Introduction
Increased expression and alteration in sulfation
pattern of chondroitin sulfate glycosaminoglycans (CS-GAGs) are
observed in various neoplastic tissues, including pancreatic, lung,
ovarian and breast (1–3). Several studies, including ours,
suggest involvement of particular CS chains in tumor progression
and metastasis (2,4–7). The
sulfation by site-specific chondroitin sulfotransferases controls
the biological function of CS-GAGs (8–11).
CHST11 is a key enzyme in the biosynthesis of chondroitin sulfates
(CS) and its action on chondroitin chains can lead to the
production of chondroitin sulfate A (CS-A), B (CS-B) and E (CS-E)
(12,13). We have shown that the expression of
CHST11 is upregulated in tumor tissues of breast cancer
patients compared to their normal tissues and that the expression
levels of the gene in breast cancer cell lines correlate with CS-A
expression, P-selectin binding and aggressive phenotype (14). However, the clinical significance
of CHST11 expression is yet to be established and more studies are
needed to evaluate the expression of this gene in relation to
cancer progression.
We have observed that the expression of
CHST11 is low to none in MCF7 cells with considerably higher
expression in MDA-MB-231 (14).
The expression levels correlated with P-selectin reactivity and
general aggressiveness. The expression of P-selectin ligands on
tumor cells plays a role in distant metastasis by facilitating
tumor cell extravasation (15–18).
Because of its potential role in breast cancer metastasis,
determining mechanisms controlling CHST11 expression is of
great interest. Aberrant DNA methylation is a mechanism that
controls gene expression and is involved in tumor initiation and
progression (19–23). In a gene profiling study of head
and neck cancer it was suggested that DNA methylation may affect
CHST11 expression in laryngeal carcinoma (24). In another study on breast cancer
cell lines using genome wide methylation profiling, the authors
reported that CHST11 is hypermethylated in ER-positive and
hypomethylated in ER-negative cell lines (25). Therefore, variation in the
expression of CHST11 in breast cancer cells may be
controlled by DNA methylation.
Our current data suggest that the expression levels
of CHST11 can predict progression of breast cancer and more
aggressive phenotypes. Our investigation of the methylation status
of CHST11 in breast cancer cells clearly demonstrates an
association between lack of expression of this gene and
hypermethylation of a CpG island of its DNA. Treatment of MCF7
cells with 5AzadC led to a dose-dependent increase in the
expression of the gene and its product CS-A. Given the role of CS
and the significance of CHST11 expression in tumor growth
and metastasis, these findings have significant implications for
the development of novel prognostic strategies in breast
cancer.
Materials and methods
Reagents
Anti-CS-A mAb 2H6 was from Associates of Cape
Cod/Seikagaku America (Falmouth, MA, USA). Fluorescence-conjugated,
anti-mouse IgM and 5-aza-2′-deoxycytidine (5AzadC) were from Sigma
(St. Louis, MO, USA). Primers were from Integrated DNA Technologies
(IDT, Coralville, IA, USA). Real-time PCR reagents were from
Applied Biosystems (Foster City, CA, USA). TRIzol reagent was from
Invitrogen (Carlsbad, CA, USA), FailSafe PCR PreMix Selection kits
and Enzyme Mix were from Epicentre Biotechnologies (Madison, WI,
USA). Alkaline phosphatase (rAPid) was from Roche (Nutley, NJ,
USA).
Analysis of Oncomine cancer gene
microarray database
Publicly available Oncomine cancer microarray
database (Compendia Biosciences; Ann Arbor, MI, USA; www.oncomine.com) was used to examine the expression
of CHST11 in cancer tissue and determine association of
CHST11 expression with breast cancer outcomes. Richardson
et al (GEO accession GSE3744) (26), Finak et al (GEO accession
GSE9014) (27), The Cancer Genome
Atlas (TCGA, http://tcga-data.nci.nih.gov/tcga/) invasive breast
carcinoma gene expression data and Gluck et al (GEO
accession GSE22358) (28) dataset
were used to compare CHST11 expression levels between cancer
and normal tissues. Hoeflich et al (GEO accession GSE12777)
(29) dataset was used to compare
expression levels of CHST11 among a panel of cell lines that
represent luminal, Her-2-amplified and basal-like molecular
subtypes of breast cancer. Schuetz et al dataset (GEO
accession GSE3893) (30) was used
to compare CHST11 expression levels between matched ductal
carcinoma in situ (DCIS) and invasive ductal carcinoma
(IDC). Log-transformed, median-centered and normalized expression
values (31) were extracted,
analyzed and graphed accordingly.
Cell lines and tissue specimen
Cell lines MCF7, MDA-MB-231, MDA-MB-468, T47-D, and
ZR-75-1 were from ATCC (Manassas, VA, USA). MDA-MB-231, MDA-MB-468,
T47-D, and ZR-75-1 cells were cultured in DMEM supplemented with
10% heat-inactivated fetal bovine serum (Life Technologies), 50
U/ml penicillin and 50 μg/ml streptomycin. MCF7 were grown as
described before (14). Cells are
checked every six months to be free from Mycoplasma contamination
using the MycoAlert® Mycoplasma Detection kit (Lonza
Rockland Inc., Rockland, ME, USA).
De-identified paraffin-embedded specimens from 5
female breast cancer patients were provided by the Department of
Pathology of the University of Arkansas for Medical Sciences
(UAMS). For this study, an active human tissue use protocol
approved by the UAMS Institutional Review Board was used.
Total RNA isolation and quantitative
real-time RT-PCR (real-time RT-qPCR)
RNA isolation and quantitative real-time was
performed as described earlier (14). Briefly, total RNA was isolated from
cultured cells using TRIzol reagent (Life Technologies, Grand
Island, NY, USA) following the manufacturer’s instructions. The
quantity and quality of the isolated RNA was determined using an
Agilent 2100 Bioanalyzer (Palo Alto, CA, USA). Total RNA (1 μg) was
reverse-transcribed using random-hexamer primers with TaqMan
Reverse Transcription reagents (Applied Biosystems).
Reverse-transcribed RNA was amplified with SYBR Green PCR Master
Mix (Applied Biosystems) plus 0.3 μM of gene-specific upstream and
downstream primers during 40 cycles on an Applied Biosystems 7900
HT Fast Real-time system. Data were analyzed by absolute and
relative quantification. In absolute quantification, data were
expressed in relation to 18S RNA, where the standard curves were
generated using pooled RNA from the samples assayed. In relative
quantification, the 2−ΔΔCT method was used to
assess the target transcript in a treatment group to that of
untreated control using expression of an internal control
(reference gene) to normalize data (32). Expression of GAPDH was used as
internal control. The primer sequences are shown in Table I.
 | Table IPrimers used in this study. |
Table I
Primers used in this study.
| Primer | Sequence |
|---|
| 18S forward |
5′-TTCGAACGTCTGCCCTATCAA-3′ |
| 18S reverse |
5′-ATGGTAGGCACGGCGACTA-3′ |
| CHST11
forward |
5′-TCCCTTTGGTGTGGACATCT-3′ |
| CHST11
reverse |
5′-CACGTGTCTGTCACCTGGTC-3′ |
| GAPDH forward |
5′-ACAGTCAGCCGCATCTTCTT-3′ |
| GAPDH reverse |
5′-ACGACCAAATCCGTTGACTC-3′ |
| CHST11
bisulfite outside, forward |
5′-TTTGATTATTGTAGTTTTGGAGGAAAT-3′ |
| CHST11
bisulfite reverse |
5′-CCTTACAATTAAAAAAACAAATTATTACTA-3′ |
| CHST11
bisulfite nested, forward |
5′-TTTTTGTGGTTTAGGTAAAGTTTTA-3′ |
5AzadC treatment
5AzadC was dissolved in ice-cold phosphate-buffered
saline, filter sterilized at 4°C and the resulting solution used to
treat the MCF7 cell line. For dose–response experiments, cells were
harvested 5 days after the initial treatment. Cell growth medium
was refreshed every other day.
Extraction and bisulfite modification of
DNA
DNA from cells was extracted as described before
(33). DNA from the
paraffin-embedded tissues of breast cancer patients was extracted
using Ex-Wax DNA extraction kit (Chemicon International, Temecula,
CA, USA), following the manufacturer’s instructions with the
additional steps of extracting recovered DNA with phenol (Amresco,
Solon, OH, USA) and then 1-bromo-3-chloropropane (Molecular
Research Center Inc., Cincinnati, OH, USA). DNA was bisulfite
modified with an Epitect kit (Qiagen, Valencia, CA, USA) using 300
ng of DNA per reaction. PCR was performed using a FailSafe PCR
PreMix Selection kit and FailSafe Enzyme Mix. Each 25-μl PCR
reaction included 1.0 μM of each primer, 2.5 U of the FailSafe
Enzyme Mix and 12.5 μl of the FailSafe PCR PreMixes A or C.
Bisulfite-modified genomic DNA was amplified by semi-nested PCR
using two sets of primers for part of intron 1 that is within a CpG
island spanning exon 1 of the CHST11 gene (Genbank NM_000012 and
exon 1 located with NM_018413). The same amplification profile was
used for both reactions of the semi-nested PCR: 1 cycle at 80°C for
1 min, 1 cycle at 94°C for 1 min; 1 cycle at 95°C for 1 min, 54°C
for 1 min, 72°C for 1 min; 1 cycle at 95°C for 1 min, 53°C for 1
min, 72°C for 1 min; 1 cycle at 95°C for 1 min, 52°C for 1 min,
72°C for 1 min; 1 cycle at 95°C for 1 min, 51°C for 1 min, 72°C for
1 min; 36 cycles at 95°C for 1 min, 50°C for 1 min, 72°C for 1 min;
72°C for 5 min and cooling to 4°C.
A forward, outside primer and reverse primer were
used for CHST11 for the first reaction (Table I). A second, semi-nested, PCR was
then performed on 1 μl of the amplificate (in a 25-μl PCR reaction)
using a forward nested primer and the reverse primer from the first
reaction (346-bp PCR product, Table
I). The primers were designed using MethPrimer web software
(34) (http://www.urogene.org/methprimer/). The CpG island
was defined using CpG Island Searcher set on the default criteria
for defining CpG islands (35)
(http://cpgislands.usc.edu/cpg.aspx).
Bisulfite genomic sequencing (BGS) and
methylation level quantification
BGS was performed as described before (33) with the following minor
modifications. We used rAPid alkaline phosphatase and PCR products
were sequenced using the nested forward (upstream) CHST11
primer. We also analyzed DNA methylation in the region spanning
>1.5 kb of the CHST11 CpG island with reduced
representation bisulfite sequencing [RRBS (36,37)]
data generated on breast cancer cell lines for the Illumina Idea
Challenge (http://www.illumina.com/landing/idea) (25). RRBS selects DNA fragments ≤220 bp
in the vicinity of MspI recognition sites (C.CGG) (36). We only considered CpG dinucleotides
with coverage >10x. The percentage of methylation was inferred
from the number of methylated reads divided by the sum of
methylated and unmethylated reads. All reads were mapped to the
hg18 release of the human genome (http://genome.ucsc.edu/cgi-bin/hgGateway?db=hg18).
Second generation sequencing DNA methylation results were plotted
in Matlab (http://www.mathworks.com).
Statistical analysis
One-way ANOVA with Tukey’s post hoc procedure
test was used to compare gene expression between subtypes of cell
lines. For comparison of gene expression data generated by
real-time PCR, the raw amount for each mRNA was log transformed and
normalized to the control mRNA (18S) amount, and analyzed via
one-way ANOVA with Tukey’s post hoc procedure. The
Mann-Whitney U test was performed to compare gene expression
between DCIS and IDC. For 5AzadC induced fold change in gene
expression, the mRNA levels of the non-zero doses for each
transcript and experimental replication were normalized to that of
the zero-dose control, transformed to their base-10 logarithms and
analyzed for trend with dose via one-way ANOVA. Statistical
analyses were performed using Excel (Microsoft, Seattle, WA, USA)
or GraphPad Prism version 5.00 for Windows (GraphPad Software, San
Diego, CA, USA). All P-values were 2-sided.
Results
CHST11 is overexpressed in basal-like and
HER2-amplified cell lines and the elevated expression correlates
with tumor progression
We have shown that the expression of CHST11
is elevated in tumor cells compared to normal cells of breast
cancer patient tissue specimens (14). We used Oncomine database to confirm
our original data. The data from several studies were analyzed to
compare CHST11 expression in breast carcinoma versus normal
breast tissue (Table II). The
comparison showed average increases of 2–3.6-fold in CHST11
expression in breast cancer compared to normal tissue.
| Table IICHST11 differential transcript
expression in human breast carcinomas extracted from multiple
studies in the Oncomine microarray database. |
Table II
CHST11 differential transcript
expression in human breast carcinomas extracted from multiple
studies in the Oncomine microarray database.
| Studya | Comparison
(specimen number in each group) | Average fold
increase | P-valueb |
|---|
| TCGAc | Invasive breast
carcinoma (n=76) vs normal (n=61) | 2.6 | 2.51E-24 |
| Finak et al
(27) | Invasive breast
carcinoma (n=53) vs normal (n=6) | 3.6 | 9.43E-20 |
| Richardson et
al (26) | Ductal breast
carcinoma (n=40) vs normal (n=7) | 2.2 | 2.00E-4d |
| Gluck et al
(28) | Invasive breast
carcinoma (n=154) vs. normal (n=4) | 2 | 0.02 |
Our previously published data analyzing
CHST11 expression in a set of commonly used human breast
cancer cell lines suggest that CHST11 expression is low in
luminal and high in basal-like cell lines (14). The data relate CHST11
expression to basal-like cancer cells. To further investigate such
an association, we analyzed an Oncomine dataset that screened 50
breast cancer cell lines representative of the molecular subtypes
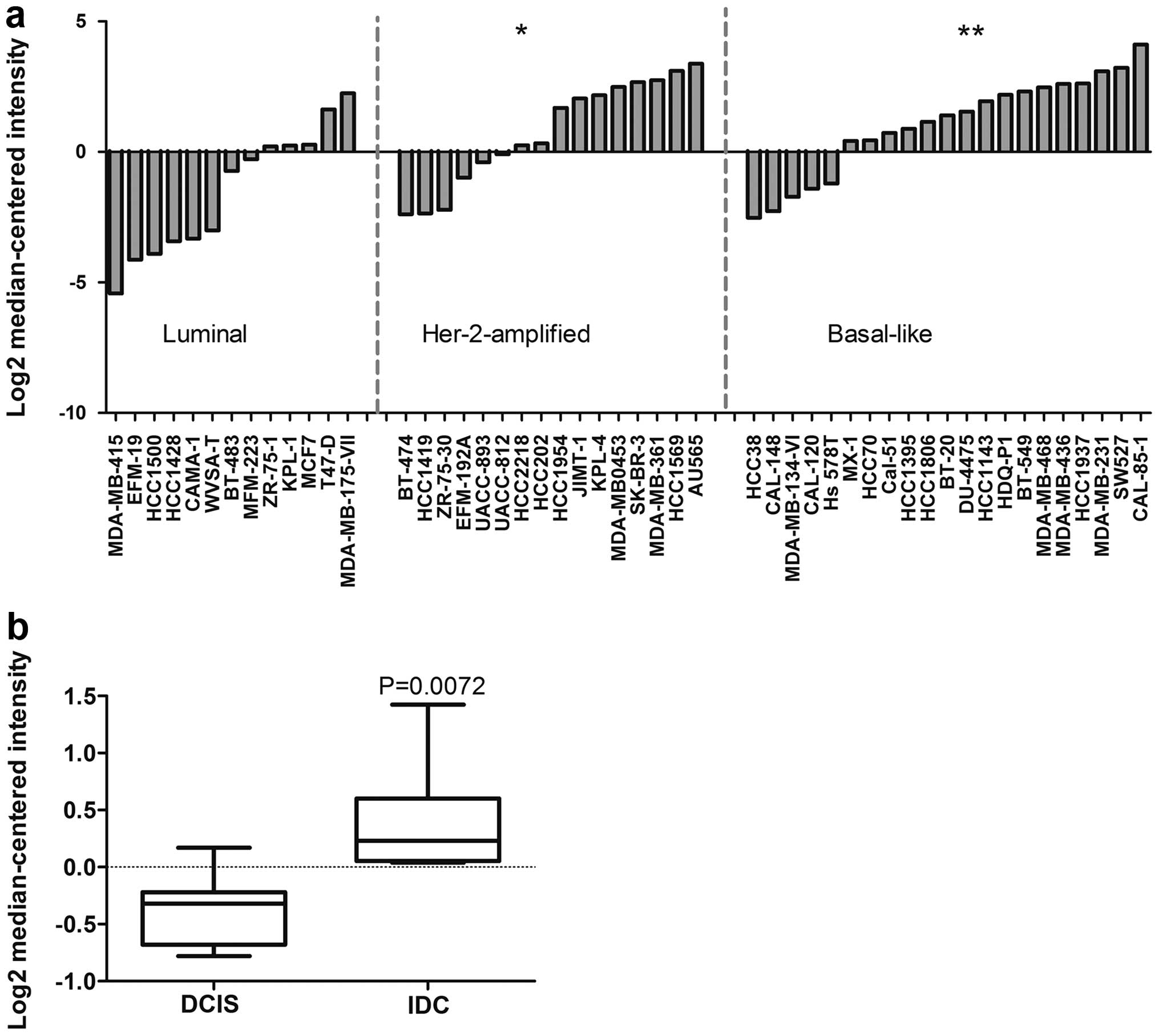
luminal, Her2-amplified, and basal-like (29). We observed that the expression of
CHST11 in basal-like cell lines was significantly higher
than in luminal cell types (Fig.
1a). CHST11 was also significantly overexpressed in
Her-2-amplified cell lines. Considering cells with above average
expression as positive, 75% of basal-like and 56% Her-2-amplified
cell lines were positive; while only 15% of luminal cell lines were
positive for CHST11 expression.
To investigate the association of CHST11
overexpression with tumor progression, we interrogated another
dataset in the Oncomine database, comparing the expression of
CHST11 between ductal carcinoma in situ (DCIS) and
invasive ductal carcinoma (IDC) (30). DCIS and IDC samples were isolated
by laser capture microdissection (LCM) and matched for each
individual specimen (30). We
found a significant increase in the expression of CHST11 in
IDC compared to DCIS samples (Fig.
1b). Notably, both groups displayed a similar distribution
pattern for ER status (5 ER-positive specimens in each group) and
HER2-amplification pattern (3 HER2-amplified specimens in each
group) with histological grades 2 and 3. The data link the elevated
expression of CHST11 to aggressive subtypes and progression
of the disease.
A CpG island in the CHST11 gene sequence
is hypermethylated in MCF7 and hypomethylated in MDA-MB-231
cells
Our current and previously published data suggest
that CHST11 expression plays a role in tumor progression and
metastasis. Therefore understanding the mechanisms controlling
CHST11 gene expression will help formulate strategies for
development of biomarkers and drug targets. We demonstrated
previously that CHST11 and CS-A expression was high in
highly metastatic MDA-MB-231 cells, while it was significantly
lower in MCF7 cells as assayed by qRT-PCR and flow cytometry
(14). To examine whether DNA
methylation controls variation and regulation of CHST11 in
breast cancer cell lines, we examined DNA methylation of the
CHST11 sequence in a section of its CpG island covering part
of its promoter, its first exon, and a portion of its first intron
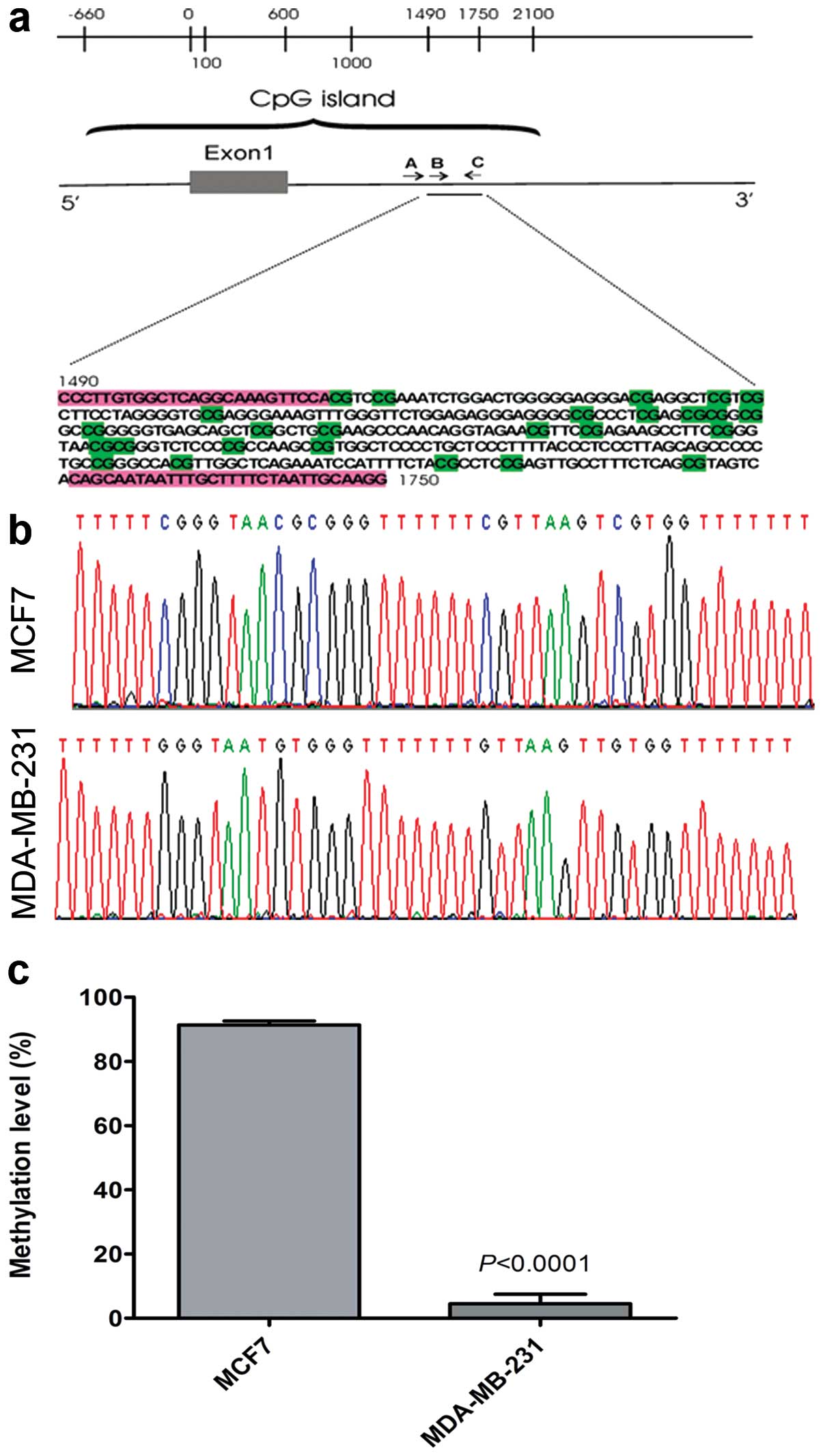
(Fig. 2a). Using BGS and the
Mquant algorithm (33) we
determined methylation levels of ten CpG sites for MCF7 and
MDA-MB-231 cells (Fig. 2b). This
revealed very high methylation levels (91%) averaged >10 CpGs in
the CHST11 sequence in MCF7 cells, and very low levels (5%)
over the same 10 CpGs in MDA-MB-231 cells (Fig. 2c). These observations suggest that
low expression of CHST11 in MCF7 cells is due to DNA
hypermethylation.
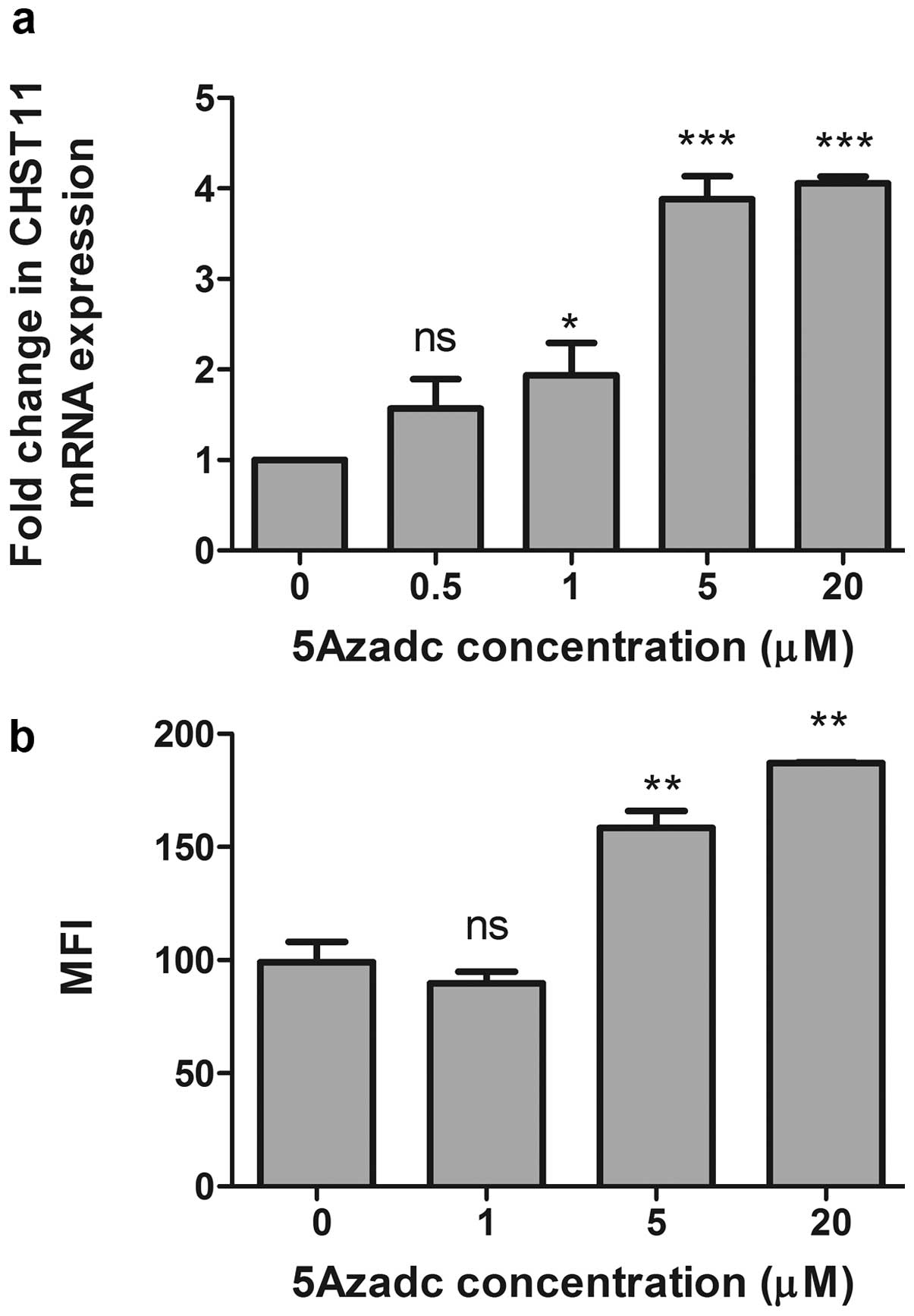
Treatment of MCF7 cells with 5AzadC
increases the expression of CHST11 and CS-A
To further validate our data and confirm that low
expression of CHST11 in the MCF7 cell line is due to
hypermethylation status of the CHST11 CpG island, MCF7 cells
were treated with 5AzadC and CHST11 gene expression was
determined. Upon 5AzadC treatment, we observed a significant
increase in the expression of CHST11 mRNA (Fig. 3a) that paralleled surface
expression of CS-A as assayed by flow cytometry using anti-CS-A mAb
2H6 (Fig. 3b). Together, the data
suggest that low expression of CHST11 in the MCF7 cells is
due to promoter hypermethylation, and that DNA hypomethylation in
the more aggressive mesenchymal-like MDA-MB-231 cell line is
permissive for overt CHST11 expression.
Methylation of the CHST11 CpG island in
other cell lines with low expression of CHST11
To confirm a role for methylation of the
CHST11 CpG island in the expression control of
CHST11, we further analyzed second generation DNA
methylation sequencing data on breast cancer cell lines (Illumina
Idea Challenge and ref. 25). This
method provided information on DNA methylation at single nucleotide
resolution across 162 out of 186 CpGs in the ~2.5 kb long
CHST11 CpG island. Methylation analysis of Illumina Idea
Challenge data of these cell lines indicates a close relationship
between expression levels and methylation status. A very
hypomethylated state was observed for MDA-MB-231 (Fig. 4a). The methylation status of two
other triple-negative cell lines MDA-MB-468 and BT-20, was similar
to MDA-MB-231 (Fig 4b and c).
Similar to MCF7 cells, the CpG island of the CHST11 gene in
the T47-D and ZR-75-1 cell lines, was hypermethylated (Fig. 4d–f). The expression of
CHST11 in T47-D and ZR-75-1 cell lines was further examined
by qRT-PCR (Fig. 4g) and flow
cytometry (Table III), and
compared with CHST11 expression levels in MCF7 and
MDA-MB-231. Consistent with the methylation data, the expression of
the CHST11 gene and CS-A in both of these cell lines was
significantly lower than in MDA-MB-231 and comparable with that of
MCF7 cells. MDA-MB-468 displayed a very hypomethylated state but
the expression of CHST11 in these cells was moderate and
less than what we observed for MDA-MB-231 and MDA-MET (4,14).
The expression of CHST11 in tumor tissue may also be
controlled by DNA methylation similar to that observed in cell
lines.
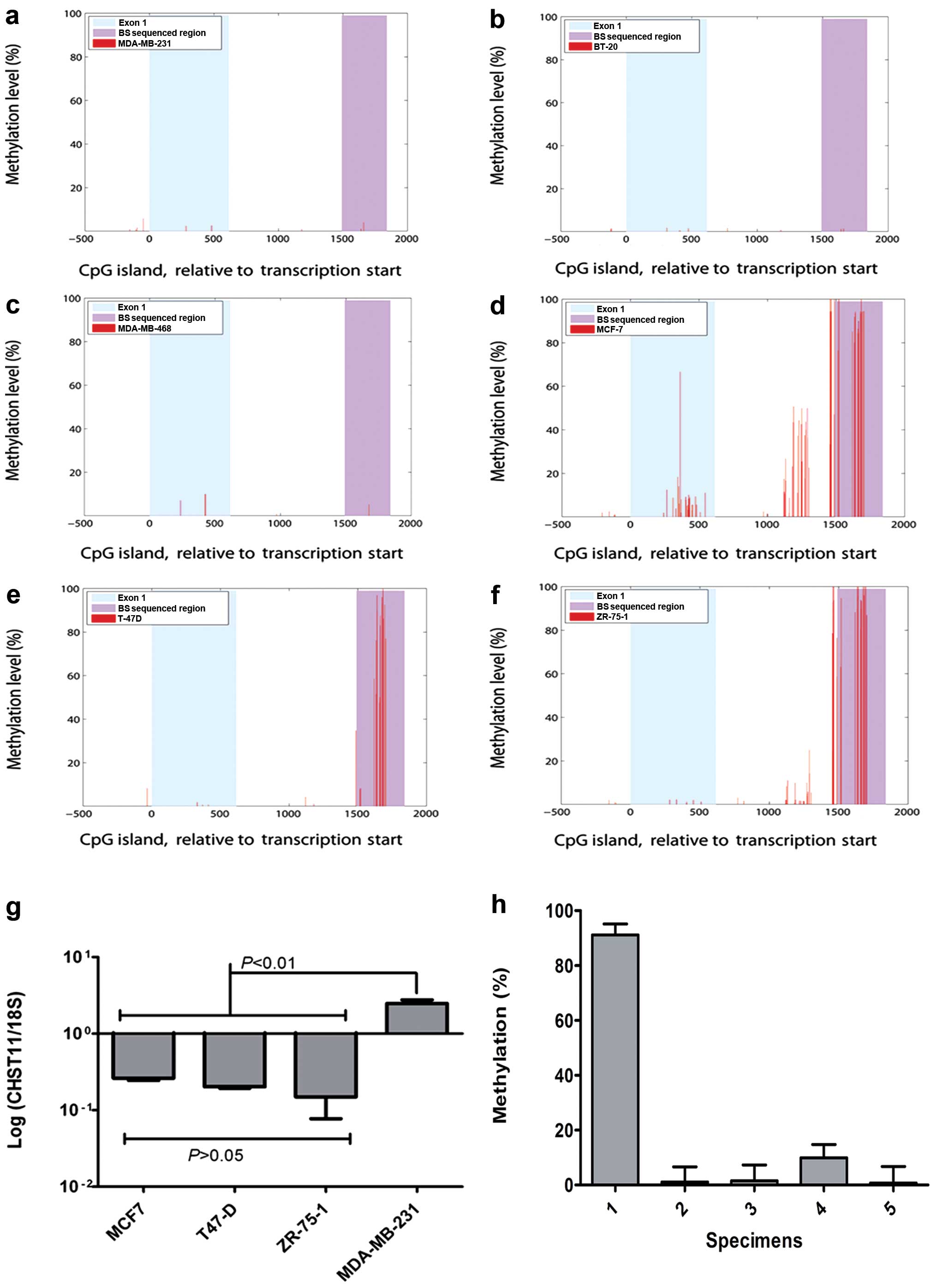 | Figure 4CHST11 expression and
methylation status of its CpG island in basal-like and luminal-like
breast cancer cell lines and the patient tissues. Single nucleotide
resolution of DNA methylation in the CHST11 CpG island of
MDA-MB-231 (a), BT-20 (b), MDA-MB-468 (c), MCF-7 (d), T-47D (e),
and ZR-75-1 (f) was determined by RRBS (36). The red peaks denote Cs covered with
≥10 reads. Methylation level percentage was calculated from the
number of Cs after bisulfite treatment divided by the sum of Cs
(methylated sites) and Ts (unmethylated sites). There are a total
of 186 CpG dinucleotides in the CpG island, of which ≥75% were
accessible to RRBS with a minimum coverage of 10 (average coverage
≥40). The x-axis shows CHST11 in the coordinates relative to the
transcription start site. The first exon and the genomic region we
sequenced after the bisulfite treatment (BS sequenced region) are
shown as indicated in the plots. (g) The expression of
CHST11 was measured in MCF7, T-47D, ZR-75-1 and MDA-MB-231
cells by qRT-PCR and normalized using 18S. Data were analyzed by
one-way ANOVA and post hoc analysis using data from three
independent experiments. Means, standard deviations, statistically
significant differences and P-values are shown. (h) Methylation
levels of the CpG island in five specimens from breast cancer
patients diagnosed with invasive ductal carcinoma. Methylation
analysis was performed and methylation levels were quantified as
described in legend to Fig. 2.
Percentages were averaged >20 CpGs and ≥2 replications. Means
and SEM are shown. Data were Arcsin transformed and subjected to
one-way ANOVA with Tukey’s post hoc analysis to compare
means. Specimen 1 is ER-positive and the other four specimens are
triple negative. Specimen 1 displayed a significantly higher
methylation level than the other specimens (P≤0.001). |
| Table IIISummary of CHST11 gene
expression and methylation status in human breast cancer cell lines
tested. |
Table III
Summary of CHST11 gene
expression and methylation status in human breast cancer cell lines
tested.
| Cell line | ER status | Molecular
subtype | CHST11
(qRT-PCR) | CS-A expression
(2H6 mAb binding by flow cytometry) | DNA methylation
status |
|---|
| MCF7 | Positive | Luminal | Low | Lowb |
Hypermethylated |
| T47-D | Positive | Luminal | Low | Lowc |
Hypermethylated |
| ZR-75-1 | Positive | Luminal | Low | Lowc |
Hypermethylated |
| BT-474 | Positive | Her-2
amplified | Lowa | ND |
Hypermethylated |
| MDA-MB-468 | Negative | Basal-like | Intermediate | Intermediate to
highb | Hypomethylated |
| BT-20 | Negative | Basal-like |
Intermediatea | ND | Hypomethylated |
| MDA-MB-231 | Negative | Basal-like | High | Highb | Hypomethylated |
| MDA-MET | Negative | Basal-like | High | Highb | Hypomethylated |
To determine whether the same CpG island can be
methylated differently in actual breast cancer, we examined the
methylation status of the CpG island of CHST11 gene in 5
de-identified clinical breast cancer specimens. We observed that in
4 triple negative (TN, i.e., negative for ER, Her2/neu and
progesterone receptor) specimens the CpG island was highly
hypomethylated, very similar to what was observed for ER-negative
basal-like cell lines (Fig. 4h).
The CpG island was hypermethylated in one ER-positive specimen
tested. Therefore, methylation status of CHST11 CpG might be
a useful marker to differentiate between luminal and basal-like
breast cancer subtypes, however, a larger sample size is required
to confirm this.
Discussion
We have previously shown that removal or blocking of
CS on the surface of breast cancer cells inhibits metastasis
(5,14). We have demonstrated that the
expression levels of CHST11 correlate with the expression of
P-selectin-reactive surface CS and aggressiveness of human breast
cancer cells (14). Here, we
confirm that the CHST11 gene is overexpressed in cancer
tissues compared to normal breast tissues and its expression
correlates with more aggressive basal-like and HER2-amplified
phenotypes in cell lines. The expression analysis of CHST11
in DCIS and IDC specimens suggest a progression specific role for
this gene that needs to be further validated. An association
between CHST11 expression and distant metastasis in breast
cancer patients has been reported (6). The data suggest that the expression
of CHST11 correlates well with properties associated with
prognosis and progression and should be investigated as a potential
biomarker.
Our data suggest that the expression of
CHST11 is controlled by DNA methylation. Aberrant DNA
methylation is involved in tumor initiation and progression
(22,23,38).
Hypermethylation of tumor suppressor genes is a common mechanism of
gene silencing observed in cancer, and similarly, DNA
hypomethylation can contribute to overexpression of tumor-promoting
genes. DNA hypomethylation is associated with advanced stages,
metastatic phenotypes, and drug-resistant variants of breast cancer
(39–41). We find that induced hypomethylation
by 5Azadc led to CHST11 overexpression and increased levels
of the CHST11’s immediate cell surface product CS-A. Based
upon our results, it seems that a combination of hypomethylation
and high expression occurs in basal-like cancer cells.
Hypermethylation of CHST11 (and very low to no expression)
clearly differentiates the least aggressive, luminal cells with
epithelial morphology from the more aggressive cells we tested. We
observed a hypomethylated CpG island in a limited number of triple
negative specimens from patients. Triple-negative cancers are
usually classified as basal-like while ER-positive cells are
considered luminal. The data suggest that methylation analysis of
this CpG island of CHST11 might be potentially used as a
surrogate for detection of expression levels of this gene in
clinical samples. However, more experiments are needed to evaluate
the correlation of the expression of this gene with its DNA
methylation levels among subtypes of breast cancer.
Our study illustrates an example of a gene, where
methylation is lower and expression is higher as the cancer
phenotype becomes more aggressive. In contrast, the main trend is
widespread gene hypermethylation as cancers go from less aggressive
to more aggressive phenotypes (42,43).
Our data are consistent with some other models of metastatic and/or
more aggressive cancers exemplifying tumor promoting genes like
urokinase plasminogen activator (uPA), PAX3 and Ezrin that are less
methylated and/or more highly expressed in the more aggressive
cancer forms (40,44,45).
This less frequent pattern of gene methylation between cancers with
low and high aggressiveness suggests that the expression of these
genes is necessary for the more aggressive phenotypes (because
their change in gene specific methylation is counter to the trend
of gene hypermethylation in cancer).
It needs to be pointed out that in less aggressive
basal B cells that are epithelial-like (e.g., MDA-MB-468),
hypomethylation is accompanied by intermediate expression of
CHST11, suggesting involvement of other mechanisms in
controlling the expression levels of this gene such as TGFβ1
(46,47). As we suggested before (14), in order for CHST11 activity to
express the proper receptor it might need to be co-expressed with a
specific proteoglycan. Future studies on well-characterized
clinical specimens are needed to determine a role for the
CHST11 gene alone or in combination with other genes in
patient outcome. More studies are also needed to determine whether
DNA methylation status can replace gene expression for
prognosis.
The recognition that silencing of tumor suppressor
genes through promoter hypermethylation plays a significant role in
tumorigenesis (48,49) has led to the clinical use of
hypomethylating agents including 5AzadC (50). However, the expression of several
pro-tumor genes is induced by DNA hypomethylation (40,45).
The expression of such genes, including CHST11, may be
activated by the clinical use of hypomethylating agents and this
may promote more aggressive forms of breast cancer. In this regard,
our data, in agreement with others (51), suggest that therapeutic use of such
demethylating agents may promote tumor progression and metastasis.
Histone deacetylase inhibitors can also hypomethylate genes and
change their expression levels in breast cancer cell lines
(52) and their combination with
metabolic therapies may modify their action (53). Therefore, additional studies are
needed to determine if some agents or mechanisms of hypomethylation
are more or less likely to promote tumor metastasis.
Our findings suggest that the expression of
CHST11 correlates with aggressive phenotypes and progression
of DCIS to IDC. We found out that the expression of CHST11
is modulated by DNA methylation. Therefore, DNA methylation plays
an important role in the remodeling of CS in breast tumors. Our
data strongly suggest that the expression of CHST11 and its
role in defining metastatic potential of tumor cells should be
seriously considered when demethylating agents are used for medical
treatment of breast cancer patients. Moreover, the methylation and
expression of the CHST11 gene have potential to be developed
into novel prognostic biomarkers.
Acknowledgements
This study was supported in part by a pilot project
grant to B.M.K. from the UAMS Translational Research Institute
through the Clinical and Translational Science Award 1UL1RR029884
from the National Center for Research Resources.
References
|
1
|
Masuda H, Ozeki T, Takazono I and Tanaka
Y: Composition of glycosaminoglycans in human pancreatic cancer.
Biochem Med Metab Biol. 41:193–200. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li F, Ten Dam GB, Murugan S, Yamada S,
Hashiguchi T, Mizumoto S, Oguri K, Okayama M, van Kuppevelt TH and
Sugahara K: Involvement of highly sulfated chondroitin sulfate in
the metastasis of the Lewis lung carcinoma cells. J Biol Chem.
283:34294–34304. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Svensson KJ, Christianson HC, Kucharzewska
P, Fagerstrom V, Lundstedt L, Borgquist S, Jirstrom K and Belting
M: Chondroitin sulfate expression predicts poor outcome in breast
cancer. Int J Oncol. 39:1421–1428. 2011.PubMed/NCBI
|
|
4
|
Iida J, Wilhelmson KL, Ng J, Lee P,
Morrison C, Tam E, Overall CM and McCarthy JB: Cell surface
chondroitin sulfate glycosaminoglycan in melanoma: role in the
activation of pro-MMP-2 (pro-gelatinase A). Biochem J. 403:553–563.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Monzavi-Karbassi B, Stanley JS, Hennings
L, Jousheghany F, Artaud C, Shaaf S and Kieber-Emmons T:
Chondroitin sulfate glycosaminoglycans as major P-selectin ligands
on metastatic breast cancer cell lines. Int J Cancer.
120:1179–1191. 2007. View Article : Google Scholar
|
|
6
|
Martinez P, Vergoten G, Colomb F, Bobowski
M, Steenackers A, Carpentier M, Allain F, Delannoy P and Julien S:
Over-sulfated glycosaminoglycans are alternative selectin ligands:
insights into molecular interactions and possible role in breast
cancer metastasis. Clin Exp Metastasis. 30:919–931. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vallen MJ, van der Steen SC, van Tilborg
AA, Massuger LF and van Kuppevelt TH: Sulfated sugars in the
extracellular matrix orchestrate ovarian cancer development: ‘when
sweet turns sour’. Gynecol Oncol. Aug 23–2014.(Epub ahead of
print). pii: S0090-8258(14)01276-1. View Article : Google Scholar
|
|
8
|
Sugahara K, Mikami T, Uyama T, Mizuguchi
S, Nomura K and Kitagawa H: Recent advances in the structural
biology of chondroitin sulfate and dermatan sulfate. Curr Opin
Struct Biol. 13:612–620. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kusche-Gullberg M and Kjellen L:
Sulfotransferases in glycosaminoglycan biosynthesis. Curr Opin
Struct Biol. 13:605–611. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Afratis N, Gialeli C, Nikitovic D,
Tsegenidis T, Karousou E, Theocharis AD, Pavao MS, Tzanakakis GN
and Karamanos NK: Glycosaminoglycans: key players in cancer cell
biology and treatment. FEBS J. 279:1177–1197. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mikami T and Kitagawa H: Biosynthesis and
function of chondroitin sulfate. Biochim Biophys Acta.
1830.4719–4733. 2013.
|
|
12
|
Mikami T, Mizumoto S, Kago N, Kitagawa H
and Sugahara K: Specificities of three distinct human
chondroitin/dermatan N-acetylgalactosamine 4-O-sulfotransferases
demonstrated using partially desulfated dermatan sulfate as an
acceptor: implication of differential roles in dermatan sulfate
biosynthesis. J Biol Chem. 278:36115–36127. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Uyama T, Ishida M, Izumikawa T, Trybala E,
Tufaro F, Bergstrom T, Sugahara K and Kitagawa H: Chondroitin
4-O-sulfotransferase-1 regulates E disaccharide expression of
chondroitin sulfate required for herpes simplex virus infectivity.
J Biol Chem. 281:38668–38674. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cooney CA, Jousheghany F, Yao-Borengasser
A, Phanavanh B, Gomes T, Kieber-Emmons AM, Siegel ER, Suva LJ,
Ferrone S, Kieber-Emmons T and Monzavi-Karbassi B: Chondroitin
sulfates play a major role in breast cancer metastasis: a role for
CSPG4 and CHST11 gene expression in forming surface P-selectin
ligands in aggressive breast cancer cells. Breast Cancer Res.
13:R582011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim YJ, Borsig L, Varki NM and Varki A:
P-selectin deficiency attenuates tumor growth and metastasis. Proc
Natl Acad Sci USA. 95:9325–9330. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Borsig L, Wong R, Feramisco J, Nadeau DR,
Varki NM and Varki A: Heparin and cancer revisited: mechanistic
connections involving platelets, P-selectin, carcinoma mucins, and
tumor metastasis. Proc Natl Acad Sci USA. 98:3352–3357. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Garcia J, Callewaert N and Borsig L:
P-selectin mediates metastatic progression through binding to
sulfatides on tumor cells. Glycobiology. 17:185–196. 2007.
View Article : Google Scholar
|
|
18
|
Stubke K, Wicklein D, Herich L, Schumacher
U and Nehmann N: Selectin-deficiency reduces the number of
spontaneous metastases in a xenograft model of human breast cancer.
Cancer Lett. 321:89–99. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Narayan A, Ji W, Zhang XY, Marrogi A,
Graff JR, Baylin SB and Ehrlich M: Hypomethylation of
pericentromeric DNA in breast adenocarcinomas. Int J Cancer.
77:833–838. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Szyf M, Pakneshan P and Rabbani SA: DNA
methylation and breast cancer. Biochem Pharmacol. 68:1187–1197.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ehrlich M: DNA methylation in cancer: too
much, but also too little. Oncogene. 21:5400–5413. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bardowell SA, Parker J, Fan C, Crandell J,
Perou CM and Swift-Scanlan T: Differential methylation relative to
breast cancer subtype and matched normal tissue reveals distinct
patterns. Breast Cancer Res Treat. 142:365–380. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Weisenberger DJ: Characterizing DNA
methylation alterations from The Cancer Genome Atlas. J Clin
Invest. 124:17–23. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kalathas D, Triantaphyllidou IE,
Mastronikolis NS, Goumas PD, Papadas TA, Tsiropoulos G and Vynios
DH: The chondroitin/dermatan sulfate synthesizing and modifying
enzymes in laryngeal cancer: expressional and epigenetic studies.
Head Neck Oncol. 2:272010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun Z, Asmann YW, Kalari KR, Bot B,
Eckel-Passow JE, Baker TR, Carr JM, Khrebtukova I, Luo S, Zhang L,
Schroth GP, Perez EA and Thompson EA: Integrated analysis of gene
expression, CpG island methylation, and gene copy number in breast
cancer cells by deep sequencing. PLoS One. 6:e174902011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Richardson AL, Wang ZC, De Nicolo A, Lu X,
Brown M, Miron A, Liao X, Iglehart JD, Livingston DM and Ganesan S:
X chromosomal abnormalities in basal-like human breast cancer.
Cancer Cell. 9:121–132. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Finak G, Bertos N, Pepin F, Sadekova S,
Souleimanova M, Zhao H, Chen H, Omeroglu G, Meterissian S, Omeroglu
A, Hallett M and Park M: Stromal gene expression predicts clinical
outcome in breast cancer. Nat Med. 14:518–527. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gluck S, Ross JS, Royce M, McKenna EF Jr,
Perou CM, Avisar E and Wu L: TP53 genomics predict higher clinical
and pathologic tumor response in operable early-stage breast cancer
treated with docetaxel-capecitabine ± trastuzumab. Breast Cancer
Res Treat. 132:781–791. 2012. View Article : Google Scholar
|
|
29
|
Hoeflich KP, O’Brien C, Boyd Z, Cavet G,
Guerrero S, Jung K, Januario T, Savage H, Punnoose E, Truong T,
Zhou W, Berry L, Murray L, Amler L, Belvin M, Friedman LS and
Lackner MR: In vivo antitumor activity of MEK and
phosphatidylinositol 3-kinase inhibitors in basal-like breast
cancer models. Clin Cancer Res. 15:4649–4664. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schuetz CS, Bonin M, Clare SE, Nieselt K,
Sotlar K, Walter M, Fehm T, Solomayer E, Riess O, Wallwiener D,
Kurek R and Neubauer HJ: Progression-specific genes identified by
expression profiling of matched ductal carcinomas in situ and
invasive breast tumors, combining laser capture microdissection and
oligonucleotide microarray analysis. Cancer Res. 66:5278–5286.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
ONCOMINE: a cancer microarray database and integrated data-mining
platform. Neoplasia. 6:1–6. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
33
|
Leakey TI, Zielinski J, Siegfried RN,
Siegel ER, Fan CY and Cooney CA: A simple algorithm for quantifying
DNA methylation levels on multiple independent CpG sites in
bisulfite genomic sequencing electropherograms. Nucleic Acids Res.
36:e642008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li LC and Dahiya R: MethPrimer: designing
primers for methylation PCRs. Bioinformatics. 18:1427–1431. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Takai D and Jones PA: The CpG island
searcher: a new WWW resource. In Silico Biol. 3:235–240.
2003.PubMed/NCBI
|
|
36
|
Meissner A, Mikkelsen TS, Gu H, Wernig M,
Hanna J, Sivachenko A, Zhang X, Bernstein BE, Nusbaum C, Jaffe DB,
Gnirke A, Jaenisch R and Lander ES: Genome-scale DNA methylation
maps of pluripotent and differentiated cells. Nature. 454:766–770.
2008.PubMed/NCBI
|
|
37
|
Smith ZD, Gu H, Bock C, Gnirke A and
Meissner A: High-throughput bisulfite sequencing in mammalian
genomes. Methods. 48:226–232. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Feinberg AP, Gehrke CW, Kuo KC and Ehrlich
M: Reduced genomic 5-methylcytosine content in human colonic
neoplasia. Cancer Res. 48:1159–1161. 1988.PubMed/NCBI
|
|
39
|
Soares J, Pinto AE, Cunha CV, Andre S,
Barao I, Sousa JM and Cravo M: Global DNA hypomethylation in breast
carcinoma: correlation with prognostic factors and tumor
progression. Cancer. 85:112–118. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pakneshan P, Szyf M, Farias-Eisner R and
Rabbani SA: Reversal of the hypomethylation status of urokinase
(uPA) promoter blocks breast cancer growth and metastasis. J Biol
Chem. 279:31735–31744. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chekhun VF, Kulik GI, Yurchenko OV,
Tryndyak VP, Todor IN, Luniv LS, Tregubova NA, Pryzimirska TV,
Montgomery B, Rusetskaya NV and Pogribny IP: Role of DNA
hypomethylation in the development of the resistance to doxorubicin
in human MCF-7 breast adenocarcinoma cells. Cancer Lett. 231:87–93.
2006. View Article : Google Scholar
|
|
42
|
Andrews J, Kennette W, Pilon J, Hodgson A,
Tuck AB, Chambers AF and Rodenhiser DI: Multi-platform whole-genome
microarray analyses refine the epigenetic signature of breast
cancer metastasis with gene expression and copy number. PLoS One.
5:e86652010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wolff EM, Chihara Y, Pan F, Weisenberger
DJ, Siegmund KD, Sugano K, Kawashima K, Laird PW, Jones PA and
Liang G: Unique DNA methylation patterns distinguish noninvasive
and invasive urothelial cancers and establish an epigenetic field
defect in premalignant tissue. Cancer Res. 70:8169–8178. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kurmasheva RT, Peterson CA, Parham DM,
Chen B, McDonald RE and Cooney CA: Upstream CpG island methylation
of the PAX3 gene in human rhabdomyosarcomas. Pediatr Blood Cancer.
44:328–337. 2005. View Article : Google Scholar
|
|
45
|
Yu Y, Zeng P, Xiong J, Liu Z, Berger SL
and Merlino G: Epigenetic drugs can stimulate metastasis through
enhanced expression of the pro-metastatic Ezrin gene. PLoS One.
5:e127102010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kluppel M, Vallis KA and Wrana JL: A
high-throughput induction gene trap approach defines C4ST as a
target of BMP signaling. Mech Dev. 118:77–89. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Willis CM, Wrana JL and Kluppel M:
Identification and characterization of TGFbeta-dependent and
-independent cis-regulatory modules in the C4ST-1/CHST11 locus.
Genet Mol Res. 8:1331–1343. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
MacLeod AR, Rouleau J and Szyf M:
Regulation of DNA methylation by the Ras signaling pathway. J Biol
Chem. 270:11327–11337. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ramchandani S, MacLeod AR, Pinard M, von
Hofe E and Szyf M: Inhibition of tumorigenesis by a cytosine-DNA,
methyltransferase, antisense oligodeoxynucleotide. Proc Natl Acad
Sci USA. 94:684–689. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fenaux P: Inhibitors of DNA methylation:
beyond myelodysplastic syndromes. Nat Clin Pract Oncol. 2(Suppl 1):
S36–S44. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ateeq B, Unterberger A, Szyf M and Rabbani
SA: Pharmacological inhibition of DNA methylation induces
proinvasive and prometastatic genes in vitro and in vivo.
Neoplasia. 10:266–278. 2008.PubMed/NCBI
|
|
52
|
Meeran SM, Patel SN and Tollefsbol TO:
Sulforaphane causes epigenetic repression of hTERT expression in
human breast cancer cell lines. PLoS One. 5:e114572010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Cooney CA: Drugs and supplements that may
slow aging of the epigenome. Drug Discov Today Ther Strateg.
7:57–84. 2010. View Article : Google Scholar
|