Introduction
Melanoma is an aggressive disease with an
ever-increasing incidence rate and is the deadliest known form of
skin cancer. In 2015, an estimated 80,100 new cases and 13,340
deaths from skin cancer are expected in the United States, with
melanoma accounting for 75% of all skin cancer deaths (1). Although targeted therapies and
immunotherapies have revolutionized melanoma treatment and improved
the quality of life of advanced melanoma patients, few drugs are
available (only seven as of 2014), and drug resistance is
increasing (2). Therefore, further
study of the progression of metastatic melanoma and the innovation
of new, targeted therapies are desperately needed.
Long non-coding RNAs (lncRNAs) are RNA molecules
with lengths of more than 200 nucleotides and little probability of
translation into a protein (3).
Considered useless and subsequently ignored for several decades,
lncRNA is now attracting more and more attention, and increasing
evidence has suggested that lncRNAs participate in universal
biological processes such as cell differentiation, proliferation,
migration and invasion (4).
Studies have also revealed that some lncRNAs, such as HOTAIR and
growth arrest specific transcript 5 (GAS5), can function as tumor
promoters or suppressors, respectively (5,6).
Other lncRNAs function by providing a scaffold through which
proteins can adhere, accumulate and exert their functions,
suggesting that specific RNA secondary structures are needed for
lncRNAs to carry out their functions (7,8).
Located at 1q25 and with a mature length of 650 bases, the lncRNA
growth arrest-specific transcript 5 (GAS5) comprises 12 exons and
encodes 10 box C/D snoRNAs within its introns (9). GAS5 was originally isolated from NIH
3T3 cells by Schneider et al with the purpose of identifying
potential tumor suppressor genes enriched during growth arrest
induced by serum starvation (10).
GAS5 transcript levels are significantly reduced in a spectrum of
cancers including breast, prostate, glioblastoma, renal clear cell,
bladder, hepatocellular, pancreatic, non-small cell lung,
colorectal, cervical and multiple myeloma (11–21).
Importantly, aberrant GAS5 expression was significantly correlated
with metastasis in cervical cancer (21) and hepatocellular carcinoma
(15).
However, despite the past research, the expression
profile and biological role of GAS5 and its mechanism in melanoma
metastasis remain largely unknown. The tumorigenesis process and
progression of melanoma to metastasis is generally recognized as a
multistep process in which the cellular and molecular mechanisms
change. The first step of invasion, which is the degradation of the
extracellular matrix, is regulated by the matrix metalloproteinase
(MMP) family, which plays a key role in tumor cell invasion and
metastasis (22). MMP2, which was
shown to be specifically involved in type IV collagen degradation,
is also secreted by a wide range of melanoma cells (23). Specifically, MMP2 is detected in
the early stages of melanocytic tumor progression (24). Although MMP13 is also gelatinolytic
and type IV collagenolytic, it is prone to degrading type I, II and
III collagens (25). Previous
studies have shown that upregulated MMP13 and its activity in
melanoma cell lines are involved in the metastasis of melanoma
(26,27). A recent study revealed that MMP13
not only cleaves laminin-5 into small fragments to accelerate tumor
metastasis but also disrupts vasculogenic mimicry in the presence
of cleaved laminin-5, exerting dual effects on melanoma (28).
The SK-Mel-110 melanoma cell line, isolated from a
primary melanoma patient, has been cultured in vitro for
nearly 30 years and has been divided many times, which is
indicative of an immortal cell line (29). SK-Mel-110 cells grow actively, with
a doubling time of ~20 h, and are tumorigenic in immunodeficient
nude mice (30). They have been
used widely at several melanoma research centers for the study of
melanogenesis, which investigates the role that autophagy and
apoptotic resistance play in the pathogenesis of melanoma, and in
various pharmacological and toxicological studies (31–33).
In the present study, substantial downregulation of GAS5 was
detected in SK-Mel-110 cells, and therefore it was selected for
further study.
Based on these reports, we hypothesized that GAS5
may function as a tumor suppressor by inhibiting melanoma cell
invasiveness via the regulation of MMP2 or MMP13. To further
understand the role of GAS5 in the metastasis of melanoma, we
overexpressed GAS5 in SK-Mel-110 cells. Using this model, the
effects of GAS5 on melanoma migration and invasion, and the
potential correlation between GAS5 and MMPs, were investigated.
Materials and methods
Cell culture
The human melanoma cell line A375 was purchased from
ATCC (Manassas, VA, USA) and Hacat, SK-Mel-110, SK-Mel-28 and M21
were purchased from the Shanghai Institutes for Biological Science
(SIBS), China Academy of Science (CAS). Cells were cultured in
Dulbecco's modified Eagle's medium (DMEM, Invitrogen, Carlsbad, CA,
USA) supplemented with 5% fetal bovine serum (FBS, Life
Technologies, Burlington, Ontario, Canada), 100 U/ml penicillin and
100 μg/ml streptomycin at 37°C in 5% (v/v) CO2. Cells
were maintained without exceeding 0.5×106 cells/ml. 293T
human embryonic kidney (HEK) cells were obtained from the Cell Bank
at the Chinese Academy of Science and cultured in DMEM containing
10% (v/v) FBS and 100 μg/ml streptomycin at 37°C in 5% (v/v)
CO2.
Predicting secondary structure of
GAS5
Secondary structural prediction was performed using
the RNA Vienna Package on the webserver (http://rna.tbi.univie.ac.at/) (34,35).
This program predicts RNA secondary structure based on minimum
energy requirements and pair probabilities.
Total RNA extraction and detection of
GAS5 by qRT-PCR
Total RNA was isolated from the transfected cells
using TRIzol reagent (Invitrogen) according to the manufacturer's
protocol. Briefly, after removing the culture medium, TRIzol
reagent was added to each well, followed by incubation at room
temperature for 10 min and centrifugation at 12,000 g for 10 min.
The supernatant was removed to obtain the precipitated RNA. After
washing with 75% ethanol, the dried RNA pellet was then resuspended
in RNase-free water and stored at −80°C until use. cDNA was
synthesized using Oligo(dT) and M-MLV reverse transcriptase (M1701,
Promega). GAS5 primers (F, 5′-GCACACAGGCATTAGAC AGA-3′; R,
5′-AAGCCGACTCTCCATACCTT-3′), and the U6 primers (F,
5′-CTCGCTTCGGCAGCACA-3′; R, 5′-AACG CTTCACGAATTTGCGT-3′) were
synthesized by Shengon (Shanghai, China). Real-time PCR was
performed using a sequence detection system (ABI 7300; Applied
Biosystems, Foster City, CA, USA). The 20-μl reaction mixture
contained 10 μl of SYBR green PCR master mix (A6020, Promega), 1 μl
of cDNA template, 1 μl of PCR forward primer, 1 μl of PCR reverse
primer and 7 μl of RNA-free water. PCR running conditions were as
follows: 15 sec at 95°C for the initial denaturation, 30 cycles of
5 sec at 95°C and 30 sec at 60°C and 1 min at 95°C for annealing.
The threshold cycle (Ct value), which is the cycle number at which
the amount of amplified gene of interest reaches a fixed threshold,
was subsequently determined. GAS5 gene expression was compared to
control cells by qRT-PCR using the 2−ΔΔCt method with U6 as an
internal control. The results are expressed as the mean ± SD from
three independent experiments.
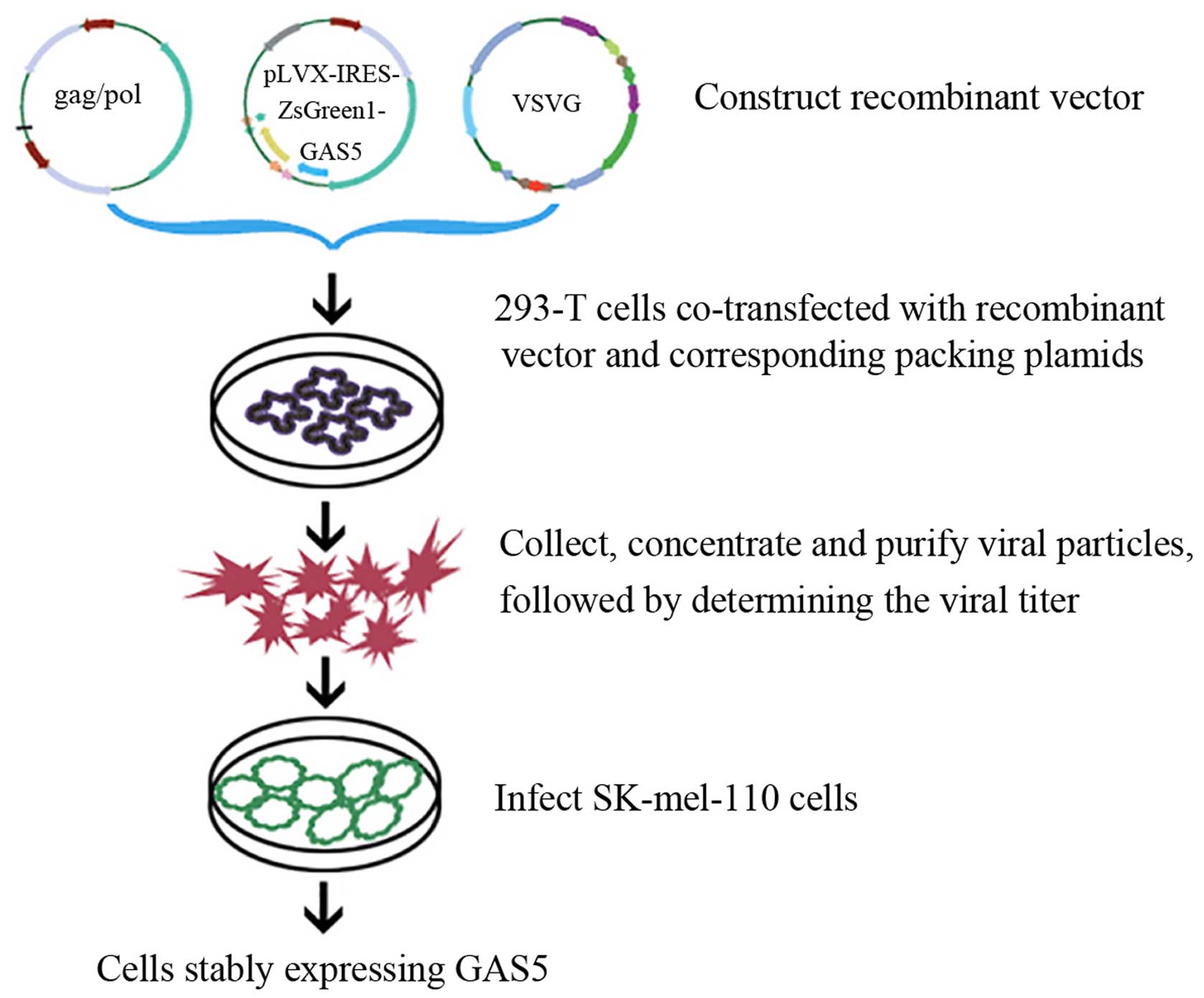
Constructing a recombinant vector
containing the GAS5 gene
The mature human GAS5 gene (NM_002578.2) was
synthesized by Genewiz (Suzhou, China). The primers are as follows:
GAS5-XhoI-F, 5′-ccgctcgagTTTCGAGGTAGGA
GTCGACTCCTGTG-3′; and GAS5-BamHI-R, 5′-cgcggatcc
TTTTTTTTTTTTTTTTTTTTGGATTGCAAA-3′. These primers contain both
XhoI and BamHI restriction sites (underlined). The
25-μl reaction mixture contained 2.5 μl of 2 mM dNTP mixture, 2.5
μl of 10X KOD buffer, 1.5 μl of 25 mM MgSO4, 0.5 μl of
synthesized template, 0.3 μl of PCR forward primer, 0.3 μl of PCR
reverse primer, 0.3 μl of KOD Plus Neo and 17.1 μl of RNA-free
water. PCR running conditions were as follows: 3 min at 94°C for
the initial denaturation, 30 cycles of 15 sec at 98°C and 15 sec at
58°C and 30 sec at 95°C for annealing. The resulting product was
kept at 16°C until further use. After isolation and purification of
the PCR product, the GAS5 gene was inserted into the corresponding
site of the linearized expression vector pLVX-IRES-ZsGreen1 (data
not shown), downstream of the cytomegalovirus (CMV) promoter. The
recombinant plasmid was then transformed into DH5α competent cells
(36,37) (Fig.
1).
Identifying positive recombinant vectors,
packaging lentivirus and determining lentivirus titers
Positive recombinant clones were examined and
verified by DNA sequencing (data not shown). To generate
lentiviruses, three individual lentiviral vectors were required:
the pLVX-IRES-ZsGreen1-GAS5 vector, the pHelper 1.0 vector (gag/pol
element) and the pHelper 2.0 vector (VSVG element). Lentiviruses
were generated in HEK293T cells as described previously, with minor
modifications (38). In brief, 20
μg of pLVX-IRES-ZsGreen1-GAS5 vector, 10 μg of pHelper 1.0 and 10
μg of pHelper 2.0 were cotransfected into 293T cells with the help
of a transfection reagent (Lipofectamine 2000, Invitrogen).
Seventy-two hours after transfection, supernatants were removed and
filtered (0.45-μm filter, Millipore). Virus was collected by
ultracentrifugation for 60 min at 20,000 rpm (4°C) and stored at
−80°C until further use. The virus-containing supernatant was
titrated onto SK-Mel-110 cells to determine the titers needed to
transduce 95% of the cells (6×106 transduction
units/ml). Mock viruses were generated by the same procedure using
otherwise identical vectors lacking GAS5 cDNA.
The GAS5 gene was observed as a band between 500–750
bp, which correlated with the expected size of 651 bp (data not
shown). DNA sequencing confirmed that the tested sequence was
identical to that in GenBank/NCBI (data not shown).
Establishing stable GAS5 overexpression
in human melanoma cells
For transduction, SK-Mel-100 cells were inoculated
in a 24-well plate and cultured until they reached 50–60%
confluence. The experimental virus solution,
pLVX-IRES-ZsGreen1-GAS5 (GAS5 overexpression group), and the
control virus solution, ZsGreen 1 (control group), were thawed on
ice and added to the culture at a multiplicity of infection (MOI)
of 5 for 72 h in the presence of polybrene (Santa Cruz Biotech).
Infection was carried out in humidified air containing 5%
CO2 at 37°C. ZsGreen1 expression was visualized using a
fluorescence microscope. The fluorescent signal indicating ZsGreen1
expression in the stably-transfected cells was observed after
repeated freeze-thaw cycles to determine the stability of the
transfected genes, after which the experimental procedures were
performed.
RNA fluorescence in situ
hybridization
Oligonucleotide modified probes for human lncRNA
GAS5 (5′-TAMRA-CAGGAGCAGAACCATTAAGCTGGTCCAGGCAAGT-3′-TAMRA) and a
negative/scramble control were used for RNA fluorescence in
situ hybridization (FISH). Procedures were carried out based on
a previous study (39), with
slight modification. Briefly, the cell suspension was pipetted onto
autoclaved glass slides. The following day, the slides were washed
in PBS and fixed in 4% paraformaldehyde. To permeabilize the cell
membrane, 0.2 M HCl was used. The slides were then treated with the
protease reagent; this activity was stopped by the addition of
glycine. After dehydration with 70, 85 and 100% ethanol,
hybridization was performed at 37°C overnight in a dark moist
chamber. After hybridization, slides were washed three times in 50%
formamide/2X SSC for 5 min each at room temperature. These slides
were counterstained with DAPI. The images were acquired using a
confocal microscope.
Cell mobility and invasion using a
Transwell assay
Cell mobility was examined using a 24-well chamber
plate containing a polycarbonate membrane with an 8-μm pore size.
DMEM with 10% FBS (0.8 ml) served as a chemoattractant in the lower
chamber. First, 1×104 cells in 0.2 ml of serum-free
medium were added to the inserts and incubated for 24 h at 37°C and
5% CO2. Cells remaining on the upper membrane surface of
the inserts were then removed with a cotton swab, and the total
number of cells that migrated into the lower chamber was counted. A
cell invasion assay was performed in a similar manner to the cell
motility assay with some modifications. Briefly, 5×104
cells in serum-free DMEM were added to the upper chambers of
transwells with 8-μm pore size polycarbonate membranes and
precoated with a thin layer of Matrigel. DMEM with 10% FBS was used
as a chemoattractant in the lower chamber. The total numbers of
cells in the lower chamber were counted after 36 h of incubation at
37°C with 5% CO2.
Western blotting to demonstrate MMP2 and
MMP13 protein expression
For western blots, cell lysates and culture
supernatants were prepared using RIPA buffer (50 mM Tris-HCl, pH
7.4) with 1 mM PMSF and incubated at 4°C for 30 min. The lysates
were centrifuged at 12,000 rpm for 10 min at 4°C. Protein
concentrations were determined using a NanoDrop ND-1000
spectrophotometer (Thermo Scientific). Samples (20 μg of protein)
were separated by SDS-PAGE using 5% stacking and 12% separating
gels and subsequently transferred onto a PVDF membrane (Millipore).
After blocking in Tris-buffer (50 mM Tris, pH 7.5) containing 5%
skim milk, the membrane was incubated overnight at 4°C with
antibodies to detect MMP2 (ab110186, 1:2000; Abcam), MMP13
(ab39012, 1:1000; Abcam) and GAPDH (Ab8227, 1:1000; Abcam),
followed by incubation for 1 h at room temperature with a
corresponding HRP-conjugated goat anti-rabbit IgG secondary
antibody (sc-2004, 1:1000; Santa Cruz). Protein expression was
visualized after extensive washing using an enhanced
chemiluminescence advanced detection kit (34077, Thermo Scientific)
and quantified with analysis software (Quantity One; Bio-Rad
Laboratories Inc., Hercules, CA, USA).
MMP2 activity assay
MMP2 activity in cell extracts and culture
supernatants was detected using an MMP2 activity assay kit
(GMS50070.1, Genmed, Shanghai, China) according to the
manufacturer's instructions. For this assay, MMP2 activity was
detected by the fluorescence resonance energy transfer method
(FRET), which involves the transfer of excited energy from a donor
(with a short wavelength) to an acceptor (with a long wavelength)
so that the emitted photon from the donor, whose reaction is
dependent on proximity to the acceptor, is quenched. The donor
fluorescent probe, 7-methoxy-coumarin, labeled with the peptide
substrate PLGLAR, was quenched by the acceptor using dinitrobenzene
diamino isobutyryloxy. With the hydrolysis of the polypeptide
Gly-Leu bond by MMP2, the strongly fluorescent 7-methoxycoumarin
polypeptide fragment was released. Based on this fluorescence
intensity (excited at 330 nm and emitted at 400 nm) and standard
curve, we obtained the 7-methoxycoumarin concentration. The
specific activity of MMP2 was then calculated using the following
formula: MMP2 activity (nmol/mg/min) = 7-methoxycoumarin
concentration (μmol/l) × dilution times ÷ 5 min ÷ the concentration
of protein sample (mg/ml).
Statistical analysis
Data are expressed as the mean ± standard deviation
of triplicate samples. Statistical analysis was performed using
Student's t-test. Differences were considered to be statistically
significant at P<0.05 (two-tailed). All statistical analysis was
performed with GraphPad Prism version 6.0e for Mac OS X (GraphPad
Software, La Jolla, CA, USA).
Results
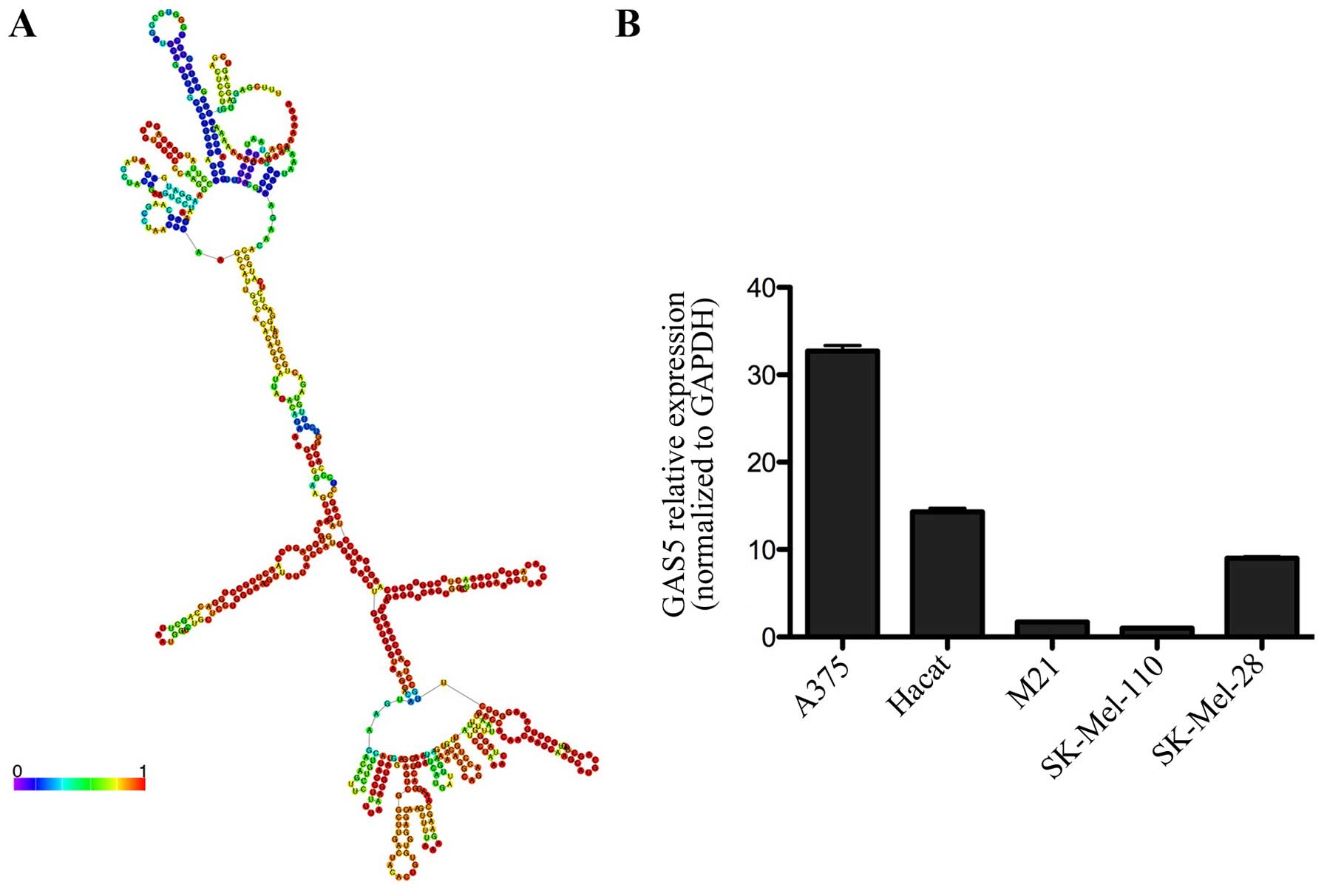
The secondary structure of GAS5 and its
mRNA levels in the SK-Mel-110 cell line
We first predicted the secondary structure of GAS5
using RNAfold and found that it contains a complex stem-loop
structure, indicating that GAS5 may perform a complex and important
biological function (Fig. 2A).
Next, we examined lncRNA GAS5 expression levels in HaCaT, A375,
SK-Mel-28, SK-Mel-110 and M21 cells by qRT-PCR. After normalizing
to GAPDH, the GAS5 expression level in SK-Mel-110 cells was
significantly lower than that detected in other melanoma cell lines
(Fig. 2B).
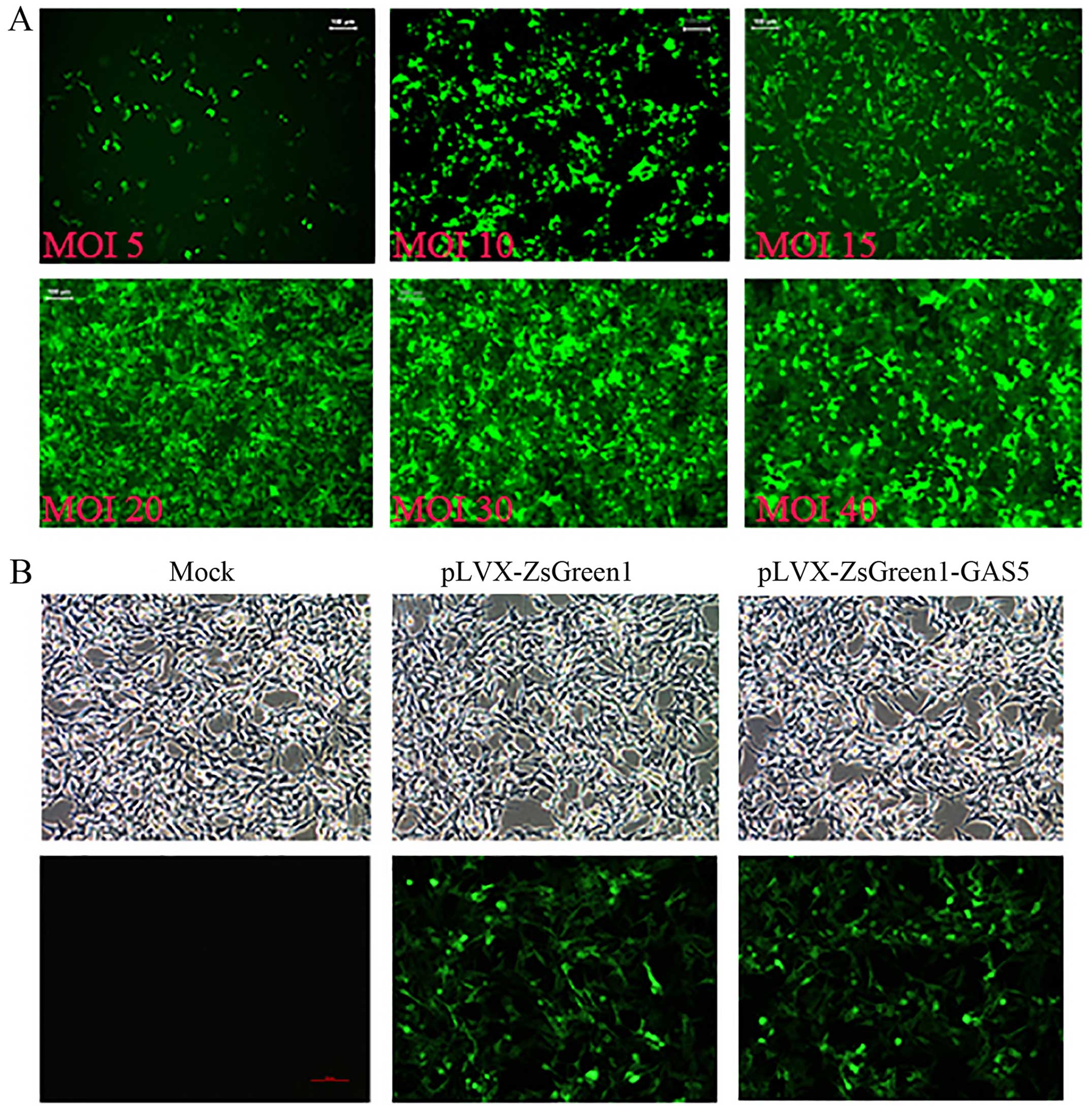
Transfection efficiency and selection of
a stably expressing GAS5 cell line
To investigate GAS5 function in SK-Mel-110
cell lines, we overexpressed this gene by infecting SK-Mel-110
cells with pLVX-IRES-ZsGreen1-GAS5-containing lentivirus. The
GAS5 fragment was amplified by reverse transcription
polymerase chain reaction (RT-PCR) and verified by DNA sequencing.
The use of different MOIs was designed to verify the most efficient
dose of virus. After infection, green fluorescence in the cytoplasm
of SK-Mel-110 cells was observed by a fluorescence microscope, and
the efficiency of the infection, or the fraction of SK-Mel-110
cells with ZsGreen1 fluorescence, was quantified. As the number of
viral particles increased, the number of SK-Mel-110 cells with
ZsGreen1 fluorescence also increased. When the MOI reached >40,
the number of green fluorescent SK-Mel-110 cells did not increase
significantly (Fig. 3A).
Consequently, a MOI of 40 was chosen for subsequent experiments.
The transfection efficiency was calculated as the ratio of the
number of fluorescence-positive cells to the total number of cells.
Cells were transfected with pLVX-IRES-ZsGreen1-GAS5-free or
transduced with GAS5-containing virus at a MOI of 40; 95% of
cells maintained these plasmids after 3 passages (Fig. 3B).
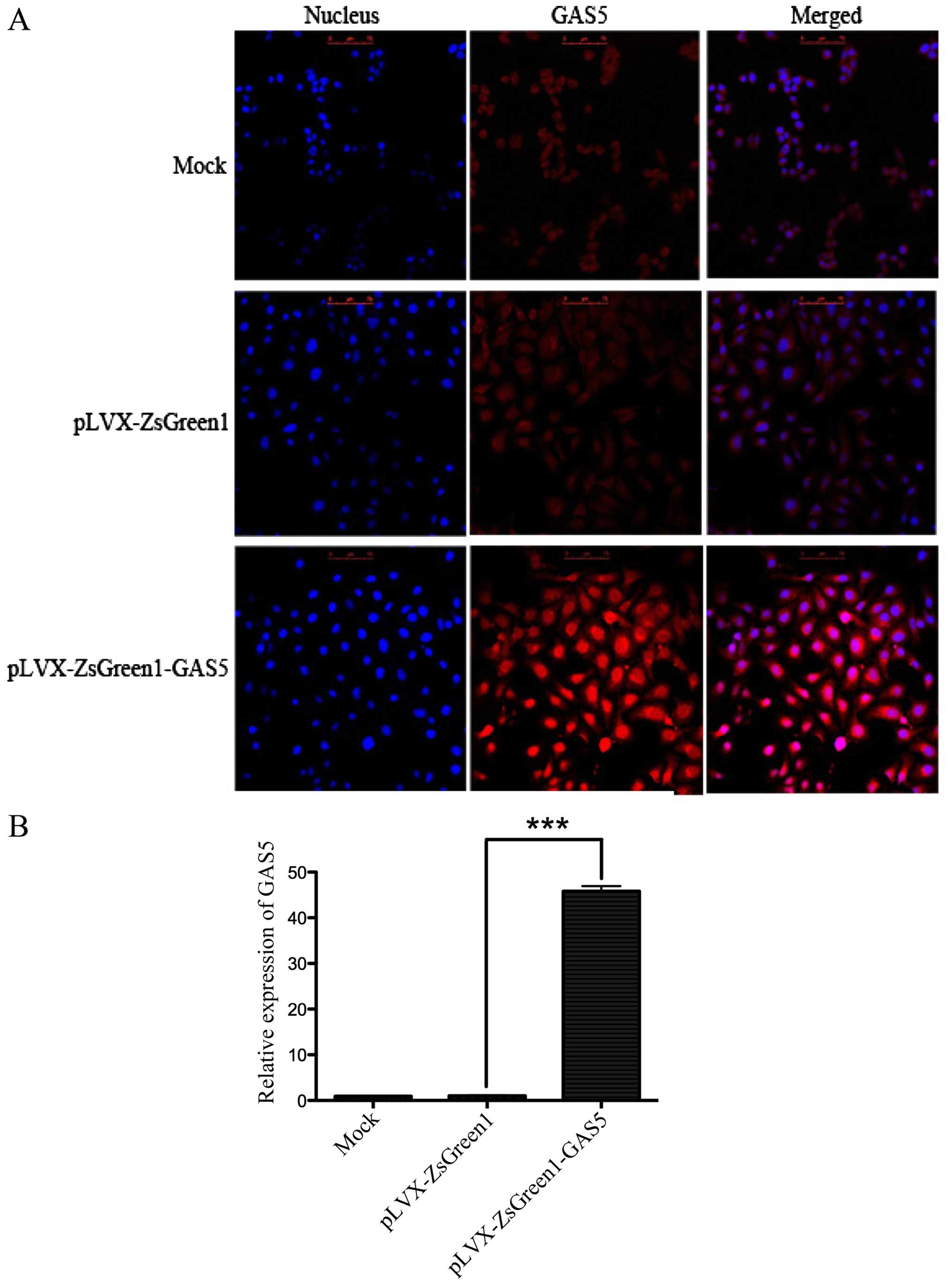
GAS5 localizes within both the nucleus
and cytoplasm
Expression of GAS5 mRNA from wild-type and
overexpressed GAS5 SK-Mel-110 cells was determined by FISH and
real-time PCR. FISH showed that GAS5 is mainly localized to the
cytoplasm in untreated cells but accumulates in the nucleus in
GAS5-overexpressed SK-Mel-110 cells (Fig. 4A). The efficiency of the lentivirus
transductions was also confirmed by examining GAS5 mRNA levels via
real-time PCR. The relative expression of GAS5 mRNA was remarkably
increased (45-fold) in GAS5-overexpressing SK-Mel-110 cells,
compared with levels in the negative pLVX-ZsGreen1 and mock control
groups (ANOVA; F=1535, P<0.01) (Fig. 4B).
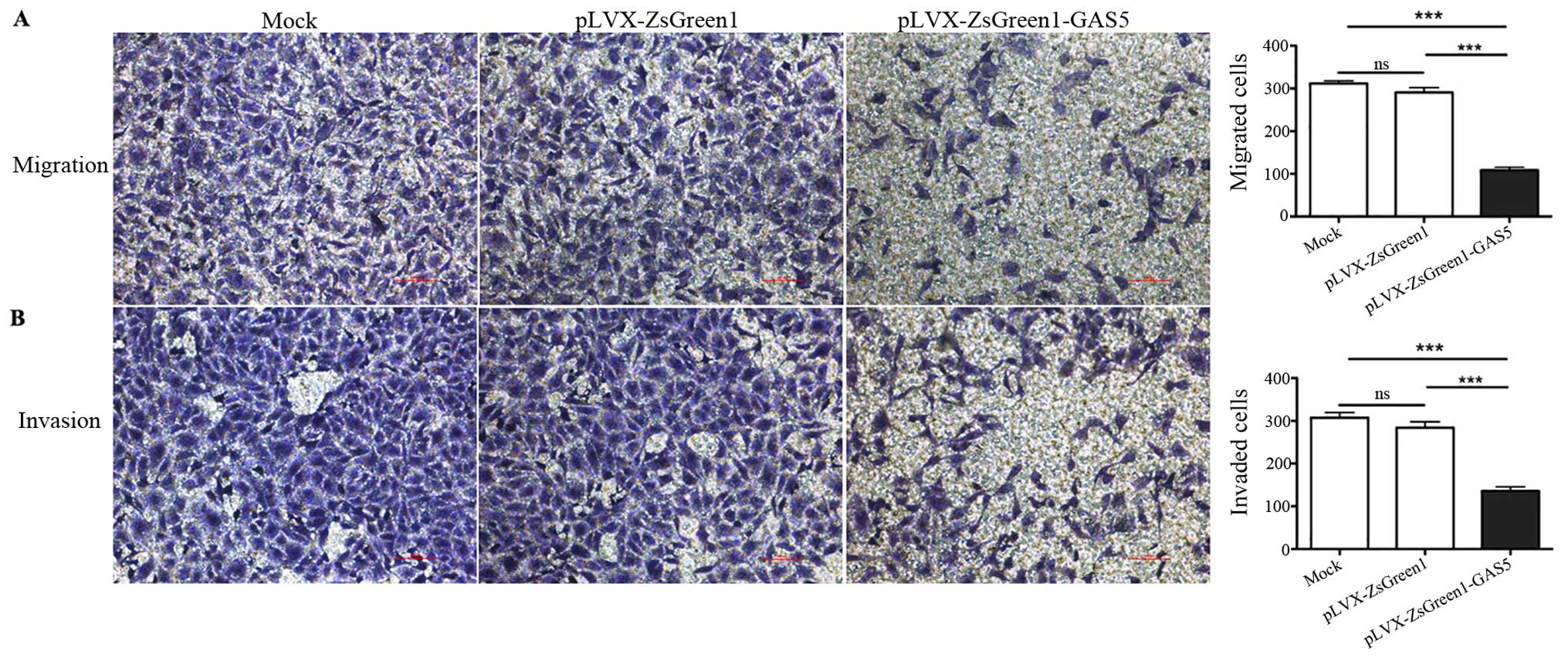
GAS5 inhibits the migration and invasion
of SK-Mel-110 cells
Both migration and invasion play crucial roles in
the metastasis of malignant tumors, which is the main driver behind
cancer-related deaths. A classic transwell chamber assay was
performed to examine the role of GAS5 in regulating the migratory
ability of the melanoma cells. Compared to mock and pLVX-ZsGreen1
cells, pLVX-ZsGreen1-GAS5 cells showed a significant decrease in
migratory ability (65.3%, P<0.001, Fig. 5A). Furthermore, the effects of GAS5
on cell invasion were investigated using a Transwell invasion
assay. Three types of cells were cultured in Boyden chambers
covered with a Matrigel-coated polycarbonate membrane. Twenty-four
hours later, pLVX-ZsGreen1-GAS5 cells migrated through the
Matrigel, and a significant decrease in migrated cells was observed
in these chambers compared to chambers containing mock and
pLVX-ZsGreen1 cells (55.6%, P<0.001, Fig. 5B). Taken together, these results
indicate that overexpressed GAS5 not only reduces the migration of
melanoma cells but also inhibits their ability to invade a matrix,
reminiscent of their movement during metastasis.
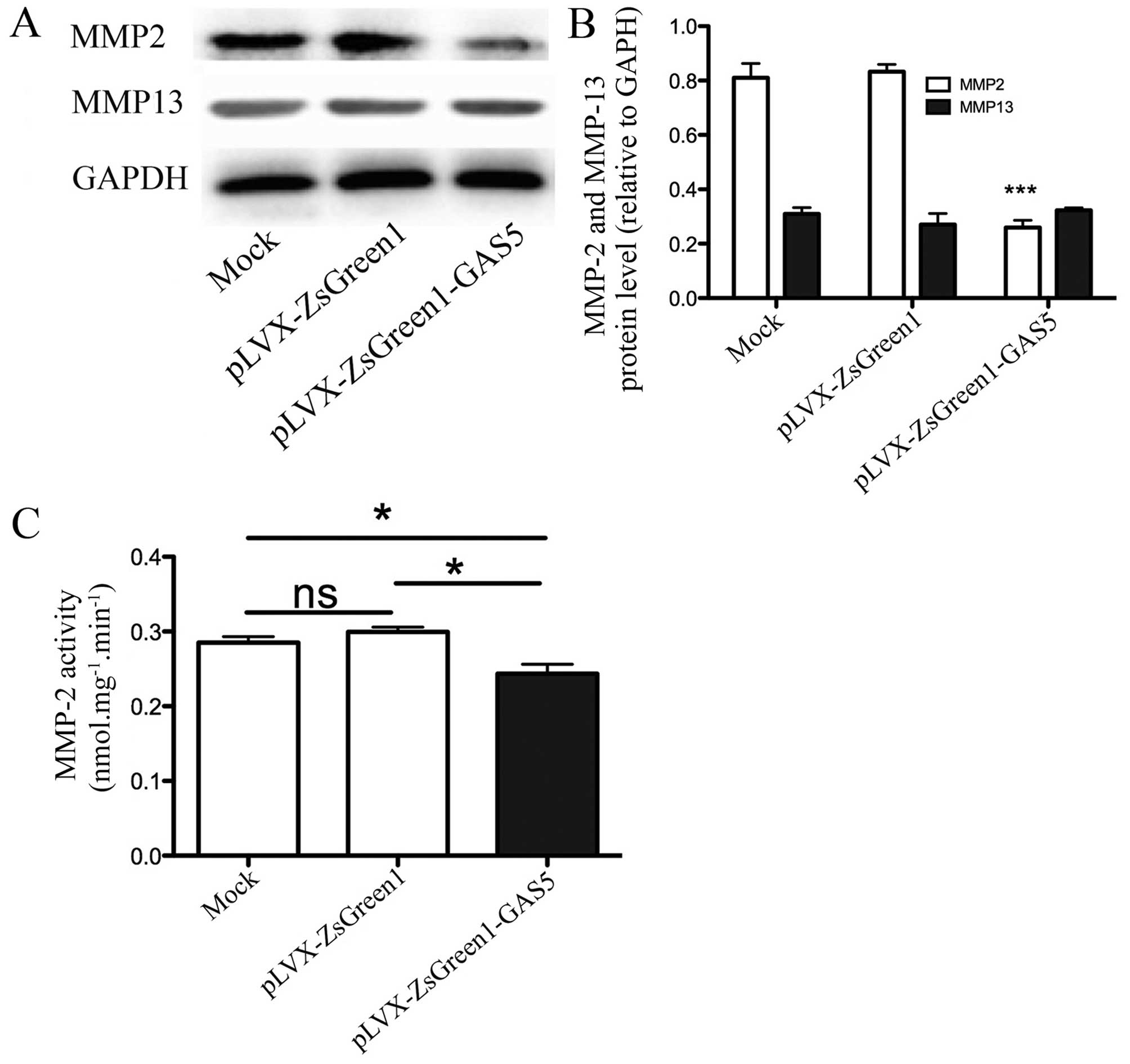
MMP2 expression levels and activity are
reduced in GAS5 overexpressing cells
We examined MMP2 and MMP13 expression levels using
western blot analysis. Overexpressing GAS5 significantly
downregulated MMP2 expression by 67.9% (P<0.001) (Fig. 6A and B). Given the importance of
MMP activity, we also examined how it was affected by GAS5
overexpression. A decrease in MMP2 activity was detected after
overexpressing GAS5 (15.8%, P=0.013) (Fig. 6C), consistent with its reduced
protein expression. However, there was no significant difference
between SK-Mel-110 and SK-Mel-110-GAS5 cells.
Discussion
The present study aimed to examine the role of
lncRNA GAS5 in the regulation of migration and invasion of melanoma
cells and to discover the potential underlying mechanisms in this
regulation.
We constructed stably expressing GAS5 melanoma cells
with which we explored the underlying function of GAS5 in the
process of melanoma metastasis. We not only found that GAS5
expression was significantly downregulated in SK-Mel-110 cells
compared to HaCaT cells, but we also showed that the overexpression
of GAS5 reduced the migration and invasiveness of SK-Mel-110 cells.
Further study revealed that overexpressed GAS5 reduced cell
invasion, at least partially, by inhibiting MMP2 function. To the
best of our knowledge, this is the first study of the relationship
between GAS5 and melanoma metastasis.
It has been reported that lncRNAs play important
roles in carcinogenesis and the aggressive progression of a
spectrum of human cancers (40,41).
Some lncRNAs have been implicated in melanoma, such as antisense
noncoding RNA in the INK4B locus (ANRIL), BRAF activated non-coding
RNA (BANCR), metastasis associated lung adenocarcinoma transcript 1
(MALAT1) and SPRYT-4 intronic transcripts (SPRY4-IT1) (39,42–45).
Specifically, chromosomal translocation involving the 1q25 locus,
which contains GAS5, has been identified in melanoma (42). Of note, lncRNA GAS5 has gained
increasing attention in cancer research because of its ubiquitous
down-regulation in many cancers (5). However, little is known about its
expression in melanoma. Here we show that GAS5 is down-regulated in
SK-Mel-110 melanoma cells, identified through qRT-PCR and FISH.
It has been demonstrated that lower GAS5 expression
is correlated with lymph node metastasis in cervical cancer and
hepatocellular carcinoma (15,21).
It is thought that low expression of GAS5 is associated with a
higher possibility of lymph node metastasis and distant metastasis
in non-small cell lung cancer and gastric cancer, although the
differences are not significant (P=0.652 and P=0.056, respectively)
(17,19). These findings further suggest that
lncRNA GAS5 may function as a tumor suppressor and may represent a
potential biomarker and new therapeutic target for some cancers.
Our results demonstrate that lncRNA GAS5 is downregulated in
SK-Mel-110 cells, more so than in HaCaT cells, and that
overexpression of GAS5 inhibits the migration and invasion of these
cells.
Migration and invasion are two crucial steps in the
process of cancer cell metastasis, which has a high likelihood of
leading to a poor prognosis. The invasion of malignant tumors
requires the proteolytic degradation of extracellular matrix (ECM)
components. Although many proteolytic enzymes and proteases have
been implicated in matrix degradation, MMPs are most closely
associated with the process of tumor invasion and metastasis
(46,47). MMP2, a 72-kDa type IV collagenase
also known as gelatinase A, is significantly increased in
metastatic melanoma (43%) compared to normal (5%) tissue. Compared
to patients with negative-to-moderate MMP2 expression, patients
with high MMP2 levels have significantly lower survival rates, and
this information could predict patient survival, independent of
tumor thickness and ulceration (48). Other studies have shown that higher
MMP13 expression, which belongs to the collagenase group, is
notably associated with metastasis and poorer survival of melanoma
patients (26–28).
Previous studies have provided limited insight
regarding the mechanisms by which lncRNA GAS5 inhibits melanoma
migration and invasion. We hypothesized that overexpressed GAS5
reduces the invasion of SK-Mel-110 cells by targeting MMP2 and
MMP13. In parallel with the reduced cell invasion, a decrease in
MMP2 protein expression levels was also observed in these cells,
consistent with the predominant role of the 72-kDa gelatinase. MMP2
is an important type IV basal membrane collagenase in human
melanoma (24,49). Moreover, both the expression level
of MMP2 and its activity, rather than that of MMP13, were
negatively associated with the expression of GAS5, which indicates
that the lncRNA GAS5 may regulate invasion, at least partially, by
modulating MMP2 protein expression levels and activity in
SK-Mel-110 cells. MMP2 and MMP13 differ from each other in terms of
their amino acid sequences, peptide domain structure and substrate
specificity, likely explaining their diverse expression profile in
SK-Mel-110 cells (50). Exploring
the potential pathway (s) by which GAS5 regulates MMP2 expression
and its activity will facilitate the understanding to lncRNA and
possibly benefit the prevention and treatment of melanoma. It also
provides signposts for identification of targets and predictive
biomarkers, as well as potentially useful guidance for decisions
about therapy. Therefore, aiming to detect whether there is any
direct interaction between GAS5 and MMP2 (both pro- and
active-MMP2), we carried out the RNA pull-down assay. Indeed, there
is a protein band close to the 63 kDa marker (active-MMP2 molecule
weight is 64 kDa) yet the mass spectrometry results revealed that
this molecule is not MMP2 (data not shown). Given that there is
marked accumulation of nucleus GAS5, we suspect that GAS5 may be
involved in the transcription of MMP2, our further studies will
focus on this potential hypothesis.
Cellular localization plays an important role in
understanding the functional roles of lncRNAs, as many lncRNAs
regulate nuclear events and thus must localize to the nucleus.
Zhang et al discovered that a novel RNA motif, a pentamer
sequence of AGCCC with special sequence restrictions at positions
-8 and -3, mediated the nuclear localization of lncRNA BORG,
demonstrating that diverse cellular localization influences cell
function and biological processes (51). A previous study showed that GAS5 is
predominantly located in the cytoplasm of breast cancer cells
(52). Similarly, the present
study also found that GAS5 locates in the cytoplasm in SK-Mel-110
cells. However, overexpressing GAS5 leads to a noticeable
accumulation of GAS5 both in the cytoplasm and nucleus, especially
in the nucleus. Considering the previous study that GAS5 can
compromise the transcription efficiency of cIAP2 by acting as a
‘GRE mimic’ in the nucleus, an explanation for its location in the
nucleus is that GAS5 exerts its effect on MMP2 possibly by
inversely regulating the transcription of MMP2 (52). We hypothesize that accumulation of
GAS5 in the nucleus may be one of crucial processes of exerting its
function. In additon, Renganathan et al revealed that GAS5
is slightly more abundant in the nucleus than that in the cytoplasm
in the absence of HhAntag and prevalence in the nucleus was
examined after treatment with HhAntag, indicating that
HhAntag-induced growth arrest involves accumulation of nuclear GAS5
(18). This study reinforced our
hypothesized model. Another potential explanation is that the
central dogma of molecular biology holds that ‘information’ flows
from the genes to the structure of the proteins through the formula
DNA→RNA→Protein and exoporting different RNAs from the nucleus to
the cytoplasm need various pathways. Large RNAs (rRNA, mRNA, lncRNA
and so on) assemble into complicated ribonucleoprotein (RNP)
particles and recruit the corresponding exporters via
class-specific adaptor proteins (53). Overexpressing GAS5 leads to
extremely increased GAS5, resulting in a temporary shortage of the
corresponding adaptors, thereby giving rise to the accumulation of
nuclear GAS5.
In conclusion, the present study established cell
models that overexpressed GAS5 to examine its effects on melanoma
cell invasion and migration, as well as other potential functions.
Our data solidified an important inhibitive role for GAS5 in the
migration and invasion of SK-Mel-110 melanoma cells, which possibly
involves MMP2. Our results show that lenti-virus-mediated
overexpression of lncRNA GAS5 decreases the invasive ability of
SK-Mel-110 cells by inhibiting MMP2 expression and activity, but
not that of MMP13, possibly at the level of transcription. Our
findings provide a new avenue for exploring how lncRNA GAS5
functions in solid tumors. Moreover, these data suggest new drug
targets and diagnostic biomarkers for malignant melanoma, and may
better inform on the choice of therapies for melanoma patients.
Acknowledgements
This study was supported by the National Science
Foundation of China (nos. 81160246 and 81160421) and the Science
and Technology Fund of Yunnan Province (no. 2013FB102). We also
appreciate American Journal Experts (AJE) for their language
editing service.
Abbreviations:
|
GAS5
|
growth arrest specific transcript
5
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lo JA and Fisher DE: The melanoma
revolution: From UV carcinogenesis to a new era in therapeutics.
Science. 346:945–949. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guil S and Esteller M: RNA-RNA
interactions in gene regulation: The coding and noncoding players.
Trends Biochem Sci. 40:248–256. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bonasio R and Shiekhattar R: Regulation of
transcription by long noncoding RNAs. Annu Rev Genet. 48:433–455.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pickard MR and Williams GT: Molecular and
cellular mechanisms of action of tumour suppressor GAS5 LncRNA.
Genes (Basel). 6:484–499. 2015.
|
|
6
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mercer TR and Mattick JS: Structure and
function of long noncoding RNAs in epigenetic regulation. Nat
Struct Mol Biol. 20:300–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Smith CM and Steitz JA: Classification of
gas5 as a multi-small-nucleolar-RNA (snoRNA) host gene and a member
of the 5′-terminal oligopyrimidine gene family reveals common
features of snoRNA host genes. Mol Cell Biol. 18:6897–6909. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schneider C, King RM and Philipson L:
Genes specifically expressed at growth arrest of mammalian cells.
Cell. 54:787–793. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Z, Zhu Z, Watabe K, Zhang X, Bai C,
Xu M, Wu F and Mo YY: Negative regulation of lncRNA GAS5 by miR-21.
Cell Death Differ. 20:1558–1568. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Romanuik TL, Wang G, Morozova O, Delaney
A, Marra MA and Sadar MD: LNCaP Atlas: Gene expression associated
with in vivo progression to castration-recurrent prostate cancer.
BMC Med Genomics. 3:432010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang XQ, Sun S, Lam KF, Kiang KM, Pu JK,
Ho AS, Lui WM, Fung CF, Wong TS and Leung GK: A long non-coding RNA
signature in glioblastoma multiforme predicts survival. Neurobiol
Dis. 58:123–131. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu Z, Wang W, Jiang J, Bao E, Xu D, Zeng
Y, Tao L and Qiu J: Downregulation of GAS5 promotes bladder cancer
cell proliferation, partly by regulating CDK6. PLoS One.
8:e739912013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tu ZQ, Li RJ, Mei JZ and Li XH:
Down-regulation of long non-coding RNA GAS5 is associated with the
prognosis of hepatocellular carcinoma. Int J Clin Exp Pathol.
7:4303–4309. 2014.PubMed/NCBI
|
|
16
|
Lu X, Fang Y, Wang Z, Xie J, Zhan Q, Deng
X, Chen H, Jin J, Peng C, Li H, et al: Downregulation of gas5
increases pancreatic cancer cell proliferation by regulating CDK6.
Cell Tissue Res. 354:891–896. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shi X, Sun M, Liu H, Yao Y, Kong R, Chen F
and Song Y: A critical role for the long non-coding RNA GAS5 in
proliferation and apoptosis in non-small-cell lung cancer. Mol
Carcinog. 54(Suppl 1): E1–E12. 2015. View
Article : Google Scholar
|
|
18
|
Renganathan A, Kresoja-Rakic J, Echeverry
N, Ziltener G, Vrugt B, Opitz I, Stahel RA and Felley-Bosco E: GAS5
long non-coding RNA in malignant pleural mesothelioma. Mol Cancer.
13:1192014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun M, Jin FY, Xia R, Kong R, Li JH, Xu
TP, Liu YW, Zhang EB, Liu XH and De W: Decreased expression of long
noncoding RNA GAS5 indicates a poor prognosis and promotes cell
proliferation in gastric cancer. BMC Cancer. 14:3192014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yin D, He X, Zhang E, Kong R, De W and
Zhang Z: Long noncoding RNA GAS5 affects cell proliferation and
predicts a poor prognosis in patients with colorectal cancer. Med
Oncol. 31:2532014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cao S, Liu W, Li F, Zhao W and Qin C:
Decreased expression of lncRNA GAS5 predicts a poor prognosis in
cervical cancer. Int J Clin Exp Pathol. 7:6776–6783.
2014.PubMed/NCBI
|
|
22
|
Jacob A and Prekeris R: The regulation of
MMP targeting to invadopodia during cancer metastasis. Front Cell
Dev Biol. 3:42015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shi H, Liu L, Liu L, Geng J, Zhou Y and
Chen L: β-Elemene inhibits the metastasis of B16F10 melanoma cells
by down-regulation of the expression of uPA, uPAR, MMP-2, and
MMP-9. Melanoma Res. 24:99–107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Toschi E, Rota R, Antonini A, Melillo G
and Capogrossi MC: Wild-type p53 gene transfer inhibits invasion
and reduces matrix metalloproteinase-2 levels in p53-mutated human
melanoma cells. J Invest Dermatol. 114:1188–1194. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Airola K, Karonen T, Vaalamo M, Lehti K,
Lohi J, Kariniemi AL, Keski-Oja J and Saarialho-Kere UK: Expression
of collage-nases-1 and -3 and their inhibitors TIMP-1 and -3
correlates with the level of invasion in malignant melanomas. Br J
Cancer. 80:733–743. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zigrino P, Kuhn I, Bäuerle T, Zamek J, Fox
JW, Neumann S, Licht A, Schorpp-Kistner M, Angel P and Mauch C:
Stromal expression of MMP-13 is required for melanoma invasion and
metastasis. J Invest Dermatol. 129:2686–2693. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Corte MD, Gonzalez LO, Corte MG, Quintela
I, Pidal I, Bongera M and Vizoso F: Collagenase-3 (MMP-13)
expression in cutaneous malignant melanoma. Int J Biol Markers.
20:242–248. 2005.
|
|
28
|
Zhao X, Sun B, Li Y, Liu Y, Zhang D, Wang
X, Gu Q, Zhao J, Dong X, Liu Z, et al: Dual effects of
collagenase-3 on melanoma: Metastasis promotion and disruption of
vasculogenic mimicry. Oncotarget. 6:8890–8899. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Houghton AN, Real FX, Davis LJ,
Cordon-Cardo C and Old LJ: Phenotypic heterogeneity of melanoma.
Relation to the differentiation program of melanoma cells. J Exp
Med. 165:812–829. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Albino AP, Juan G, Traganos F, Reinhart L,
Connolly J, Rose DP and Darzynkiewicz Z: Cell cycle arrest and
apoptosis of melanoma cells by docosahexaenoic acid: Association
with decreased pRb phosphorylation. Cancer Res. 60:4139–4145.
2000.PubMed/NCBI
|
|
31
|
Corazzari M, Rapino F, Ciccosanti F,
Giglio P, Antonioli M, Conti B, Fimia GM, Lovat PE and Piacentini
M: Oncogenic BRAF induces chronic ER stress condition resulting in
increased basal autophagy and apoptotic resistance of cutaneous
melanoma. Cell Death Differ. 22:946–958. 2015. View Article : Google Scholar
|
|
32
|
Tabolacci C, Cordella M, Turcano L, Rossi
S, Lentini A, Mariotti S, Nisini R, Sette G, Eramo A, Piredda L, et
al: Aloe-emodin exerts a potent anticancer and immunomodulatory
activity on BRAF-mutated human melanoma cells. Eur J Pharmacol.
762:283–292. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li J, Cheng Y, Tai D, Martinka M, Welch DR
and Li G: Prognostic significance of BRMS1 expression in human
melanoma and its role in tumor angiogenesis. Oncogene. 30:896–906.
2011. View Article : Google Scholar :
|
|
34
|
Hofacker IL: RNA secondary structure
analysis using the Vienna RNA package. Curr Prot Bioinform. Jun
1–2009. View Article : Google Scholar
|
|
35
|
Hofacker IL: Vienna RNA secondary
structure server. Nucleic Acids Res. 31:3429–3431. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Du Y, Zhu H, Li D, Wang L, Zhang L, Luo Y,
Pan D and Huang M: Lentiviral-mediated overexpression of Akt1
reduces anoxia-reoxygenation injury in cardiomyocytes. Cell Biol
Int. 38:488–496. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li Z, Wang H, Huang S, Zhou L, Wang L, Du
C and Wang C: Establishment of stable MRP1 knockdown by
lentivirus-delivered shRNA in the mouse testis Sertoli TM4 cell
line. Toxicol Mech Methods. 25:81–90. 2015. View Article : Google Scholar
|
|
38
|
Li L, Li B, Zhang H, Bai S, Wang Y, Zhao B
and Jonas JB: Lentiviral vector-mediated PAX6 overexpression
promotes growth and inhibits apoptosis of human retinoblastoma
cells. Invest Ophthalmol Vis Sci. 52:8393–8400. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Khaitan D, Dinger ME, Mazar J, Crawford J,
Smith MA, Mattick JS and Perera RJ: The melanoma-upregulated long
noncoding RNA SPRY4-IT1 modulates apoptosis and invasion. Cancer
Res. 71:3852–3862. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gibb EA, Brown CJ and Lam WL: The
functional role of long non-coding RNA in human carcinomas. Mol
Cancer. 10:38–55. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Spizzo R, Almeida MI, Colombatti A and
Calin GA: Long non-coding RNAs and cancer: A new frontier of
translational research? Oncogene. 31:4577–4587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Smedley D, Sidhar S, Birdsall S, Bennett
D, Herlyn M, Cooper C and Shipley J: Characterization of chromosome
1 abnormalities in malignant melanomas. Genes Chromosomes Cancer.
28:121–125. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Flockhart RJ, Webster DE, Qu K,
Mascarenhas N, Kovalski J, Kretz M and Khavari PA: BRAFV600E
remodels the melanocyte transcriptome and induces BANCR to regulate
melanoma cell migration. Genome Res. 22:1006–1014. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tian Y, Zhang X, Hao Y, Fang Z and He Y:
Potential roles of abnormally expressed long noncoding RNA UCA1 and
Malat-1 in metastasis of melanoma. Melanoma Res. 24:335–341. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wapinski O and Chang HY: Long noncoding
RNAs and human disease. Trends Cell Biol. 21:354–361. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bonnans C, Chou J and Werb Z: Remodelling
the extracellular matrix in development and disease. Nat Rev Mol
Cell Biol. 15:786–801. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shay G, Lynch CC and Fingleton B: Moving
targets: Emerging roles for MMPs in cancer progression and
metastasis. Matrix Biol. 44–46:200–206. 2015. View Article : Google Scholar
|
|
48
|
Rotte A, Martinka M and Li G: MMP2
expression is a prognostic marker for primary melanoma patients.
Cell Oncol (Dordr). 35:207–216. 2012. View Article : Google Scholar
|
|
49
|
Ray JM and Stetler-Stevenson WG:
Gelatinase A activity directly modulates melanoma cell adhesion and
spreading. EMBO J. 14:908–917. 1995.PubMed/NCBI
|
|
50
|
Brown GT and Murray GI: Current
mechanistic insights into the roles of matrix metalloproteinases in
tumour invasion and metastasis. J Pathol. 237:273–281. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhang B, Gunawardane L, Niazi F, Jahanbani
F, Chen X and Valadkhan S: A novel RNA motif mediates the strict
nuclear localization of a long noncoding RNA. Mol Cell Biol.
34:2318–2329. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Mourtada-Maarabouni M, Pickard MR, Hedge
VL, Farzaneh F and Williams GT: GAS5, a non-protein-coding RNA,
controls apoptosis and is downregulated in breast cancer. Oncogene.
28:195–208. 2009. View Article : Google Scholar
|
|
53
|
Köhler A and Hurt E: Exporting RNA from
the nucleus to the cytoplasm. Nat Rev Mol Cell Biol. 8:761–773.
2007. View Article : Google Scholar : PubMed/NCBI
|