Introduction
Hepatocellular carcinoma (HCC) ranks as the fifth
most common malignant disorder and the third leading cause of
cancer-related deaths worldwide (1). The most important causes leading to
HCC are the HBV and HCV infections, heavy alcohol consumption,
aflatoxin B1, gender (males are more susceptible than females),
obesity associated with non-alcoholic fatty liver disease, and
α1-antitrypsin deficiency (2).
Surgical resection, liver transplantation and ablative procedures
are considered the curative procedures especially for early-stage
HCC but also for these patients, bearing the most favorable
conditions, the final prognosis is not completely satisfactory
(3). The increasing understanding
of the tumor biology of HCC could be helpful for the development of
future targeted HCC therapeutics. The basic research community has
to pursue this objective to include into the clinical practice some
new molecular markers reflecting the biological aggressiveness of
HCC.
LIM and SH3 protein 1 (LASP-1) was initially
identified from a cDNA library of metastatic axillary lymph nodes
of breast cancer patients. The human gene is located on 17q21
chromosome, it encodes a protein of 261 amino acids containing an
N-terminal LIM domain followed by two actin binding domains in the
core of the LASP-1 protein mediating an interaction between LASP-1
and the actin cytoskeleton (4,5). The
SH3 domain at the C-terminus is involved in protein-protein
interactions specifically with zyxin, pallidin, lipoma preferred
partner (LPP) and vasodilator-stimulated phosphoprotein (VASP). The
exact functions of LASP-1 are still not completely elucidated.
LASP-1 is localised in dynamic actin assembly such as focal
contacts, focal adhesions, lamellipodia membrane ruffles and
pseudopodia where it interacts with motility-associated proteins
and functions as structural scaffold (6–8).
LASP-1 is overexpressed in several human
malignancies included human metastatic breast cancer, ovarian
cancer, colorectal cancer, malignant childhood medulloblastoma,
hepatocellular carcinoma, bladder and oral cancer, and prostate
carcinoma (9–15). In vitro studies showed that
LASP-1 plays an important role in tumor development and metastases.
The knock-down of LASP-1 by RNA interference resulted in a strong
inhibition of the proliferation and migration of various cancer
cells, such as breast, ovarian, colorectal and prostate cancer cell
lines (9,15,16).
In certain types of malignant cells the nuclear localization of the
protein was observed. LASP-1 expression and nuclear localization
correlated significantly with tumor size, nodal positivity and a
poor long-term survival of the patients affected by breast cancer
(17–19). In a previous study, we found that
LASP-1 is a downstream protein of the urokinase type plasminogen
activator (uPA) and its mediator in HCC cell migration likely
taking part in the cytoskeleton changes that occur during this
process (20). Ectopic uPA
overexpression induced LASP-1 upregulation and cell motility in HCC
cells. However, ectopic LASP-1 overexpression did not upregulate
uPA expression. In the present study, we investigated the
biological role of LASP-1 in HCC by a molecular and biological
characterization of LASP-1 expression in human HCC specimens and in
cultured HCC cells. We ascertained the heterogenous expression
level of LASP-1 mRNA in HCC with different hepatic background
disease and we have biologically characterized the ectopic LASP-1
overexpression in HCC cells.
Materials and methods
Cell cultures
SKHep1Clone3 (SKHep1C3), SKHep1C3 nod.69.2, selected
from human HCC-derived cells (SKHep1: ATCC HTB-52), AB15 and AB19
human dermal fibroblasts were maintained in Earle’s MEM (Life
Technologies, Carlsbad, CA, USA) supplemented with 10% foetal
bovine serum (Life Technologies) at 37°C in a 5% CO2
incubator. Differentiated human HCC-derived cells (HepG2, ATTC
HB-8065; HuH-6; HuH7) and HA22T/VGH undifferentiated HCC-derived
cells were maintained in RPMI-1640 (Life Technologies) supplemented
with 10% foetal bovine serum and 1 mM sodium pyruvate at 37°C in a
5% CO2 incubator. The HuH-6 and HA22T/VGH cells were
kindly provided by N. D’Alessandro (University of Palermo,
Italy).
Tissues and clinicopathological features
of HCC and evaluation of LASP-1 expression in tumoral and
peri-tumoral (PT) human tissues by qPCR
All human HCC samples (n=55) as well as the
corresponding PT non-tumor samples (resected 1–2 cm from the
malignant tumor) were obtained from HCC patients for pathological
examination. Each biopsy specimen was obtained with the patient’s
informed consent under standard conditions of sampling and
processing (40). Each specimen
was determined to be HCC or PT by pathological examination. In this
study, 55 HCC subjects underwent surgical resection. The subjects
consisted of 35 men and 20 women (54 Italian and 1 Chinese) ranging
from 38 to 82 years of age (mean age: 68.7±8.4 years). The subjects
did not have any apparent distant metastases, and none had been
previously treated for HCC. We subdivided the cases on the basis of
presence or absence of liver cirrhosis (31 HCC with cirrhosis, 24
HCC without cirrhosis). Twenty-three patients were HCV positive, 12
were HBV positive, 3 were both HBV and HCV positive, and 16 were
both HBV and HCV negative, for 1 patient no information was
available (Table I). The total RNA
from tissue samples was isolated using TRIzol reagent (Invitrogen),
according to the manufacturer’s instructions. The expression of
LASP-1 mRNAs in the tissues was evaluated using TaqMan Gene
Expression Assay (Applied Biosystem). GAPDH was used as an internal
standard.
 | Table IClinical and pathological
characteristics of the studied population. |
Table I
Clinical and pathological
characteristics of the studied population.
| Case | Gender | Years | Grading | TNM | Background
disease | HBV | HCV |
|---|
| LV 136 | F | 79 | G2 | NA | Cirrhosis with
active chronic hepatitis | − | + |
| LV 185 | M | 66 | G1 | T1N0M0 | Chronic
hepatitis | + | − |
| LV 197 | M | 70 | G2 | T1N0M0 | Cirrhosis with
micro- and macrovescicular steatosis | − | − |
| LV 204 | M | 74 | G3 | NA | Active
cirrhosis | − | + |
| LV 216 | F | 67 | G1 | NA | Cirrhosis | − | − |
| LV 218 | M | 64 | G2 | NA | Cirrhosis with
active chronic hepatitis | NA | NA |
| LV 219 | M | 57 | G1 | T1N0M0 | Cirrhosis with
active chronic hepatitis | + | − |
| LV 224 | M | 55 | G3 | T3bN0M0 | Cirrhosis with
active chronic hepatitis | + | + |
| LV 225 | M | 49 | G3 | T3bN0M0 | Microvescicular
steatosis | − | − |
| LV 227 | F | 72 | G2/G3 | T1N0M0 | Cirrhosis with
active chronic hepatitis | − | + |
| LV 228 | M | 59 | G2 | T1N0M0 | Active chronic
hepatitis of severe level with necrosis and bridging porto-portal
fibrosis (HBsAg) | + | − |
| LV 229 | F | 79 | G2/G3 | T3bN0M0 | Cirrhosis with
active chronic hepatitis | − | − |
| LV 232 | M | 76 | NA | NA | Cirrhosis with
chronic hepatitis | − | + |
| LV 235 | F | 82 | G3 | T2N0M0 | Cirrhosis with
active chronic hepatitis | − | + |
| LV 236 | F | 76 | G1 | T1N0M0 | Cirrhosis with
active chronic hepatitis | − | + |
| LV 237 | M | 68 | G2/G3 | T1N0M0 | Mildly active
chronic hepatitis | − | + |
| LV 240 | M | 71 | G3 | T3bN0M0 | Active chronic
hepatitis with necrosis and bridging and portal fibrosis | + | − |
| LV 241 | F | 38 | G2 | T3N0M0 | Reactive
hepatitis | + | − |
| LV 242 | F | 63 | G2 | T2N0M0 | Active chronic
hepatitis with focal fibrosis and with bridging porto-portal
fibrosis | − | + |
| LV 262 | M | 73 | G2 | NA | Mild hepatitis | − | − |
| LV 268 | F | 68 | G1 | T1N0M0 | Cirrhosis with
active chronic hepatitis | + | − |
| LV 273 | M | 73 | G2 | T1N0M0 | Cirrhosis with
active chronic hepatitis and mild macro- and microvescicular
steatosis | − | + |
| LV 274 | F | 81 | G2 | T1N0M0 | Mildly active
chronic hepatitis with micro- and macrovescicular steatosis (30% of
parenchyma) | − | − |
| LV 276 | M | 72 | G2 | T1N0M0 | Cirrhosis with
active chronic hepatitis | − | + |
| LV 277 | F | 75 | G2 | NA | Chronic
hepatitis | − | − |
| LV 278 | M | 72 | G2 | NA | Cirrhosis | − | + |
| LV 279 | M | 70 | NA | NA | Chronic
hepatitis | − | + |
| LV 280 | F | 74 | G2/G3 | T1N0M0 | Cirrhosis with
active chronic hepatitis | − | + |
| LV 281 | M | 74 | G1 | NA | Normal
parenchyma | − | − |
| LV 283 | M | 78 | G2 | T1N0M0 | Mildly active
chronic hepatitis | + | − |
| LV 284 | M | 76 | G2 | T1N0M0 | Active chronic
hepatitis | + | − |
| LV 285 | M | 77 | G2 | T2N0M0 | Active chronic
hepatitis with moderate/severe necrosis | − | + |
| LV 286 | M | 69 | G3 | T4N0M0 | Active
cirrhosis | − | + |
| LV 287 | M | 63 | G2 | T2N0M0 | Active
cirrhosis | − | − |
| LV 288 | F | 64 | G2 | T1N0M0 | Active
cirrhosis | − | − |
| LV 290 | M | 65 | G2/G3 | T1N0M0 | Active
cirrhosis | + | − |
| LV 291 | M | 69 | G1 | NA | Hepatitis | + | + |
| LV 292 | M | 66 | G3 | NA | Active
cirrhosis | − | + |
| LV 293 | M | 61 | G3 | NA | Active
cirrhosis | + | + |
| LV 294 | M | 61 | G2 | NA | Hepatitis | − | − |
| LV 295 | M | 72 | G3 | NA | Active
cirrhosis | − | − |
| LV 296 | M | 67 | G3 | NA | Active
cirrhosis | + | − |
| LV 297 | M | 73 | G3/G4 | NA | Hepatitis | + | − |
| LV 302 | F | 61 | G2 | NA | Cirrhosis | − | − |
| LV 303 | M | 76 | G3/G4 | NA | Cirrhosis | − | − |
| LV 304 | M | 70 | G2 | NA | Cirrhosis | − | + |
| LV 305 | F | 76 | G3 | NA | Hepatitis | − | + |
| LV 306 | M | 48 | G3 | NA | Hepatitis | − | + |
| LV 307 | M | 64 | G2 | NA | Steatosis | − | − |
| LV 308 | M | 73 | G2 | NA | Active
cirrhosis | − | − |
| LV 310 | F | 71 | G1 | NA | Active
cirrhosis | − | + |
| LV 317 | F | 69 | G1 | NA | Cirrhosis | + | − |
| LV 318 | F | 71 | G2 | NA | Cirrhosis with
ECA | − | + |
| LV 323 | F | 78 | G3 | NA | Hepatitis | − | + |
| LV 325 | F | 65 | G3 | NA | Chronic hepatitis
with ECA | − | + |
The PCR mixture (25 μl) containing 1 μl of the
specific probe, 11.25 μl of cDNA and 13.75 μl of Taq-Man 2X
Universal PCR Master Mix were incubated in a 7500 Applied
Biosystems instrument initially at 55°C for 2 min, then at 95°C for
10 min, followed by 40 cycles of 95°C for 15 sec and 60°C for 60
sec. The expression of LASP-1 mRNAs (RQ) was based on the
ΔΔCT method.
For each case the ratio (R) between the relative
levels in HCC (RQHCC) and those in PT (RQPT)
was calculated. The mRNA expression level was considered to be
decreased for a R-value ≤0.8 and increased for a R-value ≥1.2. A
value between 0.8 and 1.2 was defined as having no change in
expression level.
Immunohistochemical analysis
Tissue sections (5 μm) were de-paraffinized in
xylene, rehydrated in ethanol, incubated in 0.3%
H2O2 in methanol for 20 min to block
endogenous peroxidase activity; 3% BSA was used to block
non-specific staining. The sections were washed with 1X PBS and
incubated with rabbit anti-human LASP-1 (1:50 v/v) overnight (Santa
Cruz Biotechnology, Biosource, CA, USA). The biotinylated secondary
antibodies were added for 15 min (Super Sensitive IHC Detection
Systems, BioGenex, San Ramon, CA, USA). After extensive washing,
the sections were incubated with horseradish peroxidase complex
(ABC complex) for 15 min. The chromogen DAB was used to localize
the peroxidase in tissues. The slides were counterstained with
H&E and analyzed with an optical microscope at ×10, ×40 and ×60
magnification.
Lasp-1 cloning and transient transfection
of HA22T/VGH with pGPF-LASP1
The full lenght coding region (CDS) of LASP-1 was
amplified from SKHep1C3 cDNA using the LASP-1 cl Forward:
5′-ggaaccatgaaccccaac-3′ and SH3 Reverse: 5′-cctccacgtagttggccg-3′
primers and then directly cloned in the pcDNA3.1-CT-GFP-TOPO vector
(Life Technologies) upstream the GFP gene following manufacturer’s
instructions. The correct sequence and the orientation of the
insert were ascertained by direct automatic sequencing of the
plasmid. The vector, named pGFP-LASP1, was transiently transfected
in HA22T/VGH cells. Briefly, 2,900,000 HA22T/VGH cells were seeded
in 10-cm diameter Petri dishes and 24 h later, when the cells
reached 80% confluency, they were transfected with 24 μg/dish of
pGFP-LASP1 plasmid using 60 μl/dish of Lipofectamine (Life
Technologies) following the manufacturer’s instructions. After 72 h
of transfection, cell lysates were collected for
immunoprecipitation analysis in NP-40 buffer (Life Technologies)
containing phosphatase (Sigma) and protease (Roche) inhibitor
cocktails or in 0.05% SDS for routine analysis by western blotting
(WB).
LASP-1-GFP immunoprecipitation
Protein G Dynabeads® (50 μl) (Life
Technologies) per sample were incubated overnight at 4°C with 10 μg
of mouse monoclonal anti-human LASP-1 (Chemicon International) or
mouse monoclonal anti-human vimentin (Santa Cruz Biotechnology) or
mouse IgG1 (negative control). The day after the Ab-Dynabead
complexes were incubated overnight with cell extracts from
pGFP-LASP1 transfected HA22T/VGH or HA22T/VGH control cell extracts
in NP-40 buffer (1 ml, containing ~1 mg of total proteins). The day
after, the Ag-Ab-Dynabead complexes were placed on the magnet and
the supernatants were kept for further analysis (immunodepleted =
ID). After 3 washes of the Ag-Ab-Dynabeads with washing buffer, 30
μl of elution buffer per sample were added and the complex was
incubated at room temperature for 2 min. The Dynabeads were
separated on the magnet and the supernatants containing the Ab-Ag
complex (IP CTRL, IP α-LASP-1) were placed in a clean tube for
proteomic analysis.
Proteomic identification of LASP-1
partner proteins
Prior to the mass spectrometry (MS) analysis and in
order to confirm that we had obtained precipitated proteins, we
analyzed aliquots of our samples on 4–12% gels by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and Western
blotting according to the manufacturer’s instructions.
The IP CTRL and IP α-LASP-1 and the corresponding
antibodies were loaded on one-dimensional 4–12% NuPAGE®
precast gels (Life Technologies). Proteins were visualized with
G250 Coomassie Blue (Bio-Rad) or with silver staining by standard
procedures. For protein profiling, protein bands were excised from
Coomassie-stained preparative gels and processed as previously
described (42). MALDI-TOF-MS was
carried out using a Voyager-DE STR (Applied Biosystems), equipped
with a nitrogen laser (337 nm). Monoisotopic peptide masses were
analyzed using the Aldente software http://www.expasy.org/tools/webcite. The input was
searched according to: Aldente, UniProtKB/SwissProt; predefined
taxon, Mammalia; Spectrometer internal error max, 25. Only proteins
identified in two separate experiments were considered.
Western blot analysis
The samples were electrophoresed in 4–12% Bis-Tris
gels at 100 V using MES running buffer. The proteins were
electrophoretically transferred to a nitrocellulose membrane at 100
V for 1.5 h. The membrane was blocked with 3% milk in PBS 1X at
37°C for 2 h. The blots were incubated with primary antibodies:
rabbit anti human LASP-1 polyclonal antibody, 1:500 in 0.3% BSA-PBS
(Millipore); rabbit anti-green fluorescent protein polyclonal
antibody, 1:100 in 0.3% BSA-PBS (Santa Cruz Biotechnology); mouse
anti-vimentin monoclonal antibody, 1:1000 in 0.3% BSA-PBS (Santa
Cruz Biotechnology) at room temperature overnight, washed three
times with PBS, then incubated with an
alkaline-phosphatase-conjugated secondary antibody (1:7500 in 0.3%
BSA-PBS) for 4 h at 37°C. The results of the immunoreaction were
detected with Nitroblue tetrazolium and bromochloroindolyl
phosphate (Promega).
Immunofluorescence and confocal
immunofluorescence analysis
For the immunofluorescence detection of GFP,
HA22T/VGH were seeded and cultured (75,000 cells/22×22 mm glass
coverslips in 3-cm diameter Petri dishes) in growth medium. After
24h the cells were transfected with pGFP-LASP1 as described above
and after 48h the cultures were fixed in cold methanol × 20 min at
4°C. The coverslips were directly mounted on glass slides in
mounting medium and photographed with a Leitz Fluorescence
microscope (magnification, ×63).
For the immunofluorescence detection of vimentin,
HA22T/VGH were seeded and cultured (75,000 cells/22×22 mm glass
coverslips in 3-cm diameter Petri dishes) in growth medium. After
24 h the cells were fixed in cold methanol ×20 min at 4°C, after
one washing in PBS (1×3 min) and one treatment with 0.3% BSA in PBS
(1×3 min) the cells were immunoreacted with the first antibody,
mouse monoclonal anti-vimentin (1:50 in 0.3% BSA) for 1 h at room
temperature and washed 3×5 min in PBS. The cells were then
immunoreacted with the secondary rhodamine-conjugated anti-mouse
IgG (1:100 in 0.3% BSA) (Calbiochem, San Diego, CA, USA) for 30 min
at room temperature. The coverslips were mounted on glass slides in
mounting medium and photographed with a Leitz fluorescence
microscope (magnification, ×63).
For the confocal microscopy, AB15 and HA22T/VGH
cells were grown, fixed and permeabilized as described for
immunofluorescence microscopy. The samples were incubated with
primary polyclonal antibody against LASP-1 and with primary
monoclonal antibody against VIM (1:100 in 0.3% BSA) for 30 min at
room temperature. After the last washing step, the unconjugated
primary antibodies were recognized with secondary antibodies (1:400
in 0.3% BSA) conjugated with Alexa-Fluor 555 and Alexa-Fluor 488
dyes (Life Technologies, Invitrogen) for 30 min at RT. DNA was
counterstained with DAPI (1:3000) (Calbiochem) for 10 min. The
coverslips were mounted on glass slides in SlowFade Gold antifade
reagent (Life Technologies, Invitrogen). The images were acquired
on Zeiss LSM 510 Meta confocal microscope (Carl Zeiss, Milan,
Italy).
Statistical analysis
Each experiment was carried out at least twice. The
histograms represent the mean values, and bars indicate standard
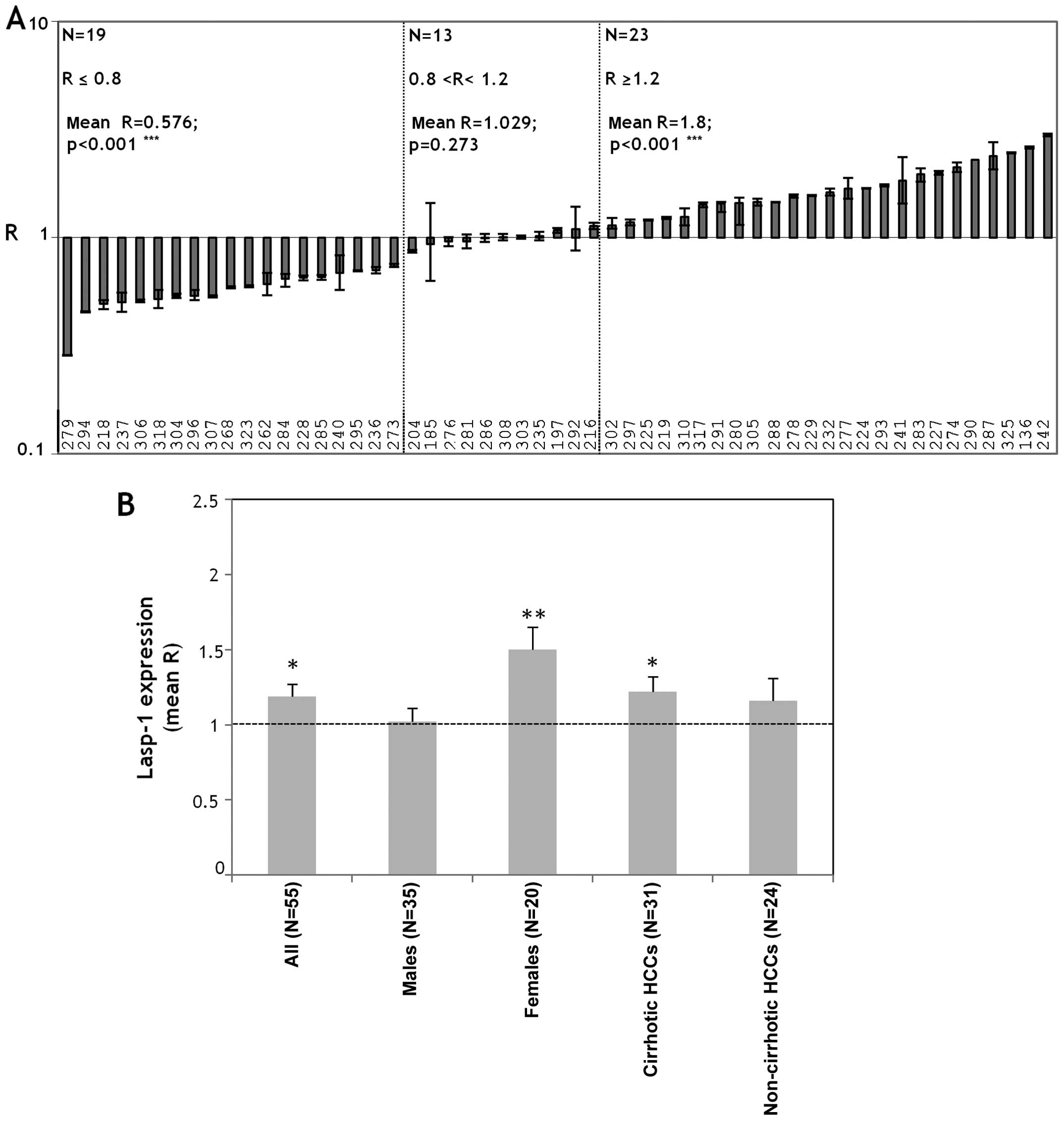
errors (SE) of the mean. For the data shown in Fig. 1 statistical significance of the
results was determined using Student’s t-test for single group mean
(expected value=1) and data were considered significant when
P≤0.05. Statistical analysis was performed with kyplot, version 2.0
beta 13 (http://www.woundedmoon.org/win32/kyplot.html).
Results
LASP-1 mRNA is significantly
overexpressed in HCC, particularly in female HCC patients and in
cirrhotic HCCs
The expression levels of LASP-1 mRNA were evaluated
by qPCR in 55 pairs of HCC and PT tissues. The R-value for each
sample was the ratio between the LASP-1 expression level in the HCC
tissue (RQHCC) and the expression level detected in the
relative non-tumor counterpart PT (RQPT). We arbitrary
assumed LASP-1 upregulation when R≥1.2, LASP-1 downregulation when
R≤0.8 and no variation when 0.8<R<1.2. So doing the cases
were subdivided into 3 groups: R≥1.2 (n=23); R≤0.8 (n=19);
0.8<R<1.2 (n=13) (Fig. 1A).
Thus, the first group was defined by higher LASP-1 mRNA levels in
HCC than PT (mean R=1.8; p<0.001); the second group was
characterized by lower LASP-1 mRNA levels in HCC than PT (mean
R=0.576, p<0.001); the third group was defined by equal levels
of LASP-1 mRNA in HCC and PT tissues (mean R=1.029). No clinic
parameter correlated with the LASP-1 mRNA expression (gender, age,
tumor grading, background hepatic disease, viral hepatitis
infection) in the 3 groups. Considering the mean LASP-1 expression
in all the HCCs and PTs (N=55) we evidenced increased levels in HCC
tissues compared to PTs (mean R=1.20, p<0.05) (Fig. 1B). The female HCC patients (N=20)
overexpressed LASP-1 mRNA in HCC tissues (mean R=1.5, p<0.01)
(Fig. 1B); the males (N=35) did
not show the disregulation of LASP-1 mRNA between HCC and PT
tissues (mean R=1.02) (Fig. 1B).
The statistical analysis performed by using the Mann-Whitney test
for unpaired data between mean R-values of males and females showed
a significant differential expression of LASP-1 mRNA in male and
female HCC patients (p=0.0147).
We further stratified the cases on the basis of the
presence (31/55) or absence (24/55) of cirrhosis as a background
liver disease. LASP-1 expression levels increased in HCCs compared
to PTs in the cirrhotic cases (mean R=1.22, p<0.05) (Fig. 1B). No change in LASP-1 expression
levels was observed in non-cirrhotic cases (mean R=1.15) (Fig. 1B).
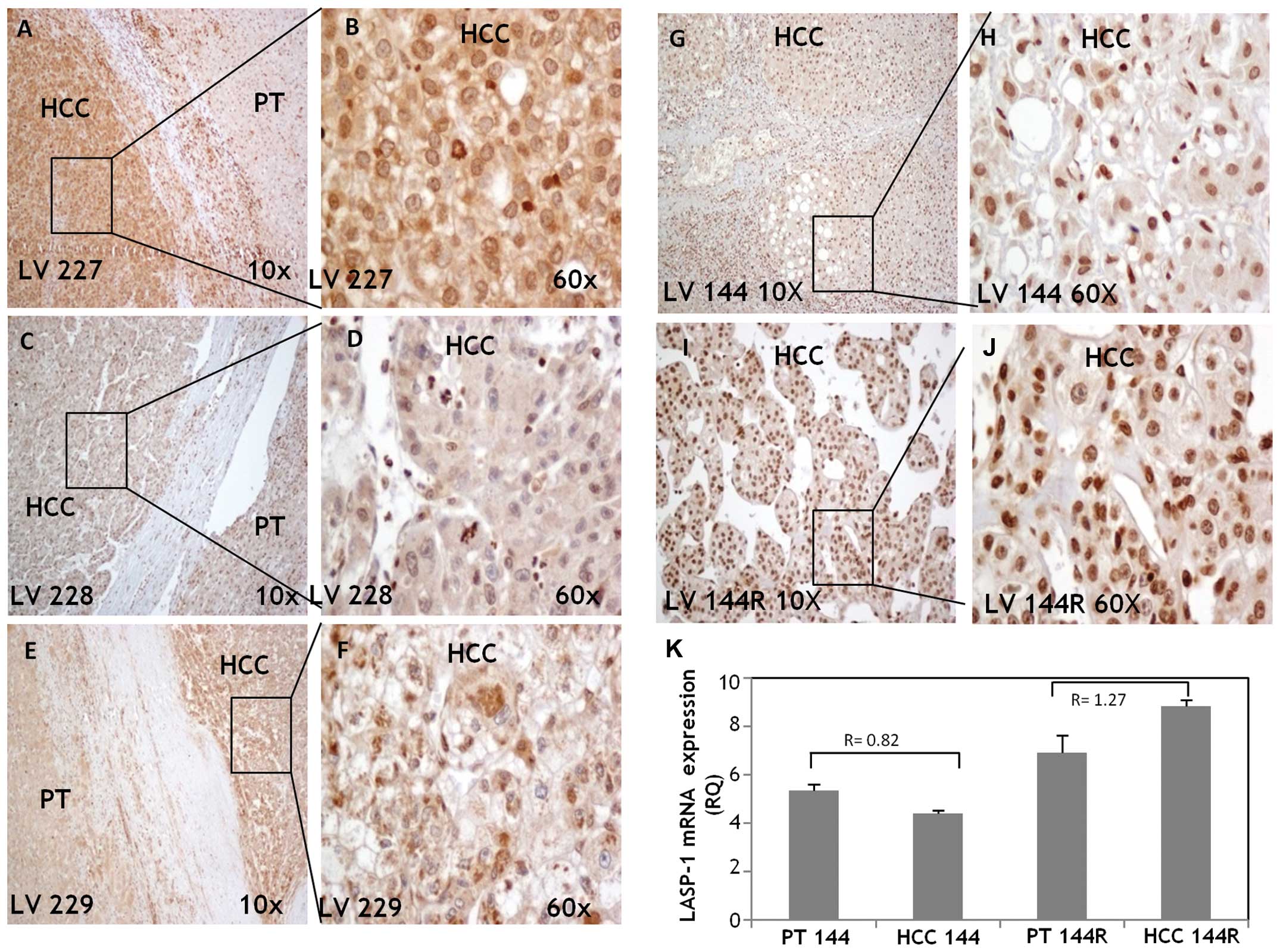
Levels of LASP-1 protein and mRNA
expression are comparable and they increase in recurrent HCC
To compare LASP-1 mRNA and protein expression levels
and to ascertain its cellular localization in some samples,
immunohistochemistry (IHC) was performed in paired HCC tumors and
their matched adjacent non-tumor tissues. We did not include all
the samples because, during our work, IHC evaluation of LASP-1
protein expression was assessed by other authors (12). The tissues analyzed here were
obtained from the biopsy specimens of 4 different HCC patients.
Three cases (LV 227, LV 228, LV 229) were selected among the 55
cases analyzed for LASP-1 mRNA expression, 1 case (LV 144) was
selected since the primary tumor and the intra-hepatic tumor
recurrence tissues were available (the intrahepatic recurrent HCC
was developed 28 months after the primary tumor resection). The
LASP-1 protein expression level reflected the LASP-1 mRNA levels in
the cases examined.
In LV 227 and LV 229 (Fig. 2A and B, E and F, respectively)
LASP-1 protein was greatly upregulated in HCC compared to PT with a
cytoplasmic staining and weak/moderate nuclear positivity. In LV
228 (Fig. 2C and D) LASP-1 was
essentially detectable in the inflammatory cells of the PT tissues
while in HCC LASP-1 was weakly expressed in the cytoplasm and
nuclei. LV 144 displayed moderate nuclear staining, particularly in
differentiated areas (Fig. 2G and
H); its intra-hepatic metastasis LV 144R (Fig. 2I and J) showed LASP-1 strong
nuclear staining and very weak cytoplasmic positivity. LASP-1 mRNA
levels were higher in PT 144R and HCC 144R than PT 144 and HCC 144
and the LASP-1 level in HCC 144R increased 2 fold compared to HCC
144 (RQHCC144R=8.821; RQHCC144=4.402)
(Fig. 2K).
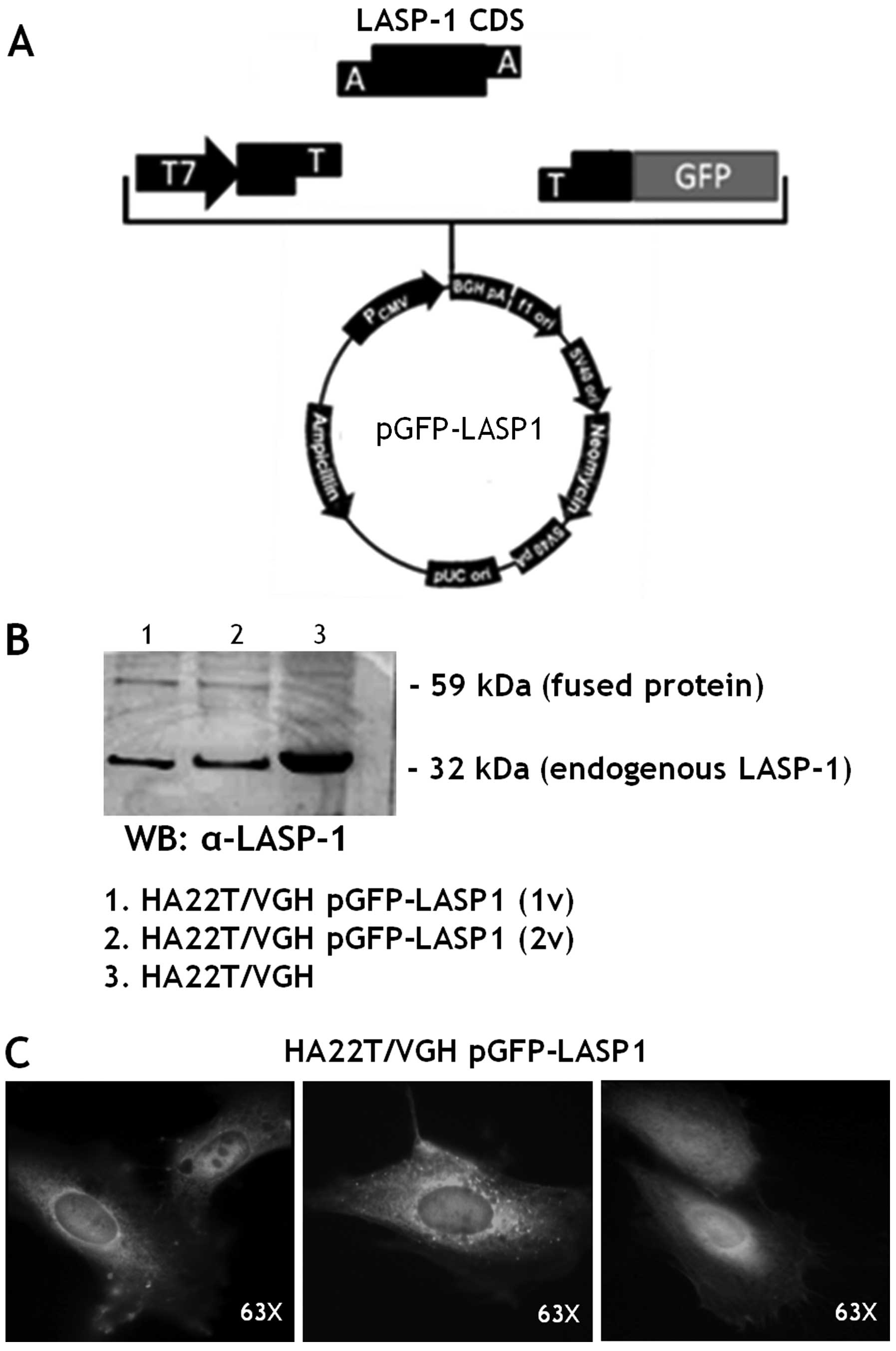
Immunoprecipitation of LASP-1-GFP fused
protein and MALDI-TOF-MS identification of its molecular
interactors in HA22T/VGH cells
LASP-1 CDS was cloned upstream the GFP coding gene
in the pCDNA3.1 expression vector (Fig. 3A). The plasmid, named pGFP-LASP1,
was sequenced and then transiently transfected in the HA22T/VGH
cell line (HA22T/VGH pGFP-LASP1).
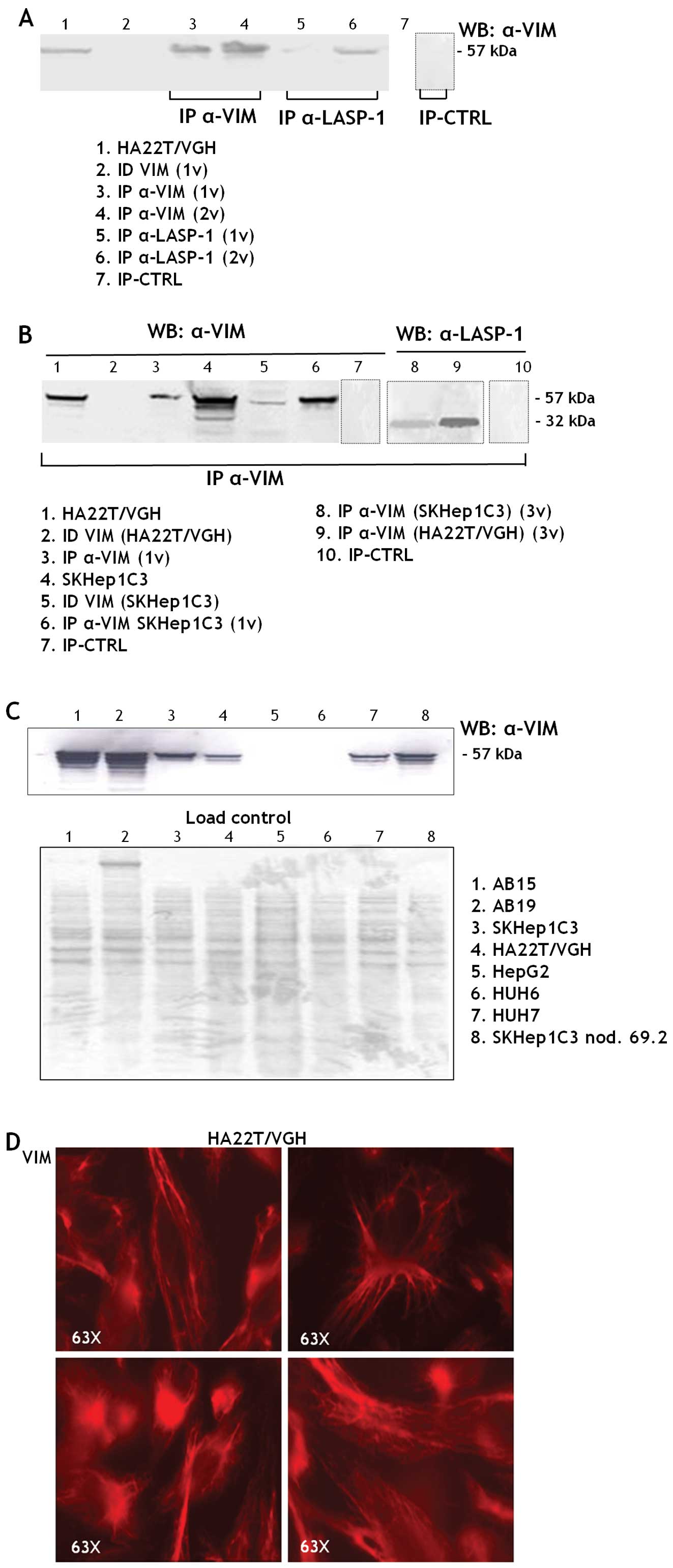
The fused protein (59 kDa) was detected in the
transfected cells while the endogenous LASP-1 (32 kDa) was detected
in transfected and untransfected HA22T/VGH cells (Fig. 3B) as expected. The fluorescent
fused protein LASP-1-GFP was directly observed under a fluorescence
microscope. It was mainly localized in the cytoplasm and in the
peri-nuclear area of the HA22T/VGH cells (Fig. 3C).
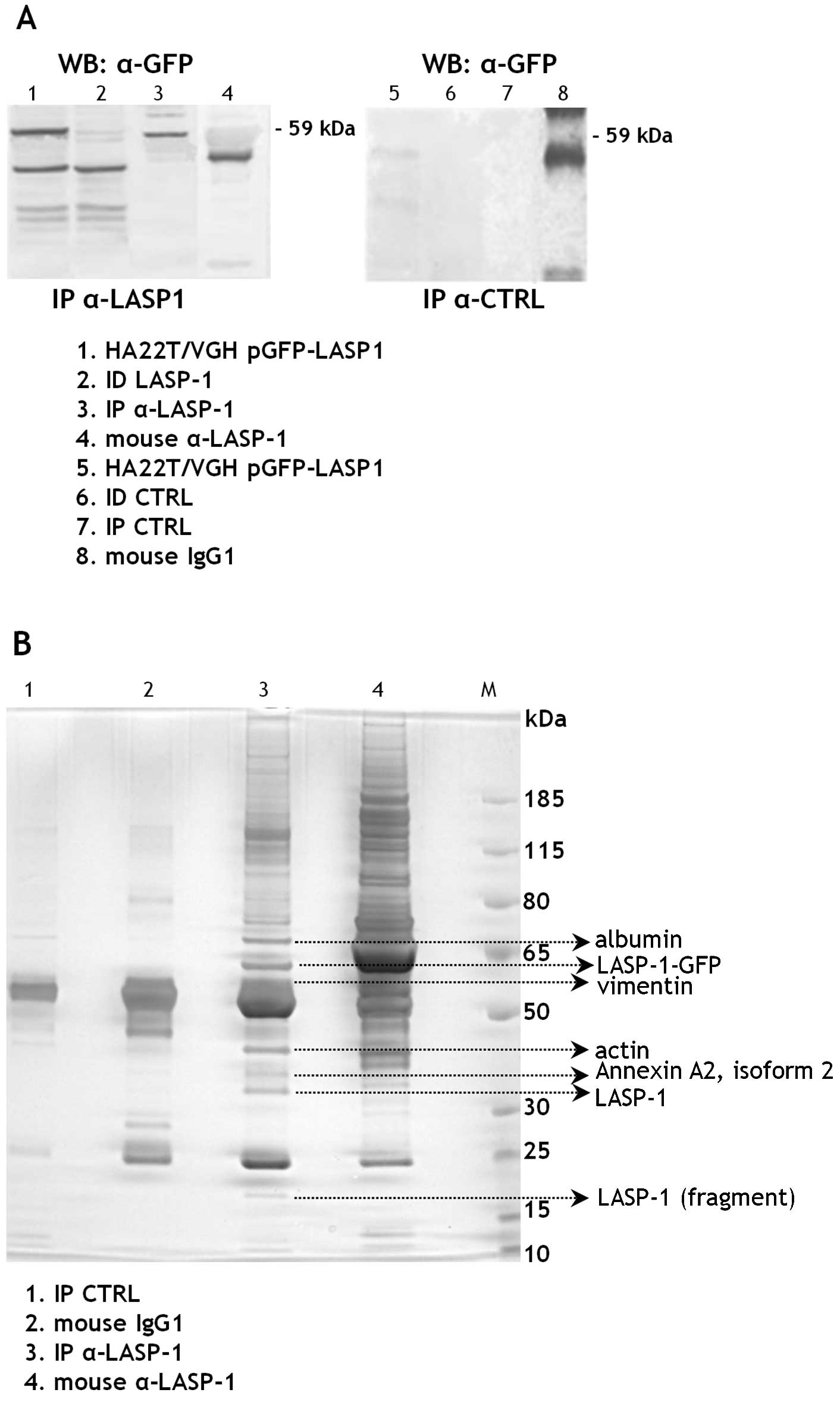
To identify new molecular partners of LASP-1 in HCC
cells we first immunoprecipitated the proteins from the cell
extracts of the transfected HA22T/VGH cells using the mouse
monoclonal anti-LASP-1 antibodies immobilized on magnetic beads.
The presence of the fused 59 kDa protein in the LASP-1
immunoprecipitated fraction (IP α-LASP1) was tested by WB with
anti-GFP antibodies (Fig. 4A). No
bands were detected in the control cell extracts immunoprecipitated
with the mouse IgG1 (IP CTRL) and this demonstrated the absence of
proteins aspecifically bound to the mouse antibody used or to the
magnetic beads.
The LASP-1 (IP α-LASP1), the control (IP CTRL)
immunoprecipitated fractions and the antibodies employed to
immunoprecipitate were separated on a bidimensional polyacrylamide
gel. The differential protein bands present in the IP α-LASP1
fraction were excised and analyzed with MALDI-TOF-MS. The
experiment was performed twice and vimentin (VIM) was detected in
IP α-LASP1 both times with statistically significant scores. The
same for actin, albumin and annexin (Fig. 4B).
To validate vimentin as a new molecular partner of
LASP-1 we immunoprecipitated the proteins from HA22T/VGH cell
extracts with anti-VIM and anti-LASP1 antibodies. The VIM protein
(57 kDa) was detected in the IP α-VIM and in the IP α-LASP1 of the
HA22T/VGH cells (Fig. 5A); the
LASP-1 protein was detected in the IP α-VIM of both HA22T/VGH and
SKHep1C3 cells (Fig. 5B, lanes 8
and 9); this demonstrates that VIM could be a molecular partner of
LASP-1 in both HCC cell lines. VIM, a member of the intermediate
filaments, is variably expressed in human HCC cells, in particular
at high levels in the undifferentiated HCC cells (HA22T/VGH,
SKHep1C3, SKHep1C3 nod. 69.2) and in fibroblasts (AB15 and AB19)
(Fig. 5C). Like other cells of
connective tissue, fibroblasts are derived from the primitive
mesenchyme. Thus, they express the intermediate filament protein
VIM. In more differentiated HCC cells VIM is not expressed (HepG2,
HuH6) or it is expressed at lower levels (HuH7). The
immunofluorescence analysis performed in HA22T/VGH cells revealed a
cytoskeletal localization of VIM (Fig.
5D).
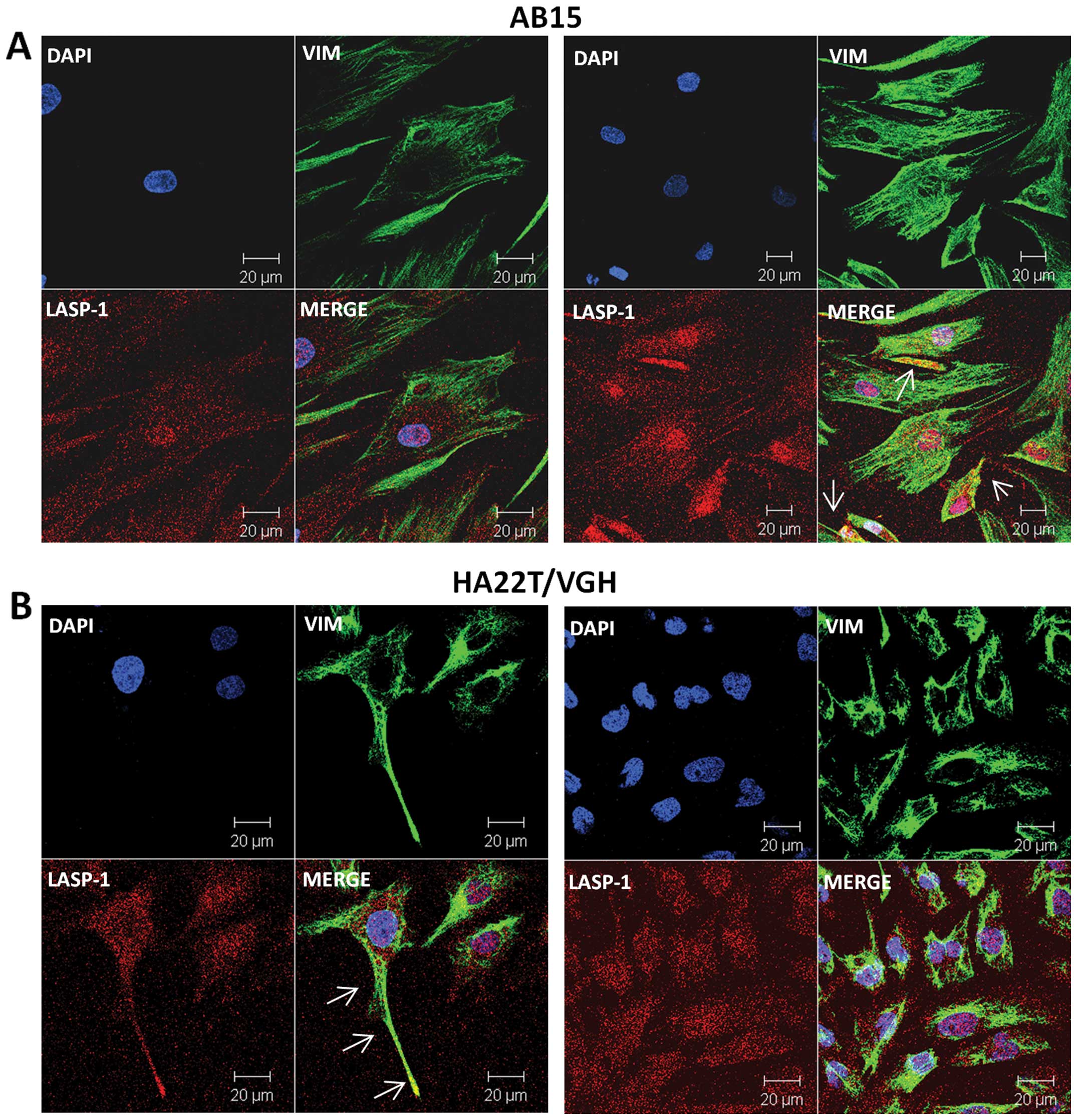
We further analyzed the eventual co-localization of
VIM and LASP-1 in human cell lines, dermal fibroblasts (AB15) and
HCC derived cells (HA22T/VGH) by confocal immunofluorescence. The
Z-stack observation evidenced that LASP-1 in AB15 cells was
localized both in the membrane and in the nuclei (Fig. 6A) and that VIM is mainly localized
in the cytoplasm probably in the cytoskeleton structure. VIM and
LASP-1 co-localized in AB15 cells as shown by white arrows
(Fig. 6A, right). In HA22T/VGH
cells LASP-1 is localized into the nucleus and in the cytoplasm and
VIM in the cytoplasm (Fig. 6B).
VIM and LASP-1 are co-localized in several portions of the
cytoplasm and this supported the results obtained by
co-immunoprecipitation experiments. VIM and LASP-1 also
co-localized in the cellular extensions of HA22T/VGH (white
arrows).
Discussion
LASP-1 is a key protein that explicates its
functions in the lamellipodia, filipodia, pseudopodia and focal
adhesions during cell movement and it contributes in maintaining
the cytoskeleton architecture by binding actin filaments (21). In various cancers, LASP-1 is
overexpressed, it influences the aggressive behavior of the cells
promoting proliferation, migration, invasion and metastasis
(9–13). Concerning LASP-1 and HCC, it is
known from global transcriptional profiles of matched pairs of HBV
associated HCC tumor and non-tumor liver tissue specimens that
LASP-1 is a gene significantly upregulated in this subgroup of HCC
(22,23). The same authors reported qPCR
validation data showing LASP-1 upregulation in 8/8 HBV associated
HCC cases and further demonstrated that LASP-1 is transcriptionally
repressed by p53 (23).
During the accomplishment of our study Wang et
al (12) demonstrated the
overexpression of LASP-1 protein in HCC; they showed no correlation
between LASP-1 protein expression and liver cirrhosis of HCC
patients while they found correlation between cytosolic and nuclear
LASP-1 protein expression with hepatitis B surface antigen (HBsAg)
of HCC patients. In this regard it is known that HBx could
upregulate LASP-1 through PI3-K (24). In recent years, using a proteomics
analysis, we found that LASP-1 is a mediator of uPA during
migration of HCC cells (20) and
that LASP-1 expression is coordinated with the overexpression of
uPA, a negative prognostic marker for this type of cancer (25). In the present study, we wanted to
investigate further the role of LASP-1 expression in HCC by
determining for the first time the expression levels of LASP-1 mRNA
in HCC tumors with different hepatic background disease and by
characterizing the ectopic LASP-1 overexpression in HCC cells using
MALDI-TOF mass spectrometry.
From the determinations of LASP-1 expression levels
in all cases of HCC tested, LASP-1 mRNA levels were generally and
significantly upregulated in HCCs compared to their adjacent
non-tumor counterpart. The analysis of differential R-values
displayed three HCC subgroups, the first with LASP1 up regulation
(23/55, 42%), the second with LASP-1 downregulation (19/55, 34%)
and the third with similar LASP-1 expression (13/55, 24%). No
clinic parameter (gender, age, tumor grading, TNM, viral hepatitis
infection and background hepatic disease) correlated with the
LASP-1 mRNA levels in these three subgroups.
By subdividing the HCC patients according to the
gender, LASP-1 mRNA was significantly overexpressed in HCC compared
to the PT tissues in female HCC patients while in males the levels
were comparable. We have not yet investigated the reasons of this
finding. In general, men are two to four times more often
associated with HCC than women (26) and in our study the number of males
and females enrolled reflected this proportion. It is known that
men and women have a different risk in developing HCC since
estrogens prevent while androgens promote liver cancer but the
molecular mechanism of action remains unclear (27). We cannot exclude that the
overexpression of LASP-1, at least at mRNA level, in the HCC tissue
may be regulated by different mechanisms in males and females and
that sex hormones might be involved. Note that LASP-1 is known to
be upregulated in breast cancer, where it was first identified, and
in ovarian cancer that are malignancies promoted by female hormonal
components (9,17). Foxa1 and Foxa2 were reported as
focal transcription factors for the sexual dimorphism of HCC
(28). Using bioinformatics
(29) we actually verified that
the LASP-1 promoter could be recognized by both Foxa1 and Foxa2 in
multiple sites. Whether the LASP-1 differential expression in
female and male HCCs may be controlled by these two transcription
factors remains a possibility and an interesting point for further
investigation.
By subdividing the HCC patients in cirrhotic and
non-cirrhotic HCCs, in the present work, for the first time, we
found a significant LASP-1 overexpression in the HCCs developed in
cirrhotic livers. Therefore, the evaluation of LASP-1 mRNA levels
in cirrhotic livers (i.e. in liver biopsies) might be a useful tool
to monitor the progression of the disease from hepatic cirrhosis to
the HCC stage. Most of the HCCs occur in the setting of cirrhosis
that is present in approximately 80–90% of HCC patients and
represents the largest single risk factor and hence can be
considered a premalignant condition (1). Its presence impacts survival,
strongly influences treatment decisions, and clearly needs the
increasingly common multidisciplinary approach to HCC
management.
When we were carrying out this study, Wang et
al provided IHC evidence for LASP-1 protein expression
generally higher in HCC than in adjacent non-tumoral tissues and
its expression level was associated with HBsAg and AFP levels
(12). In our study, we did not
find any correlation between LASP-1 mRNA expression and HBsAg and
AFP, but we tested and verified in few cases that in our specimens
the mRNA and protein expression of LASP-1 were comparable and
generally displayed the same trend of expression. This was not
obvious since it is well known that a given mRNA and the
correspondent protein not necessary follow the same trend of
expression. Hence we can argue that the regulatory mechanisms
underlying the differential expression of LASP-1 in HCC might act
at mRNA level. For IHC determinations, the protein levels were more
difficult to determine and compare than mRNA levels since the
localization of LASP-1 protein was nuclear, cytoplasmic or both; we
believe that LASP-1 mRNA levels are better quantifiable and in this
study it allowed the stratification of the cases on the basis of
the gender and the presence of hepatic cirrhosis as a background
liver disease. The stratification of HCC cases in more homogeneous
groups defined by clinical features and molecular parameters can be
of help to study and to identify targeted therapies, one of the
ambitious goals of basic molecular oncology.
Concerning the molecular characterization of LASP-1
in HCC cells, with the use of MALDI-TOF mass spectrometer we found
that vimentin (VIM) is a new molecular partner of LASP-1 in two
undifferentiated HCC cell lines. VIM is a member of the
intermediate filaments (IF) and it is important in determining the
cytoskeleton structure and its changes in both physiological and
pathological conditions (30). VIM
also acts as signal transducer, relaying information from the
extracellular matrix (ECM) to nuclei and it is an important
hallmark of the EMT (epithelial-mesenchymal transition) that
results in the loss of cellular adhesion and increased migratory
and invasive activities of several types of tumor cells (31). It is known that the overexpression
of VIM is significantly associated with HCC metastasis and it is a
circulating molecular biomarker for HCC (32,33)
and that LASP-1 induces TGF-β-mediated EMT transition in human
colorectal cancer (34). As
mentioned above, in previous studies, we demonstrated that uPA is
overexpressed in HCC and that it is a responsive therapeutic target
since its inhibition provoked a decrease of HCC cellular migration
and invasion (25,35–37).
We also demonstrated LASP-1 as a mediator of uPA in cell motility
(20). With the results obtained
in the present study, we can add new knowledge and hypothesize that
VIM can directly interact with LASP-1 probably during the
cytoskeleton dynamics necessary for the HCC cell motility. It will
be of interest to assess in the future whether the mRNA expression
of VIM and LASP-1 is coordinated in HCC.
For cell localization of LASP-1 protein, data in the
literature point the attention to the nuclear localization of
LASP-1 in some cancers (e.g. breast cancer and HCC) (17–19).
Utilizing IHC and confocal immunofluorescence analysis we observed
the nuclear localization of LASP-1 in HCC tissues and in human
cells. Among the cases analyzed by IHC, one presented intra-hepatic
recurrence (LV 144) and it showed almost exclusive nuclear
localization with enhanced expression compared to the primary
tumor. This could be in line with the consideration that the
nuclear localization of LASP-1 may be associated with a less
favorable prognosis of cancer patients (18).
In our in vitro studies the use of confocal
immunofluorescence displayed the nuclear localization of LASP-1
both in normal human fibroblasts and in HCC derived cells, but at
higher level in cancer cells as expected. To the best of our
knowledge, no data on nuclear localization in the normal biological
context exist as yet in the literature; therefore our observations
exclude that the nuclear localization of LASP-1 may be specifically
cancer-associated. LASP-1 in the nucleus might be involved in
controlling gene expression possibly as a co-transcription factor
and it may contribute in defining the nuclear F-actin architecture
(19). Nuclear localization in
human fibroblasts suggests that LASP-1 could be involved in the
functions described above not only in the cancer context but also
in normal conditions. Further studies are needed to better
understand the functions of LASP-1 in the nucleus.
In conclusion, the present study has evidenced the
upregulation of LASP-1 mRNA expression particularly in female HCCs
and in cirrhotic HCCs. The identification of groups of HCC patients
with shared molecular and clinical characteristics is important to
set up the follow-up of the patients and to study better therapies
(37). Moreover, we have
identified VIM as a new molecular partner of LASP-1. Most probably,
the overexpression of uPA, LASP-1 and VIM in HCC triggers the
malignant ability of the HCC cells, in particular migration,
because this requires the cytoskeleton remodeling. There are some
anticancer drugs in current clinics that directly affect vimentin,
such as silibinin and withaferin A (38,39).
The finding that LASP-1 can collaborate with VIM and uPA in
aggressive HCC cells may be of help in future studies of innovative
therapies targeting these molecules alone or in combination or by
miR-mediated negative regulation (40,41).
Acknowledgements
The authors would like to thank Dr Jenovia Smith for
the linguistic revision of the manuscript. This study was partially
supported by AIRC, the Ministero dell’Istruzione, dell’Università e
della Ricerca (MIUR), MIUR PRIN 2007 (MWCEAL_003), by MIUR local
funds of the University of Brescia, by Regione Lombardia NEDD, by
Lega Italiana per la Lotta contro i Tumori (LILT), by Centro per lo
studio, la prevenzione e la cura delle patologie epatiche di
interesse chirurgico, Brescia, Italy.
References
|
1
|
Caldwell S and Park SH: The epidemiology
of hepatocellular cancer: From the perspectives of public health
problem to tumor biology. J Gastroenterol. 44(Suppl 19): 96–101.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bertino G, Di Carlo I, Ardiri A, Calvagno
GS, Demma S, Malaguarnera G, Bertino N and Malaguarnera M, Toro A
and Malaguarnera M: Systemic therapies in hepatocellular carcinoma:
Present and future. Future Oncol. 9:1533–1548. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
El-Serag HB: Hepatocellular carcinoma. N
Engl J Med. 365:1118–1127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tomasetto C, Moog-Lutz C, Régnier CH,
Schreiber V, Basset P and Rio MC: Lasp-1 (MLN 50) defines a new LIM
protein subfamily characterized by the association of LIM and SH3
domains. FEBS Lett. 373:245–249. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schreiber V, Moog-Lutz C, Régnier CH,
Chenard MP, Boeuf H, Vonesch JL, Tomasetto C and Rio MC: Lasp-1, a
novel type of actin-binding protein accumulating in cell membrane
extensions. Mol Med. 4:675–687. 1998.PubMed/NCBI
|
|
6
|
Chew CS, Chen X, Parente JA Jr, Tarrer S,
Okamoto C and Qin HY: Lasp-1 binds to non-muscle F-actin in vitro
and is localized within multiple sites of dynamic actin assembly in
vivo. J Cell Sci. 115:4787–4799. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lin YH, Park ZY, Lin D, Brahmbhatt AA, Rio
MC, Yates JR III and Klemke RL: Regulation of cell migration and
survival by focal adhesion targeting of Lasp-1. J Cell Biol.
165:421–432. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Stölting M, Wiesner C, van Vliet V, Butt
E, Pavenstädt H, Linder S and Kremerskothen J: Lasp-1 regulates
podosome function. PLoS One. 7:e353402012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Grunewald TG, Kammerer U, Winkler C,
Schindler D, Sickmann A, Honig A and Butt E: Overexpression of
LASP-1 mediates migration and proliferation of human ovarian cancer
cells and influences zyxin localisation. Br J Cancer. 96:296–305.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Traenka C, Remke M, Korshunov A, et al:
Role of LIM and SH3 protein 1 (LASP1) in the metastatic
dissemination of medulloblastoma. Cancer Res. 70:8003–8014. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Payton S: Bladder cancer: LASP-1 - a
promising urine marker for detection of bladder cancer. Nat Rev
Urol. 9:2402012. View Article : Google Scholar
|
|
12
|
Wang H, Li W, Jin X, Cui S and Zhao L: LIM
and SH3 protein 1, a promoter of cell proliferation and migration,
is a novel independent prognostic indicator in hepatocellular
carcinoma. Eur J Cancer. 49:974–983. 2013. View Article : Google Scholar
|
|
13
|
Shimizu F, Shiiba M, Ogawara K, et al:
Overexpression of LIM and SH3 protein 1 leading to accelerated G2/M
phase transition contributes to enhanced tumourigenesis in oral
cancer. PLoS One. 8:e831872013. View Article : Google Scholar
|
|
14
|
Hailer A, Grunewald TGP, Orth M, Reiss C,
Kneitz B, Spahn M and Butt E: Loss of tumor suppressor mir-203
mediates overexpression of LIM and SH3 protein 1 (LASP1) in
high-risk prostate cancer thereby increasing cell proliferation and
migration. Oncotarget. 5:4144–4153. 2014.PubMed/NCBI
|
|
15
|
Grunewald TG, Kammerer U, Schulze E,
Schindler D, Honig A, Zimmer M and Butt E: Silencing of LASP-1
influences zyxin localization, inhibits proliferation and reduces
migration in breast cancer cells. Exp Cell Res. 312:974–982. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao L, Wang H, Liu C, et al: Promotion of
colorectal cancer growth and metastasis by the LIM and SH3 domain
protein 1. Gut. 59:1226–1235. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Grunewald TG, Kammerer U, Kapp M, Eck M,
Dietl J, Butt E and Honig A: Nuclear localization and cytosolic
overexpression of LASP-1 correlates with tumor size and
nodal-positivity of human breast carcinoma. BMC Cancer. 7:198–207.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Frietsch JJ, Grunewald TG, Jasper S,
Kammerer U, Herterich S, Kapp M, Honig A and Butt E: Nuclear
localisation of LASP-1 correlates with poor long-term survival in
female breast cancer. Br J Cancer. 102:1645–1653. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mihlan S, Reiß C, Thalheimer P, Herterich
S, Gaetzner S, Kremerskothen J, Pavenstädt HJ, Lewandrowski U,
Sickmann A and Butt E: Nuclear import of LASP-1 is regulated by
phosphorylation and dynamic protein-protein interactions. Oncogene.
32:2107–2113. 2013. View Article : Google Scholar
|
|
20
|
Salvi A, Bongarzone I, Miccichè F, Arici
B, Barlati S and De Petro G: Proteomic identification of LASP-1
down-regulation after RNAi urokinase silencing in human
hepatocellular carcinoma cells. Neoplasia. 11:207–219.
2009.PubMed/NCBI
|
|
21
|
Grunewald TG and Butt E: The LIM and SH3
domain protein family: Structural proteins or signal transducers or
both? Mol Cancer. 7:31–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Neo SY, Leow CK, Vega VB, Long PM, Islam
AFM, Lai PB, Liu ET and Ren EC: Identification of discriminators of
hepatoma by gene expression profiling using a minimal dataset
approach. Hepatology. 39:944–953. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang B, Feng P, Xiao Z and Ren EC: LIM and
SH3 protein 1 (Lasp1) is a novel p53 transcriptional target
involved in hepatocellular carcinoma. J Hepatol. 50:528–537. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tang R, Kong F, Hu L, You H, Zhang P, Du W
and Zheng K: Role of hepatitis B virus X protein in regulating LIM
and SH3 protein 1 (LASP-1) expression to mediate proliferation and
migration of hepatoma cells. Virol J. 9:163–175. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
De Petro G, Tavian D, Copeta A, Portolani
N, Giulini SM and Barlati S: Expression of urokinase-type
plasminogen activator (u-PA), u-PA receptor, and tissue-type PA
messenger RNAs in human hepatocellular carcinoma. Cancer Res.
58:2234–2239. 1998.PubMed/NCBI
|
|
26
|
Yeh YT, Chang CW, Wei RJ and Wang SN:
Progesterone and related compounds in hepatocellular carcinoma:
Basic and clinical aspects. Biomed Res Int. 2013:2905752013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yeh SH and Chen PJ: Gender disparity of
hepatocellular carcinoma: The roles of sex hormones. Oncology.
78(Suppl 1): 172–179. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li Z, Tuteja G, Schug J and Kaestner KH:
Foxa1 and Foxa2 are essential for sexual dimorphism in liver
cancer. Cell. 148:72–83. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rosenbloom KR, Dreszer TR, Long JC, et al:
ENCODE whole-genome data in the UCSC Genome Browser: Update 2012.
Nucleic Acids Res. 40:D912–D917. 2012. View Article : Google Scholar :
|
|
30
|
Ivaska J, Pallari HM, Nevo J and Eriksson
JE: Novel functions of vimentin in cell adhesion, migration, and
signaling. Exp Cell Res. 313:2050–2062. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Savagner P: The epithelial-mesenchymal
transition (EMT) phenomenon. Ann Oncol. 21(Suppl 7): vii89–vii92.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hu L, Lau SH, Tzang CH, et al: Association
of vimentin overexpression and hepatocellular carcinoma metastasis.
Oncogene. 23:298–302. 2004.
|
|
33
|
Pan TL, Wang PW, Huang CC, Yeh CT, Hu TH
and Yu JS: Network analysis and proteomic identification of
vimentin as a key regulator associated with invasion and metastasis
in human hepatocellular carcinoma cells. J Proteomics.
75:4676–4692. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang H, Shi J, Luo Y, Liao Q, Niu Y, Zhang
F, Shao Z, Ding Y and Zhao L: LIM and SH3 protein 1 induces
TGFβ-mediated epithelial-mesenchymal transition in human colorectal
cancer by regulating S100A4 expression. Clin Cancer Res.
20:5835–5847. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Salvi A, Arici B, Alghisi A, Barlati S and
De Petro G: RNA interference against urokinase in hepatocellular
carcinoma xenografts in nude mice. Tumour Biol. 28:16–26. 2007.
View Article : Google Scholar
|
|
36
|
Salvi A, Arici B, De Petro G and Barlati
S: Small interfering RNA urokinase silencing inhibits invasion and
migration of human hepatocellular carcinoma cells. Mol Cancer Ther.
3:671–678. 2004.PubMed/NCBI
|
|
37
|
Salvi A, Abeni E, Portolani N, Barlati S
and De Petro G: Human hepatocellular carcinoma cell-specific miRNAs
reveal the differential expression of miR-24 and miR-27a in
cirrhotic/non-cirrhotic HCC. Int J Oncol. 42:391–402. 2013.
|
|
38
|
Wu KJ, Zeng J, Zhu GD, Zhang LL, Zhang D,
Li L, Fan JH, Wang XY and He DL: Silibinin inhibits prostate cancer
invasion, motility and migration by suppressing vimentin and MMP-2
expression. Acta Pharmacol Sin. 30:1162–1168. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Satelli A and Li S: Vimentin in cancer and
its potential as a molecular target for cancer therapy. Cell Mol
Life Sci. 68:3033–3046. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Salvi A, Conde I, Abeni E, Arici B, Grossi
I, Specchia C, Portolani N, Barlati S and De Petro G: Effects of
miR-193a and sorafenib on hepatocellular carcinoma cells. Mol
Cancer. 12:162–176. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Salvi A, Sabelli C, Moncini S, Venturin M,
Arici B, Riva P, Portolani N, Giulini SM, De Petro G and Barlati S:
MicroRNA-23b mediates urokinase and c-met downmodulation and a
decreased migration of human hepatocellular carcinoma cells. FEBS
J. 276:2966–2982. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gorla L, Mondellini P, Cuccuru G, Miccichè
F, Cassinelli G, Cremona M, Pierotti MA, Lanzi C and Bongarzone I:
Proteomics study of medullary thyroid carcinomas expressing RET
germ-line mutations: Identification of new signaling elements. Mol
Carcinog. 48:220–231. 2009. View Article : Google Scholar
|