Introduction
Esophageal carcinoma is one of alimentary canal
malignancies with high incidence of approximately 0.3104 million
new malignancies worldwide each year. Despite extensive application
in its diagnosis and treatment, recurrence of malignancy remains
the main cause of high mortality after treatment for esophageal
carcinoma patients. Current treatments, including surgical
intervention, radiotherapy and chemotherapy, are moderately
effective in the early-stage cases, but are less effective in more
advanced cases (1). For example,
patients with esophageal carcinoma may benefit from radiotherapy
(RT) to a certain degree, however, patient prognosis remains
unsatisfactory and unpredictable due to profound radioresistance.
Thus, identification of key molecules or pathways specifically
expressed in esophageal carcinoma that are essential for the growth
of cancer cells may provide novel therapeutic targets and
ultimately lead to improved survival. Research over the past years
clearly implicates multiple genetic alterations in the development
and progression of esophageal carcinoma (2), including those that have important
functions in cell growth, metastasis, DNA damage and repair
(3).
Ring finger protein (RNF2), a member of the polycomb
group of transcriptional repressors, has been detected in a variety
of human carcinoma specimens in various stages. In particular, RNF2
is overexpressed in a number of malignancies (4,5).
GeneChip analysis also showed that RNF2 can be used to predict
cancer metastasis (6), promote
cancer cell proliferation and invasion, resist apoptosis and
enhance transfer capabilities (7).
Moreover, as an active heterodimer E3 ligase, RNF2 may be
associated with the ubiquitination and phosphorylation of H2AX,
which is thought to be a critical sensor that can initiate DNA
damage response (DDR) (8,9). It has been also shown that H2AX was
associated with radio-sensitization of esophageal cancer cells
(10). However, how the function
of RNF2 in radiosensitivity is regulated remains a fundamental
question that needs to be answered to elucidate the essential
mechanisms controlling DDR.
To identify new and crucial factors regulating the
function of RNF2 in DDR, we conducted a proteomic analysis to
systematically identify RNF2 interacting proteins. To the best of
our knowledge, the mechanisms by which RNF2 promoted the growth of
esophageal carcinoma cells after radiotherapy has not been reported
thus far. Given the important role of RNF2 in ionizing
radiation-induced DDR, we hypothesize that silencing of RNF2 may
bring in DNA damage pathway defects and thus increase the
radiosensitivity. In the present study, we attempted verification
of this hypothesis through different cells lines in vitro
and explored the mechanisms through which RNF2 induces oncogenesis
and tumor progression.
Materials and methods
Tissue specimens and immunohistochemical
analysis
Sixty-four esophageal carcinoma tissues and five
normal esophageal tissues were obtained from the Department of
Thoracic Surgery, Hebei Hospital of the Fourth affiliated Medical
University (Hebei, China), from January 2008 to December 2009.
Patient consent was obtained for the collection of specimens, and
all study protocols were approved by the Ethics Committee for
Clinical Research of the Fourth affiliated Medical University.
Immunohistochemistry protocols were performed as previously
described (11). In brief, slides
were incubated with an anti-RNF2 monoclonal antibody (1:100,
EPR12245; Upstate, Lake Placid, NY, USA) followed by incubation
with a horse-radish peroxidase-conjugated antimouse secondary
antibody (1:200; Dako, Glostrup, Denmark). Antibody binding was
visualized using 3,3′-diaminobenzidine and counterstained with
hematoxylin. Negative control sections were incubated with PBS
instead of the primary antibody. Immunostained results were
independently evaluated by two pathologists who were blinded to the
clinical and histopathological features. Ring finger protein 2
(RNF2) cytoplasm accumulation was quantified as a percentage of the
total number of cytoplasm detected in at least 4–5 random high
power fields (x400) in each section. Cases with >10% of cells
staining for RNF2 were scored as positive samples (12).
Cell lines and cell culture
Five human esophageal cancer cell lines, TE13, TE1,
KYSE30, EC9706 and ECA109, and the normal esophageal cell line
HEEC, were maintained at 37°C and 5% CO2 in RPMI-1640
medium (Gibco, Gaithersburg, MD, USA) supplemented with 10% fetal
bovine serum (FBS; Invitrogen, Carlsbad, CA, USA), penicillin (100
U/ml), and streptomycin (0.1 mg/ml).
X-ray irradiation
Irradiation was performed at room temperature with a
6MV Siemens linear accelerator (Siemens, Concord, CA, USA) at a
dose rate of 2 Gy/min. After irradiation, cells were recovered in
an incubator for the indicated time until harvesting.
shRNA transfection
For the shRNA analyses, human RNF2 small interfering
RNA (shRNA) with the nucleotide sequence
5′-GGUAACGCCACUGUUGAUCACUUAU-3′ (sense) and
5′-AUAAGUGAUCAACAGUGGCGUUACC-3′ (antisense), corresponding to part
of the RNF2 mRNA, and the negative control scrambled shRNA
(NC-shRNA: sense, 5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACAGGUUCGGAGAATT-3′) were designed and purchased from
Invitrogen. All of the shRNA sequences were subjected to basic
local alignment search tool (BLAST) to confirm the absence of
homology to any additional known coding sequences in the human
genome. Cells were transfected using Lipofectamine RNAiMAX
(Invitrogen) according to the manufacturer's protocol. Briefly, one
day prior the transfection, ECA109 and TE13 cells
(5×105/well) were cultured in 6-well tissue culture
plates until they reached 50% confluence, then the cells were
transiently transfected with either RNF2-shRNA (Shanghai Genechem
Co., Ltd., Shanghai, China) or NC-shRNA (100 nM). Cells were
collected 24 h after transduction with shRNA and were selected in
puromycin for stable clones. Subsequently, the cells were used for
irradiation as indicated.
Quantitative real-time reverse
transcription-polymerase chain reaction (qRT-PCR)
Total RNAs were extracted by using TRIzol reagent.
Real-time PCR was then performed using Platinum SYBR-Green qPCR
SuperMix-UDG (Invitrogen) according to the manufacturer's protocol.
Briefly, after the reverse transcription reaction at 42°C for 60
min and 70°C for 5 min, cDNAs was synthesized using the ReverAid
First Strand cDNA Synthesis kit (Thermo Fisher Scientific, Waltham,
MA USA), then initially denatured at 94°C for 30 sec, and followed
by 40 cycles of repeated procedure as follows: denatured at 94°C
for 5 sec, annealed at 56°C for 15 sec and extended at 72°C for 10
sec. As a control, the levels of glyceraldehyde phosphate
dehydrogenase (GAPDH) expression were also analyzed. Incorporation
of the SYBR-Green dye into PCR products was monitored in real-time
with LightCycler real-time PCR detection system (Roche Applied
Science, Indianapolis, IN, USA), thereby allowing determination of
the threshold cycle (Ct) at which exponential amplification of
products begins. Independent experiments were repeated three times
for each sample and the relative expression levels of genes were
analyzed by using 2−ΔΔCt method (13).
Western blot analysis
The cellular total protein was solubilized in RIPA
lysis buffer (1% Triton X-100, 150 mM NaCl, 10 mM Tris-HCl, pH 7.4,
1 mM EDTA, 1 Mm EGTA, pH 8.0, 0.2 mM Na3VO4,
0.2 mM phenylmethylsulfonylfluride and 0.5% NP-40). The protein
amount was evaluated by using BCA assays (Pierce, Rockford, IL,
USA) and separated on 10% SDS-PAGE gel, and electrotransferred to a
PVDF membrane (Pierce). The membrane was incubated overnight at 4°C
with the indicated antibodies. The specific antibodies were rabbit
RNF2 antibody (1:10,000; Abcam, Cambridge, USA), anti-rH2AX
(1:10,000; Abcam), anti-H2AK119ub (1:10,00; Millipore, Billerica,
MA, USA), rabbit Bcl-2 (1:1,000; Abcam), and anti-Bax (1:500;
Abcam), anti-H2AX (1:1,000; Abcam), anti-p16 (1:50,00; Abcam),
anti-cyclin D2 (1:500; Abcam), anti-CDK4 (1:1,000; Abcam) and
rabbit β-actin (1:10,000; Bioworld Technology, Inc., Louis Park,
MN, USA). After washing for 3×5min with TBS-T, the membrane was
incubated with secondary anti-rabbit IgG for 1 h at room
temperature away from light. The membrane was scanned for the
relative value of protein expression in gray scale by Image-Pro
plus software 6.0 (Media Cybernetics, Sliver Spring, MD, USA). The
levels of the proteins were calculated as the ratio of the
intensity of protein to that of β-actin. Experiments were carried
out in triplicate wells each time and repeated three times.
Cell proliferation assay
The cells (2×103/well) were cultured in
96-well tissue culture plates until they reached 50% confluence,
then transfected with a final concentration of 100 nM. After the
cells had been transfected for 24 h, viability of the cells were
determined at 24, 48, 72 and 96 h after irradiation using the Cell
Titer 96 AQueous One Solution cell proliferation assay
(MTS) that was purchased from the Dojindo Molecular Technologies
(Gaithersburg, MD, USA). Briefly, cells were collected and
incubated in medium containing 5 mg/ml MTS reagent (Promega,
Madison, WI, USA) at 37°C for 2 h. The absorbance at 492 nm was
measured by an enzyme linked immunosorbent assay (ELISA) plate
reader. This experiment was repeated three times.
Co-immunoprecipitation (Co-IP) assay
Cell lysates were first pre-cleared with 25 ml of
Protein A-agarose (Santa Cruz Biotechnology). The supernatants were
immunoprecipitated with 2 mg of anti-rH2AX (1:100; Abcam),
anti-H2AK119ub (1:100; Millipore), anti-RNF2 (1:100; Abcam)
antibody for 2 h at 4°C, followed by incubation with protein
A-agarose overnight at 4°C. Protein A-agarose-antigen antibody
complexes were pelleted by centrifugation at 12,000 x g for 60 sec
at 4°C. The pellets were washed five times with 1 ml IPH buffer,
for 20 min each time at 4°C. Bound proteins were resolved by
SDS-PAGE, followed by western blot analysis with antibodies against
rH2AX, H2AK119ub and RNF2.
Analysis of cell cycle and apoptosis
Both cell cycle distribution and spontaneous
apoptosis events were detected by using a FACSCalibur II sorter and
CellQuest FACS system (BD Biosciences, San Jose, CA, USA). To
analyze cell cycle distribution, cells were transfected by using
shRNA for 24 h and stimulated with irradiation for 24 h before
being harvested. Cells were fixed with 70% ethanol at 4°C
overnight, washed twice with PBS, and resuspended in staining
solution (50 μg/ml propidium iodide, 1 mg/ml RNase A, 0.1% Triton
X-100 in PBS) for 30 min at 37°C in the dark. To detect the extent
of apoptosis under stress conditions, cells were transfected using
shRNA for 24 h before irradiated and stained with fluorescein
isothiocyanate (FITC)-conjugated Annexin V and 7-amino-actinomycin
D (7-AAD) using the Annexin V-FITC apoptosis detection kit (BD
Biosciences) according to the manufacturer's protocol.
Colony formation assay
Cell survival curves were generated by a standard
colony formation assay with minor modifications (14). Precooled tumor cells were
irradiated by graded single doses (0–8 Gy) in control, NC and RNF2
shRNA groups, seeded in Petri dishes, and then cultivated in
RPMI-1640 supplemented with 10% FBS. The experiments were repeated
at least twice. Two weeks later, the cells were fixed and stained
with crystal violet (0.6%). Colonies of exceeding 50 cells were
scored as survivors.
Statistical analysis
Statistical analysis was conducted with the SPSS
software package (version 13.0). All data were presented as the
mean ± standard deviation (SD), and analyzed using ANOVA with the
SPSS 13.0. For all tests, a P-value <0.05 and 0.01 was
considered to be statistically significant and indicated by
asterisks in the figures. All P-values given were the results of
two-sided tests. Data were obtained from at least three independent
experiments with a similar pattern.
Results
RNF2 expression is increased in
esophageal cancer cells and specimens
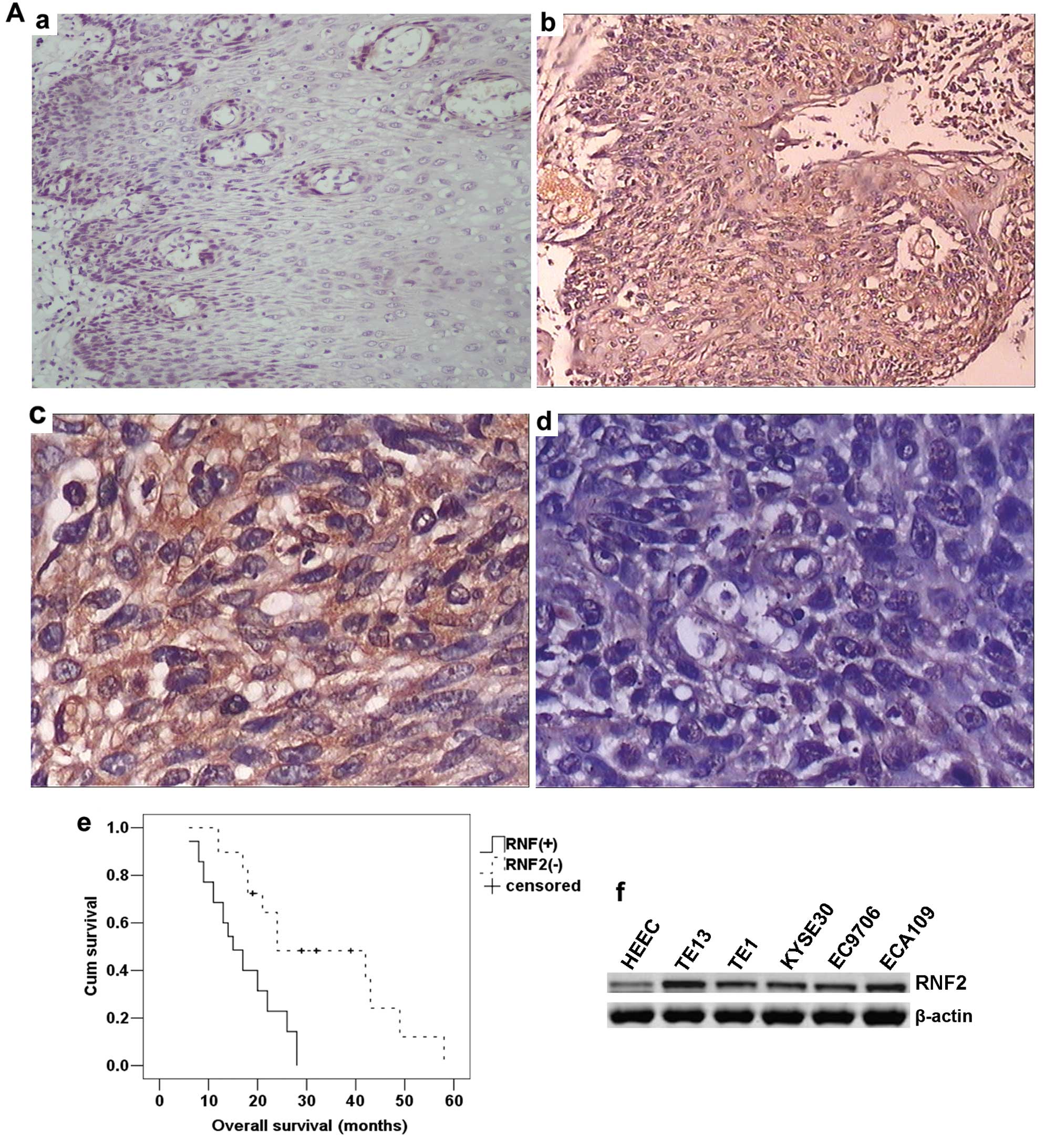
The expression of RNF2 in esophageal cancer and
normal esophageal tissues was analyzed by immunohistochemistry. In
contrast to previous reports (15), RNF2 was mainly expressed in
neoplastic epithelial cytoplasm, plasma-lemma and occasionally in
the cell nuclei (Fig. 1A-b and
-c). In addition, no staining or only weak staining was seen in
normal epithelium cells (Fig.
1A-a). For the 64 esophageal cancer patients, increased
cytoplasm accumulation of RNF2 was found in 35 (54.69%) specimens.
Furthermore, we detected RNF2 expression in five esophageal cancer
cell lines (TE13, TE1, KYSE30, EC9706 and ECA109) and the immortal
esophageal epithelial cell line HEEC. The levels of RNF2 expression
in the esophageal cell lines were higher than that for the HEEC
cell line (Fig. 1A-f).
RNF2 overexpression is associated with
the progression and adverse prognosis of esophageal cancer
To investigate the clinical role of RNF2 during
esophageal cancer development, we analyzed the correlation between
RNF2 expression and the clinicopathological features of patients
with esophageal cancer. As shown in Table I, the cytoplasm accumulation of
RNF2 in esophageal cancer was significantly associated with tumor
size, clinical stage and lymph node metastases (P<0.05),
indicating a correlation between RNF2 expression and esophageal
cancer invasion, and metastasis. However, no evident correlation
were observed between the expression of RNF2 and other clinical
features such as patient age, gender and histological grade
(P>0.05 for all comparisons). Furthermore, Kaplan-Meier survival
analysis demonstrated that patients harboring RNF2 overexpression
had a significantly shorter overall survival than patients with
lower levels of RNF2 expression (P<0.001, log-rank test;
Fig. 1A-e). Taken together, these
observations indicate that RNF2 could contribute to the evaluation
of the prognosis in patients with esophageal cancer.
 | Table IAssociation between RNF2 expression
and clinico-pathological variables of patients with esophageal
cancer. |
Table I
Association between RNF2 expression
and clinico-pathological variables of patients with esophageal
cancer.
| RNF2 expression | |
|---|
|
| |
|---|
| Variables | Positive | Negative | P-value |
|---|
| Age (years) |
| >60 | 19 | 13 | 0.451 |
| ≤60 | 16 | 16 | |
| Gender |
| Male | 18 | 14 | 0.802 |
| Female | 17 | 15 | |
| Tumor size
(cm) |
| ≤2 | 13 | 19 | 0.024 |
| >2 | 22 | 10 | |
| Histological
grade |
| Well or
moderated | 16 | 17 | 0.304 |
| Poorly | 19 | 12 | |
| TNM stage |
| I–II | 12 | 18 | 0.027 |
| III–IV | 23 | 11 | |
| Lymph node
metastasis |
| Positive | 25 | 12 | 0.015 |
| Negative | 10 | 17 | |
Specific knockdown of RNF2 expression by
shRNA inhibits the growth and improves the radiosensitivity of
esophageal cancer cells after IR in vitro and in vivo
To address the functional importance of the RNF2
gene, we first employed RNF2-specific shRNA to deplete its
expressions in ECA109 and TE13 cells, both of which were treated
with negative control (NC) shRNA or shRNA targeting the RNF2 gene
owing to their high levels of RNF2. After transfected for 24 h, the
expression of RNF2 in cells was subsequently determined by
real-time quantitative reverse transcription-polymerase chain
reaction (qRT-PCR) and western blot analysis. The qRT-PCR analysis
confirmed that the levels of glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) were unaffected by transfection of RNF2 shRNA
or NC shRNA. As shown in Fig. 1B,
qRT-PCR showed that the shRNA RNF2 effectively suppressed the
expression of RNF2 in both ECA109 and TE13 cells (P<0.01).
Furthermore, the empty vector had no effect on the expression of
RNF2 in either cell line. Similar results were observed in the
western blot analysis (Fig. 1C).
Therefore, ECA109-RNF2 shRNA and TE13-RNF2 shRNA cells were further
characterized.
To evaluate the effect of RNF2 depletion on cell
proliferation, cell viability was first examined by using MTS
assay. As shown in Fig. 2A, the
proliferation rate between ECA109 and TE13 cells was not
significantly different in three groups at each time point without
irradiation. In contrast, RNF2 depletion significantly inhibited
the growth of ECA109 and TE13 cells compared with cells in control
group and NC group after IR (P<0.05) (Fig. 2B). Moreover, the proliferation
level in each group was obviously higher at 48, 72 and 96 h after
IR than corresponding unirradiated groups. In addition, we also
further detected the radiatiosensitivity in different groups after
exposure to IR and found that there was no significant difference
between control and NC groups. However, cells in the RNF2 shRNA
group had higher sensitivity than the other groups (Fig. 2C). To confirm these findings in
vivo, xenograft tumor growth assays were performed in nude
mice. Compared with the control cells, injection of ECA109 and TE13
RNF2 shRNA cells led to markedly decreased tumor weight (P<0.05;
Fig. 2D). Collectively, these data
suggested that the knockdown of RNF2 after irradiation
significantly inhibited cells proliferation and tumorigenicity and
increased radiosensitivity.
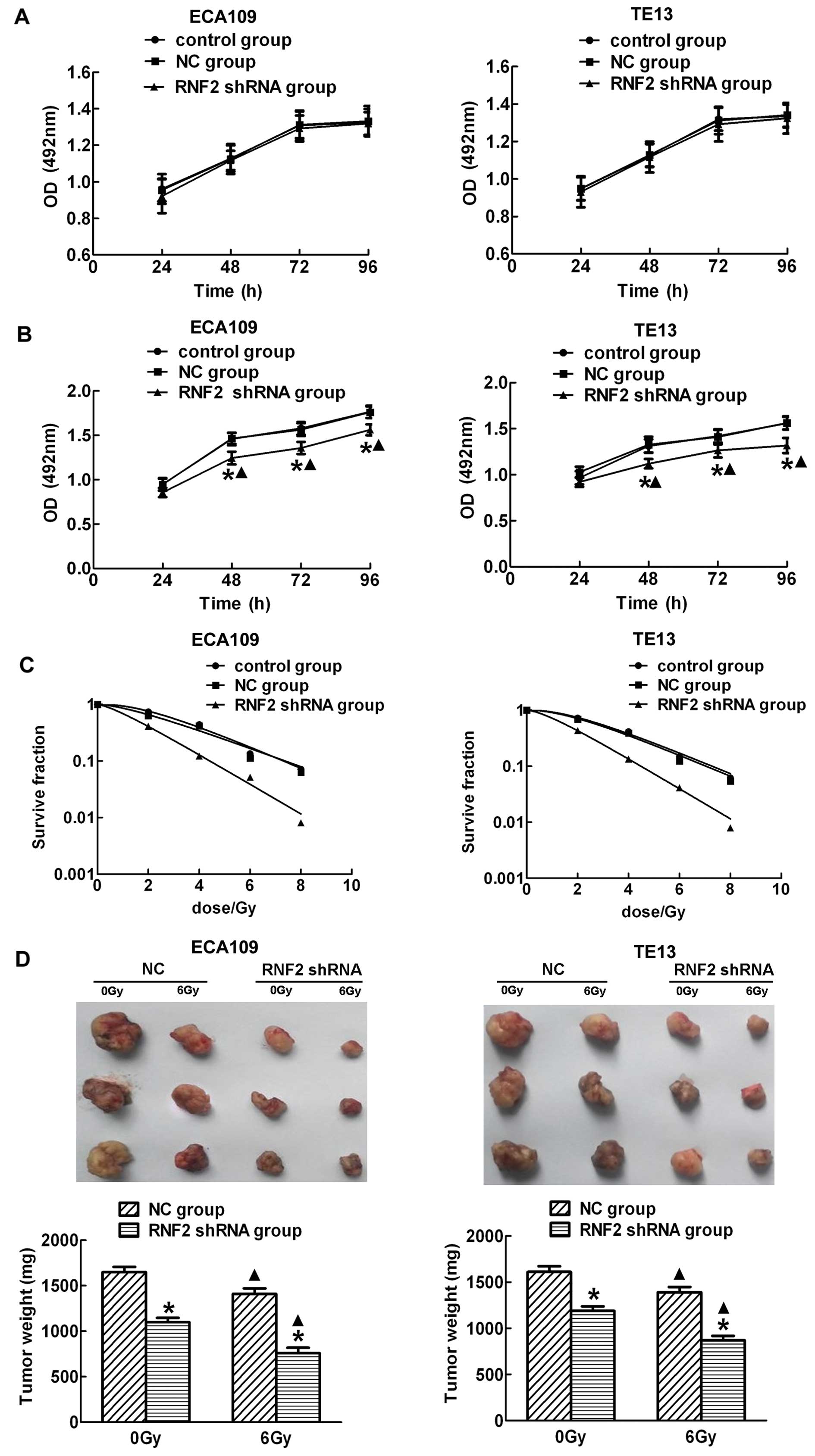 | Figure 2RNF2 depletion causes inhibition of
growth and tumorigenesis of esophageal carcinoma cells. There were
no obvious difference among control group, NC group and RNF2 shRNA
group without irradiation both in ECA109 and TE13 cells (A), while
downregulation of RNF2 expression by RNF2 shRNA significantly
inhibited the growth of ECA109 and TE13 cells after irradiation
(B). Besides, the proliferation levels in the three groups at 48,
72 and 96 h after IR were higher than before irradiation. Survival
curves of treated with IR were drawed, respectively. GraphPad Prism
5.0 was used to fit cell survival curves, as well as the radiation
biological parameters. Control and NC group cells had a broader
shoulder compared to RNF2 shRNA group cells, indicating that they
were more radioresistant than the latter (C). The nude mice were
sacrificed, the tumors were dissected, and tumor weights were
measured, the data are shown as the means ± standard error for each
group (n=5) (D). *P<0.05 compared with control group
or NC group. ▲P<0.05 compared with corresponding
unirradiated group. |
RNF2 interacts with the ubiquitination
and phosphorylation of H2AX in ECA109 and TE13 cells in a DNA
damage-induced manner
In our experiment, ionizing radiation induced the
expression of RNF2, H2AK119ub, r-H2AX in a dose-dependent manner.
Moreover, the expression of three kinds of proteins were consistent
in ECA109 and TE13 cells (Fig. 3A and
B). All were highest at 1 and 2 h, and their expression
gradually reduced after this time. By 24 h, levels were restored to
unirradiated levels. These data led us to believe that there was
some relationship between RNF2 and H2AK119ub, r-H2AX. Therefore,
the specificity of this interaction was confirmed by
co-immunoprecipitation of H2AK119ub, r-H2AX and RNF2. Notably,
there was no significant interaction without irradiation (Fig. 3C), while their interaction was
obviously enhanced in both ECA109 and TE13 cells upon IR (Fig. 3D), indicating that RNF2 may play an
important role in DNA damage repair through inducing the
ubiquitination or phosphorylation of H2AX.
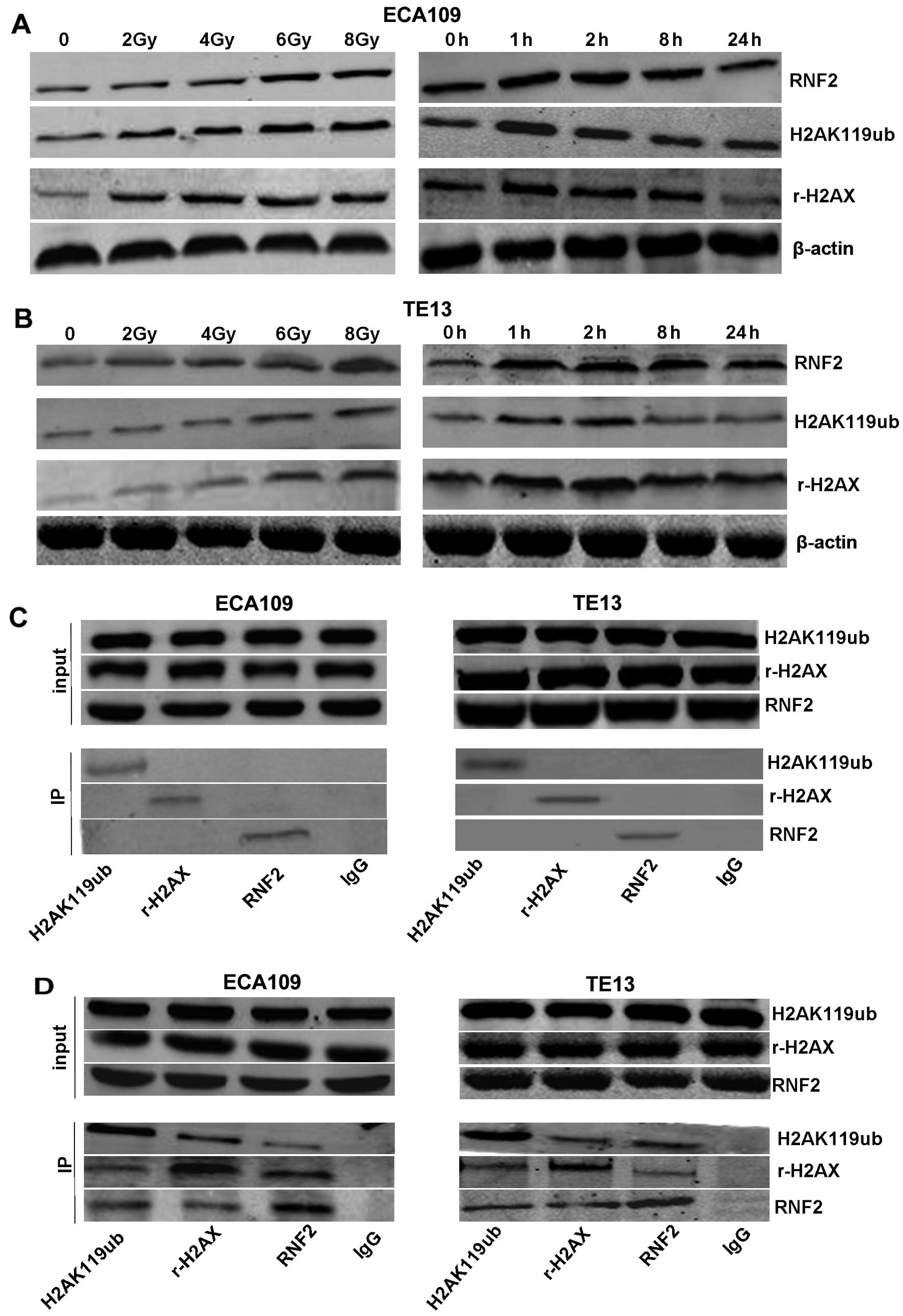 | Figure 3The interaction of RNF2 and H2AK119ub,
r-H2AX in ECA109 and TE13 esophageal cancer cells after irradiation
or not. The expression of RNF2, r-H2AX and H2AK119ub were observed
in ECA109 (A) and TE13 (B) cells by western blotting. Cells were
irradiated with 2, 4, 6 or 8 Gy, and fixed for 1 h after IR,
followed by being irradiated with 6 Gy at different times. In
addition, there was no significant interaction without irradiation
in cells (C), while the interaction between RNF2 and H2AK119ub,
r-H2AX was obviously enhanced in cells after irradiation (D),
implying that RNF2 may be related to the phosphorylation and
ubiquitination of H2AX. |
Mechanisms of radiosensitization by
silencing of RNF2
In order to further explore the mechanisms by
silencing of RNF2 in promoting radiosensitization of esophageal
cancer cells, flow cytometry was performed demonstrating that
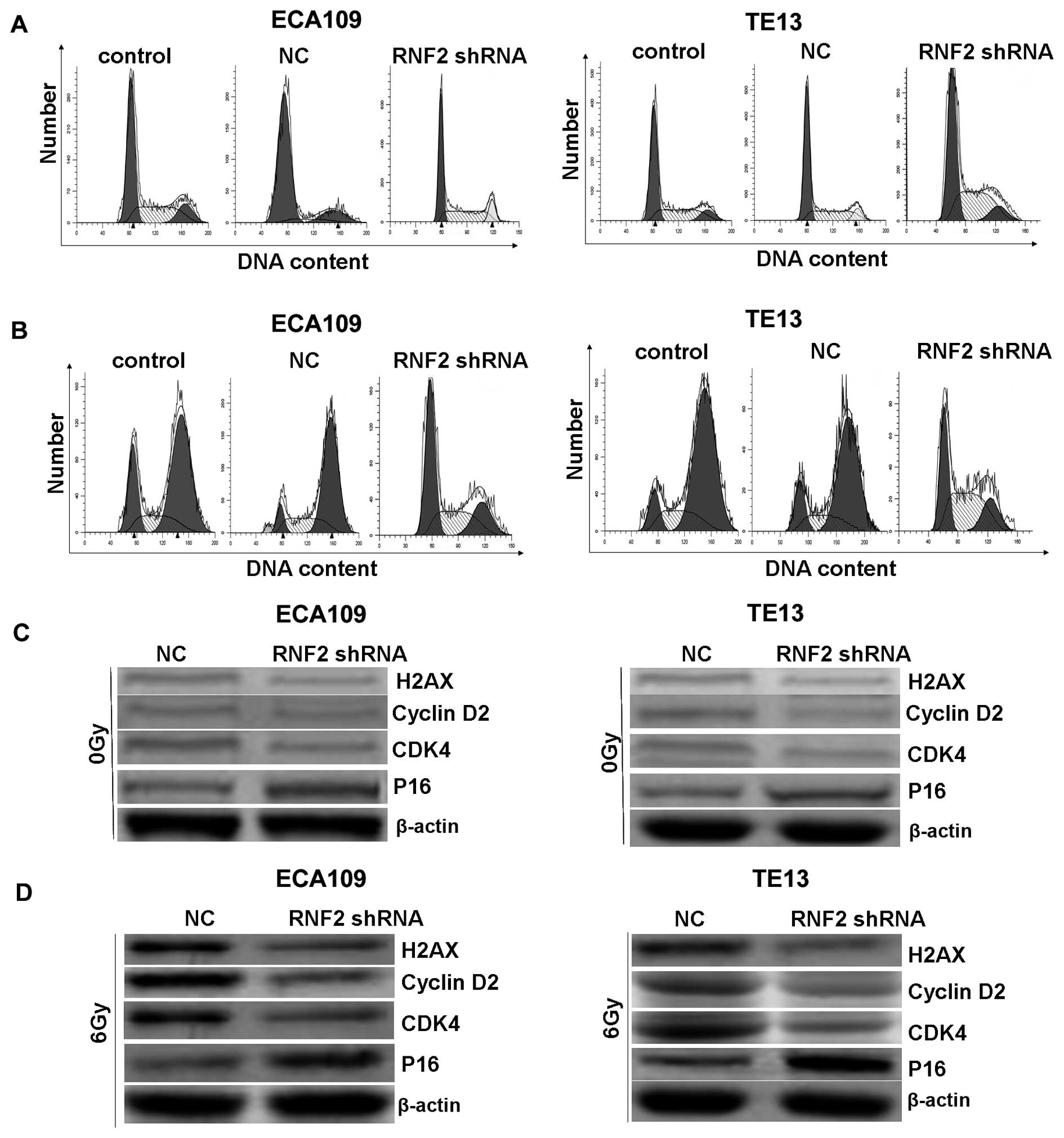
irradiation could obviously induce cell cycle arrest at G2/M phase
after irradiation for 24 h in both ECA109 and TE13 cells in
vitro (Fig. 4B). However,
silencing RNF2 by shRNA could cause G1 arrest in cells and reduce
compensatorily G2/M phase, which allowed much time to kill more
tumor cells, and induced radiosensitivity. If no irradiation, there
was no significant difference among RNF2 shRNA group, control group
and NC group (Fig. 4A). Our
results showed that RNF2 gene knockdown suppressed the entry of
cells from G1 into S phase, inhibited cell growth, and therefore
enhanced radiosensitivity. Furthermore, western blotting was
performed to explore the cell cycle regulatory role of RNF2.
Interestingly, the levels of these indexes were also detected but
were not obviously altered before IR (Fig. 4C). However, after RNF2 knockdown,
the expression of cyclin D2, cyclin dependent kinase 4 (CDK4) and
H2AX was significantly decreased, while the level of p16 was
obviously increased after IR (Fig.
4D). Collectively, these data demonstrated that RNF2 may
regulate proteins associated with cell cycle through interaction
with H2AX.
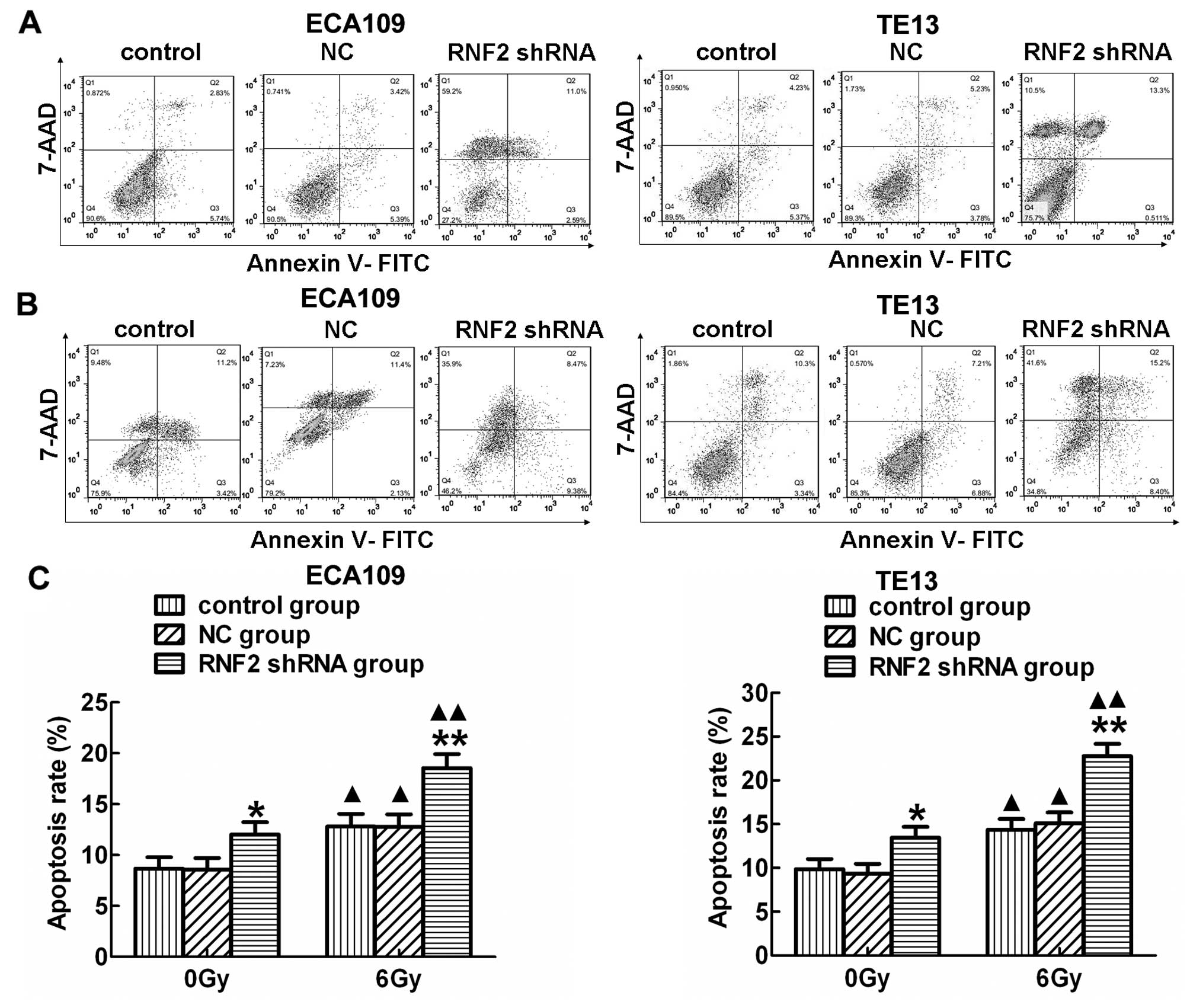
In addition, the apoptosis rate in RNF2 shRNA group
was significantly higher than the other two groups in both ECA109
and TE13 cells before (Fig. 5A)
and after IR (Fig. 5B; P<0.05).
Moreover, a strong increase in apoptosis was detected in both cell
lines after IR compared to the corresponding irradiated groups
(P<0.05). Furthermore, the expression of several cell
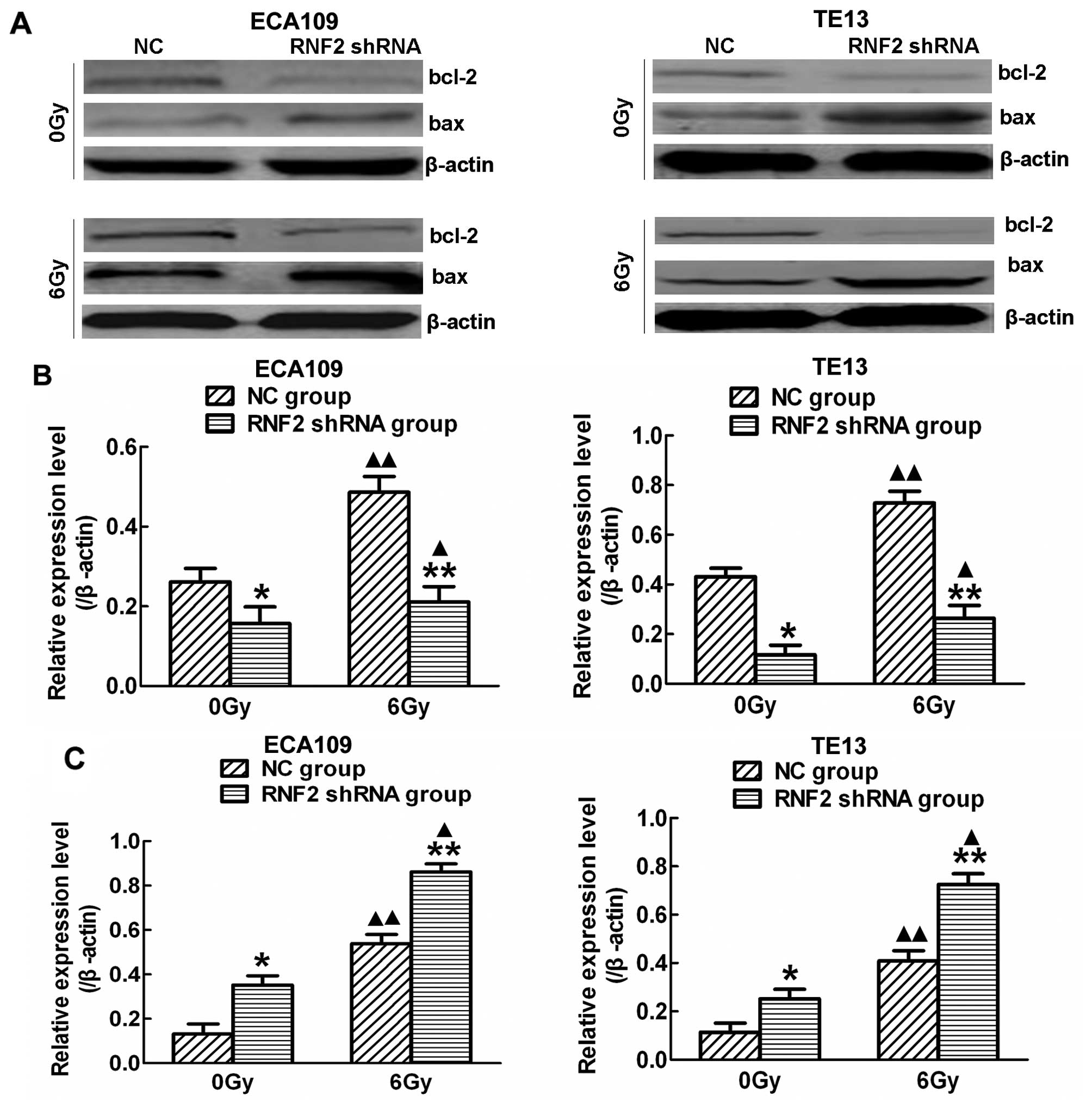
survival-related proteins was evaluated by western blot analysis
(Fig. 6A). Following RNF2
silencing, the expression of Bcl-2 was decreased (Fig. 6B), while the expression of Bax was
increased before and after IR (Fig.
6C), and the level of these indexes was higher after IR than
before IR (P<0.05). Thus, RNF2 has an ability to regulate the
cell cycle, and induce apoptosis thus promoting radiosensitivity
after IR.
Discussion
Esophageal carcinoma is one of the most frequent
malignant tumors, with high morbility and mortality worldwide. For
example, the incidence of esophageal carcinoma is as high as
26.3/100,000 individuals in America, which is higher than in Asia.
Radiotherapy is one of the most important therapeutic stragies for
esophageal carcinoma, however, the 5-year survival rate is only
from 10 to 15%, accompanied high recurrence rate from ~60 to 80%.
Although the local control rate is improved after extensive
application of new techniques in radiotherapy, local failure is
still the main cause of death. Moreover, the molecular mechanisms
behind the development and progression of esophageal carcinoma
after RT are uncertain.
RNF2, one of core components in the polycomb
repressive complex 1, may play an important role in the
immortalization of normal epithelial cells, early malignant
transformation, the maintenance of the self-renewal of stem cells,
and carcinogenesis (16–18). In addition to their role in
development, RNF2 has been reported to be overexpressed in a
variety of human cancers, such as various malignant solid tumors
(19). In the present study, we
provided additional clinicopathological evidence and a mechanistic
basis for the role of RNF2 in human esophageal carcinoma
progression. We found that RNF2 was overexpressed in both
esophageal carcinoma tissues and cell lines compared with normal
esophageal tissues and a normal esophageal epithelial cell line.
Here, we further demonstrated that aberrant cytoplasm
overexpression of RNF2 was positively correlated with tumor size
and lymph node metastases, but negatively correlated with patient
survival, underscoring the role of RNF2 in the progression of
esophageal carcinoma. Indeed, this association between RNF2
upregulation and tumor size, pathological grade, and an adverse
prognosis is not unprecedented, as similar findings have been
reported in other tumor types (20). However, there is no statistically
significant correlation between RNF2 overexpression and the age or
gender of esophageal carcinoma patients. The reason may be due to
the small number of cases examined in the present study, and
therefore additional specimens would need to be analyzed to confirm
our results.
To further confirm the significance of RNF2
overexpression in the progression of esophageal carcinoma, we
designed and constructed shRNA recombinant expression vector
targeting RNF2 to downregulate its expression in both the protein
and mRNA level in two esophageal carcinoma cell lines. We
investigated the effect of RNF2 shRNA on the proliferation and
radiosensitivity of esophageal carcinoma cells upon irradiation.
RNF2 gene knockdown in ECA109 and TE13 cells significantly
inhibited cell proliferation and improved radiosensitivity after
the cells were treated with shRNA and irradiation at different
times. Previous studies have reported that the downregulation of
RNF2 by the adenovirus-mediated delivery of RNF2 shRNA reduced the
proliferation, and invasion of cancer cells, and decreased the
radioresistance (15). Our results
were in accordance with previous studies. Notably, there was no
obvious difference in cell proliferation of different groups
without irradiation, manifesting that radiotherapy combined with
shRNA can effectively inhibit the proliferation of tumor cells and
enhance the radiosensitization.
It has been reported that RNF2 was associated with
ubiquitination and phosphorylation of H2AX (8,21).
Similar studies identified that RNF2 possesses the E3 ligase
activity for H2AX monoubiquitination and interacts with H2AX in a
DNA damage-induced manner (22,23).
In accordance with previous studies, we found RNF2 regulated
monoubiquitination and phosphorylation of H2AX in the DNA damage
response induced by ionizing radiation in a dose-dependent and
time-dependent manner. In addition, the interaction between RNF2
and H2AK119ub, r-H2AX was obviously enhanced in esophageal
carcinoma after exposure to IR, while there was no significant
interaction without irradiation, indicating a vital role of RNF2 in
DNA damage response and its potential mechanism. However, we did
not assess whether RNF2 is involved in ubiquitination or
phosphorylation of H2AX. Thus, we will evaluate this aspect in
order to better explore its related mechanism of RNF2 in DNA damage
response.
Cellular responses to DNA damage include cell cycle
arrest and apoptosis. Previous studies have reported that
downregulation of RNF2 caused G0/G1 arrest in Tera-1 cells
(7). Our data demonstrated that
inhibiting the expression of RNF2 by shRNA in ECA109 and TE13 cells
significantly induced spontaneous cell apoptosis and cell cycle
arrest at the G0/G1 phase after IR. However, given no ionizing
radiation, there was no statistically significant difference among
the control group, NC group and RNF2 shRNA group. We also detected
the expression of several known G0/G1 cell cycle-related proteins
in different groups before and after IR. As a result, obvious
decreases in the levels of cyclin D2, CDK4 and marked increases of
p16 were observed after RNF2-depletion and IR. However, their
levels had no significant alteration before IR. P16 is a member of
CDK inhibitors which mediated negative effects on cell cycle
progression through the binding of various cyclin-CDK complexes and
suppression of their viability (24). Prior studies demonstrated that
rH2AX, phosphorylation of H2AX and a marker of DNA damage response,
was associated with p16 and p53 (25,26).
DNA damage can induce the expression of p53 and p16, enhancing cell
cycle arrest at G1 phase (27). In
the present study, RNF2 depletion downregulated the level of cyclin
D2, CDK and upregulated the expression of p16, and then altered
cell cycle distribution. Interestingly, knockdown of RNF2 also
decreased the expression of H2AX. As described above, the
expression of RNF2 was related to ubiquitination or phosphorylation
of H2AX. Therefore, these results suggest that RNF2-mediated
promotion of tumor progression is related to increased cell growth
via interaction with H2AX to regulate the expression of some
cyclins and p16, and further to regulate the cell cycle.
In the present study, our data showed that apoptosis
of cells was higher in RNF2 shRNA group than control group and NC
group before and after exposure to irradiation. Moreover, our
results also detected the promotion effect of RNF2 shRNA on
apoptosis in ECA109 and TE13 cells after treated with irradiation
which was accompanied by downregulated expression of Bcl-2 and
upregulation of Bax, which are the downstream targets of the
nuclear factor-kappa B (NF-κB) and associated with cell apoptosis
after DNA damage (28). Thus,
these data demonstrated the downregulation of RNF2 shRNA enhances
IR-induced apoptosis. RNF2-mediated Bcl-2 and Bax expression may
serve as a novel pathway to contribute to esophageal carcinoma
progression. Moreover, a recent study showed that RNF2 silencing
significantly sensitized glioma cells to irradiation in
vitro through increasing apoptosis (29). Therefore, our results suggest the
mechanism of RNF2-mediated promotion of tumor progression may be
associated with Bcl-2 family proteins to enhance cell survival.
In summary, the present study detected RNF2
expression and its clinical significance in esophageal carcinoma.
RNF2 was found to be overexpressed in esophageal carcinoma cell
lines and was associated with the poor prognosis of esophageal
carcinoma patients. Our data reflect the important role of RNF2 in
the growth of esophageal carcinoma cells by shRNA silencing of RNF2
expression. In addition, downregulation of RNF2 decreased Bcl-2
expression, enhanced Bax expression and induced IR-induced G0/G1
arrest by inhibiting RNF2 expression. To the best of our knowledge,
we demonstrated the suppression of RNF2 expression by shRNA in
esophageal carcinoma for the first time after treatment with
irradiation, indicating that RNF2 may be an important gene for
targeted therapy of human esophageal carcinoma.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (no. 81372416).
References
|
1
|
Sun Y, Liu M, Yang B, Li B and Lu J: Role
of siRNA silencing of MMP-2 gene on invasion and growth of
laryngeal squamous cell carcinoma. Eur Arch Otorhinolaryngol.
265:1385–1391. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhao JX and Xie XL: Regulation of gene
expression in laryngeal carcinama by microRNAs. Int J Pathol Clin
Med. 32:222–225. 2012.
|
|
3
|
Nacerddine K, Beaudry JB, Ginjala V,
Westerman B, Mattiroli F, Song JY, van der Poel H, Ponz OB,
Pritchard C, Cornelissen-Steijger P, et al: Akt-mediated
phosphorylation of Bmi1 modulates its oncogenic potential, E3
ligase activity, and DNA damage repair activity in mouse prostate
cancer. J Clin Invest. 122:1920–1932. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sánchez-Beato M, Sánchez E,
González-Carreró J, Morente M, Díez A, Sánchez-Verde L, Martín MC,
Cigudosa JC, Vidal M and Piris MA: Variability in the expression of
polycomb proteins in different normal and tumoral tissues. A pilot
study using tissue microarrays. Mod Pathol. 19:684–694. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Martínez-Romero C, Rooman I, Skoudy A,
Guerra C, Molero X, González A, Iglesias M, Lobato T, Bosch A,
Barbacid M, et al: The epigenetic regulators Bmi1 and Ring1B are
differentially regulated in pancreatitis and pancreatic ductal
adenocarcinoma. J Pathol. 219:205–213. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bosch A, Panoutsopoulou K, Corominas JM,
Gimeno R, Moreno-Bueno G, Martín-Caballero J, Morales S, Lobato T,
Martínez-Romero C, Farias EF, et al: The Polycomb group protein
RING1B is overexpressed in ductal breast carcinoma and is required
to sustain FAK steady state levels in breast cancer epithelial
cells. Oncotarget. 5:2065–2076. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Su WJ, Fang JS, Cheng F, Liu C, Zhou F and
Zhang J: RNF2/Ring1b negatively regulates p53 expression in
selective cancer cell types to promote tumor development. Proc Natl
Acad Sci USA. 110:1720–1725. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pan MR, Peng G, Hung WC and Lin SY:
Monoubiquitination of H2AX protein regulates DNA damage response
signaling. J Biol Chem. 286:28599–28607. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu CY, Kang HY, Yang WL, Wu J, Jeong YS,
Wang J, Chan CH, Lee SW, Zhang X, Lamothe B, et al: Critical role
of monoubiquitination of histone H2AX protein in histone H2AX
phosphorylation and DNA damage response. J Biol Chem.
286:30806–30815. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hongyun S and Shuchai Z:
Radiosensitization of esophageal cancer cells ECA109 by knockdown
of H2AX. Thorac Cancer. 4:1759–7706. 2013.
|
|
11
|
Song W, Li H, Tao K, Li R, Song Z, Zhao Q,
Zhang F and Dou K: Expression and clinical significance of the stem
cell marker CD133 in hepatocellular carcinoma. Int J Clin Pract.
62:1212–1218. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Song LB, Zeng MS, Liao WT, Zhang L, Mo HY,
Liu WL, Shao JY, Wu QL, Li MZ, Xia YF, et al: Bmi-1 is a novel
molecular marker of nasopharyngeal carcinoma progression and
immortalizes primary human nasopharyngeal epithelial cells. Cancer
Res. 66:6225–6232. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
14
|
Bartek J and Lukas J: Chk1 and Chk2
kinases in checkpoint control and cancer. Cancer Cell. 3:421–429.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen J, Xu H, Zou X, Wang J, Zhu Y, Chen
H, Shen B, Deng X, Zhou A, Chin YE, et al: Snail recruits Ring1B to
mediate transcriptional repression and cell migration in pancreatic
cancer cells. Cancer Res. 74:4353–4363. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rai K, Akdemir KC, Kwong LN, Fiziev P, Wu
CJ, Keung EZ, Sharma S, Samant NS, Williams M, Axelrad JB, et al:
Dual roles of RNF2 in melanoma progression. Cancer Discov.
5:1314–1327. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
van der Stoop P, Boutsma EA, Hulsman D,
Noback S, Heimerikx M, Kerkhoven RM, Voncken JW, Wessels LF and van
Lohuizen M: Ubiquitin E3 ligase Ring1b/Rnf2 of polycomb repressive
complex 1 contributes to stable maintenance of mouse embryonic stem
cells. PLoS One. 3:e22352008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
van der Velden YU, Wang L, Querol Cano L
and Haramis AP: The polycomb group protein ring1b/rnf2 is
specifically required for craniofacial development. PLoS One.
8:e739972013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yamamoto Y, Abe A and Emi N: Clarifying
the impact of polycomb complex component disruption in human
cancers. Mol Cancer Res. 12:479–484. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen S, Chen J, Zhan Q, Zhu Y, Chen H,
Deng X, Hou Z, Shen B, Chen Y and Peng C: H2AK119Ub1 and H3K27Me3
in molecular staging for survival prediction of patients with
pancreatic ductal adenocarcinoma. Oncotarget. 5:10421–10433. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hasegawa K, Sin HS, Maezawa S, Broering
TJ, Kartashov AV, Alavattam KG, Ichijima Y, Zhang F, Bacon WC,
Greis KD, et al: SCML2 establishes the male germline epigenome
through regulation of histone H2A ubiquitination. Dev Cell.
32:574–588. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zaaroor-Regev D, de Bie P, Scheffner M,
Noy T, Shemer R, Heled M, Stein I, Pikarsky E and Ciechanover A:
Regulation of the polycomb protein Ring1B by self-ubiquitination or
by E6-AP may have implications to the pathogenesis of Angelman
syndrome. Proc Natl Acad Sci USA. 107:6788–6793. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Leung JW, Agarwal P, Canny MD, Gong F,
Robison AD, Finkelstein IJ, Durocher D and Miller KM: Nucleosome
acidic patch promotes RNF168- and RING1B/BMI1-dependent H2AX and
H2A ubiquitination and DNA damage signaling. PLoS Genet.
10:e10041782014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Boquoi A, Arora S, Chen T, Litwin S, Koh J
and Enders GH: Reversible cell cycle inhibition and premature aging
features imposed by conditional expression of p16Ink4a. Aging Cell.
14:139–147. 2015. View Article : Google Scholar :
|
|
25
|
Blanco D, Vicent S, Fraga MF,
Fernandez-Garcia I, Freire J, Lujambio A, Esteller M,
Ortiz-de-Solorzano C, Pio R, Lecanda F, et al: Molecular analysis
of a multistep lung cancer model induced by chronic inflammation
reveals epigenetic regulation of p16 and activation of the DNA
damage response pathway. Neoplasia. 9:840–852. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wen W, Peng C, Kim MO, Ho Jeong C, Zhu F,
Yao K, Zykova T, Ma W, Carper A, Langfald A, et al: Knockdown of
RNF2 induces apoptosis by regulating MDM2 and p53 stability.
Oncogene. 33:421–428. 2014. View Article : Google Scholar :
|
|
27
|
Shapiro GI, Edwards CD, Ewen ME and
Rollins BJ: p16INK4A participates in a G1 arrest
checkpoint in response to DNA damage. Mol Cell Biol. 18:378–387.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Loriot Y, Mordant P, Dugue D, Geneste O,
Gombos A, Opolon P, Guegan J, Perfettini JL, Pierre A, Berthier LK,
et al: Radiosensitization by a novel Bcl-2 and Bcl-XL inhibitor
S44563 in small-cell lung cancer. Cell Death Dis. 5:e14232014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou C, Yang F, Xi W, Wei M, Zheng G, Wang
W, Yang A, Zhang J and Wen W: Effect of RNF2 knockdown on apoptosis
and radiosensitivity in glioma U87 cells. Xi Bao Yu Fen Zi Mian Yi
Xue Za Zhi. 30:471–475. 2014.In Chinese. PubMed/NCBI
|