Introduction
Nasopharyngeal carcinoma (NPC) is one of the most
common cancers of the head and neck, particularly in southern China
and Southeast Asia (1,2). Though NPC patients are sensitive to
radio/chemo-therapy, treatment failure remains high due to the
development of local recurrence and distant metastasis (1). Epstein-Barr virus (EBV) infection,
environmental factors and genetic susceptibility are associated
with this malignancy (2). To date,
the molecular mechanisms related to the progression and clinical
outcome of NPC have not yet been fully understood. A combination of
radiotherapy and adjuvant chemotherapy has been successful for
certain NPC patients, but the 5-year survival rate is only 50–60%.
There is almost no effective treatment for those who are resistant
to radiotherapy and have tumor recurrence. Therefore, we need to
further explore the pro-oncogenic pathways that lead to or promote
NPC with the intention of identifying new targets for therapy and
improving patient survival. In addition, understanding the
molecular mechanisms and key regulatory factors exploitable in
developing adjuvant therapies to augment currently available
treatment protocols that allow decreased side-effects and toxicity
without compromising therapeutic efficacy are also required.
Recently, much attention has been focused on the use of traditional
Chinese medicinal herbs in cancer prevention and therapy due to the
pleiotropic effects of these agents on multiple
carcinogen-activated oncogenic pathways, and their equally
excellent safety profiles. Curcumin is one such potential
candidate.
Curcumin, one of the main bioactive components
extracted from a traditional Chinese medicinal herb, is associated
with a variety of functions such as immunomodulatory,
anti-inflammatory and anticancer activities (3). Substantial evidence has demonstrated
that curcumin acts as a potential chemopreventive agent as well as
a novel adjuvant treatment agent for head and neck malignancies via
its effect on a variety of biological pathways (4). However, the detailed mechanisms by
which this agent is involved in the inhibition of NPC cell growth
and induction of apoptosis have not been well elucidated.
The p53 tumor suppressor is a major regulator of
cell proliferation and apoptosis, which is activated in response to
DNA damage and is mutated in ∼50% of human cancers (5). As a tumor suppressor protein, p53
plays a pivotal role in regulating the cellular response to stress
and damage signals, and loss of p53 functionality is common in more
than 50% of cancers (6). The role
of p53 in the link of NPC has been reported, activation of p53 was
involved in the radioresponse in NPC (7) and played an important role in the
development of novel therapies for NPC treatment (8). The forkhead box, class O belongs to
the family of mammalian forkhead transcription factors, including
FOXO3a (or FKHRL1), FOXO1a (or FKHR), and FOXO4a (or AFX), which
are regulated by growth factor receptor-induced activation of the
phosphatidylinositol 3-kinase (PI3K)/AKT (or protein kinase B)
signaling pathway (9). Studies in
mammalian cells have shown that activation of FOXO3a by stimulated
the expression of proteins that are involved in apoptosis (9) and cell cycle arrest (10) in different types of cells. FoxO3a
is a transcription factor with known tumor suppressor activity and
a conserved 110-amino acid DNA-binding domain and recognize two
consensus DNA-binding sequences: 5′-TTGTTTAC-3′ and
5′-(C/A)(A/C)AAA(C/T)AA-3′ (11).
Inhibition of FOXO3a expression promotes cell transformation, tumor
progression and angiogenesis (12). Expression of FOXO3a was reported in
NPC and considered as important prognostic marker in NPC (13).
In the present study, we investigated the potential
mechanism by which curcumin inhibited NPC cell growth and induced
apoptosis. Our results showed that extracellular signal-regulated
kinase 1/2 (ERK1/2) signaling pathway-mediated increase in the
protein expression and the interaction of FOXO3a and p53
contributed to the effect of curcumin on proliferation and
apoptosis of NPC cells.
Materials and methods
Reagents
Curcumin was purchased from Sigma-Aldrich (Shanghai,
China). RPMI-1640 and penicillin/streptomycin were purchased from
Invitrogen (Carlsbad, CA, USA), fetal bovine serum (FBS) was
purchased from HyClone (UT, USA). Monoclonal antibodies specific
for p38 mitogen-activated protein kinase (p38 MAPK); ERK1/2 and
their phosphor-forms, and FOXO3a and p53 were purchased from Cell
Signaling Technology (Beverly, MA, USA). The GAPDH, p53 and FOXO3a
monoclonal antibodies were obtained from Abcam Co (Burlingame, CA,
USA). PVDF membrane was purchased from Millipore (USA). PD98059 (a
special inhibitor of ERK1/2) was purchased from Merck Millipore
(Darmstadt, Germany), MTT powder and pifithrin-α (a special
inhibitor of p53) were purchased from Sigma-Aldrich (St. Louis, MO,
USA). p53 and FOXO3a siRNAs were obtained from Santa Cruz (CA,
USA). Lipofectamine 2000 reagent was purchased from Invitrogen
(Shanghai, China). The FOXO3a-GFP and N1-GFP plasmids were kindly
provided by Frank M.J. Jacobs (Rudolf Magnus Institute of
Neuroscience, Department of Pharmacology and Anatomy, University
Medical Center, Utrecht) and was reported previously (14). Annexin V-FITC Apoptosis Kit was
purchased from Bestbio Co. (Shanghai, China).
Cell culture
Human NPC cell lines CNE2 and HNE2 were obtained
from the Cell Line Bank at the Laboratory Animal Center of Sun
Yat-sen University starting March, 2012 (Guangzhou, China). All
cell lines have been tested and authenticated for absence of
mycoplasma, genotypes, drug response, and morphology using a
commercially available kit (Invitrogen, Shanghai, China) in the
Laboratory Animal Center at Sun Yat-sen University in April, 2010.
Cells were cultured in RPMI-1640 medium, supplemented with 10%
fetal bovine serum (FBS), 5% glutamine, 100 U/ml penicillin and 100
mg/ml streptomycin. In all experiments, 60–70% of confluent cells
were washed and incubated with curcumin, PD98059, pifithrin-α for
the indicated time. Curcumin was dissolved in DMSO and the final
concentration was <1‰ (v/v) in all experiments.
MTT assay
Cell viability was analyzed by the MTT [3-(4,
5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide] assay.
Briefly, cells were seeded in 96-well plates at the density of
2×103 cells/well and were cultured with increased
concentrations of curcumin for up to 72 h, and then 10 μl of
10 mg/ml MTT solution was added to each well for an additional 4 h
according to the manufacturer’s instructions (Promega, Shanghai,
China). After centrifugation, 150 μl dimethyl sulfoxide was
added to the precipitate and the absorbance of the enzyme was
measured at 570 nm using a Microplate Reader (Bio-Rad, Hercules,
CA, USA). Cell growth rates (average absorbance of each treat group
and non-treated group) were then calculated. All experiments were
performed in triplicate and repeated at least three times.
Hoechst 33258 staining
Following treatment with curcumin at various
concentrations for up to 48 h, cells were washed twice with PBS and
fixed in 1 ml of 4% paraformaldehyde for 10 min at 4°C. After
washing twice with PBS, cells were stained with 100 μl (10
μg/ml) Hoechst 33258 (Sigma, St. Louis, MO, USA) in PBS for
15 min at room temperature in the dark, and then washed with PBS.
Afterwards, the cells were mounted and examined under fluorescence
microscopy (Olympus IX71, Tokyo, Japan). Apoptotic cells were
identified by the condensation and fragmentation of their
nuclei.
Flow cytometry
Cells (1×105 cells/dish) were seeded in
6-mm culture plate and treated with increased doses of curcumin as
indicated for up to 48 h. The adherent and floating cells were both
collected and resuspended in cold PBS for analysis. Cells were
stained with Annexin V-FITC apoptosis detection kit to monitor
apoptosis cells and propidium iodide (PI) to detect dead cells
according to the instructions from the provider. Samples were
analyzed on the FC500 Flow cytometry Systems (Beckman Coulter,
USA).
Western blot analysis
After being treated with curcumin, cells were
harvested and washed with ice-cold phosphate buffer, homogenized in
1X RIPA lysis buffer. Equal amounts of protein from whole cell
lysates were solubilized in 2X SDS-sample buffer, separated on
SDS-polyacrylamide gels. The separated proteins were transferred
onto nitrocellulose using a Bio-Rad Trans Blot semidry transfer
apparatus for 1 h at 25 V, blocked with Blotto with 5% no-nfat dry
milk and 0.1% Tween-20 overnight at 4°C, and washed with wash
buffer. Blots were incubated with polyclonal antibodies against p38
MAPK, ERK1/2 and their phosphor-forms, and FOXO3a and p53 (1:1,000)
overnight at 37°C, washed and incubated with a secondary antibody
raised against rabbit IgG conjugated to horseradish peroxidase
(1:2,000, Sigma, Beijing, China) for 1 h at room temperature. The
washed blots were transferred to freshly made ECL Prime (Pierce,
Rockford, IL, USA) and exposed to X-ray film.
Treatment with p53 and FOXO3a small
interfering RNAs
For the transfection procedure, cells were grown to
60% confluence, and p53 and FOXO3a siRNAs and control siRNA were
transfected using the oligofectamine reagent (Invitrogen). CNE2
cells (2×105 cells/ml) were seeded in RPMI-1640 medium
without antibiotics overnight. After washing the cells with PBS, 2
ml of media without antibiotics were added. Thereafter, 200
μl of Lipofectamine 2000 complex was added into each plate.
The cells were transfected with control or FoxO3a (FKHRL1) and p53
small interfering RNAs (siRNAs) for 24 h according to instructions
from the manufacturer. At 24 h after transfection, medium was
replaced for complete DMEM and cells were transfected with either
exogenous FOXO3a gene using electroporated transfection
method or treated with curcumin 40 μM for an additional 24 h
for other experiments.
Electroporated transfection assays
CNE2 cells (1×107 cells/ml) were
transferred into conical tubes and centrifuged at 1,200 rpm for 5
min. After centrifuging, the medium was removed and the cells were
washed with 1X PBS, and centrifuged again at 1,200 rpm for 5 min.
Afterwards, the PBS was aspirated and added to Bio-Rad Gene Pulser
electroporation buffer. After resuspending the cells, the desired
N1-GFP or FoxO3a-GFP plasmid DNA at a final concentration of 10 to
20 μg/ml was added and the electroporation plate was put in
the MXcell plate chamber. The electroporation conditions on the
plates to deliver 160 V/10 ms square wave were adjusted until
reaching the optimum. The conditions were set and loaded onto the
device Gene Pulser II Electroporation System (Bio-Rad, CA, USA).
After electroporation was completed, the cells were transferred to
a tissue culture plate. We typically transferred each 150 μl
electroporation sample to a 6-well tissue culture plate containing
2 ml RPMI-1640. Cells were then incubated for 48 h at 37°C, then
treated with curcumin for an additional 24 h.
Statistical analysis
Analysis of variance and Student’s t-test were used
to compare the values of the test and control samples. P<0.05
was considered a statistically significant difference. SPSS
software was used for all statistical analysis. The significance
was evaluated by the paired t-test. All the experiments were
performed at least three times, and mean values and standard
deviation were calculated.
Results
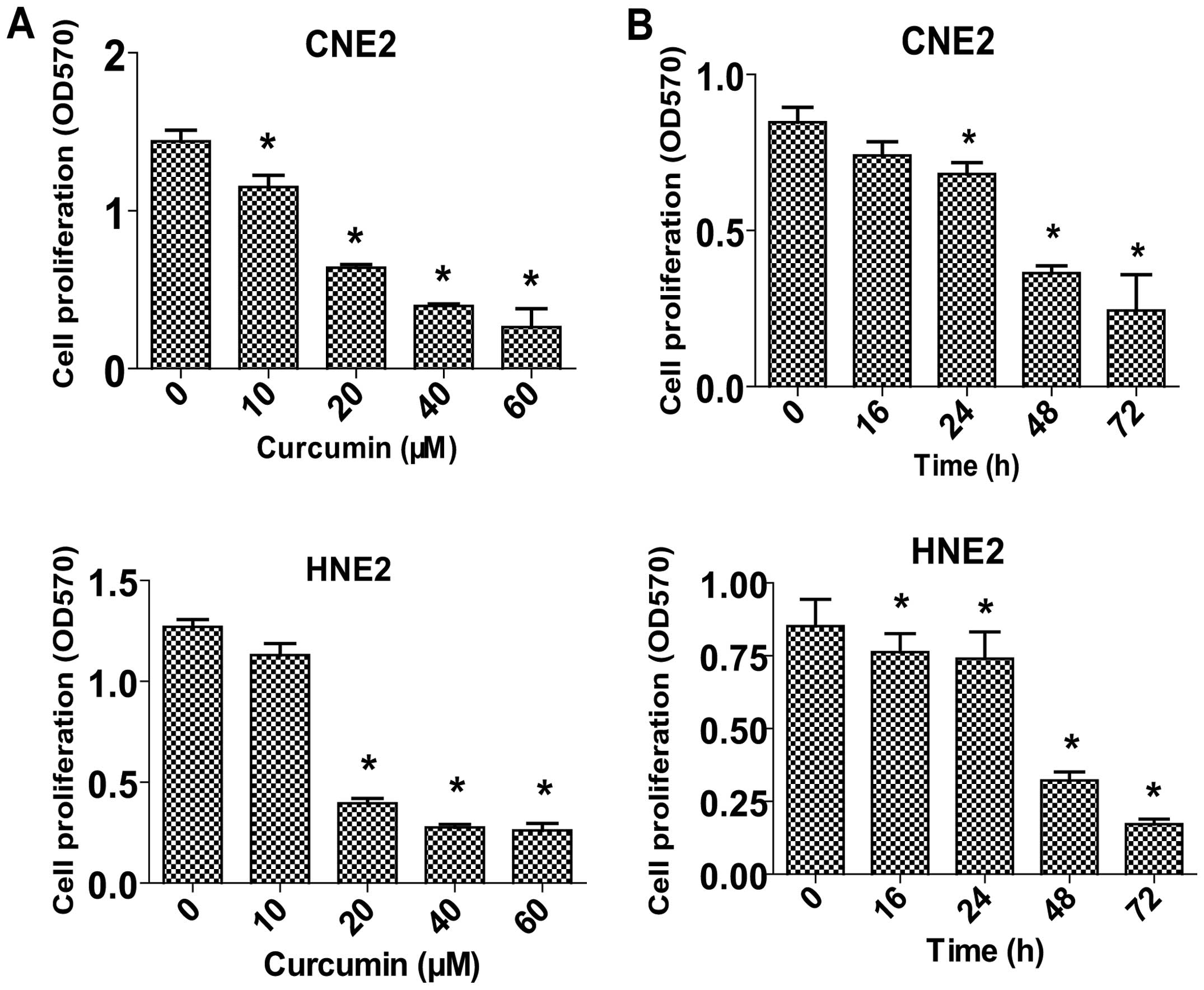
Curcumin inhibits proliferation of NPC
cells in a dose- and time-dependent manner
To determine whether curcumin may regulate the
proliferation of NPC cells, we examined the effect of curcumin on
cell proliferation in two human NPC cell lines, CNE2 and HNE2, by a
MTT assay. The results showed that treatment with increased
concentrations of curcumin for up to 72 h significantly inhibited
cell proliferation in a dose- and time-dependent manner (Fig. 1A and B).
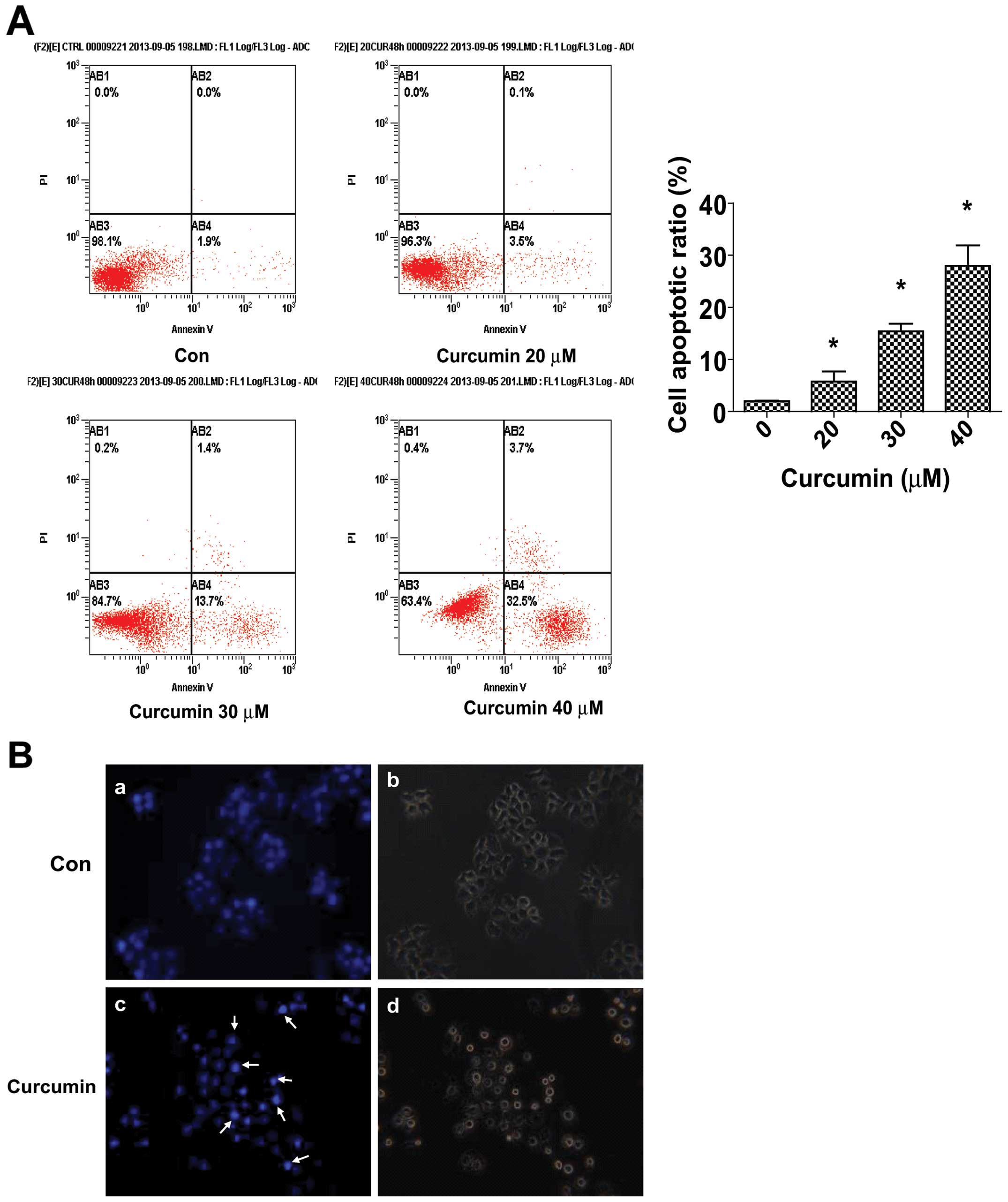
Curcumin increases apoptosis of CNE2
cells
We evaluated the effects of curcumin on apoptosis in
CNE2 cells using Annexin V-FITC and PI staining. In normal live
cells, phosphatidy serine (PS) was located on the cytoplasmic
surface of the cell membrane. However, in the apoptotic cells, PS
was found to be translocated from the inner to the outer leaflet of
the plasma membrane, thus exposing PS to the external cellular
environment. The human anticoagulant, Annexin V, is a 35–36 Kda
Ca+-dependent phospholipid binding protein that has a
high affinity for PS. Annexin V labeled with a fluorophore or
biotin can identify apoptotic cells. In the scatter plot of double
variable flow cytometry, AB3 quadrant
(FITC−/PI−) showed living cells; AB2 quadrant
(FITC+/PI+) stand for late apoptotic cells;
and AB4 quadrant (FITC+/PI−) represents early
apoptotic cells. As shown in Fig.
2A, a marked dose-dependent increase in both the early and late
stages of apoptosis was observed in CNE2 cells after curcumin
treatment compared to the control cells.
Morphological changes in the apoptotic cells were
revealed by the Hoechst 33258 staining (Fig. 2B). In the untreated CNE2 cells, the
nuclei were stained weakly, and homogeneously blue, whereas, in
cells treated with curcumin some bright chromatin condensation and
nuclear fragmentation were observed.
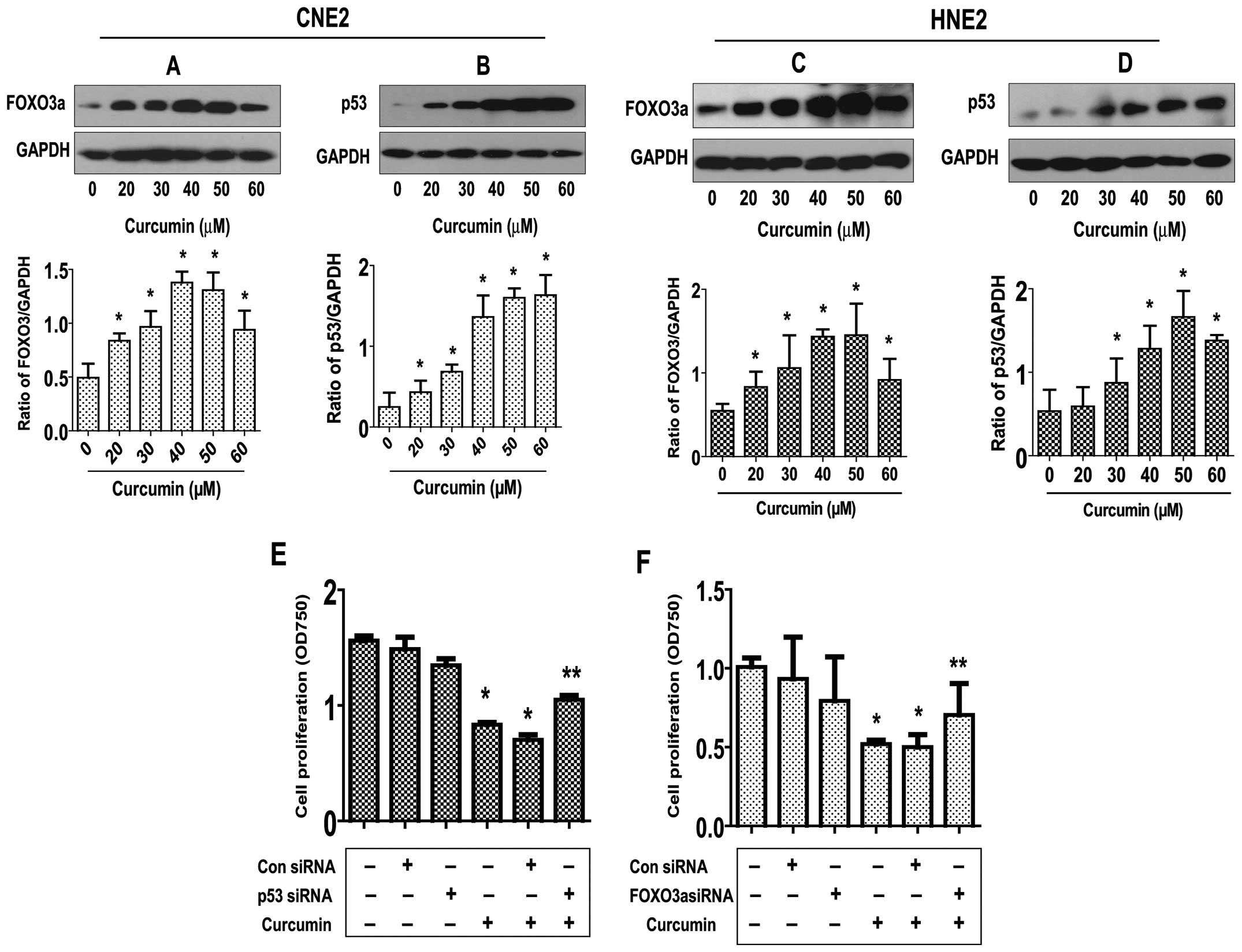
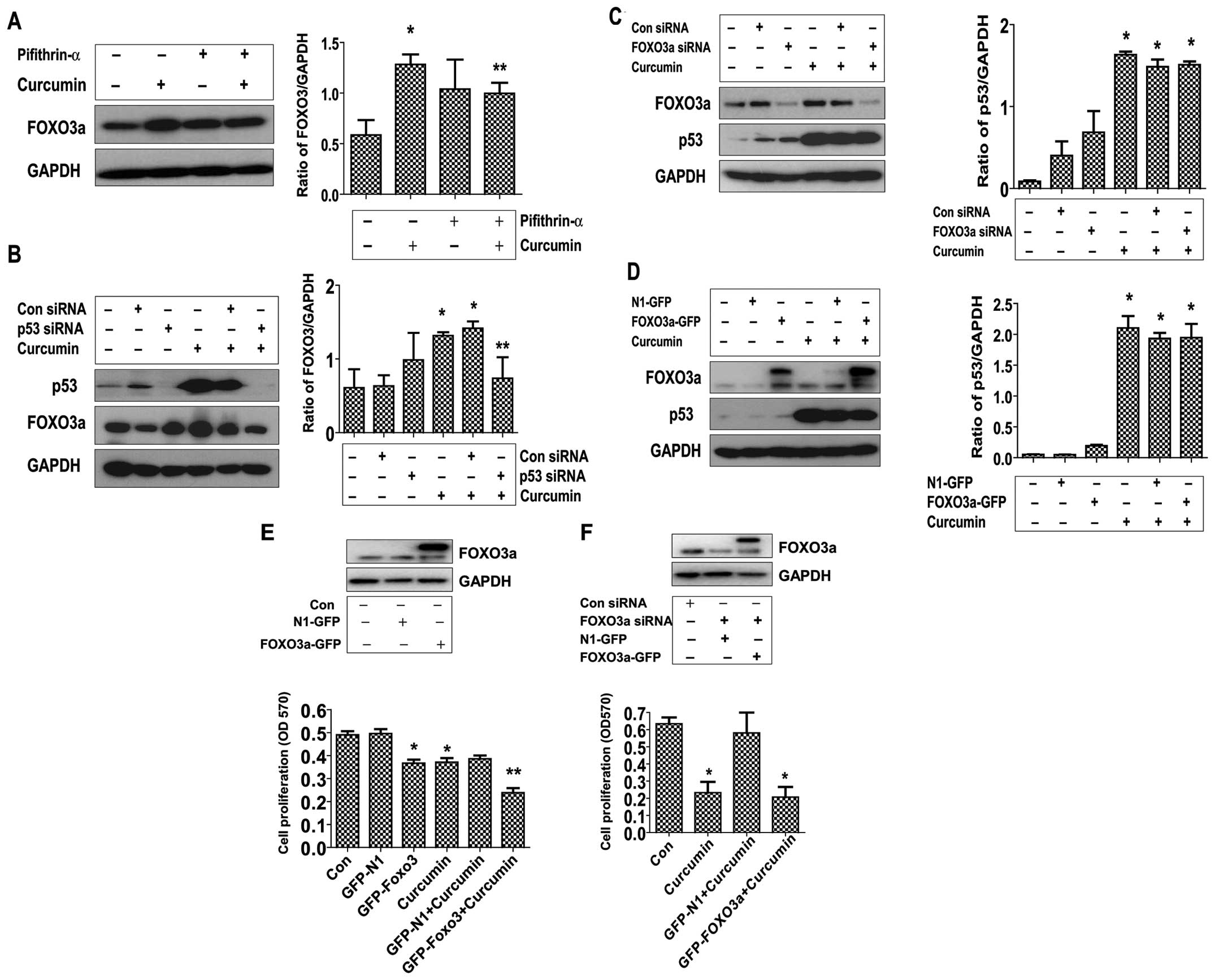
Curcumin increases FOXO3a and p53 protein
expression; silencing of FOXO3a and p53 abrogates the inhibitory
effect of curcumin on growth of NPC cells
To elucidate the potential molecular mechanism
underlying this effect, we tested the effect of curcumin on the
expression of p53 and FOXO3a active proteins, these tumor
suppressors have been shown to be involved in cell proliferation
and apoptosis (5,12). As shown in Fig. 3A and B, curcumin induced the
protein expression of both FOXO3a (Fig. 3A) and p53 (Fig. 3B) dose-dependently in CNE2 cells,
with maximal induction observed at 40–50 μM, respectively.
Note that similar results were also observed in an additional cell
line (HNE2) (Fig. 3C and D).
To further explore the role of these tumor
suppressors in mediating the curcumin-inhibited NPC cell
proliferation, we blocked p53 and FOXO3a gene expression by
transfecting CNE2 cells with p53 and FOXO3a siRNAs and evaluated
the effects of curcumin on cell proliferation. We demonstrated that
the knockdown of p53 and FOXO3a genes significantly overcame the
curcumin-mediated growth inhibition of CNE2 cells (Fig. 3E). This indicated important roles
of p53 and FOXO3a expression in curcumin-inhibited cell growth.
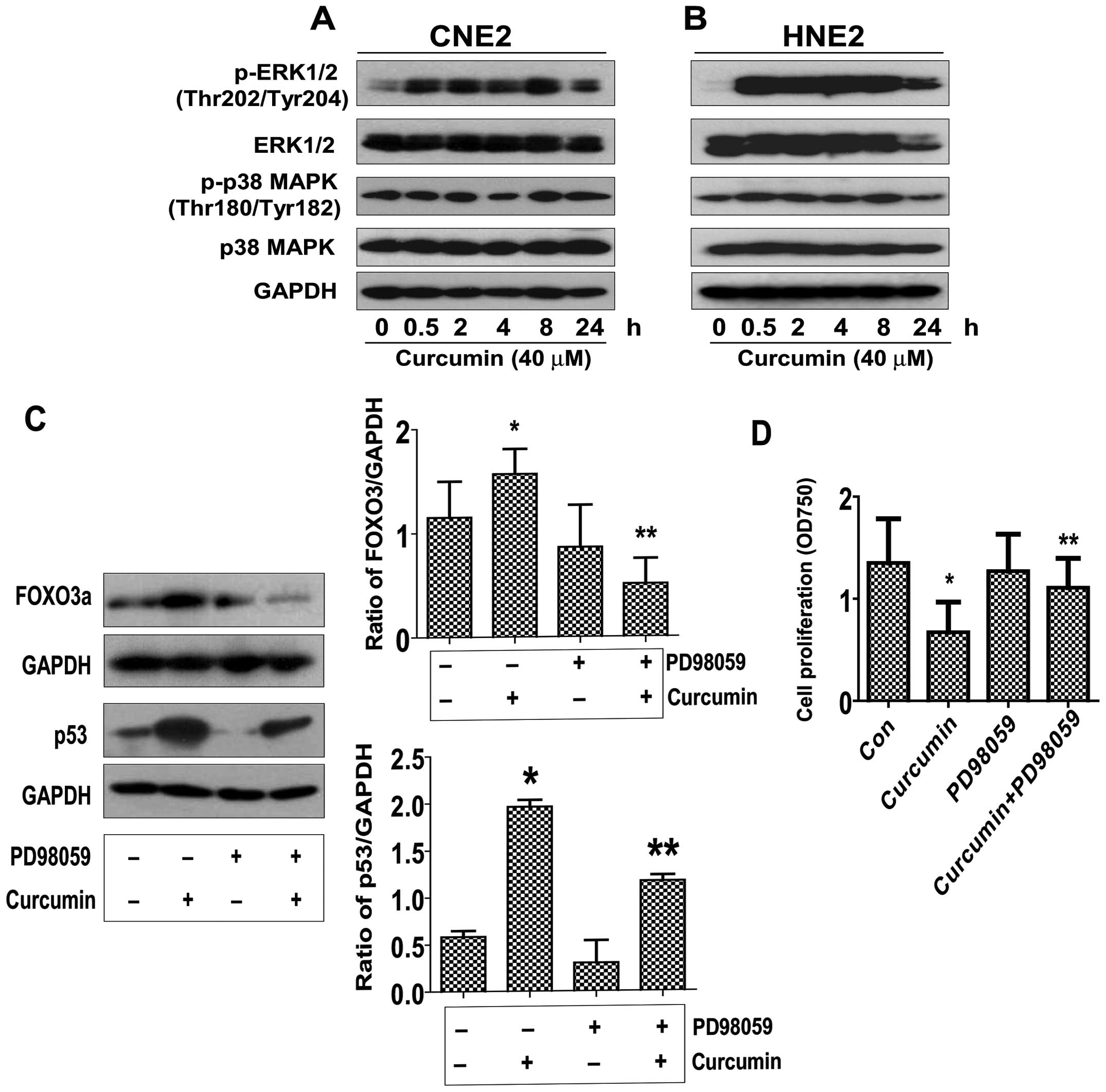
Curcumin induces ERK/MAPK activation,
blockade of ERK1/2 abrogated the effect of curcumin on FOXO3a and
p53 expression, while restoring the cell growth in the presence of
curcumin in NPC cells
In order to determine the potential signaling
pathways that are involved in the curcumin-mediated regulation of
FOXO3a and p53 expression, thereby controlling cell proliferation,
we initially evaluated the effect of curcumin on the activation of
ERK and p38 MAPK by western blot analysis. As shown in Fig. 4A, treatment with curcumin at 40
μM significantly increased the phosphorylation of ERK1/2
starting at 30 min and continued for up to 24 h, whereas the level
of total ERK1/2 protein was unchanged. However, curcumin had little
effect on the activation and protein expression of p38 MAPK.
Similar results were also observed in an additional NPC cell line
(HNE2) (Fig. 4B). Interestingly,
we showed that the ERK inhibitor (PD98059) exposed to the cells
abolished the curcumin-induced FOXO3a and p53 protein expression in
CNE2 cells (Fig. 4C). Moreover,
treatment of PD98059 restored the cell growth in the presence of
curcumin (Fig. D). Together, the
above results suggested that activation of ERK signaling pathway
was critical in mediating the effect of curcumin on regulating
FOXO3a and p53 expression, and cell growth.
p53 is upstream of FOXO3a; exogenous
expression of FOXO3a restores the effect of curcumin on growth of
cells silenced by endogenous FOXO3a gene
Transcriptional activity of FOXO3a could be
modulated though interactions with other transcriptional factors
such as tumor suppressor p53 (15), thus, we assessed if curcumin
affected the interactions of FOXO3a with p53. As shown in Fig. 5, silencing and blockade of p53
using either siRNA or specific p53 inhibitor overcame the induced
effect of curcumin on the expression of FOXO3a protein (Fig. 5A and B). On the contrary, silencing
or overexpression of FOXO3a had no effect on curcumin-induced p53
protein expression (Fig. 5C and
D), but strengthened the inhibitory effect of curcumin on cell
growth (Fig. 5E). Interestingly,
exogenous expression of FOXO3a restored the effect of curcumin on
growth of cells silenced by endogenous FOXO3a gene (Fig. 5F). Note that western blot analysis
confirmed the exogenous expression of FOXO3a protein transfected
into the cells (Fig. 5D–F).
Discussion
Studies have shown that curcumin can inhibit the
growth of a variety of tumor cells, as well as induce cell
apoptosis in several tumors including NPC cells (16–20)
suggesting that curcumin can be used as a natural antitumor agent.
However, the detailed mechanisms by which this agent targets NPC
cancer cells remained unclear. In this study, we evaluated the
response of NPC cells to curcumin treatment. Our results indicated
that curcumin inhibited NPC cell proliferation and induced
apoptosis in a dose- and time-dependent manner suggesting a tumor
suppressor property of this agent. Of note, the concentrations of
curcumin used here were consistent with or even lower then those
reported by others demonstrating significant responses in different
cell systems (21–23) although lower doses were also
reported in other studies (24,25).
We realized that a higher dose was needed to inhibit different
cancer cell growth, but this was within the range of those reported
by others and showed no toxicity (21–23).
In our study we demonstrated the role of p53 and
FOXO3a protein induction that mediated the effect of curcumin on
the inhibition of NPC cell growth. As tumor suppressors, both
transcription factors play important roles in several areas
including gene regulation, cell growth and apoptosis (5,12).
Striking similarities have been reported between p53 and FOXO, such
as post-translational modifications, common signaling pathways,
target genes, and similar mutual interactions with various proteins
(26). We found that blockade of
the activity of p53 or silencing of either p53 or FOXO3a gene
partially overcame the inhibitory effect of curcumin on NPC cell
proliferation, suggesting that induction of these two molecules
contributed to mediation of the effect of curcumin on NPC cell
growth inhibition. Whether curcumin affected the post-translational
modifications, such as phosphorylation, acetylation and
ubiquitination, of either p53 or FOXO3a, thereby regulating their
subcellular localization and transcriptional activities require
further study. Consistent with this, other studies also found the
link of curcumin and p53 or FOXO3a expression, and demonstrated the
role of these transcription factors in mediating the effect of
curcumin on controlling cell proliferation and other functions in
other cell systems (27,28). We reasoned that more studies are
required to explore the precise mechanism of p53 and FOXO3a
expression, regulation and downstream pathways in mediating the
overall response of curcumin.
The MAPK signaling pathway plays a key role in the
regulation of gene expression, cellular growth and survival
(29). Data from others indicated
that curcumin activates MAPK signaling pathways, and that
activation of MAPK, such as ERK and p38 MAPK, links
curcumin-mediated signaling to the transcriptional regulation of
genes that are crucial for cell growth inhibition (30,31).
Our result identified an important role of ERK activation in
mediating the inhibitory effect of curcumin on p53 and FOXO3a
protein expression and NPC cell growth inhibition. However, p38
MAPK played no role in our study, which differed from others
(31,32). The discrepancy remain unclear, the
use of different cell lines and the culture conditions may account
for this, which needs to be further determined. We reasoned that
curcumin may function through activating ERK/MAPK signaling
pathway, followed by upregulation of p53 and FOXO3a protein
expression, thereby inhibiting NPC cell proliferation and inducing
apoptosis. However, inhibition of ERK/MAPK reversed the effect of
curcumin on NPC proliferation and could be independent of p53 and
FOXO3a signaling. Also, the potential downstream targets of p53 and
FOXO3a may be involved in this process. Thus, more studies are
required to further elucidate this.
The aforementioned uncovered that both p53 and
FOXO3a played a causative role in mediating the effect of curcumin
in controlling NPC cell growth and perhaps inducing apoptosis.
Furthermore, our results showed that exogenous expression of FOXO3a
could enhance the effect of curcumin on cell growth, and more
importantly, restore the inhibitory effect of curcumin on growth of
NPC cells silenced by endogenous FOXO3a gene in siRNA approach,
while having no further effect on curcumin-induced p53 expression.
This further confirmed the role of FOXO3a in mediating the effect
of curcumin on cell growth; it also suggested that p53 is upstream
of FOXO3a and induction of FOXO3a by curcumin is p53-dependent. We
speculated that p53 and FOXO may have parallel functions that
worked in concert to mediate the effect of curcumin on control
growth of NPC cells. The p53 and FOXO3a interaction have been
identified (26), however, the
extent and importance of the functional interaction between these
two have not been fully explored. Consistent with our report, other
studies found that FOXO3a was a p53 target gene and that
transcriptional activity of FOXO3a was regulated by p53, while the
latter was not affected by FOXO3a (33,34).
The p53 and FOXO3a formed part of regulation transcriptional
network to control cancer cell growth and apoptosis (33,34).
In addition, curcumin induced expression of p53 or/and FOXO3a in
inhibition of cancer cell growth and other functions have been
shown in other cell systems (27,28,35,36).
However, whether curcumin affects the interaction of these proteins
to facilitate the inhibitory effect of NPC growth remain to be
determined. A number of signaling pathways mediated the expression
of FOXO3a and p53, and critically regulated apoptotic events, which
included PI3-K/Akt and cyclin-dependent kinase (CDK) (37,38).
We believed that understanding the functional interactions of p53
and FOXO3a, their regulated signaling pathways and known downstream
targets will be crucial to better elucidate their roles in
mediating the overall response of curcumin.
In summary, our data show that curcumin inhibits the
growth and induces apoptosis of NPC cells through activation of the
ERK signaling pathway; this leads to increase in protein expression
of p53 and FOXO3a. p53 is upstream of FOXO3a and induction of
FOXO3a by curcumin is p53-dependent. Both transcription factors and
tumor suppressors form a regulatory loop that work in concert to
mediate the effect of curcumin. This study unveils a new mechanism
by which curcumin inhibits the proliferation and induces apoptosis
of NPC cells.
Acknowledgements
This study was supported in part by
the Research Fund from Guangdong Province Administration of
Traditional Chinese Medicine (20132149), the Special Science and
Technology Join fund from Guangdong Provincial Department of
Science and Technology-Guangdong Academy of Traditional Chinese
Medicine (2012A032500011) and a grant from the National Nature
Scientific Foundation of China (81272614).
References
|
1.
|
Xu Y, Zhang J, Shi W and Liu Y: Anticancer
effects of 3,3′-diindolylmethane are associated with G1 arrest and
mitochondria-dependent apoptosis in human nasopharyngeal carcinoma
cells. Oncol Lett. 5:655–662. 2013.
|
|
2.
|
Pan Y, Zhou F, Zhang R and Claret FX:
Stat3 inhibitor Stattic exhibits potent antitumor activity and
induces chemo- and radio-sensitivity in nasopharyngeal carcinoma.
PloS One. 8:e545652013. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Aggarwal BB, Sundaram C, Malani N and
Ichikawa H: Curcumin: the Indian solid gold. Adv Exp Med Biol.
595:1–75. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Wilken R, Veena MS, Wang MB and Srivatsan
ES: Curcumin: A review of anti-cancer properties and therapeutic
activity in head and neck squamous cell carcinoma. Mol Cancer.
10:122011. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Nayak G and Cooper GM: p53 is a major
component of the transcriptional and apoptotic program regulated by
PI 3-kinase/Akt/GSK3 signaling. Cell Death Dis. 3:e4002012.
View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Hollstein M, Sidransky D, Vogelstein B and
Harris CC: p53 mutations in human cancers. Science. 253:49–53.
1991. View Article : Google Scholar
|
|
7.
|
Zeng GQ, Yi H, Li XH, et al:
Identification of the proteins related to p53-mediated
radioresponse in nasopharyngeal carcinoma by proteomic analysis. J
Proteomics. 74:2723–2733. 2011. View Article : Google Scholar
|
|
8.
|
Shen YA, Lin CH, Chi WH, et al:
Resveratrol impedes the stemness, epithelial-mesenchymal
transition, and metabolic reprogramming of cancer stem cells in
nasopharyngeal carcinoma through p53 activation. Evid Based
Complement Alternat Med. 2013:5903932013.
|
|
9.
|
Brunet A, Bonni A, Zigmond MJ, et al: Akt
promotes cell survival by phosphorylating and inhibiting a Forkhead
transcription factor. Cell. 96:857–868. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Schmidt M, Fernandez de Mattos S, van der
Horst A, et al: Cell cycle inhibition by FoxO forkhead
transcription factors involves downregulation of cyclin D. Mol Cell
Biol. 22:7842–7852. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Obsil T and Obsilova V: Structural basis
for DNA recognition by FOXO proteins. Biochim Biophys Acta.
1813.1946–1953. 2011.PubMed/NCBI
|
|
12.
|
Yang JY, Zong CS, Xia W, et al: ERK
promotes tumorigenesis by inhibiting FOXO3a via MDM2-mediated
degradation. Nat Cell Biol. 10:138–148. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Shou Z, Lin L, Liang J, Li JL and Chen HY:
Expression and prognosis of FOXO3a and HIF-1alpha in nasopharyngeal
carcinoma. J Cancer Res Clin Oncol. 138:585–593. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Wen Q, Duan X, Liao R, et al:
Characterization of intracellular translocation of Forkhead
transcription factor O (FoxO) members induced by NGF in PC12 cells.
Neurosci Lett. 498:31–36. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Wang F, Marshall CB, Yamamoto K, et al:
Biochemical and structural characterization of an intramolecular
interaction in FOXO3a and its binding with p53. J Mol Biol.
384:590–603. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Ye F, Zhang GH, Guan BX and Xu XC:
Suppression of esophageal cancer cell growth using curcumin,
(−)-epigallocatechin-3-gallate and lovastatin. World J
Gastroenterol. 18:126–135. 2012.
|
|
17.
|
Subramaniam D, Ponnurangam S, Ramamoorthy
P, et al: Curcumin induces cell death in esophageal cancer cells
through modulating Notch signaling. PloS One. 7:e305902012.
View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Quitschke WW: Curcuminoid binding to
embryonal carcinoma cells: reductive metabolism, induction of
apoptosis, senescence, and inhibition of cell proliferation. PloS
One. 7:e395682012. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Sahu RP, Batra S and Srivastava SK:
Activation of ATM/Chk1 by curcumin causes cell cycle arrest and
apoptosis in human pancreatic cancer cells. Br J Cancer.
100:1425–1433. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Bao B, Ali S, Banerjee S, et al: Curcumin
analogue CDF inhibits pancreatic tumor growth by switching on
suppressor microRNAs and attenuating EZH2 expression. Cancer Res.
72:335–345. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Chen CC, Sureshbabul M, Chen HW, et al:
Curcumin suppresses metastasis via Sp-1, FAK inhibition, and
E-cadherin upregulation in colorectal cancer. Evid Based Complement
Alternat Med. 2013:5416952013.PubMed/NCBI
|
|
22.
|
Wang D, Hu J, Lv L, Xia X, Liu J and Li X:
Enhanced inhibitory effect of curcumin via reactive oxygen species
generation in human nasopharyngeal carcinoma cells following
purple-light irradiation. Oncol Lett. 6:81–85. 2013.
|
|
23.
|
Sundram V, Chauhan SC, Ebeling M and Jaggi
M: Curcumin attenuates beta-catenin signaling in prostate cancer
cells through activation of protein kinase D1. PloS One.
7:e353682012. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Jiang M, Huang O, Zhang X, et al: Curcumin
induces cell death and restores tamoxifen sensitivity in the
antiestrogen-resistant breast cancer cell lines MCF-7/LCC2 and
MCF-7/LCC9. Molecules. 18:701–720. 2013. View Article : Google Scholar
|
|
25.
|
Blakemore LM, Boes C, Cordell R and Manson
MM: Curcumin-induced mitotic arrest is characterized by spindle
abnormalities, defects in chromosomal congression and DNA damage.
Carcinogenesis. 34:351–360. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
You H and Mak TW: Crosstalk between p53
and FOXO transcription factors. Cell Cycle. 4:37–38. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Qiao Q, Jiang Y and Li G: Curcumin
enhances the response of non-Hodgkin’s lymphoma cells to ionizing
radiation through further induction of cell cycle arrest at the
G2/M phase and inhibition of mTOR phosphorylation. Oncol Rep.
29:380–386. 2013.PubMed/NCBI
|
|
28.
|
Singh AT, Ghosh M, Forte TM, Ryan RO and
Gordon LI: Curcumin nanodisk-induced apoptosis in mantle cell
lymphoma. Leuk Lymphoma. 52:1537–1543. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Sebolt-Leopold JS, Herrera R and Ohren JF:
The mitogen-activated protein kinase pathway for molecular-targeted
cancer treatment. Recent Results Cancer Res. 172:155–167. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Li R, Wang Y, Liu Y, et al: Curcumin
inhibits transforming growth factor-beta1-induced EMT via PPARgamma
pathway, not Smad pathway in renal tubular epithelial cells. PloS
One. 8:e588482013. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Kang D, Park W, Lee S, Kim JH and Song JJ:
Crosstalk from survival to necrotic death coexists in DU-145 cells
by curcumin treatment. Cell Signal. 25:1288–1300. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Wang WZ, Li L, Liu MY, et al: Curcumin
induces FasL-related apoptosis through p38 activation in human
hepatocellular carcinoma Huh7 cells. Life Sci. 292:352–358. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Renault VM, Thekkat PU, Hoang KL, et al:
The pro-longevity gene FoxO3 is a direct target of the p53 tumor
suppressor. Oncogene. 30:3207–3221. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Miyaguchi Y, Tsuchiya K and Sakamoto K:
P53 negatively regulates the transcriptional activity of FOXO3a
under oxidative stress. Cell Biol Int. 33:853–860. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Guo LD, Chen XJ, Hu YH, Yu ZJ, Wang D and
Liu JZ: Curcumin inhibits proliferation and induces apoptosis of
human colorectal cancer cells by activating the mitochondria
apoptotic pathway. Phytother Res. 27:422–430. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Zingg JM, Hasan ST, Cowan D, Ricciarelli
R, Azzi A and Meydani M: Regulatory effects of curcumin on lipid
accumulation in monocytes/macrophages. J Cell Biochem. 113:833–840.
2012. View Article : Google Scholar
|
|
37.
|
Tudzarova S, Trotter MW, Wollenschlaeger
A, et al: Molecular architecture of the DNA replication origin
activation checkpoint. EMBO J. 29:3381–3394. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Amente S, Zhang J, Lavadera ML, Lania L,
Avvedimento EV and Majello B: Myc and PI3K/AKT signaling
cooperatively repress FOXO3a-dependent PUMA and GADD45a gene
expression. Nucleic Acids Res. 39:9498–9507. 2011. View Article : Google Scholar
|