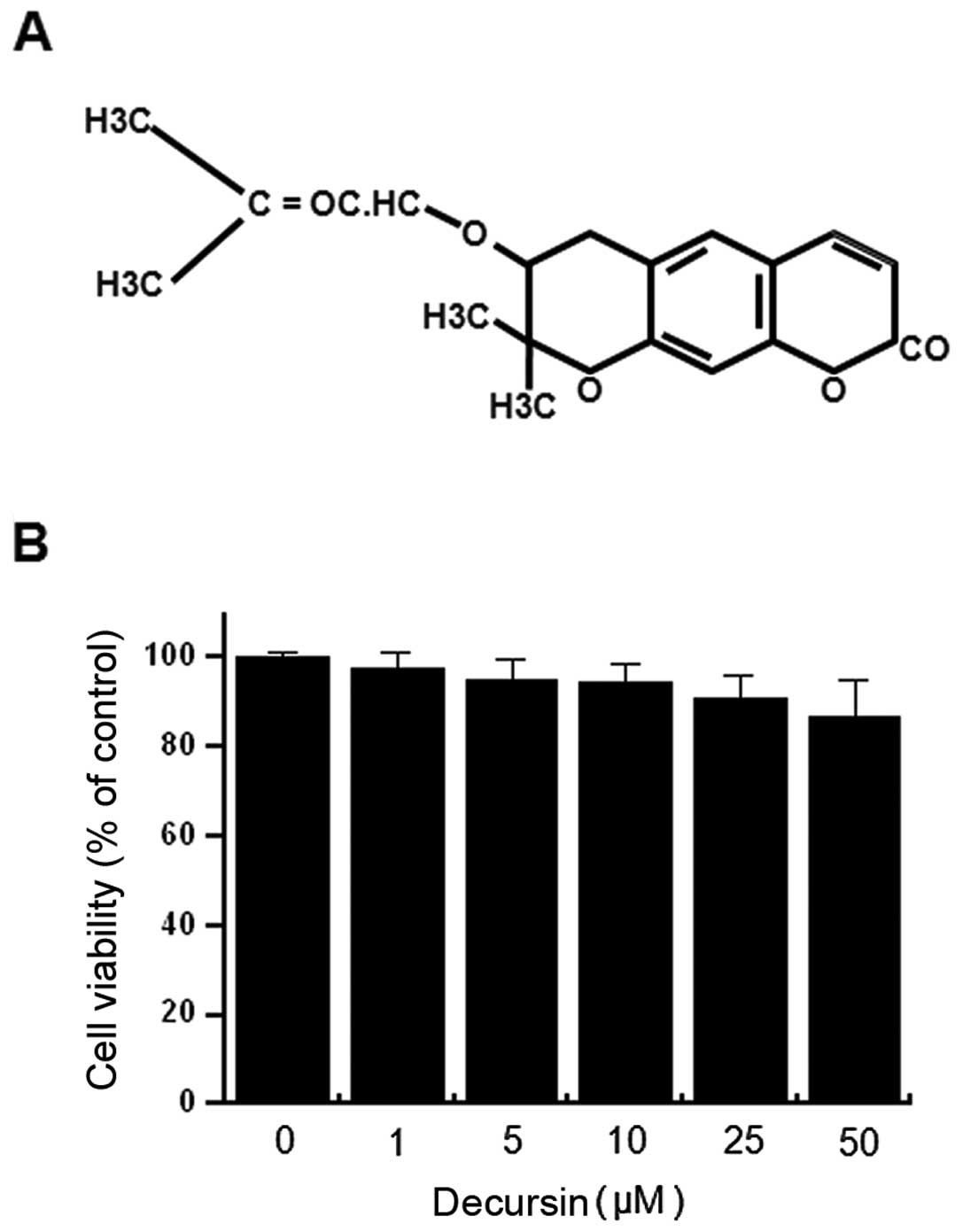
Fig. 1A is the
chemical structure of decursin. In order to investigate the
cytotoxicity of decursin on MCF-7 cells, cells were seeded into the
wells of 96-well culture plates at a density of 3×104
cells/well. The effect of decursin on MCF-7 cellular toxicity was
analyzed using the MTT assay. Treatment of MCF-7 cells with all
concentrations of decursin for 24 h did not lead to a significant
change in cell viability (Fig. 1B)
or morphology. Therefore, subsequent experiments utilized the
optimal nontoxic concentration (25 μM) of decursin.
TPA-induced MMP-9 activation was
decreased by decursin in MCF-7 cells
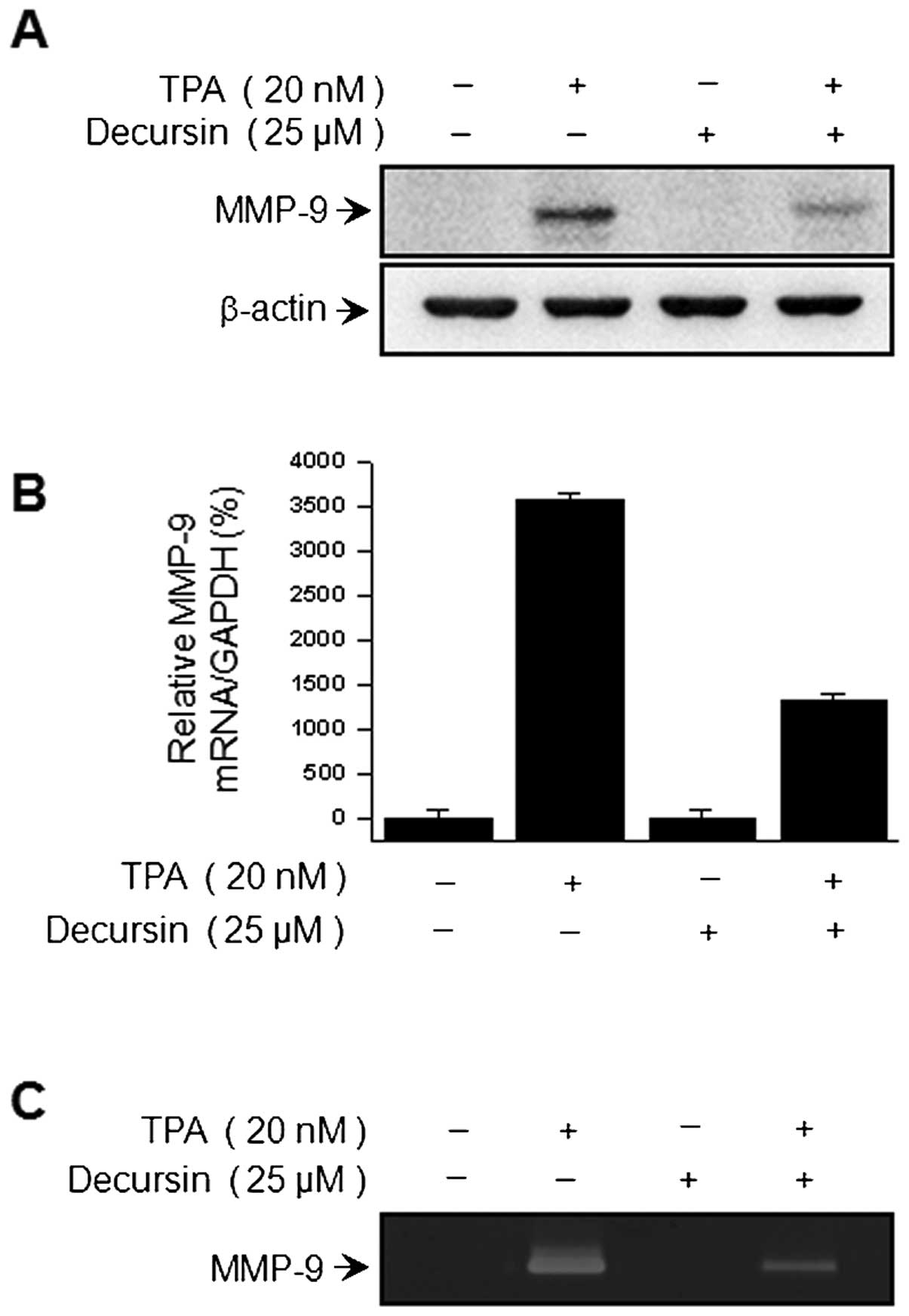
To determine the effect of decursin on TPA-induced
MMP-9 expression, we pretreated cells with the indicated
concentration of decursin prior to 20 nM TPA treatment in MCF-7
cells. After 24 h, western blot analysis, real-time PCR and
zymography were performed in MCF-7 cell-containing samples. From
our results, western blot analysis showed that the decursin
treatment of MCF-7 cells blocked the upregulation of TPA-induced
MMP-9 protein expression (Fig.
2A). Real-time PCR revealed a TPA-induced increase in the
expression of MMP-9 in MCF-7 cells, and that the addition of
decursin blocked this TPA-induced MMP-9 upregulation in a
dose-dependent manner (Fig. 2B).
In addition, we examined the level of TPA-induced MMP-9 secretion
by decursin using zymography analysis. We demonstrated that TPA
treatment of MCF-7 cells increased MMP-9 secretion, while the
addition of decursin significantly diminished TPA-induced MMP-9
secretion (Fig. 2C).
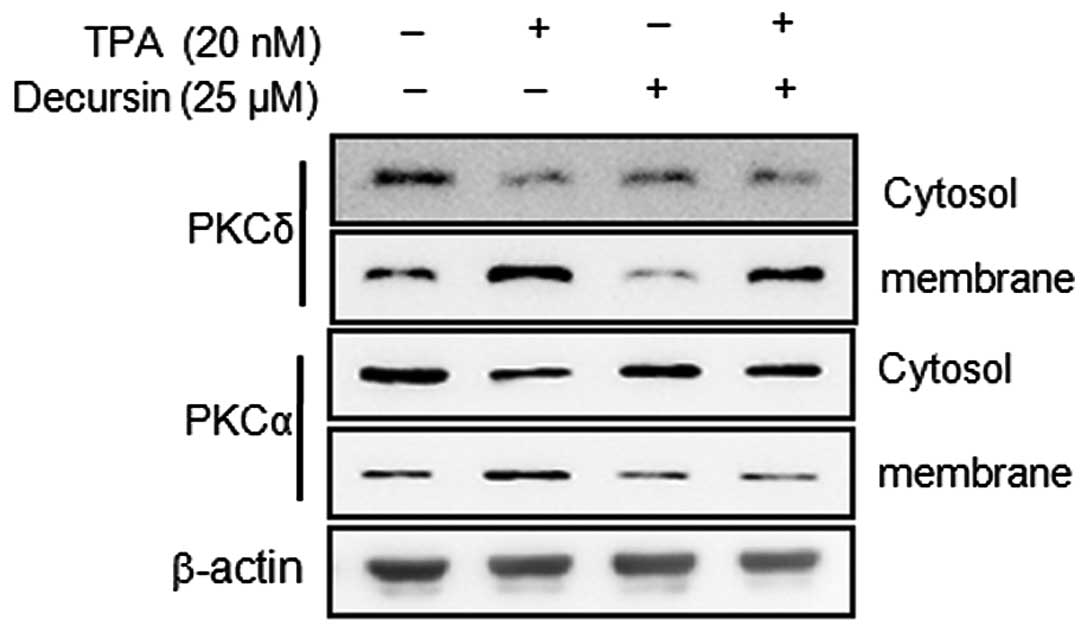
Decursin inhibits membrane localization
of TPA-induced PKCα
Activation of PKCs has been shown to correlate with
potential tumor metastasis (13,24).
TPA has been reported as a PKC activator, and activation of the PKC
isoforms, including α, β, δ and ɛ, by TPA has been identified
(25). To determine whether
decursin activates any PKC isotypes in MCF-7 cells, we analyzed the
levels of PKCα and PKCδ in the cytosol and membrane and observed
stimulation by TPA, while decursin pretreatment attenuated the
translocation of these PKCs. As shown in Fig. 3, TPA-induced membrane localization
of PKCα was blocked by pretreatment with decursin for 1 h. These
results suggest that TPA-induced MMP-9 expression and invasion may
be involved in the activation of PKCα in MCF-7 cells, while the
addition of decursin inhibits TPA-induced PKCα activation.
Decursin inhibits TPA-induced NF-κB DNA
binding activity and the MAPK pathway, but not AP-1 DNA binding
activity
To elucidate the mechanism governing
decursin-mediated inhibition of MMP-9 expression, the effect of
decursin on TPA-induced activation of NF-κB and AP-1 was confirmed
using EMSA. We investigated the NF-κB and AP-1 signaling pathways
in nuclear extracts prepared 4 h after TPA stimulation. As shown in
Fig. 4A and B, TPA substantially
increased NF-κB and AP-1 binding activity. Of note, pre-treatment
with decursin inhibited TPA-stimulated NF-κB binding activity, but
not the AP-1 pathway. These results were consistent with the view
that decursin specifically blocks NF-κB activation in MCF-7 cells.
In the western blot analysis, TPA stimulated the phosphorylation of
I-κBα and I-κBα in the cytoplasm and, accordingly, the nuclear
translocation of NF-κB subunits p50 and p65. In the case of AP-1,
c-Jun expression was considerably augmented, but c-Fos expression
was only negligibly induced in TPA-treated cells. Therefore, from
our results we observed that an increase in p-I-κBα and
translocation of p65 and p50 as a result of TPA stimulation was
significantly suppressed by decursin (Fig. 4D and E). Furthermore, we confirmed
that TPA-induced phosphorylation of c-Jun, a major subunit of AP-1,
and decursin have no effect on the phosphorylation of c-Jun
(Fig. 4E). To investigate the
inhibitory effect of decursin on the MAPK pathway, which is the
so-called upstream modulator of NF-κB and AP-1, TPA-induced
phosphorylation and activation of MAPKs was first confirmed. We
observed that decursin inhibited the phosphorylation of p38, but
not ERK and JNK 30 min after TPA treatment (Fig. 4C). These results suggest that
decursin inhibits TPA-induced MMP-9 expression through the
regulation of the p38/MAPK-NF-κB pathway in MCF-7 cells.
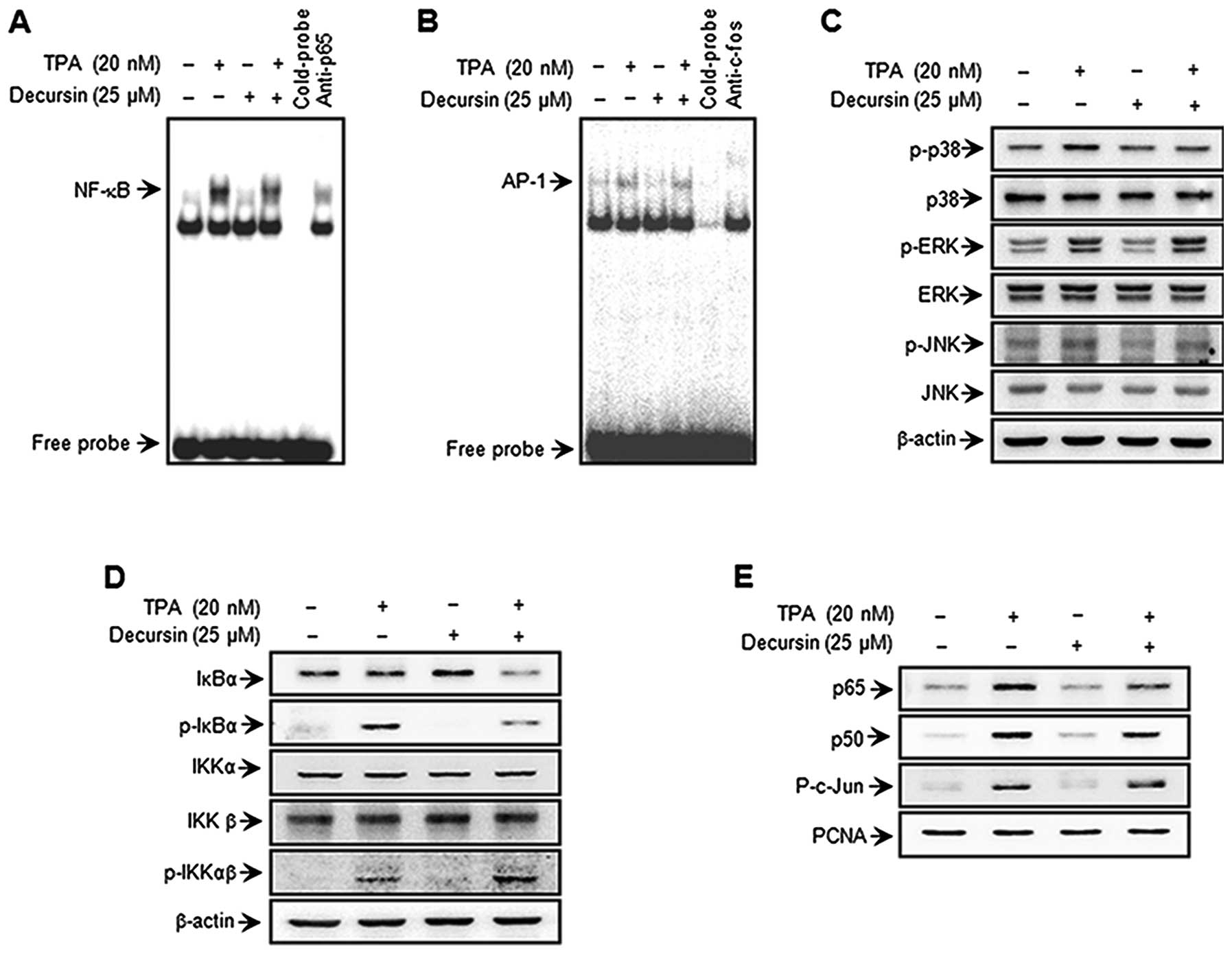 | Figure 4.Decursin inhibits TPA-induced
transcriptional activation of MMP-9 and phosphorylation of p38 MAPK
in MCF-7 cells. Cells were treated with decursin in the presence of
TPA. Following incubation for 4 h, nuclear extracts were prepared.
DNA binding of NF-κB and AP-1 was analyzed by EMSA (A and B).
Western blotting was performed to determine the cytoplasmic levels
of p-IKKαβ, IKKα, IKKβ, p-IκBα, and IκBα, as well as the nuclear
levels of the subunits of NF-κB (p50 and p65) and AP-1 (p-c-Jun) (D
and E). Cells were pretreated with TPA for 30 min in the presence
or absence of decursin. Cell lysates were prepared for western blot
analysis with specific p-p38, p38, p-JNK, JNK, p-ERK and ERK
antibodies (C). |
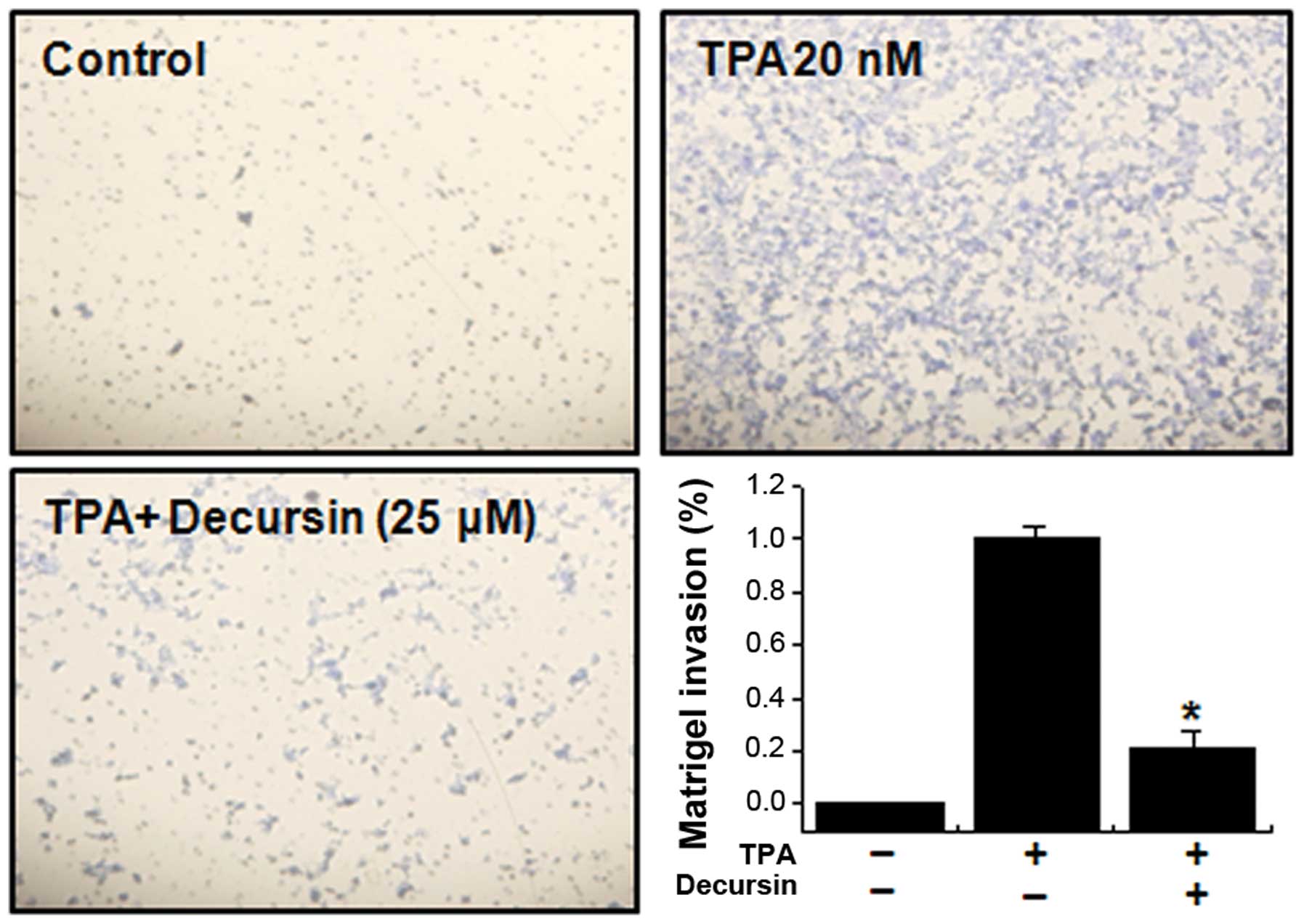
Decursin decreases TPA-induced MCF-7 cell
invasion in vitro
It has been reported that the upregulation of MMP-9
expression contributes to the invasion of cancer cells (4). Therefore, an in vitro invasion
assay was used to investigate the inhibitory effects of decursin on
the invasive potency of MCF-7 breast adenocarcinoma cells.
Treatment with TPA increased MCF-7 cell invasion compared with
untreated control cells, as determined by a Matrigel invasion
assay. Incubation of MCF-7 cells with TPA resulted in a 10-fold
increase in cell invasiveness. However, we observed that treatment
with decursin diminished TPA-induced cell invasion by 79% (Fig. 5).
Discussion
Current studies involved in developing effective
anti-invasion strategies have focused mainly on the use of natural
bio active agents in MCF-7 cells. In this study, we isolated
decursin from the roots of Angelica gigas Nakai and examined
its effects on TPA-induced MMP-9 expression and invasion in MCF-7
cells. In recent studies, decursin was found to prevent MMP-9
expression by suppression of the NF-κB pathway in cancer cells and
macrophages (22,26). Additionally, our previous study
showed that decursin inhibits UVB-induced MMP expression in human
dermal fibroblasts via regulation of NF-κB (27). However, to date, no report exists
on the anti-invasion effects of decursin in MCF-7 cells. In this
study, we examined the anti-invasive potential of decursin and
explored the molecular mechanisms underlying its activity.
Breast cancer is the most commonly detected cancer
and one of the leading causes of death in women worldwide.
Metastasis is the primary cause of breast cancer mortality. Tumor
metastasis is a complex, multistep process that includes cell
proliferation, ECM degradation, cell migration and tumor growth at
metastatic sites (28).
Morphologically, tumor invasion is associated with the presence of
a distorted edge of the primary tumor, where individual or cohorts
of tumor cells actively invade the tissue ECM surrounding the
primary tumor (29).
MMP-9 is a critical molecule in tumor invasion and
metastasis. MMP-9 activation has been shown to be associated with
tumor progression and invasion, including mammary tumors (30). In previous studies, inflammatory
cytokines, growth factors and TPA were shown to stimulate MMP-9 by
activating different intracellular-signaling pathways in breast
cancer cells (31). TPA increases
the invasiveness of MCF-7 cells by increasing transcription of
MMP-9 and activating the PKC, MAPK and phosphoinositide 3 kinase
(PI3K)/Akt pathways. Activation of PKC by TPA involves the
translocation of PKC isoforms to the plasma membrane, resulting in
proliferation, differentiation, malignant transformation, and tumor
promotion and progression in cancer cells. Our previous studies
demonstrated that PKCα and PKCδ play critical roles in MMP-9
induction and invasion in MCF-7 cells. In our previous data, TPA
stimulation resulted in the translocation of PKCα and PKCδ from the
cytosol to the cell membrane, although translocation of PKCβ was
not observed. Treatment with a non-cytotoxic dose of a PKCδ
inhibitor (rottlerin), a broad PKC inhibitor (GF109203X), and a
PKCα inhibitor (Gö6976) caused marked inhibition in TPA-induced
MMP-9 expression and secretion (32). In this study, we observed that
decursin inhibited TPA-induced membrane localization of PKCα, but
not of PKCδ.
Additionally, to understand the TPA-induced
signaling cascade underlying MMP-9 expression in MCF-7 cells, we
assessed the effects of three MAPKs and the DNA binding abilities
of transcription factors. The three major MAPK families, JNK, ERK
and p38 kinase, are expressed in MCF-7 cells, and their active
phosphorylated forms can be detected (28). MAPK signaling pathways are
important for NF-κB and AP-1 activation, which require IκB kinase,
PI3K-Akt or p38 MAPK, depending on the cell type (11,17,33).
Our results show that decursin displayed inhibitory effects on the
phosphorylation of p38 MAPK. In addition, it has been established
that TPA induces the nuclear transcription factors NF-κB and AP-1
in MCF-7 cells. NF-κB and AP-1 are transcription factors that are
important in regulating MMP-9, as the MMP-9 gene promoter contains
NF-κB and AP-1 binding sites (16). The NF-κB and AP-1 elements are
centrally involved in TPA-mediated MMP-9 gene induction (14,34).
The present results show that decursin inhibits activation of NF-κB
but not AP-1 in MCF-7 cells. Previous results confirm that
TPA-induced MMP-9 expression can be significantly inhibited by
selective inhibitors of p38 (SP600125) (35). The present results show that
decursin significantly inhibits TPA-activated p38 MAPK. Together,
these results confirm that TPA-stimulated cell invasion can be
suppressed by the inhibition of the MAPK/NF-κB pathways.
Finally, these experiments confirmed that
TPA-stimulated cell invasion was suppressed by decursin. The data
obtained from our Matrigel invasion assay showed that decursin
inhibits the TPA-induced invasion potential of MCF-7 cells
(Fig. 5). In conclusion, decursin
inhibited TPA-induced invasion by reducing MMP-9 activation mainly
through the PKCα, MAPK, and NF-κB pathways in MCF-7 cells. This is
the first study demonstrating that decursin can suppress
TPA-stimulated cancer cell invasion by inhibiting MMP-9 expression
through the suppression of PKCα. Furthermore, our results present,
for the first time, details of the molecular mechanisms in MCF-7
cells responsible for this inhibitory effect.
Acknowledgements
This work was supported by the
National Research Foundation of Korea (NRF) grant funded by the
Korea government (MEST) (no. 2011-0030130), and by Fund of Chonbuk
National University Hospital Research Institute of Clinical
Medicine.
References
|
1.
|
Jemal A, Murray T, Ward E, Samuels A,
Tiwari RC, Ghafoor A, Feuer EJ and Thun MJ: Cancer statistics,
2005. CA Cancer J Clin. 55:10–30. 2005. View Article : Google Scholar
|
|
2.
|
Friedel G, Pastorino U, Ginsberg RJ,
Goldstraw P, Johnston M, Pass H, Putnam JB and Toomes H: Results of
lung metastasectomy from breast cancer: prognostic criteria on the
basis of 467 cases of the International Registry of Lung
Metastases. Eur J Cardiothorac Surg. 22:335–344. 2002. View Article : Google Scholar
|
|
3.
|
Zwiefel K and Janni W: Current standards
in the treatment of breast cancer. Med Monatsschr Pharm.
34:280–290. 2011.(In German).
|
|
4.
|
Chambers AF and Matrisian LM: Changing
views of the role of matrix metalloproteinases in metastasis. J
Natl Cancer Inst. 89:1260–1270. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Woessner JF Jr: Matrix metalloproteinases
and their inhibitors in connective tissue remodeling. FASEB J.
5:2145–2154. 1991.PubMed/NCBI
|
|
6.
|
Nakajima M, Welch DR, Belloni PN and
Nicolson GL: Degradation of basement membrane type IV collagen and
lung subendothelial matrix by rat mammary adenocarcinoma cell
clones of differing metastatic potentials. Cancer Res.
47:4869–4876. 1987.
|
|
7.
|
Brinckerhoff CE and Matrisian LM: Matrix
metalloproteinases: a tail of a frog that became a prince. Nat Rev
Mol Cell Biol. 3:207–214. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Velinov N, Poptodorov G, Gabrovski N and
Gabrovski S: The role of matrixmetalloproteinases in the tumor
growth and metastasis. Khirurgiia. 1:44–49. 2010.(In
Bulgarian).
|
|
9.
|
Zeigler ME, Chi Y, Schmidt T and Varani J:
Role of ERK and JNK pathways in regulating cell motility and matrix
metalloproteinase 9 production in growth factor-stimulated human
epidermal keratinocytes. J Cell Physiol. 180:271–284. 1999.
View Article : Google Scholar
|
|
10.
|
Hozumi A, Nishimura Y, Nishiuma T, Kotani
Y and Yokoyama M: Induction of MMP-9 in normal human bronchial
epithelial cells by TNF-alpha via NF-kappa B-mediated pathway. Am J
Physiol Lung Cell Mol Physiol. 281:L1444–L1452. 2001.PubMed/NCBI
|
|
11.
|
Weng CJ, Chau CF, Hsieh YS, Yang SF and
Yen GC: Lucidenic acid inhibits PMA-induced invasion of human
hepatoma cells through inactivating MAPK/ERK signal transduction
pathway and reducing binding activities of NF-kappaB and AP-1.
Carcinogenesis. 29:147–156. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Newton AC: Regulation of protein kinase C.
Curr Opin Cell Biol. 9:161–167. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Lin CW, Hou WC, Shen SC, Juan SH, Ko CH,
Wang LM and Chen YC: Quercetin inhibition of tumor invasion via
suppressing PKCdelta/ERK/AP-1-dependent matrix metalloproteinase-9
activation in breast carcinoma cells. Carcinogenesis. 29:1807–1815.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Hong S, Park KK, Magae J, Ando K, Lee TS,
Kwon TK, Kwak JY, Kim CH and Chang YC: Ascochlorin inhibits matrix
metalloproteinase-9 expression by suppressing activator
protein-1-mediated gene expression through the ERK1/2 signaling
pathway: inhibitory effects of ascochlorin on the invasion of renal
carcinoma cells. J Biol Chem. 280:25202–25209. 2005. View Article : Google Scholar
|
|
15.
|
Woo MS, Jung SH, Kim SY, Hyun JW, Ko KH,
Kim WK and Kim HS: Curcumin suppresses phorbol ester-induced matrix
metalloproteinase-9 expression by inhibiting the PKC to MAPK
signaling pathways in human astroglioma cells. Biochem Biophys Res
Commun. 335:1017–1025. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Eberhardt W, Huwiler A, Beck KF, Walpen S
and Pfeilschifter J: Amplification of IL-1 beta-induced matrix
metalloproteinase-9 expression by superoxide in rat glomerular
mesangial cells is mediated by increased activities of NF-kappa B
and activating protein-1 and involves activation of the
mitogen-activated protein kinase pathways. J Immunol.
165:5788–5797. 2000.
|
|
17.
|
Madrid LV, Mayo MW, Reuther JY and Baldwin
AS Jr: Akt stimulates the transactivation potential of the RelA/p65
subunit of NF-kappaB through utilization of the Ikappa B kinase and
activation of the mitogen-activated protein kinase p38. J Biol
Chem. 276:18934–18940. 2001. View Article : Google Scholar
|
|
18.
|
Ahn Q, Jeong SJ, Lee HJ, Kwon HY, Han I,
Kim HS, Lee EO, Ahn KS, Jung MH, Zhu S, Chen CY and Kim SH:
Inhibition of cyclooxygenase-2-dependent survivin mediates
decursin-induced apoptosis in human KBM-5 myeloid leukemia cells.
Cancer Lett. 298:212–221. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Yim D, Singh RP, Agarwal C, Lee S, Chi H
and Agarwal R: A novel anticancer agent, decursin, induces G1
arrest and apoptosis in human prostate carcinoma cells. Cancer Res.
65:1035–1044. 2005.PubMed/NCBI
|
|
20.
|
Jiang C, Guo J, Wang Z, Xiao B, Lee HJ,
Lee EO, Kim SH and Lu J: Decursin and decursinol angelate inhibit
estrogen-stimulated and estrogen-independent growth and survival of
breast cancer cells. Breast Cancer Res. 9:R772007. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Kim WJ, Lee SJ, Choi YD and Moon SK:
Decursin inhibits growth of human bladder and colon cancer cells
via apoptosis, G1-phase cell cycle arrest and extracellular
signal-regulated kinase activation. Int J Mol Med. 25:635–641.
2010.
|
|
22.
|
Kim JH, Jeong JH, Jeon ST, Kim H, Ock J,
Suk K, Kim SI, Song KS and Lee WH: Decursin inhibits induction of
inflammatory mediators by blocking nuclear factor-kappaB activation
in macrophages. Mol Pharmacol. 69:1783–1790. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Lee SH, Lee JH, Kim EJ, Kim WJ, Suk K, Kim
JH, Song GY and Lee WH: A novel derivative of decursin, CSL-32,
blocks migration and production of inflammatory mediators and
modulates PI3K and NF-kappaB activities in HT1080 cells. Cell Biol
Int. 36:683–688. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Zhang J, Anastasiadis PZ, Liu Y, Thompson
EA and Fields AP: Protein kinase C (PKC) betaII induces cell
invasion through a Ras/Mek-, PKC iota/Rac 1-dependent signaling
pathway. J Biol Chem. 279:22118–22123. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Fortino V, Torricelli C, Capurro E, Sacchi
G, Valacchi G and Maioli E: Antiproliferative and survival
properties of PMA in MCF-7 breast cancer cell. Cancer Invest.
26:13–21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Kim WJ, Lee MY, Kim JH, Suk K and Lee WH:
Decursinol angelate blocks transmigration and inflammatory
activation of cancer cells through inhibition of PI3K, ERK and
NF-kappaB activation. Cancer Lett. 296:35–42. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Hwang BM, Noh EM, Kim JS, Kim JM, Hwang
JK, Kim HK, Kang JS, Kim DS, Chae HJ, You YO, Kwon KB and Lee YR:
Decursin inhibits UVB-induced MMP expression in human dermal
fibroblasts via regulation of nuclear factor-κB. Int J Mol Med.
31:477–483. 2013.PubMed/NCBI
|
|
28.
|
Lee SO, Jeong YJ, Kim M, Kim CH and Lee
IS: Suppression of PMA-induced tumor cell invasion by capillarisin
via the inhibition of NF-kappaB-dependent MMP-9 expression. Biochem
Biophys Res Commun. 366:1019–1024. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Deryugina EI and Quigley JP: Matrix
metalloproteinases and tumor metastasis. Cancer Metastasis Rev.
25:9–34. 2006. View Article : Google Scholar
|
|
30.
|
Scorilas A, Karameris A, Arnogiannaki N,
Ardavanis A, Bassilopoulos P, Trangas T and Talieri M:
Overexpression of matrix-metalloproteinase-9 in human breast
cancer: a potential favourable indicator in node-negative patients.
Br J Cancer. 84:1488–1496. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Yan C and Boyd DD: Regulation of matrix
metalloproteinase gene expression. J Cell Physiol. 211:19–26. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Kim JM, Noh EM, Kwon KB, Kim JS, You YO,
Hwang JK, Hwang BM, Kim BS, Lee SH, Lee SJ, Jung SH, Youn HJ and
Lee YR: Curcumin suppresses the TPA-induced invasion through
inhibition of PKCalpha-dependent MMP-expression in MCF-7 human
breast cancer cells. Phytomedicine. 19:1085–1092. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Yao J, Xiong S, Klos K, Nguyen N, Grijalva
R, Li P and Yu D: Multiple signaling pathways involved in
activation of matrix metalloproteinase-9 (MMP-9) by heregulin-beta1
in human breast cancer cells. Oncogene. 20:8066–8074. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Chung TW, Moon SK, Chang YC, Ko JH, Lee
YC, Cho G, Kim SH, Kim JG and Kim CH: Novel and therapeutic effect
of caffeic acid and caffeic acid phenyl ester on hepatocarcinoma
cells: complete regression of hepatoma growth and metastasis by
dual mechanism. FASEB J. 18:1670–1681. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Lee YR, Noh EM, Oh HJ, Hur H, Kim JM, Han
JH, Hwang JK, Park BH, Park JW, Youn HJ, Jung SH, Kim BS, Jung JY,
Lee SH, Park CS and Kim JS: Dihydroavenanthramide D inhibits human
breast cancer cell invasion through suppression of MMP-9
expression. Biochem Biophys Res Commun. 405:552–557. 2011.
View Article : Google Scholar : PubMed/NCBI
|