Introduction
Hepatocellular carcinoma (HCC) is one of the most
aggressive neoplasms, and is responsible for a third of the
estimated cancer-caused deaths worldwide (1). Although many new chemotherapeutic
agents and treatment modalities are now being used clinically, the
survival rate has not improved during the past two decades.
Therefore, a deeper understanding of the mechanisms controlling HCC
progression is of primary importance to develop new approaches for
treatment (2,3).
Apurinic apyrimidinic endonuclease/redox effector
factor 1 (APE/Ref-1) is a master regulator of cellular response to
oxidative stress conditions with diverse function through
transactivating numerous seminal transcription factors involved in
cell proliferation, apoptosis, and metastasis, such as AP-1 and
nuclear factor-κB (NF-κB) (4).
Therefore, the multifunctional activities of APE/Ref-1 make it a
prime candidate for gene/protein therapy in clinical settings.
Furthermore, emerging evidence indicates that APE/Ref-1 has been
uncovered elevated in various types of cancer, and its subcellular
distribution is closely correlated with tumor aggressiveness,
resistance to radiotherapy, and poor outcome (5–11).
However, attempts to achieve an APE/Ref-1−/− human cell
line or APE/Ref-1 knockout mice were embryonically lethal,
suggesting that development of an APE/Ref-1 specific inhibitor
instead would have significant therapeutic potential. In our study,
APE/Ref-1 is not only unregulated in HCC cells but also in liver
cancer tissue microarray; however, the functional role of APE/Ref-1
in tumor pathogenesis and progression is not yet clear.
In particular, it is well known that the liver is
the main organ for metabolism of copper (Cu), which has long been
considered to have carcinogenic potential (12). In just one example, Cu accumulation
in LEC rat liver leads to spontaneous HCC (13). Conversely, Cu-depleted animals
developed small, relatively avascular tumors with reduced invasive
capability (14). In vitro
studies confirmed that people with higher Cu levels are more
susceptible to cancer-related mortality (15), consistent with the finding that
both serum and tumor Cu levels are elevated in a variety of
malignancies, including HCC (16–18).
Additionally, a number of clinical trials with Cu chelators have
been conducted, and the results are promising (14,19,20).
Making use of this information, the present study was aimed at
examining the functional role of APE/Ref-1 in HCC with or without
the aid of Cu. Our data elucidate further the strong association
among Cu, APE/Ref-1 and HCC, and also reinforce the hypothesis that
APE1/Ref-1 is of great significance to facilitate HCC
progression.
Materials and methods
Cell culture
Human HCC cell line (Hep3B) and immortalized
non-malignant hepatocyte cell line (THLE3) were purchased from the
American Type Culture Collection (ATCC, Manassas, VA, USA) and
cultured as recommended by ATCC. The passage number for Hep3B used
in these experiments was ≤8, and for THLE3 was 3–5. Cells were
treated at 70% confluence with fresh medium and reagents added
simultaneously.
DNA and reporter constructs
The p3xFlag-Ref-1 is a gift from Dr Kaikobad Irani
(University of Pittsburgh Medical Center, Pittsburgh, PA, USA). The
Ref-1 promoter reporter plasmid was constructed by ligating the
pGL3-basic vector (digested with NheI and HindIII)
and an AvrII-HindIII fragment from Ref-1 promoter
plasmid pCB2 (21). The resulting
plasmid is termed pRef-1-Luc.
Reagents and antibodies
The following primary antibodies were used for
western blot analysis and immunohistochemistry: APE/Ref-1 antibody
was from Novus Biologicals (Littleton, CO, USA); AP-1/c-Fos, MMP-1,
Bcl-2 and Snail antibodies were from Santa Cruz Biotechnology
(Santa Cruz, CA, USA); p84 antibody was from GeneTex Inc. (Irvine,
CA, USA); α-tubulin antibody was from Sigma Life Sciences (St.
Louis, MO, USA). The following secondary antibodies were used:
horseradish peroxidase (HRP)-conjugated anti-mouse antibody and
anti-rabbit antibody were from Santa Cruz Biotechnology.
Cell protein extraction and western blot
analysis
Following different treatments, cells were collected
and lysed as described previously (22). By using a detergent-compatible
protein assay kit (Bio-Rad, Hercules, CA, USA), the protein
concentration was precisely measured three times. Equal amounts of
the soluble protein were subjected to SDS-PAGE and transferred to
nitrocellulose membrane (Sigma Life Sciences). Specific protein was
detected by antibody followed by the chemiluminescence detection
reagent (Bio-Rad). Measurement of signal intensity on X-ray film
was performed using an Imaging densitometer with Multi-Analyst
software (Bio-Rad). For statistical analysis, all data were
expressed as fold changes of the control based on the calculation
as the density values of the specific protein band/α-tubulin or p84
density values. All figures showing quantitative analysis include
data from at least three independent experiments.
Immunohistochemistry (IHC) and
scoring
Unstained human tissue microarrays (TMA) containing
70 cases of HCC pathologically confirmed and 10 normal tissue
samples were purchased from US Biomax Inc. (Rockville, MD, USA).
Antigen retrieval was performed by treatment with xylene and graded
alcohols. After quenching the activity of endogenous peroxidase and
blocking with 5% serum, TMA were first incubated with APE/Ref-1 as
the primary antibody and then secondary antibody. Envision and DAB
kits (Santa Cruz Biotechnology) were used for visualization and
assessments. All IHC staining was carried out in duplicate. Nuclear
and cytoplasmic staining were quantified and recorded. Three
experienced postdoctoral observers scored in a blinded manner using
a scoring system to measure staining (0, none; 1, faint; 2,
moderate; 3, strong).
Measurement of cytotoxicity
The cytotoxicity of Cu and DSF against THLE3 and
Hep3B cells was measured using the CellTiter-Glo luminescent cell
viability assay (Promega, Madison, WI, USA), based on ATP
bioluminescence as a marker of cell viability. After being exposed
to the reagents for different time periods, the luminescence
produced by the luciferase-catalyzed luciferin and ATP reaction was
detected using a Modulus™ Microplate Luminometer (Turner BioSystems
Inc., Sunnyvale, CA, USA) according to the instructions. Six
independent experiments were performed for each group and data were
normalized to the control.
Dual-luciferase reporter assay
APE/Ref-1 reporter construct was transfected into
THLE3 hepatocytes in triplicate, and luciferase activity was
analyzed after 48 h using the Dual Luciferase Reporter Assay System
(Promega) in a Modulus™ Microplate Luminometer (Turner BioSystems
Inc.). A Renilla luciferase plasmid (Promega) was also
co-transfected as an internal control for transfection efficiency.
The mean luciferase activities, normalized by cell transfection
efficiencies, were calculated relative to the activities of control
cells.
Quantitative real-time reverse
transcription (RT)-polymerase chain reaction (PCR)
Total RNA was isolated by using TRIzol according to
the manufacturer’s protocol (Invitrogen Inc., Carlsbad, CA, USA).
RNA concentration was determined by using a spectrophotometer
(Thermo Fisher Scientific Inc., Waltham, MA, USA). Equal amounts of
mRNA were amplified using First-strand cDNA Synthesis for
Quantitative RT-PCR kit (Marligen Biosciences Inc., Ijamsville, MD,
USA). Real-time-PCR was performed with iQ SYBR Green Supermix and
MYIQ5 detection system (Bio-Rad) using primer pairs: Snail, 5′-ATC
CGA AGC CAC ACA CTG-3′ (forward) and 5′-CAC TGG TAC TTC TTG ACA TCT
G-3′ (reverse); E-cadherin: 5′-GAG GAG AGC GGT GGT CAA AG-3′
(forward) and 5′-GTT CAG GGA GCT CAG ACT AG-3′ (reverse);
α-tubulin, 5′-GCG TGA TGG TGG GCA TGG GTC AG-3′ (forward) and
5′-AGG GGG GCC TCG GTC AGC AGC AC-3′ (reverse). Real-time PCR data
were analyzed using the comparative Ct method. The
2−ΔΔCt showed the difference between the threshold
cycles of the target and an internal reference (α-tubulin).
Transient transfection studies
Small interfering RNA (siRNA) duplexes directed
against APE/Ref-1 were purchased from Invitrogen Inc. Sequences of
APE/Ref-1 siRNA were as follows: sense: 5′-GUC UGG UAC GAC UGG AGU
ACC-3′, antisense: 5′-UAC UCC AGU CGU ACC AGA CCU-3′. Cells in
6-well plates were transfected at 50% confluence for siRNA and at
90% confluence for DNA plasmid using Lipofectamine 2000 (Invitrogen
Inc.) according to the manufacturer’s instructions. The final
concentration was 60 nM (siRNA) or 4 μg per well (plasmid). The
serum-free medium was replaced with regular medium 6 h later.
Cell proliferation assay
Proliferative capacity was measured using a
CellTiter AQ cell proliferation assay kit (Promega). The absorbance
of the formazan product at 490 nm was recorded in a Microplate
reader (Bio-Rad) and cell proliferation index was calculated in
comparison to the control cells. Six independent experiments were
performed for each group.
Invasion assay
The invasiveness was assessed by measuring the
number of cells migrating through a Matrigel-coated membrane with
pore size 8.0 μm (BD Biosciences, San Jose, CA, USA). According to
the manufacturer’s instructions, two groups of cells (control and
transfected cells) were collected and reconstituted in serum-free
medium (SFM) at a final concentration of 2×105 cells/ml.
Medium (750 μl) (10% serum included) was added to each well and 500
μl of prepared cells was added to the upper Matrigel-coated insert
(BD Biosciences). After incubation for 24 h, cells were fixed with
ice-cold methanol and stained with hematoxylin. Subsequently,
membranes were cut off, mounted, and visualized microscopically
(Olympus Optical Co. Ltd., Shinjuku-ku, Tokyo, Japan). The invading
cells on each of triplicate membranes were counted and averaged for
10 randomized fields at ×400 magnification.
Cell apoptosis assay
Following a 48-h transfection with APE/Ref-1 siRNA,
Hep3B cells were trypsinized and washed once with 1X PBS, fixed in
cold 70% ethanol until use. Cells were incubated in propidium
iodide (PI) staining solution in dark for 30 min: 50 μg/ml PI, 0.1%
sodium citrate, 50 μg/ml RNase A, 0.03% NP-40 in 1X PBS. Ten
thousand total events were counted for each sample which was
analyzed for apoptosis by Annexin V apoptosis detection kit
according to the manufacturer’s protocol (BD Biosciences).
Statistical analysis
Data are presented in bar plot as mean ± SD for at
least three independent experiments. Correlation between APE/Ref-1
immunostaining and tumor grade or stage was evaluated. Statistical
analyses (two-sample t-test and Pearson’s correlation) were carried
out using R-program. All p-values were two-sided, and tests with
p<0.05 were considered statistically significant.
Results
APE/Ref-1 is upregulated in HCC
cells
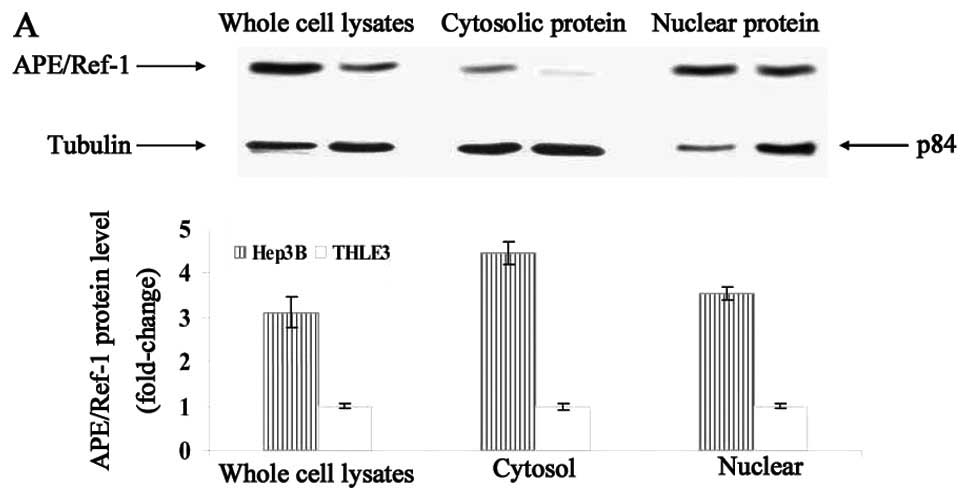
Expression of APE/Ref-1 was analyzed using western
blotting. As shown in Fig. 1A, HCC
cells exhibited a much higher level compared to hepatocytes
(3.1-fold increase in whole cell lysates; 4.5-fold increase in
cytoplasmic extracts; 3.5-fold increase in nuclear extracts).
Furthermore, increased APE/Ref-1 level was accompanied by enhanced
expression of AP-1/c-Fos, Bcl-2 and MMP-1 (Fig. 1B).
Correlation between different patterns of
APE/Ref-1 expression and clinicopathologic features
TMA was subjected to IHC analysis to study the
relationship between APE/Ref-1 expression and clinic features.
Accumulation of nuclear and cytoplasmic APE/Ref-1 was significantly
enhanced in HCC (p<0.00001, Fig. 2A
and B). The average nuclear immunostaining score of APE/Ref-1
in HCC was almost 5.5-fold of normal liver tissue (NLS), and
6.3-fold for cytoplasmic staining. APE/Ref-1 reactivity was also
compared in cases with different grade and stage. As shown in
Fig. 2C and D, cytoplasmic
staining was much higher in poorly differentiated tumors
(p<0.012) and in more aggressive tumors (p<0.001); whereas no
evident difference was observed in the nuclear reactivity of
APE/Ref-1. These findings suggested that cytoplasmic APE/Ref-1
accumulation is closely correlated with the clinical outcome of HCC
patients.
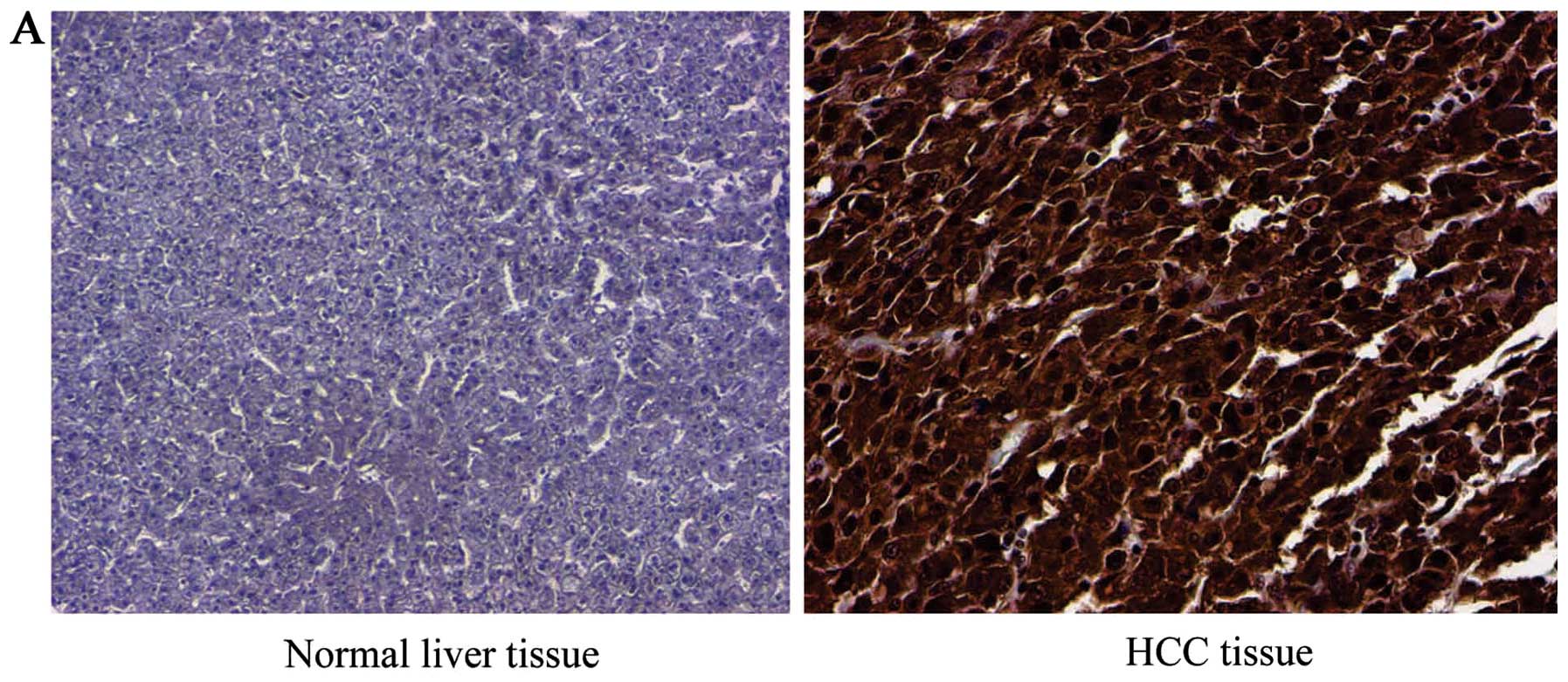 | Figure 2Correlation between different patterns
of APE/Ref-1 expression and clinicopathologic features. Human
tissue microarray was used for IHC analysis. Envision and DAB kits
were used for visualization and assessments. All IHC staining was
carried out in duplicate. Nuclear and cytoplasmic staining was
quantified and recorded. Three experienced postdoctoral observers
scored in a blinded manner at ×100 magnification based on a scoring
system to measure staining (0, none; 1, faint; 2, moderate; 3,
strong). (A and B) APE/Ref-1 immunostaining in normal liver tissue,
and HCC tissue that exhibited a significant reactivity both in
nucleus and cytoplasm (p<0.00001). Shown is a representative
image with the score of +++ and +++ in the cytoplasm and nucleus
(0, none; 1, faint; 2, moderate; 3, strong). (C and D) Cytoplasmic
reactivity is closely correlated with tumor grades (p<0.05) and
with tumor stages (p<0.001). *p<0.05;
**p<0.01 versus control. Columns, mean (n=3); bars,
SD. |
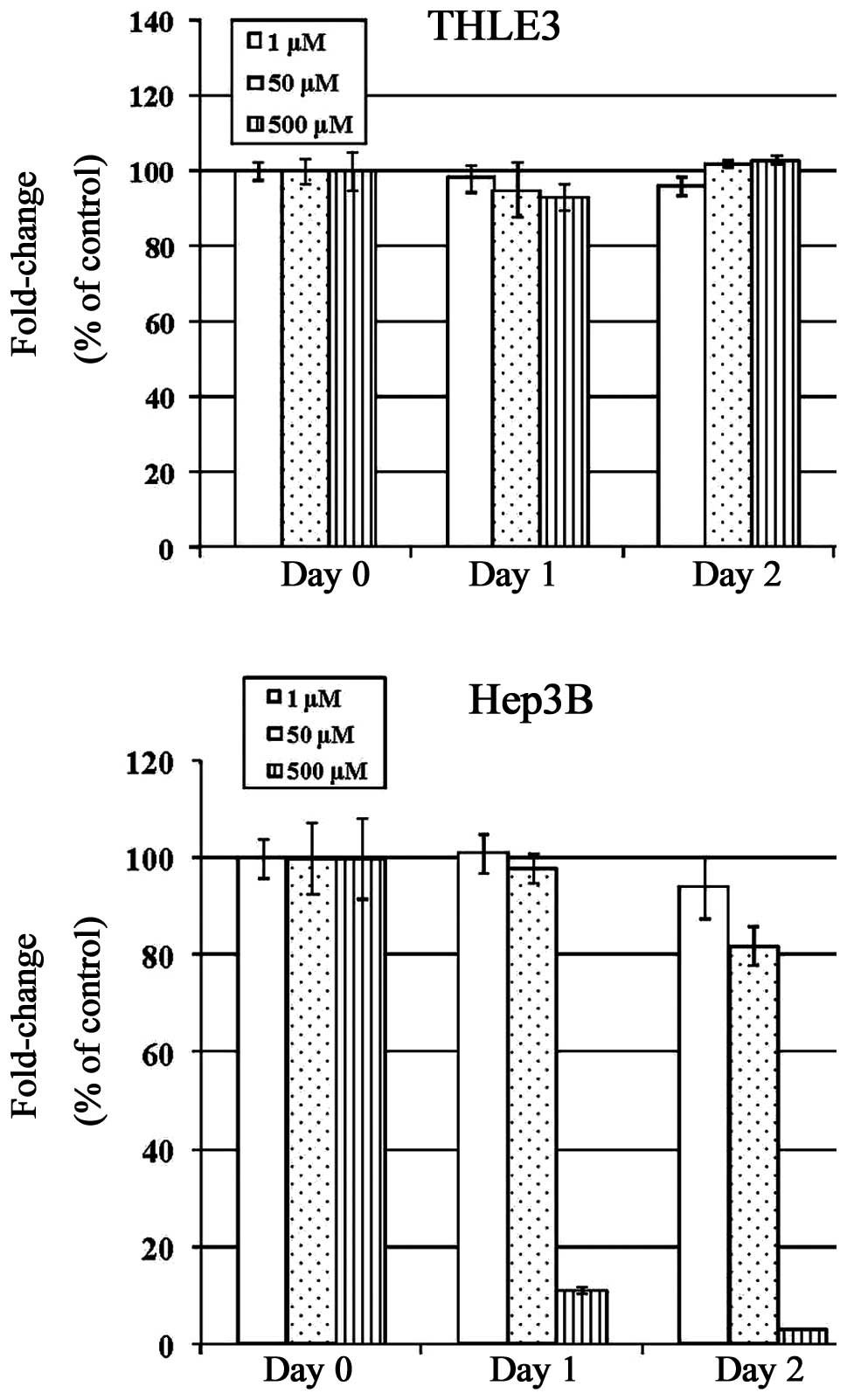
Effects of Cu treatment on cell
proliferation
Elevated Cu levels in serum and tumors have been
well documented. From our results, Cu treatment initiated the
proliferation of THLE3 hepatocytes even at a 500 μM concentration.
However, the HCC Hep3B cells were much more sensitive to Cu
treatment, probably due to the higher Cu-load existence and
consequently less buffering regulatory capacity. Based on these
finding, concentrations for subsequent experiments were determined
as 50 μM (Fig. 3).
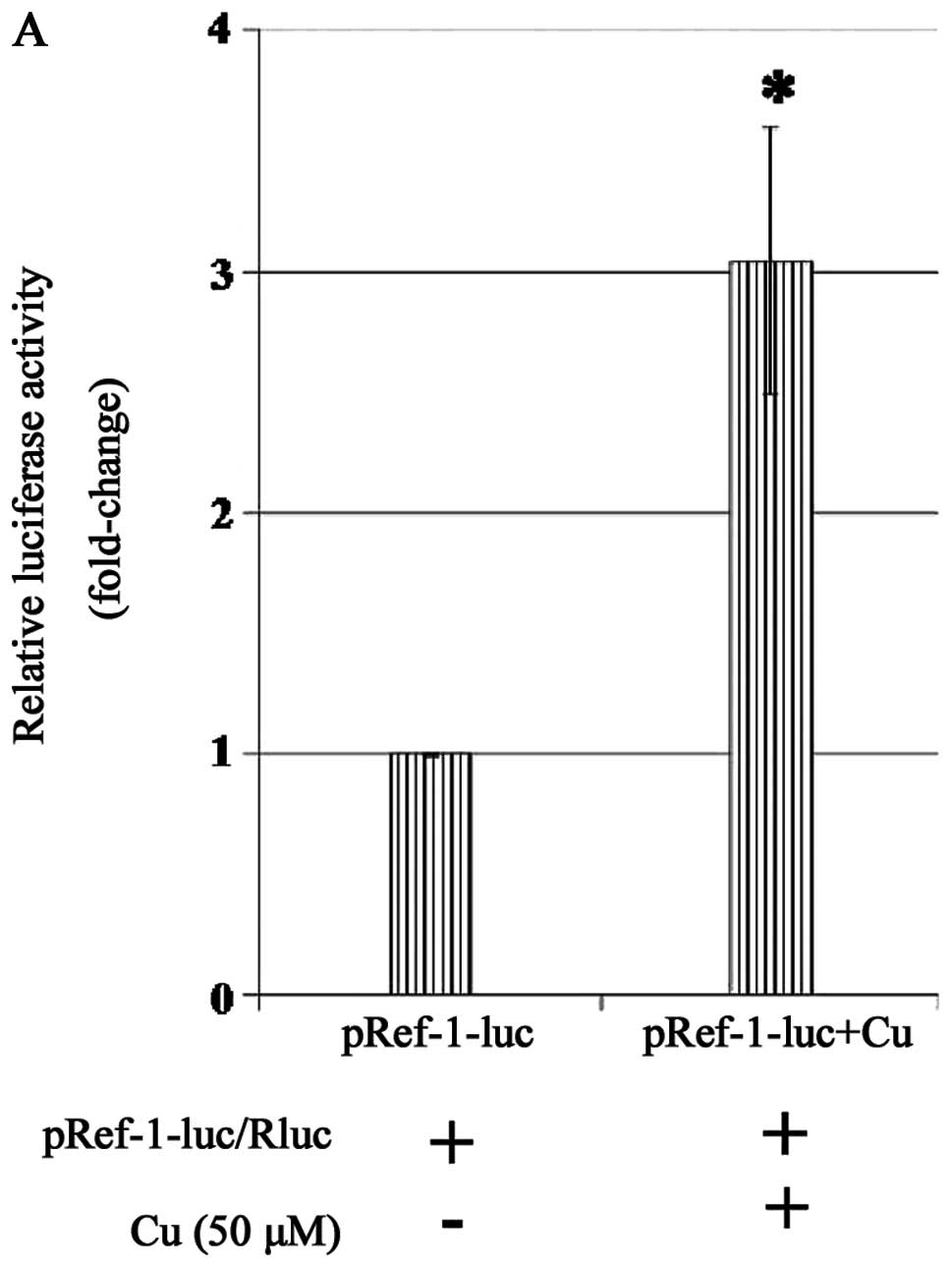
Cu treatment induces APE/Ref-1 as well as
its target genes in THLE3 hepatocytes and Hep3B cells
For a better understanding of the correlation
between Cu and APE/Ref-1, the activity of the APE/Ref-1 promoter
was measured in hepatocytes with Cu treatment using a
Dual-Luciferase Reporter assay. Cu elicited an elevated luciferase
activity ~3-fold that with the vector only (Fig. 4A), as well as the induction of
APE/Ref-1 and its downstream signals confirmed by western blotting
(Fig. 4B). In Hep3B cells, Cu
treatment also induced APE/Ref-1 but much less than that of THLE3
cells (Fig. 4C). In addition,
real-time RT-PCR was performed to detect E-cadherin and Snail mRNA
level in Hep3B cells. Following Cu treatment, mRNA level of Snail
elevated significantly along with the repression of E-cadherin
(Fig. 4D). Importantly, Cu
treatment led to the translocation of APE/Ref-1 quantified by the
ratio of cytoplasmic/nuclear relative density values (Fig. 4E). Enhanced cytoplasmic
accumulation of APE/Ref-1 was accompanied with an upregulation of
MMP-1 and Snail and a downregulation of E-cadherin, proposing that
Cu and APE/Ref-1 cytoplasmic distribution might be associated with
HCC development.
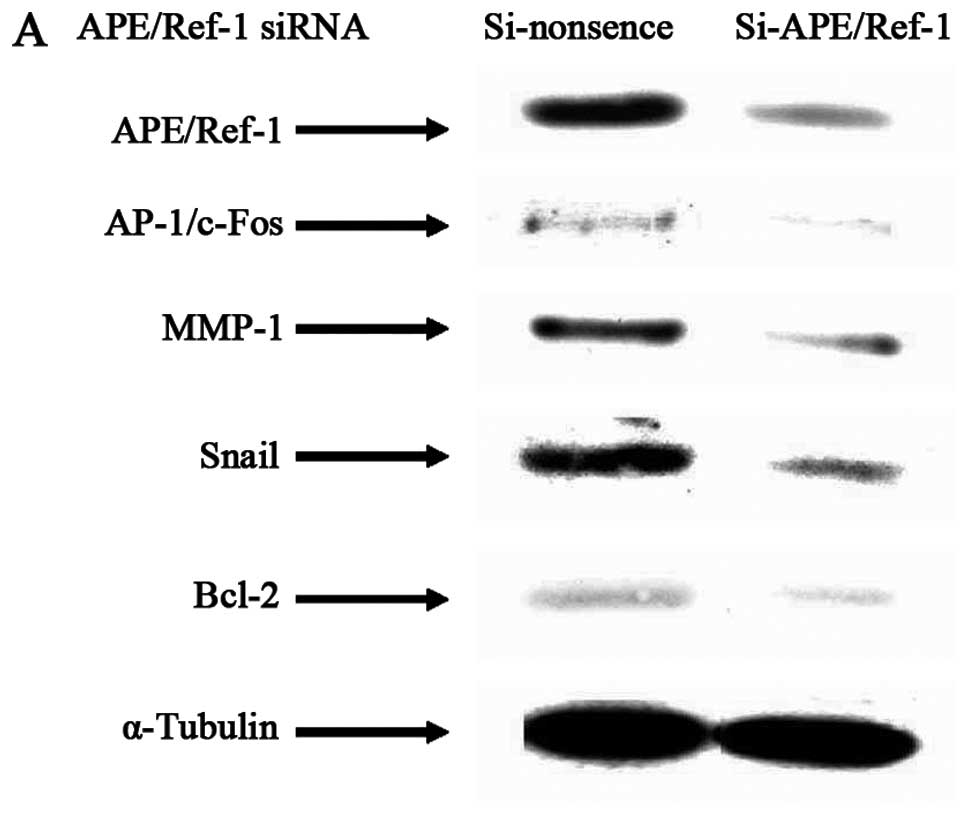
Targeted APE/Ref-1 depletion in HCC cells
leads to decreased AP-1/c-Fos, MMP-1, Snail and Bcl-2 in parallel
with corresponding alterations of proliferation, apoptosis and
invasion activities
APE/Ref-1 expression is very strong in Hep3B cells,
and transient knockdown of APE/Ref-1 using siRNA was analyzed to
explore the role of APE/Ref-1 in HCC. Knocking APE/Ref-1 down
inhibited AP-1/c-Fos, MMP-1, Snail and Bcl-2 expression, but
increased mRNA levels of E-cadherin (Fig. 5A and B). Consistently,
proliferation was suppressed, invasive activity was reduced, and
the apoptosis rate increased with depletion of APE/Ref-1 (Fig. 5C–E). After a 48-h transfection,
cell viability decreased to 52% and invasive activity to 56% of
control cells; apoptosis rate (9.6%) was 13.5-fold higher than that
of control (0.71%). These data support the notion that APE/Ref-1
potentiate HCC progression by being a key mediator and
regulator.
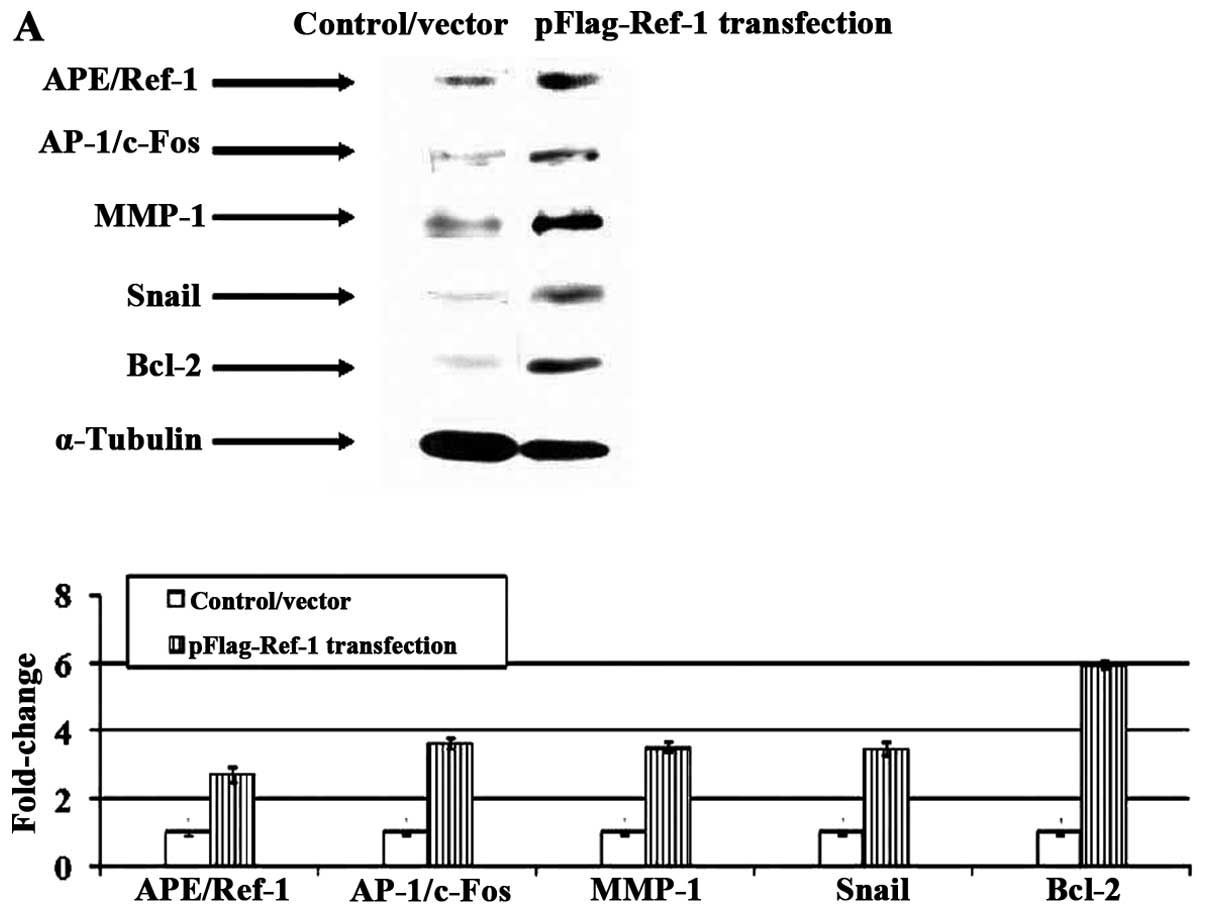
Overexpression of APE/Ref-1 in
hepatocytes potentiates the proliferative and metastatic capacities
inducing its downstream targets
To further solidify the role of APE/Ref-1 in HCC
progression, pFlag-Ref-1 plasmids were transfected into hepatocytes
in which APE/Ref-1 expression level is very low. As shown in
Fig. 6A, overexpression of
APE/Ref-1 resulted in the activation of its downstream genes. There
was a 3.6-(AP-1/c-Fos), 3.5-(MMP-1), 6.0-(Bcl-2), 3.5-(Snail)-fold
increase. Furthermore, cell proliferation and invasiveness were
assessed. By 48 h, cell viability was ~1.2-fold higher (Fig. 6B). Matrigel invasion assay
confirmed that the average number of transfected cells invading the
underside of the membrane was 2 times greater than the control
(Fig. 6C).
Discussion
The present study was designed to understand the
role of APE/Ref-1 in HCC progression. In a Cu-rich environment,
APE/Ref-1 transcription was activated, as well as the induction of
its downstream signaling cascades; additionally, translocation of
APE/Ref-1 from the nucleus to the cytoplasm was also observed along
with the activation of Snail and repression of E-cadherin. Knocking
APE/Ref-1 down reduced the expression of AP-1/c-Fos, MMP-1, Bcl-2
and Snail in parallel with corresponding decreases of cell
proliferation and invasiveness, and an increased apoptosis rate.
Consistently, overexpression of APE/Ref-1 in hepatocytes initiated
a series of intracellular signal activations that resulted in
enhanced cell proliferative and invasive capacities. Specifically,
greater cytoplasmic reactivity of APE/Ref-1 was tightly correlated
with a poorly differentiated (grade 3) and more aggressive tumors
(stage 3–4), although both nuclear and cytoplasmic APE1/Ref-1
expressions were significantly higher in HCC tissue than in normal
liver tissue. These data suggest Cu is well correlated with HCC
progression in which APE/Ref-1 occupies a crucial position as an
important mediator and regulator, and also presents evidence to
support the prognostic role of APE/Ref-1 in the outcome of HCC
patients.
It is well known that the liver is the main organ
for metabolism of Cu. Excess Cu has been demonstrated to be a
potent oxidizer causing production of reactive oxygen species
(ROS), that in turn would create the existence of an oxidative
stress related cellular disorder, such as cancer (23,24).
However, the underlying mechanisms remain to be defined. In the
present study, Cu promoted hepatocytes proliferation and enhanced
APE/Ref-1 and its target genes expression as well, suggesting that
APE/Ref-1 might be of great significance in mediating the
stimulation contributing to HCC progression.
APE/Ref-1 is well documented to be very sensitive to
oxidative stress and implicated in different areas, primarily as a
unique link between the DNA base excision repair pathway, redox
balance, transcription factor regulation and carcinogenesis
(25). Aberrant enhanced
expression of APE/Ref-1 appears to be a general phenomenon in human
cancer (5–11). In this study, targeted depletion of
APE/Ref-1 in HCC cells resulted in decreased proliferation and
invasion, and increased apoptosis rate in parallel with attenuation
in the expression of AP-1/c-Fos, MMP-1, Bcl-2 and Snail. On the
other hand, overexpressed APE/Ref-1 in hepatocytes elicited the
induction of its related signaling cascades leading cells to be
more proliferative and invasive, supporting the role of APE/Ref-1
in mediating and potentiating HCC progression. Interestingly, the
ratio of cytoplasmic to nuclear APE/Ref-1 accumulation changed with
a marked increase following Cu treatment. In addition, our tissue
array data demonstrated further that cytoplasmic intensity was
closely correlated with differentiation and invasiveness,
suggesting that different nuclear and cytoplasmic distribution of
APE/Ref-1 should be pursued as a novel prognostic factor for
clinical outcome.
As expected, AP-1/c-Fos could be regulated by
APE/Ref-1; depletion of APE/Ref-1 in Hep3B cells was associated
with a reduction in AP-1/c-Fos expression levels as well as cell
proliferative capacity, suggesting that induced APE/Ref-1
contributes to cell proliferation through AP-1/c-Fos activation.
Furthermore, depletion of APE/Ref-1 also resulted in evident
attenuation of cell invasiveness, possibly mediated by decreased
activities of MMP-1 and Snail. MMP-1 was considered to be closely
associated with tumor aggressiveness, metastatic potential and poor
prognosis (26–29). Specifically, MMP-1 expression is
largely controlled at the transcriptional level through a major
enhancer element consisting of a consensus AP-1 site (30–32).
Therefore, it is proposed that APE/Ref-1 induces AP-1, and then
MMP-1 is activated leading cells to be more aggressive and
metastatic. Additionally, Hep3B cells are well differentiated and
exhibited intensive MMP-1 expression, implying that MMP-1 is also
needed in the early phases of invasion at the site of origin and
probably plays a less critical role in the stages when such
metastases are well established (33). More importantly, evidence from the
literature showed that MMP-1 was induced in Snail overexpressed
cells (34). Snail, the
zinc-finger transcription factor, has been implicated in
accelerating cancer invasion by repressing E-cadherin and
increasing MMP gene expression, especially MMP-1 (34–36).
In agreement with this, Snail expression was found to parallel that
of MMP-1 in hepatocytes and HCC cells. Knockdown of APE/Ref-1 in
HCC cells was associated with evident decreased Snail and MMP-1,
and increased E-cadherin. Moreover, APE/Ref-1 overexpressing
hepatocytes, Snail and MMP-1 were upregulated. Taken together, Cu
exposure would induce APE/Ref-1, in turn stimulate MMP-1 and Snail,
and ultimately suppress E-cadherin expression. As a result, HCC
cells would be growing aggressive, leading to vascular invasion and
intrahepatic metastasis (36).
In the present study, knockdown of APE/Ref-1 induced
HCC cells to undergo apoptosis. We focused our attention on Bcl-2,
which is an important downstream molecule regulated by NF-κB.
Generally, its activation contributes to cell survival, while
reduced activity promotes apoptosis in response to a variety of
stimuli (37,38). In particular, it was supposed
recently that apoptosis of cancer cell is mediated by p53 (39,40);
however, the Hep3B cell line is p53-deficient (P53−/−)
with stronger Bcl-2 expression, suggesting that cells depend on the
upregulation of Bcl-2 mediated by APE/Ref-1 to prevent apoptosis
and further facilitate survival and development.
In conclusion, one of the cellular responses to Cu
stress is activation of the APE/Ref-1 pathway that regulates
cellular proliferation, apoptosis and metastatic capacity. Cu
exposure induced APE/Ref-1 and also changed the subcellular
distribution of APE/Ref-1 protein from the nucleus to the
cytoplasm. Overexpression or knockdown of APE/Ref-1 leads to
corresponding alterations both in target gene expression and cell
activities. Therefore, APE/Ref-1 overexpression in HCC might be an
important event in the progression of HCC. Further investigations
and development of APE/Ref-1 inhibitors as anticancer agents may be
promising.
Acknowledgements
We would like to thank Dr Kaikobad Irani (University
of Pittsburg) for his gift of pFlag-Ref-1, Dr Bruce Demple (Harvard
School of Public Health) for his gift of human Ref-1 promoter
construct pCB2, Farah Akhtar for technical assistance, Randal Berg,
Ryan Dellinger and Zhenyu Jia for comments on the manuscript. This
study was supported in part by National Natural Science Foundation
of China (no. 81302122) and Foundation of Shandong Health
Department (2013WS0102).
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
APE/Ref-1
|
apurinic/apyrimidinic
endonuclease-1/redox factor-1
|
|
Cu
|
copper
|
|
ROS
|
reactive oxygen species
|
|
AP-1
|
activator protein-1
|
|
MMP-1
|
matrix metalloproteinase-1
|
|
NF-κB
|
nuclear factor κB
|
|
RT-PCR
|
reverse transcription-polymerase chain
reaction
|
References
|
1
|
Scaggiante B, Kazemi M, Pozzato G, et al:
Novel hepatocellular carcinoma molecules with prognostic and
therapeutic potentials. World J Gastroenterol. 20:1268–1288. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bruix J, Boix L, Sala M and Llovet JM:
Focus on hepatocellular carcinoma. Cancer Cell. 5:215–219. 2004.
View Article : Google Scholar
|
|
3
|
Tien Kuo M and Savaraj N: Roles of
reactive oxygen species in hepatocarcinogenesis and drug resistance
gene expression in liver cancers. Mol Carcinog. 45:701–709.
2006.PubMed/NCBI
|
|
4
|
Tell G, Quadrifoglio F, Tiribelli C and
Kelley MR: The many functions of APE1/Ref-1: not only a DNA repair
enzyme. Antioxid Redox Signal. 11:601–620. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bobola MS, Blank A, Berger MS, Stevens BA
and Silber JR: Apurinic/apyrimidinic endonuclease activity is
elevated in human adult gliomas. Clin Cancer Res. 7:3510–3518.
2001.PubMed/NCBI
|
|
6
|
Kelley MR, Cheng L, Foster R, et al:
Elevated and altered expression of the multifunctional DNA base
excision repair and redox enzyme Ape1/ref-1 in prostate cancer.
Clin Cancer Res. 7:824–830. 2001.PubMed/NCBI
|
|
7
|
Moore DH, Michael H, Tritt R, Parsons SH
and Kelley MR: Alterations in the expression of the DNA
repair/redox enzyme APE/ref-1 in epithelial ovarian cancers. Clin
Cancer Res. 6:602–609. 2000.PubMed/NCBI
|
|
8
|
Russo D, Arturi F, Bulotta S, et al:
ApeI/Ref-I expression and cellular localization in human thyroid
carcinoma cell lines. J Endocrinol Invest. 24:10–12. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Robertson KA, Bullock HA, Xu Y, et al:
Altered expression of Ape1/ref-1 in germ cell tumors and
overexpression in NT2 cells confers resistance to bleomycin and
radiation. Cancer Res. 61:2220–2225. 2001.PubMed/NCBI
|
|
10
|
Wang D, Luo M and Kelley MR: Human
apurinic endonuclease 1 (APE1) expression and prognostic
significance in osteosarcoma: enhanced sensitivity of osteosarcoma
to DNA damaging agents using silencing RNA APE1 expression
inhibition. Mol Cancer Ther. 3:679–686. 2004.
|
|
11
|
Puglisi F, Aprile G, Minisini AM, et al:
Prognostic significance of Ape1/ref-1 subcellular localization in
non-small cell lung carcinomas. Anticancer Res. 21:4041–4049.
2001.PubMed/NCBI
|
|
12
|
Kawanishi S, Hiraku Y, Murata M and Oikawa
S: The role of metals in site-specific DNA damage with reference to
carcinogenesis. Free Radic Biol Med. 32:822–832. 2002.PubMed/NCBI
|
|
13
|
Li Y, Togashi Y, Sato S, et al: Abnormal
copper accumulation in non-cancerous and cancerous liver tissues of
LEC rats developing hereditary hepatitis and spontaneous hepatoma.
Jpn J Cancer Res. 82:490–492. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Goodman VL, Brewer GJ and Merajver SD:
Copper deficiency as an anti-cancer strategy. Endocr Relat Cancer.
11:255–263. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu T, Sempos CT, Freudenheim JL, Muti P
and Smit E: Serum iron, copper and zinc concentrations and risk of
cancer mortality in US adults. Ann Epidemiol. 14:195–201. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kuo KW, Chen SF, Wu CC, Chen DR and Lee
JH: Serum and tissue trace elements in patients with breast cancer
in Taiwan. Biol Trace Elem Res. 89:1–11. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zuo XL, Chen JM, Zhou X, Li XZ and Mei GY:
Levels of selenium, zinc, copper and antioxidant enzyme activity in
patients with leukemia. Biol Trace Elem Res. 114:41–53. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tashiro H, Kawamoto T, Okubo T and Koide
O: Variation in the distribution of trace elements in hepatoma.
Biol Trace Elem Res. 95:49–63. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Goodman VL, Brewer GJ and Merajver SD:
Control of copper status for cancer therapy. Curr Cancer Drug
Targets. 5:543–549. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kontoghiorghes GJ, Efstathiou A,
Ioannou-Loucaides S and Kolnagou A: Chelators controlling metal
metabolism and toxicity pathways: applications in cancer
prevention, diagnosis and treatment. Hemoglobin. 32:217–227. 2008.
View Article : Google Scholar
|
|
21
|
Harrison L, Ascione AG, Wilson DM III and
Demple B: Characterization of the promoter region of the human
apurinic endonuclease gene (APE). J Biol Chem. 270:5556–5564. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang S and Meyskens FL Jr: Alterations in
activating protein 1 composition correlate with phenotypic
differentiation changes induced by resveratrol in human melanoma.
Mol Pharmacol. 67:298–308. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gupte A and Mumper RJ: Elevated copper and
oxidative stress in cancer cells as a target for cancer treatment.
Cancer Treat Rev. 35:32–46. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Valko M, Rhodes CJ, Moncol J, Izakovic M
and Mazur M: Free radicals, metals and antioxidants in oxidative
stress-induced cancer. Chem Biol Interact. 160:1–40. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Evans AR, Limp-Foster M and Kelley MR:
Going APE over APE/Ref-1. Mutat Res. 461:83–108. 2001. View Article : Google Scholar
|
|
26
|
Merdad A, Karim S, Schulten HJ, et al:
Expression of matrix metalloproteinases (MMPs) in primary human
breast cancer: MMP-9 as a potential biomarker for cancer invasion
and metastasis. Anticancer Res. 34:1355–1366. 2014.PubMed/NCBI
|
|
27
|
Ma H, Cai H, Zhang Y, et al: Membrane
palmitoylated protein 3 promotes hepatocellular carcinoma cell
migration and invasion via up-regulating matrix metalloproteinase
1. Cancer Lett. 344:74–81. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dey S, Ghosh N, Saha D, Kesh K, Gupta A
and Swarnakar S: Matrix metalloproteinase-1 (MMP-1) promoter
polymorphisms are well linked with lower stomach tumor formation in
eastern Indian population. PLoS One. 9:e880402014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Teshima J, Doi H, Fujimori K, et al: A
human thyroid cancer cell line, DH-14–3, newly established from
poorly differentiated thyroid carcinoma. Tohoku J Exp Med.
230:75–82. 2014.PubMed/NCBI
|
|
30
|
Angel P, Baumann I, Stein B, Delius H,
Rahmsdorf HJ and Herrlich P: 12-O-tetradecanoyl-phorbol-13-acetate
induction of the human collagenase gene is mediated by an inducible
enhancer element located in the 5′-flanking region. Mol Cell Biol.
7:2256–2266. 1987.PubMed/NCBI
|
|
31
|
Gutman A and Wasylyk B: The collagenase
gene promoter contains a TPA and oncogene-responsive unit
encompassing the PEA3 and AP-1 binding sites. EMBO J. 9:2241–2246.
1990.PubMed/NCBI
|
|
32
|
Butticè G, Duterque-Coquillaud M, Basuyaux
JP, Carrère S, Kurkinen M and Stéhelin D: Erg, an Ets-family
member, differentially regulates human collagenase1 (MMP1) and
stromelysin1 (MMP3) gene expression by physically interacting with
the Fos/Jun complex. Oncogene. 13:2297–2306. 1996.
|
|
33
|
Bendardaf R, Buhmeida A, Ristamäki R,
Syrjänen K and Pyrhönen S: MMP-1 (collagenase-1) expression in
primary colorectal cancer and its metastases. Scand J
Gastroenterol. 42:1473–1478. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Miyoshi A, Kitajima Y, Sumi K, et al:
Snail and SIP1 increase cancer invasion by upregulating MMP family
in hepatocellular carcinoma cells. Br J Cancer. 90:1265–1273. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cano A, Perez-Moreno MA, Rodrigo I, et al:
The transcription factor Snail controls epithelial-mesenchymal
transitions by repressing E-cadherin expression. Nat Cell Biol.
2:76–83. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Miyoshi A, Kitajima Y, Kido S, et al:
Snail accelerates cancer invasion by upregulating MMP expression
and is associated with poor prognosis of hepatocellular carcinoma.
Br J Cancer. 92:252–258. 2005.PubMed/NCBI
|
|
37
|
Czabotar PE, Lessene G, Strasser A and
Adams JM: Control of apoptosis by the BCL-2 protein family:
implications for physiology and therapy. Nat Rev Mol Cell Biol.
15:49–63. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Han CR, Jun do Y, Lee JY and Kim YH:
Prometaphase arrest-dependent phosphorylation of Bcl-2 and Bim
reduces the association of Bcl-2 with Bak or Bim, provoking Bak
activation and mitochondrial apoptosis in nocodazole-treated Jurkat
T cells. Apoptosis. 19:224–240. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cao W, Zhang JL, Feng DY, et al: The
effect of adenovirus-conjugated NDRG2 on p53-mediated apoptosis of
hepatocarcinoma cells through attenuation of nucleotide excision
repair capacity. Biomaterials. 35:993–1003. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yang CF, Peng LX, Huang TJ, et al: Cancer
stem-like cell characteristics induced by EB virus-encoded LMP1
contribute to radioresistance in nasopharyngeal carcinoma by
suppressing the p53-mediated apoptosis pathway. Cancer Lett.
344:260–271. 2014. View Article : Google Scholar
|