Introduction
Breast cancer (BC) is one of the most prevalent
cancers in women and accounts for ~22.9% of all cancers in women
worldwide (1). BC also is the
second most prevalent malignancy and the fifth most common cause of
cancer death worldwide (1). In
China, breast cancer is one of the most common cancers in women,
and its incidence has increased by 3% annually (2). The tumorigenesis process of BC is
complicated involving many genetic and molecular alterations
(3). Although many researchers
have demonstrated signal pathways involved in BC initiation and
procession, understanding the mechanism underlying the development
of BC is challenging. Therefore, better understanding of the
molecular mechanisms underlying BC development and progression is
urgently needed.
MicroRNAs (miRNAs) are a group of endogenous
smallnon-coding RNAs that regulate the expression of their target
genes post-transcriptionally, by directly binding to the
untranslated region or the open reading frame and thus inducing RNA
degradation or the inhibition of protein translation (4,5).
Recent evidence strongly supports the finding that miRNAs
participate in the regulation of many cellular processes, including
development, differentiation, apoptosis, and proliferation
(6). Growing evidence indicated
that miRNAs are aberrantly expressed in different types of tumors
(7) and involved in human
tumorigenesis and/or metastasis by directly targeting oncogenes or
tumor suppressor genes (8). Many
miRNAs have been confirmed as modulators of cell proliferation,
apoptosis, and therapy resistance in BC (9–12).
Therefore, more extensive investigations are needed on the role of
miRNAs, which would contribute to develop novel avenues for
targeted therapy.
Recently, the expression pattern and function of the
miR-338-3p was widely studied in various cancers, and reported to
function as a tumor suppressor gene in various cancer, including
hepatocellular carcinoma (13,14),
neuroblastoma (15), ovarian
cancer (16), malignant melanoma
(17), gastric cancer (18,19)
and colorectal cancer (20,21).
However, to our knowledge, its roles and the potential mechanisms
in BC remain unclear. Hence, our study was aimed to identify the
role of miR-338-3p in BC. We found that the expression of
miR-338-3p was suppressed in both BC tissues and cancer cell lines.
Furthermore, the low expression of miR-338-3p was associated with
late TNM stage and lymph node metastatic status. In addition,
overexpression of miR-338-3p in BC cells inhibited cell
proliferation, colony formation, migration and invasion, induce
apoptosis and cell cycle arrest at G1/G0 stage in vitro, and
suppressed tumor growth in vivo by targeting SOX4, which was
identified as a direct and functional target of miR-338-3p.
Materials and methods
Patients and tissue samples
Breast cancer samples and the corresponding adjacent
ovarian tissues were obtained from 32 patients with primary BC who
underwent surgery at China-Japan Union Hospital of Jilin University
(Changchun, China) from July 2012 to August 2014. Normal breast
tissues adjacent to the tumor were taken from 3 cm away from the
tumor cells. All of the samples and matched clinical information
were collected after obtaining prior written informed consent from
the patients. The samples were immediately snap-frozen in liquid
nitrogen and stored at −80°C until use. No patients received
chemotherapy or radiotherapy prior to surgery. This study is
approved by Institutional Ethics Committees of Jilin
University.
Cell lines and cell culture
The non-cancerous human mammary epithelial cell line
MCF-10A, breast cancer cell line and human breast cancer cell lines
MCF-7, MDA-MB-231, BT-549 and MDA-MB-453 were obtained from the
Institute of Biochemistry and Cell Biology, Chinese Academy of
Sciences (Shanghai, China). All cells were maintained in RPMI-1640
medium (Gibco, Grand Island, NY, USA) containing 10% fetal bovine
serum (FBS, Gibco BRL, Gaithersburg, MD, USA), 100 U/ml penicillin
and 100 mg/ml streptomycin at 37°C in a humidified chamber
supplemented with 5% CO2.
Quantitative reverse transcription
polymerase chain reaction (qRT-PCR)
Total RNA was extracted from tissue and cells using
TRIzol (Invitrogen) according to the manufacturer's instructions.
For miRNA reverse transcription, cDNA was synthesized using One
Step Prime script miRNA cDNA Synthesis kit (Qiagen, Valencia, CA,
USA) according to the manufacturer's instructions. For mRNA reverse
transcription, cDNA was synthesized using the Primer Script RT
reagent kit (Takara Bio, Japan). Quantitative PCR was performed
using Fast SYBR Green Master Mix (Applied Biosystems, Foster City,
CA, USA) under ABI 7900 Sequence Detection System (Life
Technologies, NY, USA). U6 snRNA and GAPDH was used as an
endogenous control. The specific primers for miRNA-126 and U6 were
purchased from Applied Biosystems. The specific primers for GAPDH
and SOX4 are as follows: GAPDH (sense),
5′-TCAACGACCACTTTGTCAAGCTCA-3′; antisense:
5′-GCTGGTGGTCCAGGGGTCTTACT-3′; SOX4 (sense),
5′-AGCGACAAGATCCCTTTCATTC-3′; antisense: 5′-CGTTGCCGGACTTCACCTT-3′.
The comparative 2−ΔΔCt method was used for relative
quantification and statistical analysis. All experiments were
performed in triplicate.
Transient transfection of miRNA
mimics
miRNA-338-3p mimics and negative control mimics
(miR-NC) were purchased from GenePharma (Shanghai, China).
MDA-MB-231 cells were seeded into cell culture plates 20 h before
transfection to ensure 70% cell confluence at the moment of
transfection. Transfection of miRNA mimics into MDA-MB-231 cells
was performed using Lipofectamine 2000 (Invitrogen) according to
the manufacturer's procedure at the final concentration of 100 nM.
At 48 h post-transfection, transfection efficiencies were evaluated
in every experiment by qRT-PCR and western blot analysis.
Cell proliferation (MTT) assay and colony
formation assay
The transfected cells (5×103 cells/well)
were plated into 96-well plates. At 24, 48 and 72 h
post-transfection, MTT
(3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) were
added into cells and cultured for 4 h at 37°C, followed by removal
of the culture medium and the addition of 150 μl dimethyl sulfoxide
(DMSO, Sigma-Aldrich). The absorbance at 490 nm (OD490 nm) was
measured with a spec-trophometer.
For colony formation, the transfected cells were
seeded into a 6-well plate at a density of 1,000 cells/well. The
medium was changed every three days. Approximately 2 weeks later,
the clones were washed with 1X PBS and stained with 1.0% crystal
violet (Sigma) for 5 min. Finally the clones were photographed and
counted.
Cell cycle and apoptosis assay
Cell cycle and apoptosis assay was performed on
MDA-MB-231 cells 48 h after transfection. For cell cycle assay, the
transfected cells at 1×106 cells per well were cultured
in 12-well plates in triplicate and cultured for 24 h. Then the
cells were collected by trypsinization and washed in PBS and fixed
in ice-cold ethanol overnight at 4°C, followed by incubated in 1 ml
of staining solution (20 μg/ml propidium iodide and 10 U/ml RNaseA)
for 30 min at room temperature. Cell cycle distribution was assayed
by fluorescence-activated cell sorting based on FACSCalibur flow
cytometer (BD Biosciences, San Jose, CA, USA). All experiments were
performed in triplicate.
Cell apoptosis analysis was performed with Annexin
V-FITC Apoptosis Detection kit I (BD Biosciences) according to the
manufacturer's instructions using FACSCalibur flow cytometer (BD
Biosciences).
Wound healing assay
Transfected cells (5×103) were seeded
into 24-well tissue culture plates for 48 h. Thereafter, a linear
wound of cellular monolayer was created in the confluent cells.
After wounding, the debris was removed by washing the cells with
PBS. Migration of cells into the wound was observed at 0 and 24 h
using an IX51 inverted microscope (Olympus, Tokyo, Japan).
Individual cells were quantified as an average of at least five
fields for each experiment.
Cell invasion assay
Cell invasion assays were performed using 24-well
Transwell Permeable Supports with 8-μM pores (Corning, Lowell, MA,
USA). Briefly, 2×104 transfected cells were suspended in
serum-free medium and seeded into the Transwell inserts coated with
growth factor reduced Matrigel (BD Biosciences, Bedford, MA, USA).
Bottom wells were filled with media containing complete media. The
invasion assay was performed for 24 h at 37°C in a 5%
CO2 incubator. After incubation, migrating cells present
on the lower surface of the membrane were fixed in 70% ethanol for
30 min and stained with 2% crystal violet for 10 min on a glass
slide. Cells from 10 random fields were counted under an IX51
inverted microscope (Olympus).
miRNA target prediction
Prediction of the miR-338-3p targets was performed
using two publicly available algorithms: TargetScan6.2 (http://www.targetscan.org/), miRanda (http://www.microrna.org/).
Assay of luciferase activity
The 3′-UTR of SOX4 was amplified and cloned
downstream of pGL3/Luciferase vector (Wt SOX4 3′-UTR). Then the
mutant 3′-UTR of SOX4 (several nucleotides within the binding sites
were mutanted) was amplified using pGL3/Luc-SOX4 3′-UTR as the
template, and was cloned downstream of the pGL3/Luciferase vector
(Mut SOX4 3′-UTR). For the luciferase reporter assay, the cells
were co-transfected with miR-338-3p mimic or miR-NC and Wt SOX4
3′-UTR or Mut SOX4 3′-UTR, together with the controls. Forty-eight
hours post-transfection, cells were lysed using 1X passive lysis
buffer (Promega, Madison, WI, USA) and lysates were analyzed using
the Dual-Glo Luciferase Reporter Assay System (Promega) on the
Synergy4 multi-mode microplate reader (BioTeK).
Western blot analysis
Total protein was extracted by using RIPA buffer
(Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) from cells
harvested 48 h after transfection, separated in 10% SDS
polyacrylamide gels, and electrophoretically transferred to
nitrocellulose membranes (NC membranes, Invitrogen, Carlsbad, CA,
USA), blocked in 4% dry milk at room temperature for 1 h, and
immunostained with primary antibodies at 4°C overnight using
anti-SOX4 (1:1,000, Santa Cruz); and anti-GAPDH antibody (1;5,000,
Santa Cruz). After washing, membranes were incubated with
horseradish peroxidase (HRP)-conjugated goat anti-mouse secondary
antibodies (1:5,000; Santa Cruz Biotechnology). The protein bands
were visualized on X-ray film with a chemiluminescent detection
system (Beyotime, Shanghai, China). Blots were stripped and
reprobed with anti-GAPDH to control for loading variations.
Nude mouse xenograft assay
Twenty female BALB/c mice (18–20-g, 4–5-week-old)
were obtained from Experimental Animal Center of Changchun
Biological Institute (Changchun, China), and kept under specific
pathogen-free (SPF) conditions. This study and all experimental
protocols were approved and the methods were performed in
accordance with the guidelines of the Animal Care and Use Committee
of Jilin University.
MDA-MB-231 cells (2×106), stably
expressing miR-338-3p or miR-NC, were suspended in 100 μl PBS and
then injected subcutaneously into the posterior flank of female
BALB/c athymic nude mice. Tumor volumes in mice were measured with
a slide caliper every week until scarifice according to the
formula: Volume (mm3) = 1/2 × width2 ×
length. Five weeks after injection, mice were sacrificed, and tumor
tissues were resected and weighed. In addition, SOX4 protein
expression level of tumor tissue was determined by western blot
analysis.
Statistical analysis
All the data are shown as mean ± SD (standard
deviation) and the experiments were repeated three times. The
difference was determined by two-tailed Student's t-test.
Statistical analysis was performed with GraphPad Prism 5.0
(GraphPad Software, San Diego, CA, USA). P<0.05 was considered
statistically significant.
Results
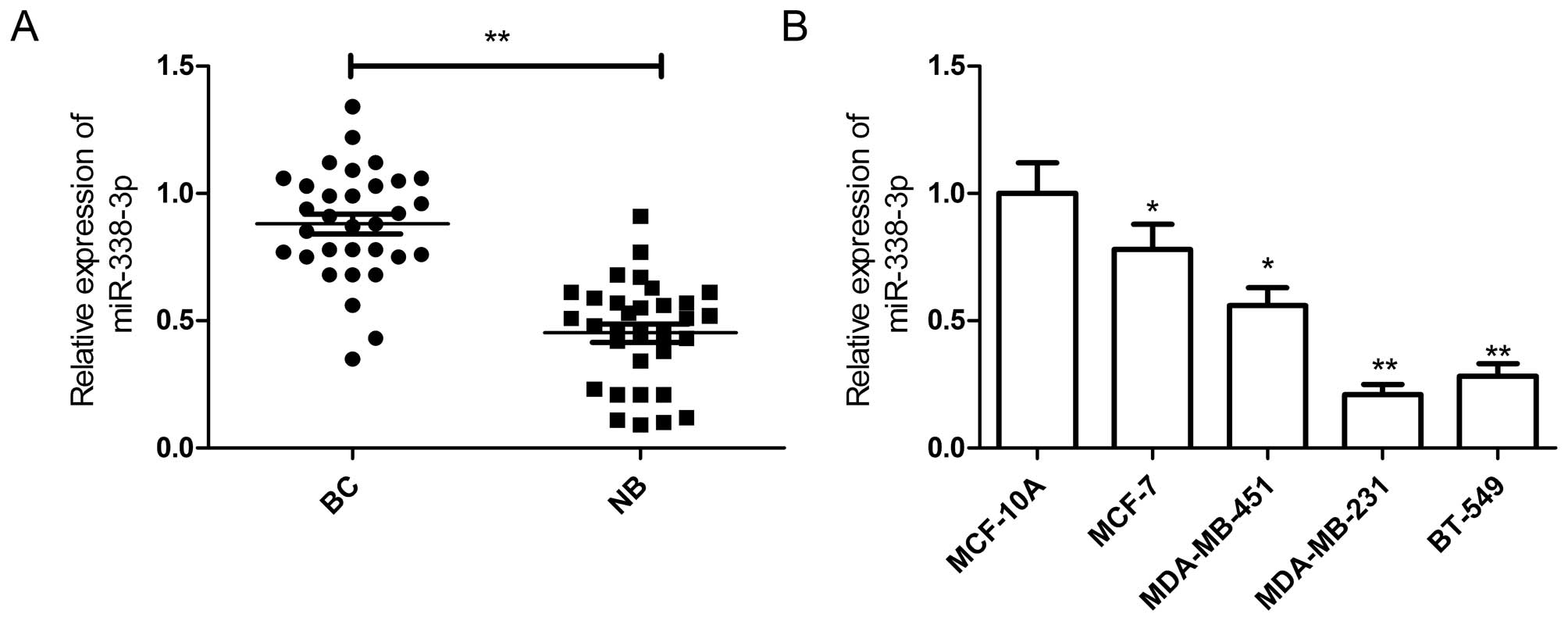
miR-338-3p is downregulated in breast
cancer tissues and cell lines
To determine the role of miR-338-3p in breast cancer
progression, we examined miR-338-3p expression in breast cancer
tissue samples and corresponding normal tissue sample by qRT-PCR.
miR-338-3p was downregulated in the breast cancer samples compared
with the paired normal breast tissues (Fig. 1A). To investigate the clinical
relevance of miR-338-3p in breast cancer, we divided the 32
patients to high miR-338-3p expression group (n=17) and low
miR-338-3p expression group (n=15) using the mean value (0.45±0.04)
of relative expression levels as a cutoff. The correlation between
the miR-338-3p expression level and clinical and pathologic
characteristics of breast cancer is listed in Table 1. In 12 cases presenting as
advanced stage III, 9 (75.0%) of the cases have low-level
miR-338-3p expression in cancer tissues; while in 20 early stages
(stages I and II), only 6 (30.0%) presented with low levels of
miR-338-3p expression. In the 13 cases of breast cancer patients
with lymph node metastasis, 10 (76.9%) exhibited low miR-338-3p
expression, however only 5 (26.3%) of 19 cases of cancers without
lymph node metastasis presented low-level miR-338-3p expression. No
correlation was observed between the miR-338-3p level and the age,
tumor size or pathologic grade status of breast cancer.
 | Table ICorrelation between
clinicopathological features and miR-338-3p expression in 32 breast
cancer tissues. |
Table I
Correlation between
clinicopathological features and miR-338-3p expression in 32 breast
cancer tissues.
| | miR-338-3p
expression | |
|---|
| |
| |
|---|
| Variables | No. of cases | Low (n %) | High (n %) | P-value |
|---|
| Age (years) | | | | 0.624 |
| <55 | 14 | 7 (50.0) | 7 (50.0) | |
| ≥55 | 18 | 8 (44.4) | 10 (55.6) | |
| Pathologic
grade | | | | 0.218 |
| I | 16 | 6 (37.5) | 10 (62.5) | |
| II,III | 16 | 9 (56.2) | 7 (43.8) | |
| TNM stage | | | | <0.01 |
| I–II | 20 | 6 (30.0) | 14 (70.0) | |
| III | 12 | 9 (75.0) | 3 (25.0) | |
| Tumor size | | | | 0.879 |
| <5 cm | 15 | 7 (46.7) | 8 (53.4) | |
| ≥5 cm | 17 | 8 (47.1) | 9 (52.9) | |
| Lymph node
metastasis | | | | <0.01 |
| No | 19 | 5 (26.3) | 14 (73.7) | |
| Yes | 13 | 10 (76.9) | 3 (23.1) | |
In addition, the expression of miR-338-3p was
detected in a panel of breast cancer cell lines and non-cancerous
breast epithelial cell line (MCF-10A). It was found that miR-338-3p
expression was downregulated in breast cancer cell lines compared
to normal non-cancerous breast epithelial cell line MCF-10A
(Fig. 1B). The data above showed
that miR-338-3p decreases in both breast cancer tissues and cell
lines, and that its expression is inversely correlated with the
metastatic abilities of breast cancer cells. We selected MDA-MB-231
cells, which showed the lowest expression of miR-338-3p, to conduct
further experiments.
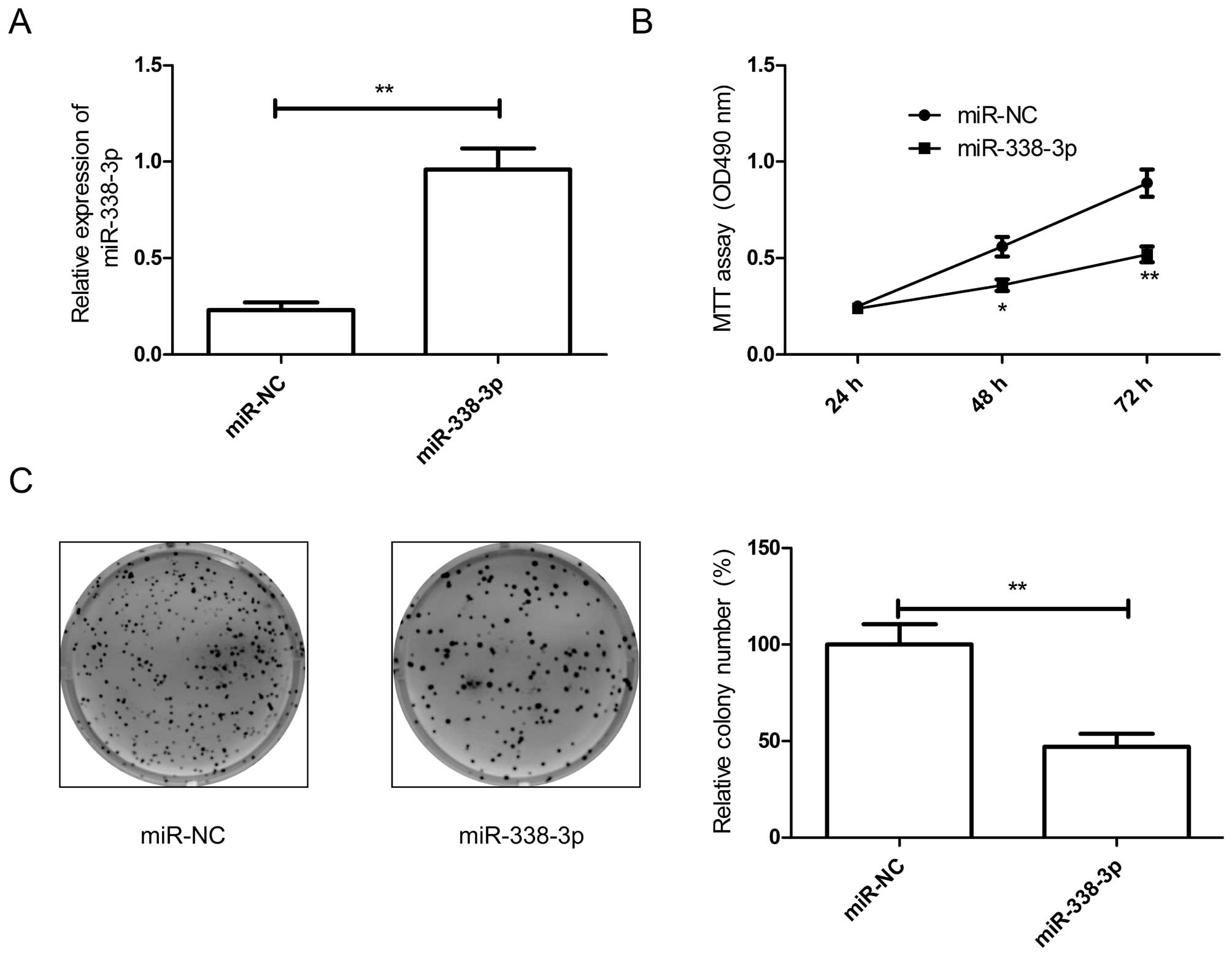
Overexpression of miR-338-3p inhibits
cell proliferation and colony formation of breast cancers
As miR-338-3p was downregulated in breast cancer
tissues and cell lines, we transfected miR-338-3p mimic into breast
cancer cells. The results from qRT-PCR assay showed that miR-338-3p
mimic could significantly upregulate the level of miR-338-3p
expression in breast cancer cells compared to miR-NC group
(Fig. 2A). To confirm miR-338-3p
effect on proliferation and colony formation in breast cancer
cells, MTT assay and colony formation assay were performed. It was
found that overexpression of miR-338-3p significantly inhibited
cell proliferation (Fig. 2B) and
colony formation (Fig. 2C) in
breast cancer cells.
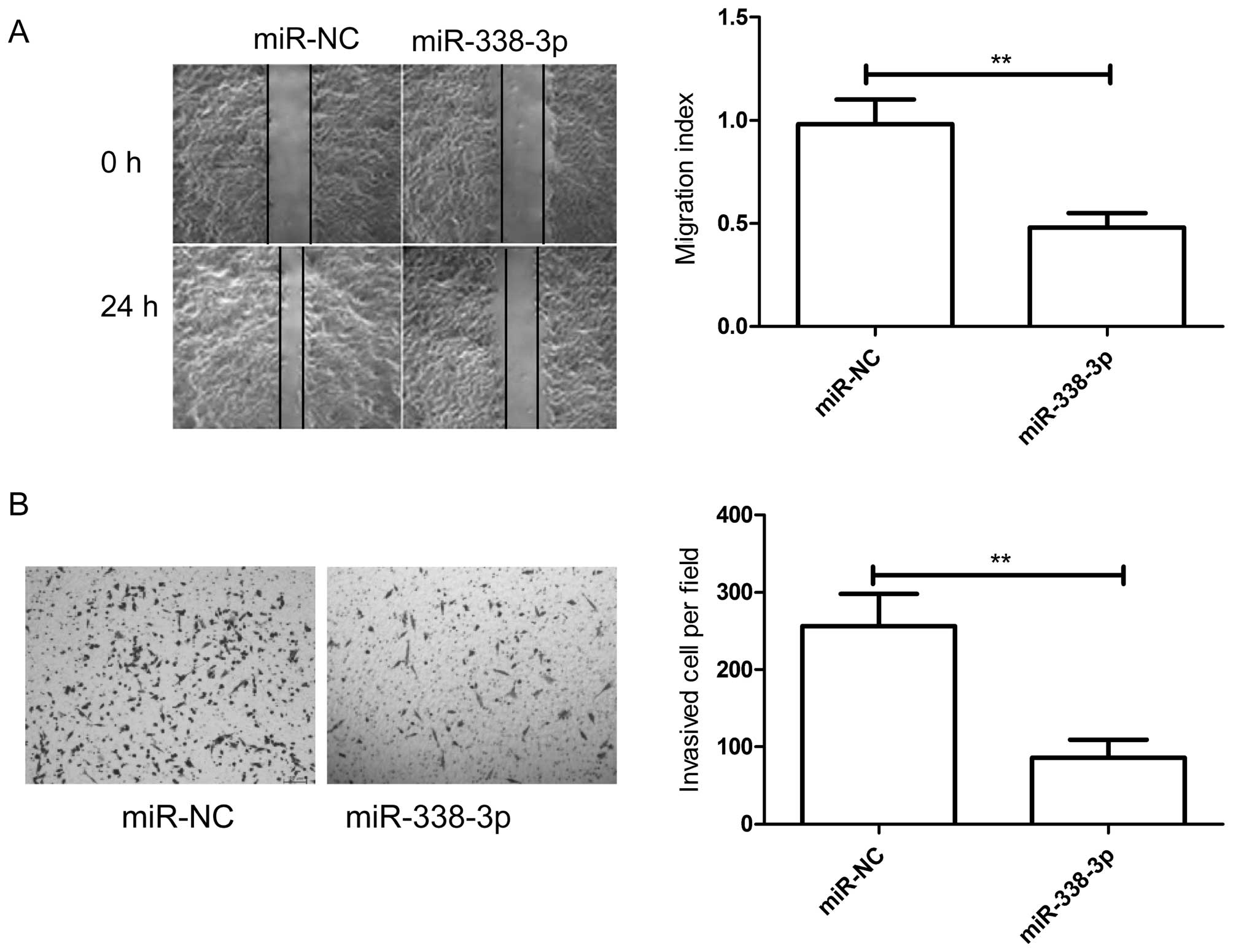
Overexpression of miR-338-3p inhibits
cell migration and invasion of breast cancers
The above results showed that miR-338-3p expression
is inversely correlated with the meta-static ability of breast
cancer cells, therefore, to investigate the miR-338-3p effect on
metastasis in vitro, migration and invasion assays were
performed in MDA-MB-231 cells transfected with miR-338-3p mimic or
miR-NC by wound healing and transwell chamber assay, respectively.
As expected, overexpression of miR-338-3p in breast cancer cells
significantly inhibited cell migration (Fig. 3A) and invasion (Fig. 3B) of breast cancer cells.
Collectively, these results suggested that miR-338-3p can
efficiently inhibited migration and invasion of breast cancer
cells.
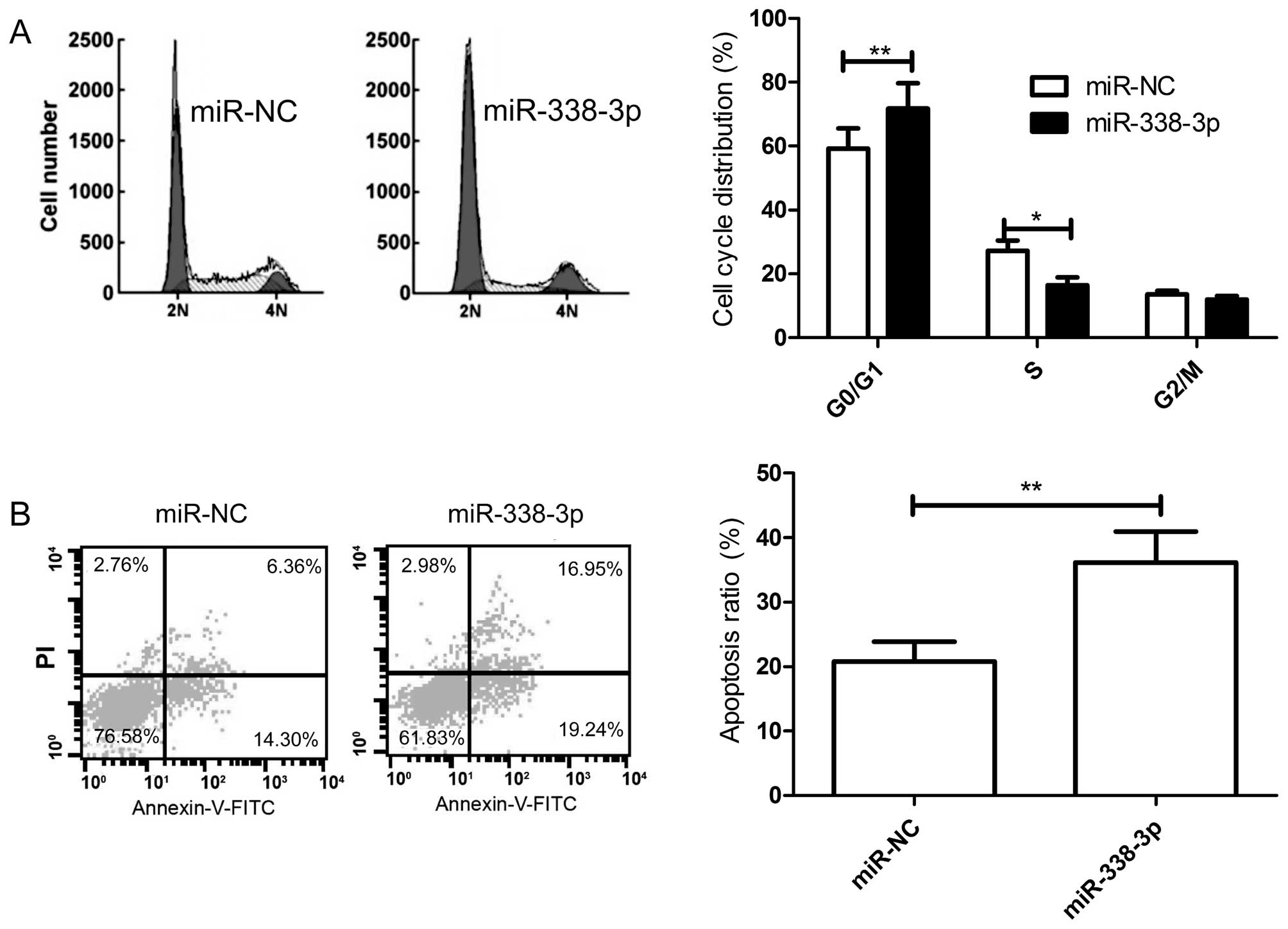
Overexpression of miR-338-3p induces cell
cycle arrest at G1/G0 stage and apoptosis of breast cancers
To further verify the role of miR-338-3p in breast
cancer cells, we tested the cell cycle and apoptosis effects in
breast cancer cells by flow cytometry. Cell cycle assay showed that
the percentage of G1 phase cells increased, and the percentage of S
phase cells decreased in breast cancer cells transfected with
miR-338-3p mimic compared to cells transfected with miR-NC
(Fig. 4A). Cell apoptosis assay
showed that overexpression of miR-338-3p significantly induced cell
apoptosis in breast cancer cells (Fig.
4B).
SOX4 is a direct target of
miR-338-3p
To explore the mechanism underlying the growth
inhibition by miR-338-3p in breast cancer cells, we used publicly
available algorithms (Targetscan6.2 and miRanda) to help identify
miR-338-3p targets in human breast cancer. We found that
sex-determining region Y-box 4 (SOX4) was a putative target of
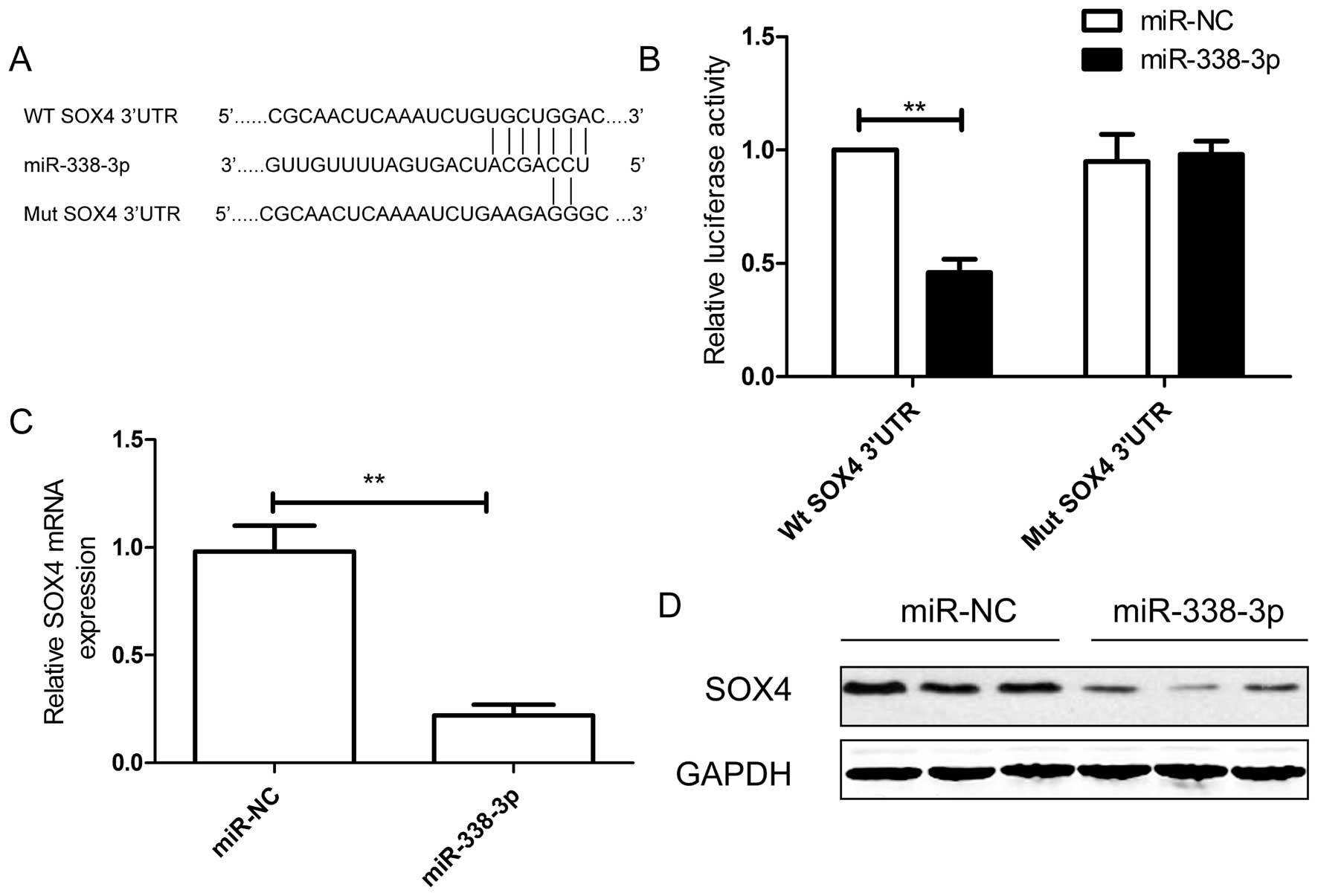
miR-338-3p. To confirm this possibility, the miR-338-3p binding
sequences present at the 3′-UTR of SOX4 mRNA (WT-3′-UTR SOX4) or
its mutant site (Mut-3′-UTR-SOX4) were subcloned downstream of the
luciferase reporter gene in pGL3 vector (Fig. 5A) and then co-transfected into
MDA-MB-231 cells, along with miR-338-3p mimic or miR-NC for
luciferase assay evaluation. Luciferase assay further revealed that
breast cancer cells transfected with miR-338-3p mimic repressed
wild-type 3′-UTR-SOX4 reporter activity (P<0.01), while
miR-338-3p mimic had no inhibition effect on the mutant 3′-UTR-SOX4
reporter activity (Fig. 5B),
indicting the direct regulation of miR-338-3p in the 3′-UTR of SOX4
mRNA. To further confirm that SOX4 acts as a target of miR-338-3p,
we examine the effect of miR-338-3p on SOX4 expression by qRT-PCR
and western blot analysis. As predicted, qRT-PCR and western
blotting showed that ectopic miR-338-3p in BC cells downregulated
SOX4 expression on mRNA level (Fig.
5C) and protein level (Fig.
5D).
SOX4 is upregulated in BC tissues and is
inversely correlated with miR-338-3p levels
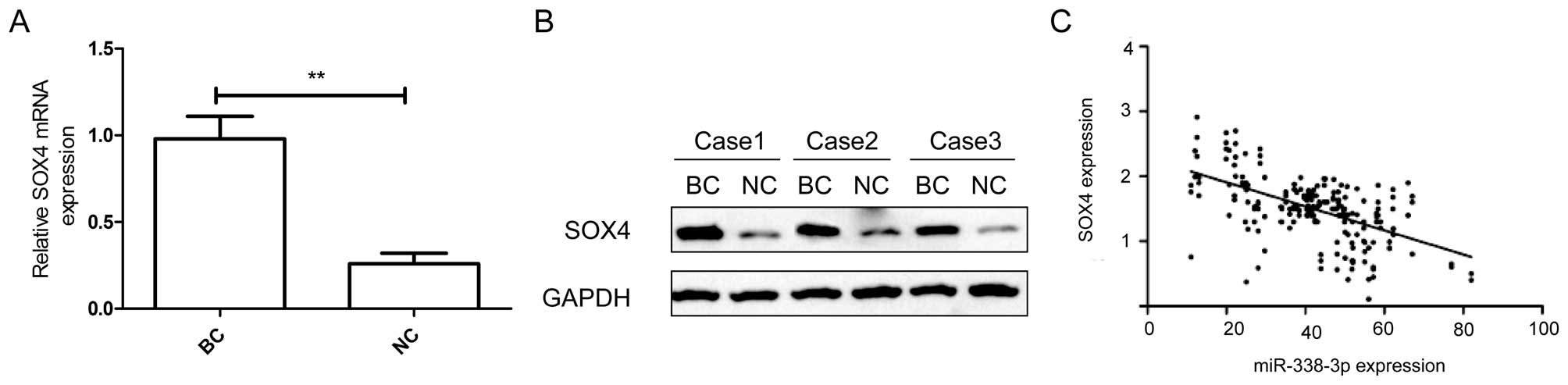
As the above results show that miR-338-3p is
downregulated in BC and targets SOX4 by binding to its 3′-UTR, we
next determined whether SOX4 expression is negatively associated
with miR-338-3p levels in the BC tissue samples. qRT-PCR and
western blot assay showed that the expression of SOX4 on mRNA and
protein level was significantly higher in BC tissues than in
matched normal tissues (P<0.05) (Fig. 6A and B). In addition, a
statistically significant inverse correlation was found between
expression levels of miR-338-3p and SOX4 mRNA in BC tissue by
Spearman's correlation analysis (r=−0.6431, P<0.001, Fig. 6C).
miR-338-3p inhibits tumorigenicity in
vivo
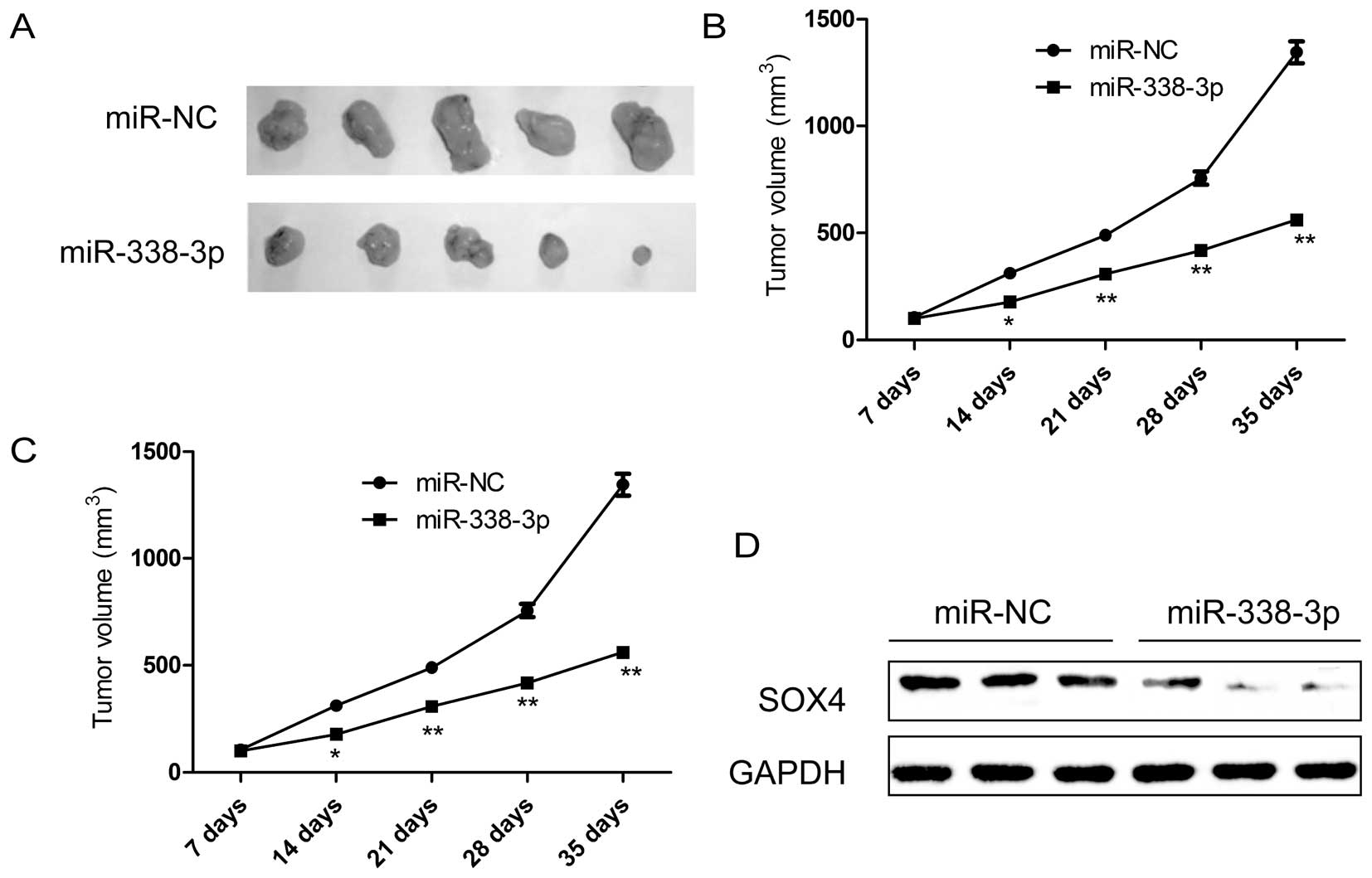
Finally, we tested whether ectopic expression of
miR-338-3p could influence the growth of breast tumor in
vivo. MDA-MB-231 cells with stable expression either of
miR-338-3p or miR-NC were selected and injected subcutaneously into
nude mice, and the tumor formation was monitored. MDA-MB-231 cells
transfected with miR-NC showed progressive growth, while MDA-MB-231
cells transfected with miR-338-3p mimic inhibited tumor growth
relative to miR-NC group. After 35 days, the nude mice were
sacrificed, and the tumor weights and volume were measured
(Fig. 7A–C). It was found that
overexpression of miR-338-3p can significantly suppress the tumor
growth of xenografts in nude mice (Fig. 7A), and decrease the tumor volume
(Fig. 7B) and tumor weight
(Fig. 7C) compared to miR-NC
group, indicating the suppressive function of miR-338-3p on breast
cancer tumorigenicity in vivo. In addition, we also
determined SOX4 protein expression in tumor tissue by western blot
analysis. We found that SOX4 protein expression was decreased in
the xenograft tumors of miR-338-3p mimic group compared to the
xenograft tumors of miR-NC group (Fig.
7D), suggesting that miR-338-3p suppressed breast cancer
tumorigenicity in vivo by targeting SOX4.
Discussion
Breast cancer is one of the most commonly diagnosed
solid malignancies and the leading cause of cancer-related deaths
among women (1). Although great
progress in surgical technique, diagnostic methods, and new
chemotherapy regimens, have significantly reduced breast
cancer-related mortality, nearly half of breast cancer patients
develop distant metastatic disease after treatment with
chemotherapeutic and/or hormonal drugs (2). Thus, there is an urgent need to
understand the molecular mechanism of breast cancer development and
metastasis for effective therapy. During the past years, microRNAs
(miRNAs) have emerged as promising prognostic and therapeutic
targets for metastatic breast cancers (22–24).
Lin et al (25) reported
that ectopic overexpression of miR-33b in highly metastatic breast
cancer cells suppresses cell self-renewal, migration and invasion
in vitro and inhibits lung metastasis in vivo by
targeting HMGA2, SALL4 and Twist1. Ahmad et al (26) found that miR-20b expression was
significantly higher in brain metastases of breast cancer patients,
compared to primary breast tumors as well as the patients without
brain metastasis, and that miR-20b significantly induced the colony
formation and invasiveness of breast cancer cells. Li et al
(27) found that ectopic
expression of miR-153 could significantly inhibit tumor growth and
impair the migration and invasion of breast cancer cells by
regulating ETM targeting metadherin (MTDH).
In the present study, to our knowledge, we first
report that miR-338-3p was downregulated in breast tumor samples
from patients compared with adjacent normal breast tissues.
miR-338-3p expression was inversely correlated with clinical stages
and metastatic status of breast cancer. Moreover, overexpression of
miR-338-3p in breast cancer cells inhibited cell proliferation and
migration and invasion in vitro and suppressed tumor growth
in vivo. These findings suggested that miR-338-3p may exert
tumor-suppressive functions and impede breast tumor growth and
metastasis.
Accumulating evidence firmly demonstrates that
miRNAs control various key cellular processes such as
proliferation, apoptosis, differentiation, metastasis, play
important roles in carcinogenesis and tumor progression functioning
as oncogene or tumor suppressor gene (28). miR-338-3p, a recently discovered
miRNA, functions as tumor suppressor in a wide range of human
malignances, including hepatocellular carcinoma, neuroblastoma,
ovarian cancer, malignant melanoma, gastric cancer and colorectal
cancer (13–21). Previously, the role of miR-338-3p
in breast cancer was poorly explored. In agreement with the above
study, we found that the overexpression of miR-338-3p in breast
cancer cells significantly inhibited cell proliferation, colony
formation, migration and invasion, and induced cell apoptosis and
cell cycle arrest at G1/G0 stage, as well as suppressed tumor
growth of breast cancer in a nude mouse model. Together with our
results, these data suggest that miR-338-3p may have potential to
serve as a tumor suppressor miRNA in various cancers including
breast cancer.
In view of the vital importance of miR-338-3p, we
further explored the molecular mechanisms underlying breast cancer
cell biological behavior by the regulation of miR-338-3p. We
selected TargetScan and miRanda algorithm to search for putative
protein-coding gene targets of miR-338-3p, especially for those
that have the ability to promote tumor cell proliferation,
migration and invasion. Based on this rationale, SOX4 was selected
as the potential target for further validation, since it has been
shown that deregulated expression of SOX4 is correlated with
increased cancer cell proliferation, cell survival, inhibition of
apoptosis and tumor progression (29,30).
Sex-determining region Y-box 4 (SOX4), located at
chromosome 6p22.3, is a member of the highly conserved SoxC
(SRY-related high-motility group box) transcription factor family,
which contains two other members, SOX11 and SOX12 (31). Notably, SOX4 has been recognized as
one of the 64 cancer signature genes (29,30).
Indeed, it is overexpressed in several types of cancer including
breast cancer (32). Genome-wide
chromatin immunoprecipitation studies have uncovered that SOX4
regulates the transcription of genes involved in TGF-β, Wnt,
Hedgehog, and Notch pathways and components of miRNA processing
machinery including Dicer, Argonaute 1 and RNA Helicase A (33,34).
More recently it was shown that SOX4 induces EMT via the polycomb
epigenetic regulator EZH2 (35).
Importantly, SOX4 has been found to be regulated by several miRNAs
such as miR-335 (36), miR-31
(37), miR-129 family (38), miR-212 (39) and miR-138 (40). Here, we first confirmed that SOX4
is a target of miR-338-3p by luciferase assay, and that
upregulation of miR-338-3p decreased the expression of SOX4 on mRNA
level and protein level. Our results also showed that SOX4
expression is upregulated in breast cancer tissue, and that high
SOX4 expression was associated with low miR-338-3p levels in breast
cancer. These finding might suggest that miR-338-3p inhibited
breast cancer growth and metastasis by targeting SOX4.
In conclusion, the present study demonstrated that
miR-338-3p expression is downregulated in breast tumor samples and
breast cancer cell lines and is inversely correlated with TNM stage
and lymph node metastatic status. miR-338 functions as tumor
suppressor, block breast cancer cell growth and metastasis in
vitro and in vivo by targeting SOX4. These finding
suggested that miR-338-3p may serve as a new diagnostic and
therapeutical agent for breast cancer.
Acknowledgements
This study was supported by the Health Department of
Jilin Province (2010SO20).
References
|
1
|
DeSantis C, Ma J, Bryan L and Jemal A:
Breast cancer statistics, 2013. CA Cancer J Clin. 64:52–62. 2014.
View Article : Google Scholar
|
|
2
|
Hong W and Dong E: The past, present and
future of breast cancer research in China. Cancer Lett. 351:1–5.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Berse B and Lynch JA: Molecular diagnostic
testing in breast cancer. Semin Oncol Nurs. 31:108–121. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fabian MR, Sonenberg N and Filipowicz W:
Regulation of mRNA translation and stability by microRNAs. Annu Rev
Biochem. 79:351–379. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Guo H, Ingolia NT, Weissman JS and Bartel
DP: Mammalian microRNAs predominantly act to decrease target mRNA
levels. Nature. 466:835–840. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McManus MT: MicroRNAs and cancer. Semin
Cancer Biol. 13:253–258. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Farazi TA, Spitzer JI, Morozov P and
Tuschl T: miRNAs in human cancer. J Pathol. 223:102–115. 2011.
View Article : Google Scholar :
|
|
9
|
Tang J, Ahmad A and Sarkar FH: The role of
microRNAs in breast cancer migration, invasion and metastasis. Int
J Mol Sci. 13:13414–13437. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Christodoulatos GS and Dalamaga M:
Micro-RNAs as clinical biomarkers and therapeutic targets in breast
cancer: Quo vadis? World J Clin Oncol. 5:71–81. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schrauder MG, Strick R, Schulz-Wendtland
R, Strissel PL, Kahmann L, Loehberg CR, Lux MP, Jud SM, Hartmann A,
Hein A, et al: Circulating micro-RNAs as potential blood-based
markers for early stage breast cancer detection. PLoS One.
7:e297702012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Le Quesne J and Caldas C: Micro-RNAs and
breast cancer. Mol Oncol. 4:230–241. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang XH, Chen JS, Wang Q, Chen XL, Wen L,
Chen LZ, Bi J, Zhang LJ, Su Q and Zeng WT: miR-338–3p suppresses
invasion of liver cancer cell by targeting smoothened. J Pathol.
225:463–472. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fu X, Tan D, Hou Z, Hu Z, Liu G, Ouyang Y
and Liu F: The effect of miR-338-3p on HBx deletion-mutant
(HBx-d382) mediated liver-cell proliferation through CyclinD1
regulation. PLoS One. 7:e432042012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen X, Pan M, Han L, Lu H, Hao X and Dong
Q: miR-338-3p suppresses neuroblastoma proliferation, invasion and
migration through targeting PREX2a. FEBS Lett. 587:3729–3737. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wen C, Liu X, Ma H, Zhang W and Li H:
miR-338-3p suppresses tumor growth of ovarian epithelial carcinoma
by targeting Runx2. Int J Oncol. 46:2277–2285. 2015.PubMed/NCBI
|
|
17
|
Caramuta S, Egyházi S, Rodolfo M, Witten
D, Hansson J, Larsson C and Lui WO: MicroRNA expression profiles
associated with mutational status and survival in malignant
melanoma. J Invest Dermatol. 130:2062–2070. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li P, Chen X, Su L, Li C, Zhi Q, Yu B,
Sheng H, Wang J, Feng R, Cai Q, et al: Epigenetic silencing of
miR-338-3p contributes to tumorigenicity in gastric cancer by
targeting SSX2IP. PLoS One. 8:e667822013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guo B, Liu L, Yao J, Ma R, Chang D, Li Z,
Song T and Huang C: miR-338-3p suppresses gastric cancer
progression through a PTEN-AKT axis by targeting P-REX2a. Mol
Cancer Res. 12:313–321. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun K, Su G, Deng H, Dong J, Lei S and Li
G: Relationship between miRNA-338-3p expression and progression and
prognosis of human colorectal carcinoma. Chin Med J (Engl).
127:1884–1890. 2014.
|
|
21
|
Sun K, Deng HJ, Lei ST, Dong JQ and Li GX:
miRNA-338-3p suppresses cell growth of human colorectal carcinoma
by targeting smoothened. World J Gastroenterol. 19:2197–2207. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang L and Wang J: MicroRNA-mediated
breast cancer metastasis: From primary site to distant organs.
Oncogene. 31:2499–2511. 2012. View Article : Google Scholar
|
|
23
|
Kusama M, Kaise H, Nakayama S, Ohta D,
Aoki T and Koyanagi Y: A case of breast cancer patient of CAF
(cyclophosphamide, adriamycin, 5-fluorouracil) resistant lung
metastasis with remarkable response to reverse drug-resistance by
toremifene. Gan To Kagaku Ryoho. 26:1171–1175. 1999.(In Japanese).
PubMed/NCBI
|
|
24
|
Tang J, Ahmad A and Sarkar FH: The role of
microRNAs in breast cancer migration, invasion and metastasis. Int
J Mol Sci. 13:13414–13437. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin Y, Liu AY, Fan C, Zheng H, Li Y, Zhang
C, Wu S, Yu D, Huang Z, Liu F, et al: MicroRNA-33b inhibits breast
cancer metastasis by targeting HMGA2, SALL4 and Twist1. Sci Rep.
5:99952015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ahmad A, Ginnebaugh KR, Sethi S, Chen W,
Ali R, Mittal S and Sarkar FH: miR-20b is up-regulated in brain
metastases from primary breast cancers. Oncotarget. 6:12188–12195.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li W, Zhai L, Zhao C and Lv S: MiR-153
inhibits epithelial-mesenchymal transition by targeting metadherin
in human breast cancer. Breast Cancer Res Treat. 150:501–509. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yi B, Piazza GA, Su X and Xi Y: MicroRNA
and cancer chemo-prevention. Cancer Prev Res (Phila). 6:401–409.
2013. View Article : Google Scholar
|
|
29
|
Vervoort SJ, Lourenço AR, van Boxtel R and
Coffer PJ: SOX4 mediates TGF-β-induced expression of mesenchymal
markers during mammary cell epithelial to mesenchymal transition.
PLoS One. 8:e532382013. View Article : Google Scholar
|
|
30
|
Vervoort SJ, van Boxtel R and Coffer PJ:
The role of SRY-related HMG box transcription factor 4 (SOX4) in
tumorigenesis and metastasis: Friend or foe? Oncogene.
32:3397–3409. 2013. View Article : Google Scholar
|
|
31
|
Bowles J, Schepers G and Koopman P:
Phylogeny of the SOX family of developmental transcription factors
based on sequence and structural indicators. Dev Biol. 227:239–255.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Song GD, Sun Y, Shen H and Li W: SOX4
overexpression is a novel biomarker of malignant status and poor
prognosis in breast cancer patients. Tumour Biol. Jan 16–2015.(Epub
ahead of print] 2015. View Article : Google Scholar
|
|
33
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
Large-scale meta-analysis of cancer microarray data identifies
common transcriptional profiles of neoplastic transformation and
progression. Proc Natl Acad Sci USA. 101:9309–9314. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Scharer CD, McCabe CD, Ali-Seyed M, Berger
MF, Bulyk ML and Moreno CS: Genome-wide promoter analysis of the
SOX4 transcriptional network in prostate cancer cells. Cancer Res.
69:709–717. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tiwari N, Tiwari VK, Waldmeier L, Balwierz
PJ, Arnold P, Pachkov M, Meyer-Schaller N, Schübeler D, van
Nimwegen E and Christofori G: Sox4 is a master regulator of
epithelial-mesenchymal transition by controlling Ezh2 expression
and epigenetic reprogramming. Cancer Cell. 23:768–783. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tavazoie SF, Alarcón C, Oskarsson T, Padua
D, Wang Q, Bos PD, Gerald WL and Massagué J: Endogenous human
microRNAs that suppress breast cancer metastasis. Nature.
451:147–152. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Koumangoye RB, Andl T, Taubenslag KJ,
Zilberman ST, Taylor CJ, Loomans HA and Andl CD: SOX4 interacts
with EZH2 and HDAC3 to suppress microRNA-31 in invasive esophageal
cancer cells. Mol Cancer. 14:242015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yu X, Song H, Xia T, Han S, Xiao B, Luo L,
Xi Y and Guo J: Growth inhibitory effects of three miR-129 family
members on gastric cancer. Gene. 532:87–93. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Luo XJ, Tang DG, Gao TL, Zhang YL, Wang M,
Quan ZX and Chen J: MicroRNA-212 inhibits osteosarcoma cells
proliferation and invasion by down-regulation of Sox4. Cell Physiol
Biochem. 34:2180–2188. 2014. View Article : Google Scholar
|
|
40
|
Yeh YM, Chuang CM, Chao KC and Wang LH:
MicroRNA-138 suppresses ovarian cancer cell invasion and metastasis
by targeting SOX4 and HIF-1α. Int J Cancer. 133:867–878. 2013.
View Article : Google Scholar : PubMed/NCBI
|