Introduction
The main function of fibroblasts is secreting the
components of extracellular matrix (ECM). Without activation,
fibroblasts are dormant in the ECM, however, once activated,
fibroblasts could be involved in secreting higher levels of ECM
components and exert a stronger proliferative effect (1,2). In
the 1970s, Ryan et al originally described the activation of
fibroblasts in granulation tissues during wound healing progression
(3,4). Activated fibroblasts are also
referred to as myofibroblasts due to their expression of α-smooth
muscle actin (α-SMA), as well as other important functions
(5,6).
Carcinoma tissues are mainly composed of tumor cells
and stromal cells, the latter include fibroblasts, endothelial and
inflammatory cells. Fibroblasts, termed cancer-associated
fibroblasts (CAFs) represent the most abundant cellular components
in cancer stroma. The heterotypic and multicellular interactions
among cells, soluble factors, signaling molecules, extracellular
matrix construct a new biological system termed the ‘tumor
microenvironment (TME)’. Researchers have demonstrated that tumor
cells and the TME coevolve through continuous paracrine
communication (7), which not only
creates a dynamic signaling circuitry that promotes cancer
initiation and progress, but also induces the activation of
fibroblasts (8). CAFs present in
cancer stroma exhibit an activated phenotype analogous to that of
fibroblasts involved in wound healing or fibrosis (9,10).
Indeed, Dvorak originally considered tumors to be ‘wounds that do
not heal’, because of the similarity with granulation tissue
(11). Increasing evidence
supports the role of CAFs as a key regulator of the paracrine
signaling required for cancer progression (12,13).
In contrast to resting fibroblasts, activated CAFs can be
identified by their expression of vimentin, α-SMA, fibroblasts
activated protein (FAP), and platelet-derived growth factor
receptor (PDGFR-β) (14–16). Kalluri has indicated that up to 80%
of stromal fibroblasts in breast cancer display this activated
phenotype (17). Furthermore,
evidence indicates that CAFs neither revert back to normal
fibroblasts nor undergo elimination via apoptosis (18).
CAFs are further characterized by their production
of abundant ECM proteins, which are responsible for the stiffening
appearance of cancer (19,20). The deposition of ECM induced by
CAFs in tumor stroma is known as desmoplasia, which has been shown
to be extensive in or around tumors, particularly in human breast
cancer and pancreatic carcinoma (21,22).
A study has shown that the compression of ECM in turn leads to the
concentration of soluble factors that promote tumorigenesis in an
autocrine and paracrine manner (13). However, Walker has reported that
the stroma in a range of primary breast carcinomas could vary from
being predominantly cellular with little collagens to being a dense
collagenous stroma with apparently few stromal cells (23). These results are contradictory to
many research results on the universal deposition of ECM in cancer
tissue. Therefore, it is necessary to elucidate the relationship
between activated CAFs and their function in ECM deposition. In
this study, we aimed to investigate the secretomics of CAFs by mass
spectrometry and other means in order to elucidate the relationship
between CAFs and ECM deposition in breast cancer.
Materials and methods
Specimens
This study was based on a well-characterized series
of TNM stage T2–T3 primary invasive breast carcinoma cases and
fibroadenomas. Invasive breast cancer was selected for study due to
its typical desmoplastic response and the presence of large numbers
of activated CAFs. The fresh specimens of invasive carcinoma were
collected from cancer patients who accepted radical mastectomy at
the Second Affiliated Hospital of Zhejiang University. Written
consent was obtained from all patients and this study was approved
by the ethics committee of Zhejiang University. Tissue specimens
used in immunohistochemistry and histochemistry were obtained from
the tissue bank of the Cancer Institute, Zhejiang University.
Immunohistochemistry (IHC) and
histochemistry
In total 160 cases of invasive breast cancer, 40
adjacent normal breast tissue and 6 fibroadenoma specimens were
obtained. After the tissues were fixed with 4% paraformaldehyde and
paraffin-embedded, sections (5 μm) were prepared, and IHC was
performed with monoclonal antibodies against α-SMA and vimentin
(Maxin). The IHC protocol was described as previously (24). H&E staining and Masson
trichrome staining (Maiwei) were performed on the three kinds of
specimens. Images were obtained using laser confocal and optical
microscopes.
Primary culture of normal fibroblasts
(NFs) and CAFs
Fresh invasive cancer specimens and paired adjacent
normal breast tissue samples (>3–5 cm away from the tumor) were
collected under sterile conditions. The specimens were sectioned
into approximately 1-mm3 pieces and placed in DMEM/F12
(Gibco) supplemented with 10% FBS (Gibco), antibiotics (100 U/ml
penicillin and 100 μg/ml streptomycin; Sigma), ascorbic acid (20
ng/ml), and FGF-basic (10 ng/ml; Sigma). After one week of
incubation in a humidified incubator with 5% CO2
atmosphere, tissue debris was removed. Once primary cells reached
80% confluence, they were harvested and reseeded in a flask.
Characterization of CAFs/NFs
CAFs and NFs were seeded into 24-well plates. The
culture medium was removed and cells were fixed for 20 min with
SafeFix solution (Sinai) once 70–90% confluence was reached,
followed by treating with 0.2% Triton X-100 for 15 min, and
incubating in non-immunone goat serum for 20 min. Rabbit
anti-vimentin, anti-FSP-1, anti-fibronectin antibodies and mouse
anti-α-SMA, anti-cytokeratin (pan) antibodies (Maxin) were used in
incubation at 4°C as the primary antibody. Anti-mouse and
anti-rabbit fluorescent secondary antibodies were then applied
accordingly. After rinsing, cells were counterstained with DAPI
(Sigma), and images were obtained with a laser scanning confocal
microscope. Immunocytochemistry by DAB staining was conducted. The
methods for western blot analysis are presented in the section
‘Western blot analysis’.
Cell growth evaluation
Briefly, 1,500 CAFs and NFs were seeded per well in
96-well plates. Cell viability was assessed daily by adding 20 μl
of 3-(4,5-dimethylthiazol-2-thiazyl)-2,5-diphenyl-tetrazolium
bromide (MTT; 5 mg/ml in PBS) to each well. MTT was also added to
the control wells without cells. Each sample included six
replicates. Following 4-h incubation at 37°C, the medium was
removed. An aliquot of 200 μl DMSO was transfered to each well, and
the plate was agitated for 15 min. The absorbance at 570 nm was
then detected. For MTT assays that were performed on the same day
with cell seeding, the cells were allowed to attach for 3 h before
the addition of MTT. Growth curves were constructed by plotting
absorbance (mean ± SD) against time.
Extraction of secretory proteins from
CAFs and NFs
When CAFs and NFs reached 70–80% confluence, the
medium was removed, and the flasks were washed gently three times
with PBS. Cells were then cultured in conditioned medium (CM;
serum-free, phenol red-free DMEM medium supplemented with ascorbic
acid 20 ng/ml). Following 18-h incubation, the CM was collected and
the cell debris was removed by centrifugation for 10 min, 4°C (200
× g), followed by sterile filtration (0.22 μm; Millipore). Protease
inhibitors were applied to prevent protein degradation. The protein
present in the CM was concentrated by an ultrafiltration (3,000 Da;
Millipore) and precipitated in acetone overnight at −20°C. The
sediment was then washed twice with acetone, followed by
resuspended in protein solution (RIPA, Beyotime). The protein
concentration was then measured with a standard Bradford protein
assay (Bio-Rad). Prior to the collection of condition medium,
β-galactosidase staining was performed to exclude senescent cells.
CM samples were collected from three pairs of homologous CAFs and
NFs.
MALDI TOF/TOF-MS for global screening of
secretory protein
In total, 200 μg of secretory proteins from CAFs and
NFs were separated by 12% SDS-PAGE, and the resulting gel was
stained with Commassie Blue Fast Staining Solution (Invitrogen).
Each lane was then cut into 15 even sections and digested as
previously reported (25).
Briefly, all sections were destained and dehydrated, and proteins
were reduced with dithiothreitol and alkylated with iodoacetamide
(IAA, Sigma). After alkylation, the samples were incubated with
sequencing-grade trypsin (Promega) at 37°C for 20 h. The peptides
were then subjected to extraction (50% acetonitrile, 5% formic
acid) and lyophilized under vacuum for MALDI TOF/TOF (Bruker
Ultraflextreme) detection. All of the peptides were retrieved from
the UniProtKB website.
Label-free quantitative assay for
secretory proteins of interest
Trypsin digestion
Protein samples from each fraction were reduced with
DTT and alkylated with IAA. After being diluted in a solution of
100 mM NH4HCO3, the protein mixture was
digested by sequencing-grade trypsin at 37°C for 20 h.
Desalting peptides and multiple
reaction monitoring (MRM) assay
The tryptic peptide mixture was desalted using a
porous C-18 reversed-phase resin (Pierce) according to the
manufacturer’s instructions. Eluted peptides were lyophilized and
redissolved in 10 μl 0.1% FA for 4000 QTRAP® LC/MS/MS
System and targeted proteomics assay (MRM assay model) (26).
Western blot analysis
Protein extraction
Whole intracellular proteins were extracted in
accordance with a standard protocol. Briefly, adherent cells
(within 8 passages) were washed twice with PBS, and then lysed in
RIPA and ultrasonicated on ice. The lysates were then centrifuged
for 30 min at 4°C (10,000 × g) and the supernatants were collected
and stored at −80°C. The extraction of secretory proteins was
performed as outlined in ‘Extraction of secretory proteins form
CAFs and NFs’ section.
Western blots
Equal amounts of protein samples from CAFs and NFs
(intracellular or secretory proteins) were separated on a 12%
SDS-PAGE gel, transferred to PVDF membranes (0.2 μm; Millipore),
and then incubated with various primary antibodies (α-SMA, PDGFR-β,
collagen α-I, collagen α-III, diluted 1:2,000 in 5% defatted milk,
GADPH, diluted 1:5,000) overnight at 4°C. The membranes were
blotted with HRP conjugated secondary antibodies (Epitomics),
developed using an ECL substrate (Pierce) and exposed to Kodak
Biomax MR film.
Bioinformatics analysis and statistical
analysis
MS data were analyzed using Analyst software
(version 1.5.1, AB Sciex), and sequences were searched in the
SwissProt database with Protein Pilot (version 4.0, AB Sciex). The
cellular localization of identified proteins was analyzed on the
basis of information available from UniprotKB. MRM propilot
(version 2.1, AB Sciex) and Protein Pilot were used to analyze the
amino acid sequence of trypsin-digested peptides and determine the
optimal peptide sequences for quantitative detection. Statistical
analyses were conducted with SPSS software (version 19.0).
Results
Fibroblast activation in breast
cancer
To characterize the activation of fibroblasts, the
expression of α-SMA was analyzed in invasive breast cancer, normal
breast tissue and fibroadenoma. We found the α-SMA positive
fibroblasts were widely present in the stroma of invasive breast
carcinoma compared to that of normal breast tissues and
fibroadenoma (Fig. 1). In normal
breast tissues, only the myoepithelial cells distributed along the
entire duct-lobular system and the smooth muscle cells around the
microvasculature were α-SMA positive, while vimentin was not
detected. It has been reported that fully differentiated
myoepithelial cells can vanished during cancer progression
(27). Therefore, the α-SMA
positive cells within the tumor stroma activated CAFs
(α-SMA+ vimentin+). A large number of
fibroblasts were detected in breast fibroadenoma with abundant ECM,
however, these fibroblasts were α-SMA-negative. In summary, our
results suggest that activation of fibroblasts only appeared in
invasive cancer, which is consistent with findings from previous
studies (13). Noteworthy, our
data showed that not all the CAFs showed α-SMA expression in the
immunofluorescence assay.
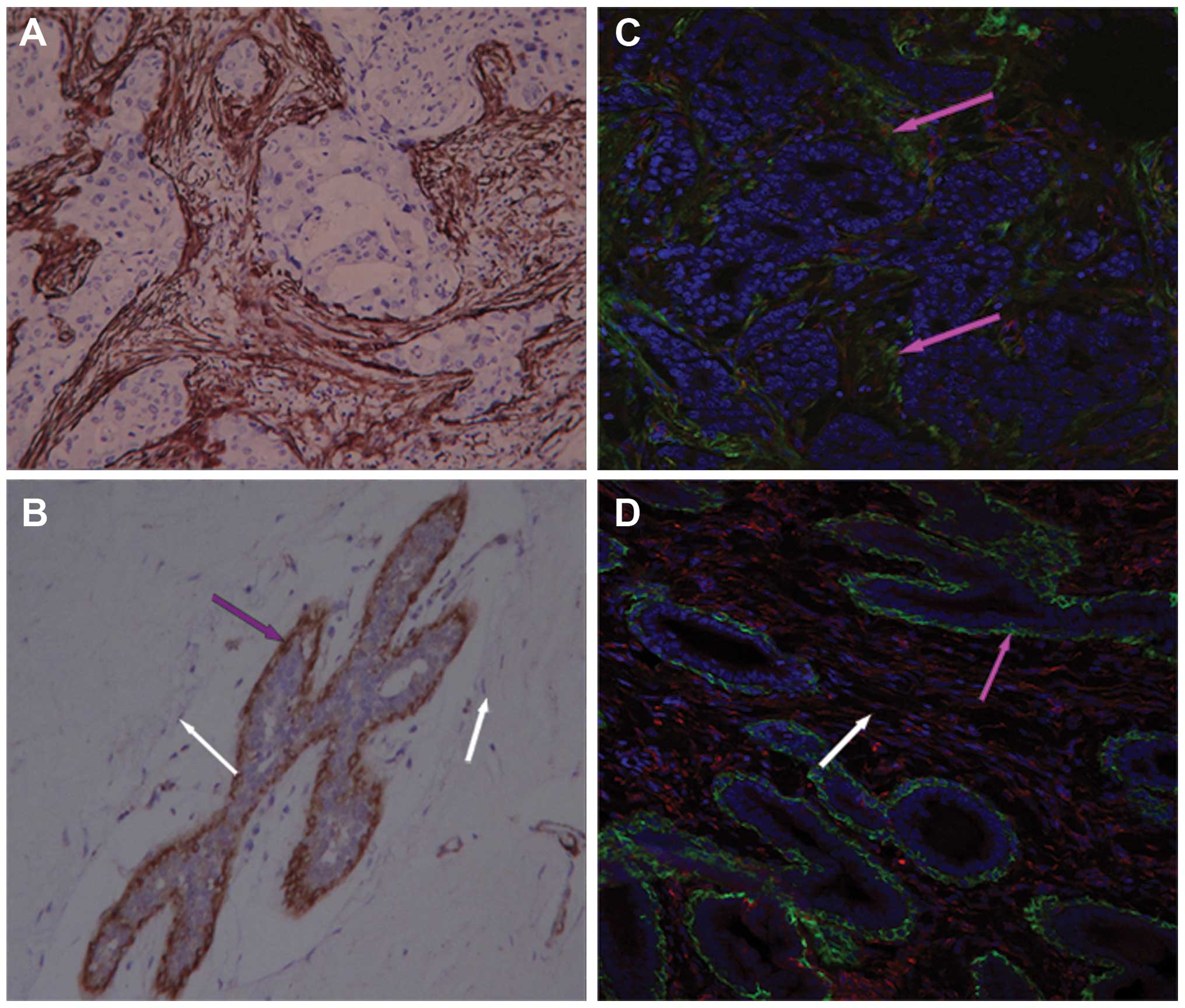 | Figure 1The difference of α-SMA expression in
invasive breast cancer, normal breast tissue and breast
fibroadenoma. Tissues detected by immunohistochemistry and
immunofluorescence showed that: α-SMA expression was widely
observed in the stroma of invasive breast cancer tissue [the brown
area in (A), DAB staining. (C) Green, α-SMA; red, vimentin, pink
arrows]. While, little α-SMA expression was observed in normal
breast/fibroadenoma stromal region (B, white arrows), and α-SMA was
only present in myoepithelium, vimentin was negative (B and D, pink
arrows, no red). (A, B, C and D), 200-fold. |
CAFs maintain an activated phenotype in
vitro
To investigate the characteristics of the primarily
cultured cells, we performed immunocytochemistry and
immunofluorescence using antibodies against α-SMA, the muscle cell
marker desmin, the epithelial cell marker cytokeratin (pan), the
mesenchymal cell marker vimentin, the fibroblast marker FSP-1 and
fibronectin (Fig. 2). The results
indicated that only CAFs were α-SMA-positive, while vimentin and
fibronectin could be detected in both CAFs and NFs. Although FSP-1
was expressed in both cell types, relatively few positive cells
were observed. Our results demonstrated that cytokeratin was not
expressed in either type of cells. It was notable that the CAFs
from one specimen showed weak expression of desmin which would be
expected to be absent in breast fibroblasts. The expression of
α-SMA was confirmed by western blot analysis (Fig. 2I). Furthermore, another activation
marker PDGFR-β was detected by western blot analysis (Fig. 2I) but the result showed no
difference between CAFs and NFs. These results demonstrated that
CAFs and NFs were kindred cell types with the nature of fibroblasts
after multiple passages in vitro, CAFs maintained an
activated phenotype.
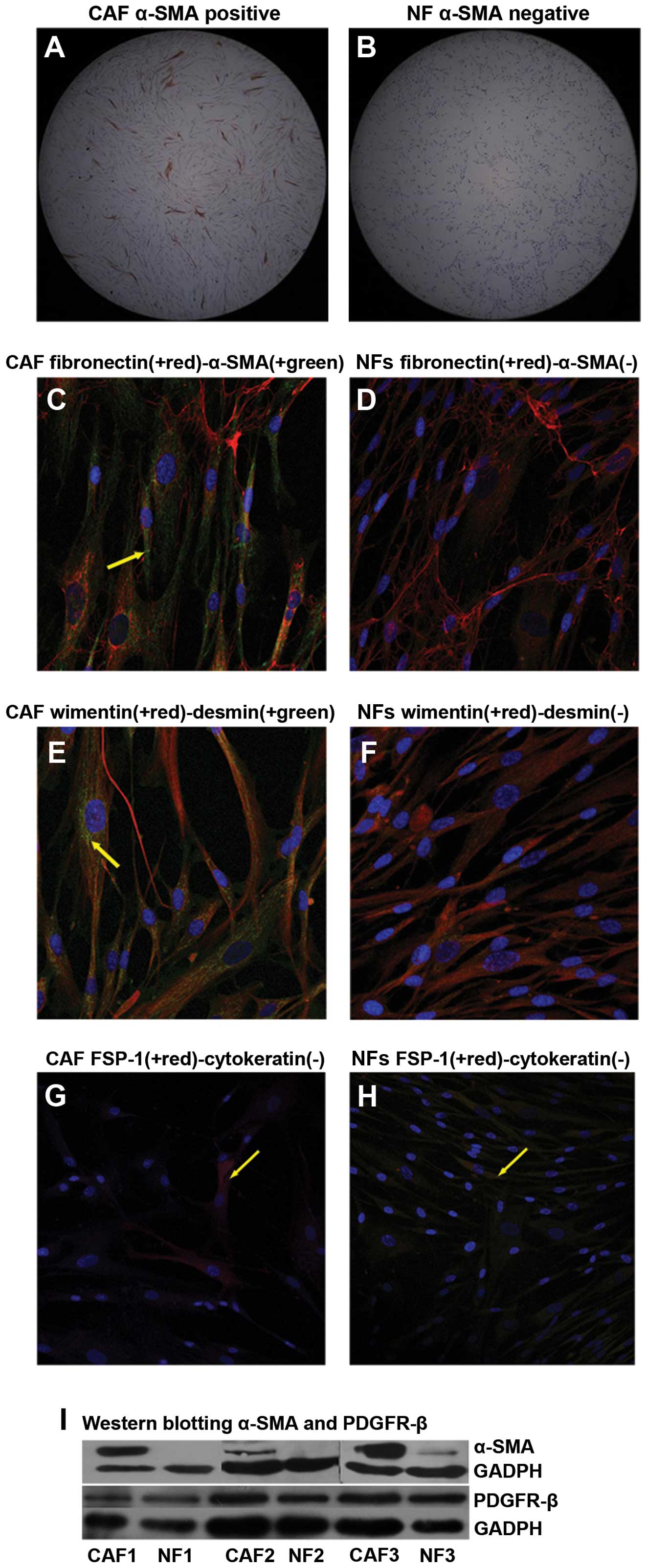 | Figure 2Characterization of CAFs and NFs
activation. 1) Immunocytochemistry: The cells were labeled by DAB,
fluorescent secondary antibodies (green, red) and DAPI (blue). a)
Immunocytochemistry showed CAFs were fractionally α-SMA-positive (A
and C), while NF completely lacked α-SMA expression (B and D). b)
CAFs and NFs both express vimentin (E and F). In (E), CAFs
demonstrated weak expression of desmin, which was absent in NF. c)
No expression of cytokeratin (pan) was observed in CAFs or NFs,
while the expression of FSP-1 was found only in a small proportion
of CAFs and NFs (G and H). (A and B), 50-fold; (C, D, E, F, G and
H), 200-fold. 2) Western blot analysis for α-SMA and PDGFR-β assay:
CAFs expressed α-SMA, while NF almost completely lacked it. But,
there was no expression difference on PDGFR-β between CAFs and NFs
(I). |
The proliferative capacity of CAFs and
NFs
The MTT assays were carried out to analyze the
proliferation capability of CAFs and NFs. The results were
normalized statistically (Fig.
3A), suggesting that the proliferation ability of CAFs were
either stronger than or equal to that of NFs (days 5, 6 and 7,
p<0.05 or p>0.05).
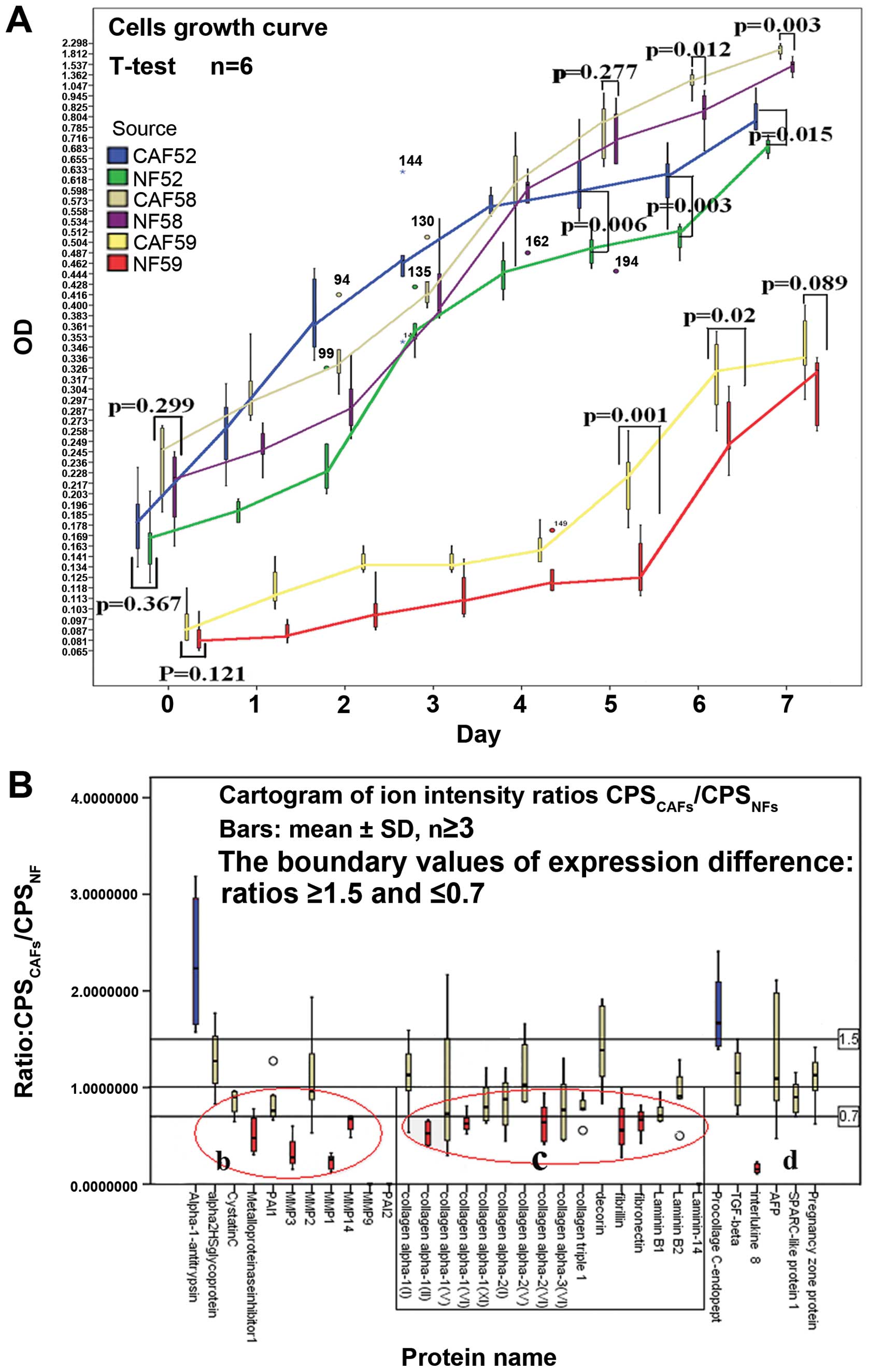 | Figure 3Cell growth curve and cartogram of
secretory protein expression from CAFs and NFs. 1) Chart A shows
the growth curve of CAFs and NFs. Three pairs of homologous CAFs
and NFs were evaluated. On days 5, 6 and 7, the data did not
provide evidence that the proliferation of CAFs in vitro was
weaker than that of NFs (p<0.05 or p>0.05). 2) Chart B is a
cartogram of peptide ion intensity ratios (CPSCAFs/ CPSNFs, mean ±
SD, n≥3). To delimit the expression difference, ratios ≥1.5 and
≤0.7 were specified as the difference boundary values of protein
expression, and ratio =1 was assigned as an equivalent reference
line. According to these rules, only two proteins in the selected
proteins had higher expression in CAFs (blue), while more proteins
showed lower expression (red). Chart B is divided into three parts:
part b, matrix-degrading enzymes and protease inhibitors; part c,
extracellular matrix components; and part d, other proteins. In
most of the ratios parts b and c are lower than 1. These proteins
included the EMC ingredient. |
The detection of secretory proteins from
CAFs and NFs
The 3 pairs of samples of secretory proteins
obtained from CM of CAFs and NFs were globally scanned by MALDI
TOF/TOF-MS. Results showed in total 2,903 proteins in CAFs and
3,023 proteins in NFs (data not shown). Among them, 2,811 proteins
were shared in these two samples. The extracellular proteins
included matrix metalloproteinases (MMP1, MMP2, MMP3, MMP9, MMP10,
MMP11 and MMP14), cathepsins, plasminogen activator inhibitors
(PAI), cystatins, collagens and other extracellular components.
Several membrane proteins were also identified, including cadherin,
integrin and growth factor-binding protein. Regarding collagens,
CAFs and NFs were found to have different expression profiles:
collagen α-1(III), collagen α-5(VI) were only discovered in NFs,
while collagen α-1(XII) and collagen α-2(V) appeared in CAFs. Four
of the six types were detected in CAFs.
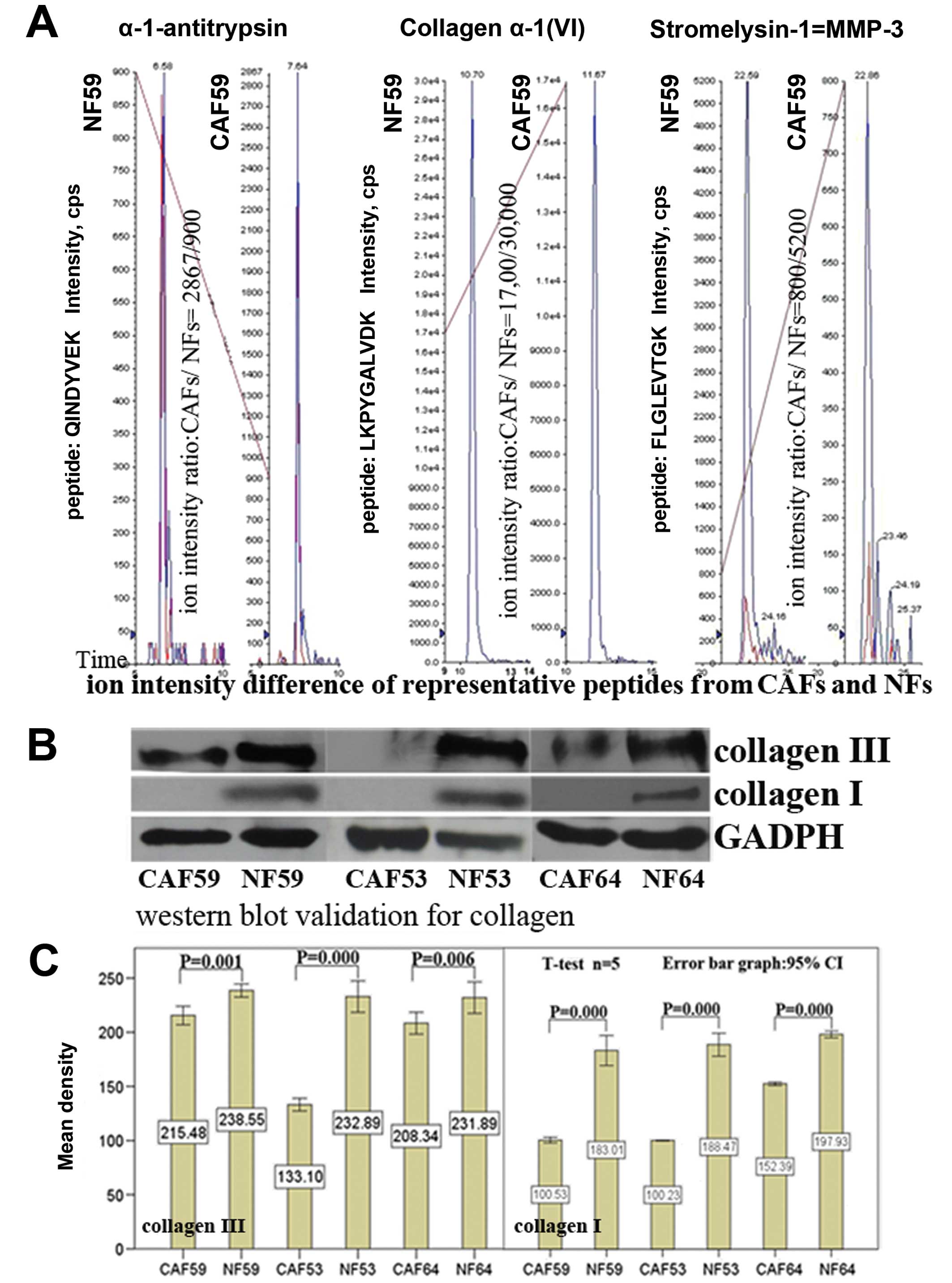
Label-free quantitative assay for
selected proteins by MRM
To quantitatively identify the differences between
secretory proteins generated by CAFs and NFs, we chose to profile
ECM-related proteins and other extracellular protein components, as
well as several cytokines (Table
I) by MRM, Applied Biosystems, and MDS Inc). The ion-intensity
values (CPS) of representative peptides from different proteins
were collected, and the CPS values of a same ion-peptide (in a same
batch testing) were calculated, ion-intensity ratio =
CPSCAFs/CPSNFs. A ratio of each protein was
then obtained from data from the same batch testing. To delimit the
expression difference, we specified ratios ≥1.5 and ≤0.7 as the
boundaries of protein expression, with ratio =1 as an equivalent
reference line. The expression differences of selected proteins are
shown in Fig. 3B.
 | Table ISelected extracellular proteins and
peptides for quantitative assay by mass spectrometry (4000 QTrap
LC/MS/MS, multiple-reaction monitoring model). |
Table I
Selected extracellular proteins and
peptides for quantitative assay by mass spectrometry (4000 QTrap
LC/MS/MS, multiple-reaction monitoring model).
| Protein ID | Protein name | Abbreviation | The sequence of
ideal peptides |
|---|
| P02452 | Collagen alpha-1(I)
chain | COL1A1 |
DGEAGAQGPPGPAGPAGER, ADDANVVR,
GPAGPQGPR |
| P02461 | Collagen
alpha-1(III) chain | COL3A1 | GPVGPSGPPGK |
| P20908 | Collagen alpha-1(V)
chain | COL5A1 | ENPGSWFSEEK |
| P12109 | Collagen
alpha-1(VI) chain | COL6A1 | IALVITDGR,
LKPYGALVDK, TAEYDVAYGESHLFR, VPSYQALLR |
| Q99715 | Collagen
alpha-1(XII) chain | COL12A1 | ALALGALQNIR,
VILTPMTAGSR, NSDVEIFAVGVK |
| P08123 | Collagen alpha-2(I)
chain | COL1A2 | GEAGAAGPAGPAGPR,
TGEVGAVGPPGFAGEK, TGHPGTVGPAGIR |
| P12110 | Collagen
alpha-2(VI) chain | COL6A2 | DYDSLAQPGFFDR,
LFAVAPNQNLK |
| P12111 | Collagen
alpha-3(VI) chain | COL6A3 | QINVGNALEYVSR,
QLGTVQQVISER, VGLEHLR |
| P05997 | Collagen alpha-2(V)
chain | COL5A2 | SLSSQIETMR |
| Q16363 | Laminin subunit
alpha-14 | LAMA14 |
AIEHAYQYGGTANSR |
| P11047 | Laminin subunit
gamma-1 | LAMC1 | LSAEDLVLEGAGLR,
LVGGPMDASVEEEGVR, NTIEETGNLAEQAR |
| P07942 | Laminin subunit
beta-1 | LAMB1 | IPSWTGAGFVR |
| P35555 | Fibrillin-1 | FBN1 | YLIESGNEDGFFK |
| P02751 | Fibronectin | FN1 | NLQPASEYTVSLVAIK,
NTFAEVTGLSPGVTYYFK, TYHVGEQWQK, VGDTYERPK |
| P14780 | Matrix
metalloproteinase-9 | MMP9 |
LGLGADVAQVTGALR |
| P03956 | Interstitial
collagenase | MMP1 | SQNPVQPIGPQTPK,
WEQTHLTYR, VTGKPDAETLK, DGFFYFFHGTR |
| Q14515 | SPARC-like protein
1 | SPARCL1 | LLAGDHPIDLLLR,
MRDWLK |
| P08254 | Stromelysin-1 | MMP3 | FLGLEVIGK |
| P50281 | Matrix
metalloproteinase-14 | MMP14 | SPQSLSAATAAMQK,
AVDSEYPK |
| Q96CG8 | Collagen triple
helix repeat-containing protein 1 | CTHRC1 | ESFEESWTPNYK |
| P20742 | Pregnancy zone
protein | PZP | ATVLNYLPK,
GPTQDFR |
| P01033 | Metalloproteinase
inhibitor 1 | TIMP1 | GFQALGDAADIR,
SEEFLIAGK |
| P01034 | Cystatin-C | CST3 | ALDFAVGEYNK,
LVGGPMDASVEEEGVR |
| P05121 | Plasminogen
activator inhibitor 1 | PAI-1 | QVDFSEVER,
TPFPDSSTHR |
| P05120 | Plasminogen
activator inhibitor 2 | PAI-2 | TPVQMMYLR |
| Q15113 | Procollagen
C-endopeptidase enhancer 1 | PCOLCE | GFLLWYSGR |
| P07585 | Decorin | DCN | VSPGAFTPLVK,
AHENEITK, DLPPDTTLLDLQNNK |
| P01009 |
Alpha-1-antitrypsin | SERPINA1 | QINDYVE |
| P10145 | Interleukin 8 | | TYSKPFHPK |
| P61812 | Transforming growth
factor-β | TGF-β | EGVYTVFAPTNEAFR,
ILGDPEALR, SPYQLVLQHSR |
| P08253 | 72 kDa type IV
collagenase | MMP2 | AFQVWSDVTPLR,
IIGYTPDLDPETVDDAFAR |
| P02771 |
Alpha-fetoprotein | AFP | YIQESQALAK |
Our data showed α-1-antitrypsin expression was
upregulated, while metalloproteinase inhibitor 1 expression was
downregulated in CAFs. No differences in protein expression were
found for other proteinase inhibitors between CAFs and NFs. When
metalloproteinases were tested, MMP1, MMP3 and MMP14 were found to
be downregulated in CAFs compared with NFs. As for the ECM
components, the data demonstrated that the expression levels of
collagen α-1(II), collagen α-1(VI), collagen α-2(VI), fibrillin and
fibronectin were all lower in CAFs than in NFs; only procollagen
C-endopeptidase enhancer 1 was highly expressed in CAFs. When ratio
= 1 was set as an equivalent reference line for the analysis, we
found a broad decrease in expression of secretory proteins in
CAFs.
Western blot analysis validation of
collagen expression
In order to verify our results, we carried out
western blotting to verify the collagen expression beyond the
threshold (≥1.5 or ≤0.7). The results showed that there was a
significant difference in expression of collagens between CAFs and
NFs (Fig. 4). These results
suggest that the setting of the boundary values was rational, and
strengthened the reliability of data from our label-free
quantitative assay. These findings also provide further evidence to
prove the decreased expression of ECM components in CAFs.
Reduction of collagen deposition in
breast cancer stroma
To further validate the collagen reduction found in
the label-free quantitative assay, we performed HE and masson
trichrome staining to assess the collagen density in breast cancer
tissue, normal tissue and breast fibroadenoma. The HE staining
sections suggested a significantly reduction of ECM amount in
cancer stroma or cancer reactive stroma (Fig. 5A–C). Masson trichrome staining
exhibited more obvious differences in collagen deposition. In order
to quantify the results, the density of the blue color in masson
trichrome staining was analyzed by ImagePro Plus6.0. Statistical
analysis showed that there were significant differences in collagen
deposition among cancer stroma, paracarcinoma, normal tissue and
fibroadenoma, whereby cancer stroma showed the least collagen
deposition (p=0.000) compared with the other three kinds of stroma.
This assay and analysis further confirmed the collagens reduction
observed in the label-free quantitative assay. As collagens are the
most abundant proteins of the ECM and offer structural support for
resident cells, the decrease of collagen deposition implies a
reduction of ECM in cancer stroma.
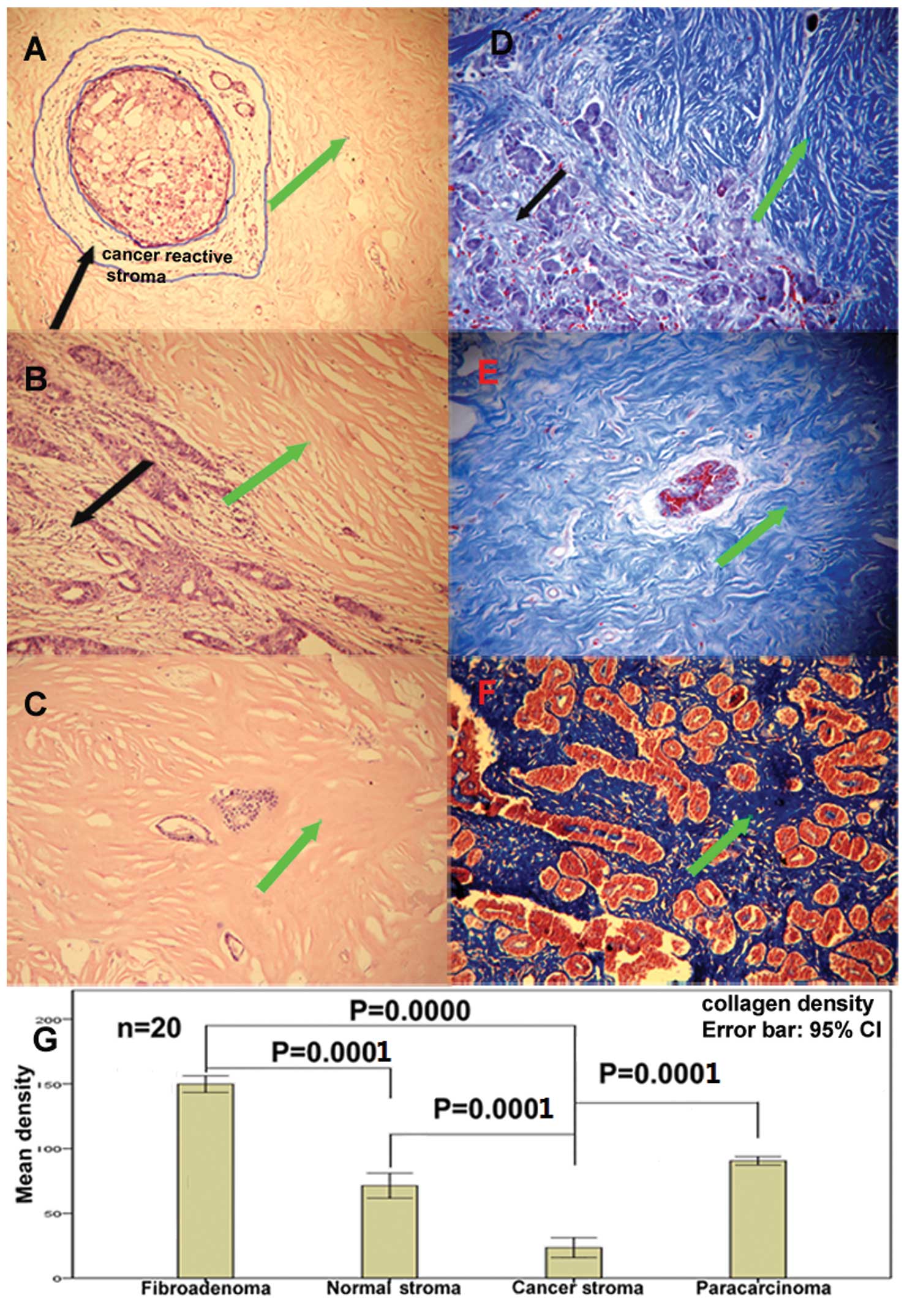 | Figure 5The deposition of ECM and collagen in
tissue. 1) (A and B) H&E staining showing that ECM is
significantly reduced in cancer stroma or cancer reactive stroma,
with or without increased numbers of stromal cells. (C) H&E
staining of normal breast tissue demonstrating a rich ECM and lack
of stromal cells (green arrow). 2) (D, E and F) Masson trichrome
staining depicting collagen in blue. In (D), the cancer stroma
(black arrow) is lighter in blue color compared with paracarcinoma
stroma, normal breast stroma (F) and fobroadenoma stroma (green
arrow). The tissue of breast fibroadenoma showed the highest
density of collagen deposition. 3) (G) Results of statistical
analysis for collagen density (the blue area shown by masson
trichrome staining. Four areas: cancer stroma, paracancer stroma,
normal tissue stroma and fibroadenoma, in D, E and F). Cancer
stroma showed the least collagen deposition comparing with the
stroma from paracarcinoma, normal tissue and fibroadenoma (error
bars represent mean ± SEM, n=20). (A, B and C) 170-fold; (D, E and
F) 130-fold. |
Discussion
Usually, the major function of fibroblasts is
maintaining the structural framework of tissues by continuously
secreting ECM components (28).
Under normal conditions, fibroblasts stay inactivated by
constitutively expressing vimentin and other markers (29). Once activated (by some pathological
factors, for example, wound healing or fibrosis), they can
immediately take on enhanced proliferative and ECM generating
capability and become markers-positive, for example, α-SMA and
PGDFR-β. The series of responses finally leads to wound healing
progression such as wound contraction, angiogenesis and stimulation
of epithelial cell proliferation (30). It has been reported that CAFs in
tumor microenvironments also acquire a similar activated phenotype,
and function in a co-evolutionary manner with cancer cells
(31,32).
In the present study, we determined the expression
of α-SMA, which is a major component of the contractile apparatus,
as a marker of fibroblast activation. While there are other
activation markers that can be used to reflect the fibroblast
activation state, α-SMA was chosen because it is the most commonly
used. Immunohistochemistry demonstrated that CAFs were the only
cellular components that expressed α-SMA in the stroma of invasive
breast cancer, but not in normal and benign tissues (Fig. 1). In agreement, Sappino et
al have found that up to 80% of breast cancers include α-SMA
positive CAFs (33) while evidence
from other studies has shown absence in breast fibroadenoma
(34). Therefore, our data
confirmed that the CAFs found in cancer stroma are in an activated
state. We also found that after undergoing multiple passages
(within 8 passages), primary cultured CAFs were still able to
maintain the expression of activation marker α-SMA (Fig. 2), suggesting that CAFs were able to
maintain the activated state in vitro as well as in
vivo.
Some researchers have indicated that activated CAFs
with PDGFR-β overexpression are responsible for the expansion of
tumor stroma and the excessive deposition of ECM (19,22).
The increased deposition of ECM in cancer is called desmoplasia,
which is a process similar to that in organ fibrosis. Usually, the
desmoplastic stroma contains increased amounts of fibrillar
collagens and other ECM components (35,36).
However, as ECM deposition in breast cancer tissues presents
different pathological types (23), further study is required to
disclose the true relationship between activated CAFs and ECM
deposition. In this study, proliferation assays of CAFs and NFs did
not provide evidence that the proliferation of CAFs in vitro
was weaker than that of NFs. While our western blot analysis
experiment showed no PDGFR-β expression difference between CAFs and
NFs. Based on the activation state of CAFs in vitro, we then
examined secretory proteins from CAFs and NFs using MS. Many
extracellular matrix components were detected, such as collagens,
fibronectin, decorin and several proteases involved in ECM
degradation (such as MMPs, cathepsin and PAI) were identified (data
not shown). Besides the differences in the expression profile of
secretory proteins between CAFs and NFs, the results of a
labeled-free quantitative assay (MRM) for intresting proteins
further demonstrated that the expression of ECM-related proteins
and proteases from CAFs diminished compared to that of NFs; this
was confirmed by western blot analysis (Fig. 4). Therefore, our experiments
suggested that activated CAFs might be attenuated in their ability
to produce ECM components, and biological characteristics of CAFs
from invasive breast cancer are different from those of fibroblasts
in normal breast tissue. This is in complete contradiction with the
theoretical anticipation that activated CAFs would secrete a large
amount of ECM components, more than NFs.
Tissue staining studies were carried out to further
clarify the relationship between CAFs and ECM deposition in
situ. In our study, the decline of ECM in malignant tissue was
observed by HE staining. Comparing to that of para-neoplastic,
normal breast and fibroadenoma stroma, Masson trichrome staining
further demonstrated that the collagen within tumor stroma was
significantly reduced. Additionally, fibroadenoma stroma was shown
to have the most abundant collagen in our experiments.
While fibroblasts with activated traits are expected
to produce more ECM components, evidence in our study demonstrated
that normal breast stroma and fibroadenoma, rather than invasive
breast cancer stroma had more extensive collagen deposition. The
fibroblasts in normal breast stroma and fibroadenoma, however, are
not α-SMA-positive. The reduction of collagens within the tumor
stroma might be possibly caused by an increase in the expression of
matrix-degrading enzymes from tumor cells and tumor stromal
fibroblasts, or by the decreased secretion of ECM components from
CAFs. However, in this study, there was no evidence to suggest that
there was an increase in the expression of MMPs and other proteases
from CAFs (Fig. 3B). Our study
therefore proposes that the reduction of collagen secretion in CAFs
was the major cause of ECM reduction in tumor stroma.
In summary, although CAFs from invasive breast
cancer obtained the activated phenotype, their capacity of
producing ECM components was significantly impaired compared with
normal fibroblasts and fibroadenoma fibroblasts. In breast cancer,
CAFs might have remodeled the stromal structure and tumor
microenvironment through changes in their biological
characteristics and the profile of secretory proteins.
Acknowledgements
This study was supported by a grant from the
National Natural Science Foundation of China (nos. 83172210 and
30973382). We thank Dr Shanwei Wang and Dr Jiaping Peng for their
technical support in pathology. We also thank Dr Jiekai Yu, Dr
Jiawei Zhang and Dr Weiting Ge for their help in Mass Spectrometry
and Laser Confocal technology.
Abbreviations:
|
CAFs
|
cancer-associated fibroblasts
|
|
NFs
|
normal bread fibroblasts
|
|
ECM
|
extracellular matrix
|
|
MAL-DI TOF/TOF-MS
|
matrix-assisted laser
desorption/ionization time of flight mass spectrometry
|
|
MRM
|
multiple reaction monitoring
|
|
CPS
|
counts per second
|
|
SMA
|
smooth muscle actin
|
|
PDGFR
|
platelet derived growth factor
receptor
|
References
|
1
|
Zeisberg M, Strutz F and Muller GA: Role
of fibroblast activation in inducing interstitial fibrosis. J
Nephrol. 13(Suppl 3): S111–S120. 2000.PubMed/NCBI
|
|
2
|
Darby IA and Hewitson TD: Fibroblast
differentiation in wound healing and fibrosis. Int Rev Cytol.
257:143–179. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ryan GB, Cliff WJ, Gabbiani G, Irle C,
Statkov PR and Majno G: Myofibroblasts in an avascular fibrous
tissue. Lab Invest. 29:197–206. 1973.PubMed/NCBI
|
|
4
|
Ryan GB, Cliff WJ, Gabbiani G, et al:
Myofibroblasts in human granulation tissue. Hum Pathol. 5:55–67.
1974. View Article : Google Scholar
|
|
5
|
Watts GT: Myofibroblasts. Lancet.
1:3351979. View Article : Google Scholar
|
|
6
|
Ratajska A and Gawlik Z: Morphological and
physiological characteristics of myofibroblasts. Patol Pol.
33:1–19. 1982.(In Polish).
|
|
7
|
Lorusso G and Ruegg C: The tumor
microenvironment and its contribution to tumor evolution toward
metastasis. Histochem Cell Biol. 130:1091–1103. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tyan SW, Kuo WH, Huang CK, et al: Breast
cancer cells induce cancer-associated fibroblasts to secrete
hepatocyte growth factor to enhance breast tumorigenesis. PLoS One.
6:e153132011. View Article : Google Scholar
|
|
9
|
Pietras K and Ostman A: Hallmarks of
cancer: interactions with the tumor stroma. Exp Cell Res.
316:1324–1331. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bissell MJ and Hines WC: Why don’t we get
more cancer? A proposed role of the microenvironment in restraining
cancer progression. Nat Med. 17:320–329. 2011.
|
|
11
|
Dvorak HF: Tumors: wounds that do not
heal. Similarities between tumor stroma generation and wound
healing. N Engl J Med. 315:1650–1659. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Vaheri A, Enzerink A, Rasanen K and
Salmenpera P: Nemosis, a novel way of fibroblast activation, in
inflammation and cancer. Exp Cell Res. 315:1633–1638. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rasanen K and Vaheri A: Activation of
fibroblasts in cancer stroma. Exp Cell Res. 316:2713–2722. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gialeli C, Nikitovic D, Kletsas D,
Theocharis AD, Tzanakakis GN and Karamanos NK: PDGF/PDGFR signaling
and targeting in cancer growth and progression: Focus on tumor
microenvironment and cancer-associated fibroblasts. Curr Pharm Des.
20:2843–2848. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bedekovics J, Kiss A, Beke L, Karolyi K
and Mehes G: Platelet derived growth factor receptor-beta
(PDGFRbeta) expression is limited to activated stromal cells in the
bone marrow and shows a strong correlation with the grade of
myelofibrosis. Virchows Arch. 463:57–65. 2013. View Article : Google Scholar
|
|
16
|
Jacob M, Chang L and Pure E: Fibroblast
activation protein in remodeling tissues. Curr Mol Med.
12:1220–1243. 2012. View Article : Google Scholar
|
|
17
|
Liao D, Luo Y, Markowitz D, Xiang R and
Reisfeld RA: Cancer associated fibroblasts promote tumor growth and
metastasis by modulating the tumor immune microenvironment in a 4T1
murine breast cancer model. PLoS One. 4:e79652009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Eyden B: The myofibroblast: an assessment
of controversial issues and a definition useful in diagnosis and
research. Ultrastruct Pathol. 25:39–50. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Conti JA, Kendall TJ, Bateman A, et al:
The desmoplastic reaction surrounding hepatic colorectal
adenocarcinoma metastases aids tumor growth and survival via
alphav integrin ligation. Clin Cancer Res. 14:6405–6413.
2008. View Article : Google Scholar
|
|
20
|
Schedin P and Keely PJ: Mammary gland ECM
remodeling, stiffness, and mechanosignaling in normal development
and tumor progression. Cold Spring Harb Perspect Biol.
3:a0032282011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Barsky SH and Gopalakrishna R: An
experimental model for studying the desmoplastic response to tumor
invasion. Cancer Lett. 35:271–279. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pandol S, Edderkaoui M, Gukovsky I, Lugea
A and Gukovskaya A: Desmoplasia of pancreatic ductal
adenocarcinoma. Clin Gastroenterol Hepatol. 7:S44–S47. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Walker RA: The complexities of breast
cancer desmoplasia. Breast Cancer Res. 3:143–145. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Katikireddy KR and O’Sullivan F:
Immunohistochemical and immunofluorescence procedures for protein
analysis. Methods Mol Biol. 784:155–167. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lai TC, Chou HC, Chen YW, et al:
Secretomic and proteomic analysis of potential breast cancer
markers by two-dimensional differential gel electrophoresis. J
Proteome Res. 9:1302–1322. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bluemlein K and Ralser M: Monitoring
protein expression in whole-cell extracts by targeted label- and
standard-free LC-MS/MS. Nat Protoc. 6:859–869. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gudjonsson T, Adriance MC, Sternlicht MD,
Petersen OW and Bissell MJ: Myoepithelial cells: their origin and
function in breast morphogenesis and neoplasia. J Mammary Gland
Biol Neoplasia. 10:261–272. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Grinnell F: Fibroblasts, myofibroblasts,
and wound contraction. J Cell Biol. 124:401–404. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
McAnulty RJ: Fibroblasts and
myofibroblasts: their source, function and role in disease. Int J
Biochem Cell Biol. 39:666–671. 2007.PubMed/NCBI
|
|
30
|
Tettamanti G, Grimaldi A, Rinaldi L, et
al: The multifunctional role of fibroblasts during wound healing in
Hirudo medicinalis (Annelida, Hirudinea). Biol Cell.
96:443–455. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Giannoni E, Bianchini F, Masieri L, et al:
Reciprocal activation of prostate cancer cells and
cancer-associated fibroblasts stimulates epithelial-mesenchymal
transition and cancer stemness. Cancer Res. 70:6945–6956. 2010.
View Article : Google Scholar
|
|
32
|
Brentnall TA, Lai LA, Coleman J, Bronner
MP, Pan S and Chen R: Arousal of cancer-associated stroma:
overexpression of palladin activates fibroblasts to promote tumor
invasion. PLoS One. 7:e302192012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sappino AP, Skalli O, Jackson B, Schurch W
and Gabbiani G: Smooth-muscle differentiation in stromal cells of
malignant and non-malignant breast tissues. Int J Cancer.
41:707–712. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Heneghan HM, Martin ST, Casey M, Tobbia I,
Benani F and Barry KM: A diagnostic dilemma in breast pathology -
benign fibroadenoma with multinucleated stromal giant cells. Diagn
Pathol. 3:332008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shao ZM, Nguyen M and Barsky SH: Human
breast carcinoma desmoplasia is PDGF initiated. Oncogene.
19:4337–4345. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Iacobuzio-Donahue CA, Argani P, Hempen PM,
Jones J and Kern SE: The desmoplastic response to infiltrating
breast carcinoma: gene expression at the site of primary invasion
and implications for comparisons between tumor types. Cancer Res.
62:5351–5357. 2002.PubMed/NCBI
|