Introduction
Mesenchymal stem cells (MSCs) derived from the
mesoderm in early development are multipotential stem cells which
have the properties of self-renewal and multilineage
differentiation. They are widely present in all types of tissues
and can differentiate in vitro into cells such as neurons,
osteoblasts, chondrocytes, myocytes and adipocytes under certain
conditions. Therefore, they are highly valuable in applications of
cell replacement therapy and tissue engineering. In 1991,
McElreavey et al isolated and cultured fibroblasts with the
capability of multilineage differentiation from Wharton's jelly of
the human umbilical cord (1).
These human umbilical cord-derived MSCs (hUMSCs) have been shown to
have greater advantages than MSCs derived from bone marrow,
placenta and other tissues. Firstly, ethical concerns regarding the
source of the stem cells (umbilical cord) are diminished, and they
can be obtained at low cost. Secondly, hUMSCs have been determined
not to cause teratomas and can inhibit cancer cells. Additionally,
due to their low immunogenicity and genetic stability, as well as
functions in immunoregulation, stroma support, paracrine signaling
and migration, hUMSCs have good clinical therapeutic potential.
There have been literature reports using
antioxidants, such as thioglycerol, 2-mercaptoethanol,
dimethylsulphoxide (DMSO) and butylhydroxsanisole, to
experimentally induce MSCs to differentiate into neuron-like cells
(2). However, these chemicals
cannot be used in live animals due to toxicity. Other researchers
proposed traditional Chinese medicine and compound preparations
with no or low cytotoxicity to induce bone marrow-derived MSCs
(BMSCs) to differentiate into neuron-like cells and achieved
satisfactory results (3). However,
reports on the use of a traditional Chinese medicine to induce
hUMSCs to differentiate into neuron-like cells are rare.
Tetramethylpyrazine (TMP) is an active alkaloid
(2,3,5,6-tetramethylpyrazine) separated and purified from a Chinese
medicine called Ligusticum wallichii, which belongs to the
family Umbelliferae. The saturation concentration of TMP in aqueous
solution at 37°C was 4.67 mg/ml. TMP has been demonstrated to
promote expansion of small arteries and enhance blood circulation
to prevent blood stasis and antiplatelet aggregation. It has also
been shown to antagonize calcium ion flow and to exhibit
antithrombotic, antioxidant and antifibrotic properties (4). Although the mechanisms of action have
not been clearly defined, it was suggested that the protective
effects and related mechanism of TMP against central lesions may
largely be attributed to inhibition of calcium channels,
antioxidation, resistance of neuronal apoptosis and increase in
heat shock protein expression (5).
In this study, we examined various doses of TMP to induce hUMSCs to
differentiate into neuron-like cells and determined the optimal
inductive concentration. Possible induction mechanism is also
discussed.
Materials and methods
Materials
Full-term pregnancy cesarean neonatal human
umbilical cords were provided by Department Obstetrics/Gynecology,
The Second Affiliated Hospital of Hebei Medical University (Hebei,
China). This study was approved by the Ethics Committee of the
Second Hospital of Hebei Medical University. The following reagents
were obtained commercially: TMP (20 mg; purity: HPLC ≥98%; batch
no. 120919 Chengdu Preferred Biological Technology Co., Ltd,
Chengdu, China); low glucose Dulbecco's Modified Eagle's Medium
(L-DMEM)/F-12, high glucose-DMEM (H-DMEM)/F-12 and fetal bovine
serum (FBS) (Gibco, Grand Island, NY, USA); trypsin (Amresco, LLC,
Solon, OH, USA); EDTA and DMSO (Sigma-Aldrich, St. Louis, MO, USA);
Triton X-100 (Sinopharm Chemical Reagent Co., Ltd., Beijing,
China); penicillin-streptomycin (Beijing Solarbio Science &
Technology Co., Ltd., Beijing, China); paraformaldehyde (Tianjin
Institute of Chemical Preparations, Tianjin, China); epidermal
growth factor (EGF) (human EGF, PHG0314; Beijing Jiamei North
Biological Technology Co., Ltd., Beijing, China); fluorescein
isothiocyanate (FITC)-CD19, FITC-CD34, phycoerythrin (PE)-CD11b,
PE-CD73, PE-CD90, PE-CD45, PE-CD105 and glial fibrillary acidic
protein (GFAP) (BD Biosciences, Franklin Lakes, NJ, USA);
neurofilament protein (NF-H), neuron-specific enolase (NSE) (Cell
Signaling Technology, Inc., Danvers, MA, USA); anti-rabbit
immunoglobulin G (IgG) (1:2,000; Affinity Biosciences, Cincinnati,
OH, USA); PS immunohistochemistry kit (Beijing Zhongshan Golden
Bridge Biotechnology Co., Ltd., Beijing, China); Taq polymerase
chain reaction (PCR) star mix (Beijing GenStar Biosolutions Co.,
Ltd., Beijing, China); EasyScript First-Strand cDNA synthesis
supermix (TransGen Biotech Co., Ltd., Beijing, China); cell RIPA
lysis buffer and phenylmethylsulfonyl fluoride (Beijing Solarbio
Science & Technology Co., Ltd.).
Isolation and culture of hUMSCs
Umbilical cords were placed in H-DMEM/F-12 culture
medium under aseptic conditions, stored at 4°C and then timely
transported to a cell culture room to carry out the following
steps. Each umbilical cord was rinsed thoroughly with D-Hank's
medium. After removing the blood sample, umbilical artery and
umbilical vein, the umbilical cord mesenchymal tissue was cut into
1 mm3 pieces, digested with 0.2% collagenase II and
placed in a culture flask containing 2 ng/ml EGF, 20% FBS, 25 mM
L-Glu and 100 U/ml penicillin streptomycin/mixture at 37°C with 5%
CO2 and saturation humidity to obtain primary cells.
Half of the culture medium was replaced after 24 h, and all the
medium was replenished every 3 days. When the cells achieved 80–90%
confluency, the medium was removed, and cells were rinsed two times
with phosphate-buffered saline (PBS) and then digested with trypsin
(0.25%)-EDTA (0.2 g/l) into single cells for passaging at the ratio
of 1:3. The culture medium was H-DMEM/F-12 containing 100 U/ml of a
penicillin-streptomycin mixture and 10% FBS.
Analysis of cellular phenotype of
hUMSCs
In the logarithmic phase of growth, hUMSCs were
digested with trypsin and rinsed with PBS, and then the single cell
suspension was aliquotted into 10 microcentrifuge tubes at
1×106/tube. Separately, mouse anti-human monoclonal
antibodies CD11-PE, CD45-PE, CD73-PE, CD90-PE, CD105-PE, human
leukocyte antigen (HLA)-DR-PE, CD19-FITC and CD34-FITC (each 5 μl)
were added to eight of the tubes, while anti-mouse IgG1-PE and
anti-mouse IgG1-FITC (each 7 μl) were added to the other two tubes
as isotype controls. After mixing the contents thoroughly, the
tubes were incubated at 4°C for 30 min. Thereafter, the cells were
rinsed with PBS and then centrifuged. The supernatant was
discarded, and the cells were resuspended with 400 μl of PBS for
analysis by flow cytometry.
Differentiation of hUMSCs into
neuron-like cells
Cells were obtained from the 3rd passage in
logarithmic growth phase with good growth conditions, hUMSCs were
rinsed twice with PBS and digested with trypsin (0.25%)-EDTA (0.2
g/l). The digestion was terminated with FBS, and then the cells
were centrifuged, resuspended in culture medium and seeded at
1×105/ml into five 6-well cell culture plates coated
with poly-lysine. The plates were divided randomly into groups A–D
and then placed in the incubator. When the cells achieved 70–80%
confluency, TMP induction medium was added to cells of groups A–C
at the concentrations of 4.67, 2.34 and 1.17 mg/ml, respectively.
Group D was treated with L-DMEM only and served as control. All the
plates were cultured in an incubator at 37°C with 5% CO2
and saturation humidity for 6 h. Morphological changes were
observed with an inverted phase contrast microscope each 0.5 h.
Identification of differentiated
cells
After induction for 6 h, expression levels of
neuronal-specific proteins NSE, NF-H and GFAP were detected by
immunocytochemistry. Briefly, after discarding the culture medium
from the culture plates, the cells were rinsed once with PBS gently
and fixed with 4% paraformaldehyde for 20 min at room temperature.
After this step and subsequent steps, the cells were rinsed with
PBS three times for 5 min each time, unless otherwise noted. The
cell membranes were disrupted with 0.5% Triton X-100 in PBS for 15
min at room temperature away from light and then incubated with 3%
H2O2 at room temperature for 5 min. Normal
goat serum was added to the cells for blocking at room temperature
for 15 min and then removed without washing. Primary antibodies to
NSE, GFAP and NF-H were added at a 1:200 dilution. After incubation
at 4°C overnight, the cells were incubated with the appropriate
biotinylated secondary antibody at room temperature for 15 min,
followed by horseradish peroxidase-conjugated streptavidin at room
temperature for 15 min. Finally, the cells were stained with
freshly prepared DAB for 1 min and counterstained with hematoxylin
before finally rinsed repeatedly with water.
RT-PCR
Total RNA was extracted by TriPure Reagent (Aidlab
Biotechnologies Co., Ltd., Beijing, China) following the
manufacturer's instructions and quantified with a Ultramicro
ultraviolet visible light meter (Gene Company, Ltd., Hong Kong,
China). cDNA was synthesized from total RNA with an EasyScript
First-Strand cDNA synthesis supermix (TransGen Biotech Co., Ltd.)
following the manufacturer's instructions. Semi-quantitative PCR
was performed using Taq PCR star mix (Beijing GenStar Biosolutions
Co., Ltd.) with the following reaction conditions: 1 cycle of 94°C
for 3 min; 30 cycles of 94°C for 30 sec, and 72°C for 30 sec; and a
final extension at 72°C for 10 min. A 10 μl sample of the PCR
product was analyzed by electrophoresis, and the gene expression
levels were calculated using GAPDH as the internal standard. The
primers were designed using Primer 5.0 software and are shown in
Table I.
 | Table IThe primer sequences used for
RT-polymerase chain reaction analysis in this study. |
Table I
The primer sequences used for
RT-polymerase chain reaction analysis in this study.
| Genes | Primer sequences | Sizes (bp) | Temperature (°C) |
|---|
| NSE | F:
GGCACTCTACCAGGACTTTG
R: GCGATGACTCACCATAACCC | 398 | 61.7 |
| NF-H | F:
TGAACACAGACGCTATGCGCTCAG
R: CACCTTTATGTGAGTGGACACAGAG | 286 | 61.2 |
| GAPDH | F:
AGAAGGCTGGGGCTCATTTG
R: AGGGGCCATCCACAGTCTTC | 258 | 64.0 |
Western blot analysis
Cell lysates were prepared with cell RIPA lysis
buffer and phenylmethylsulfonyl fluoride (Beijing Solarbio Science
& Technology Co., Ltd.), and protein concentrations were
determined with an ultramicro-ultraviolet visible light meter (Gene
Company, Ltd.). Total protein (16 mg) of each lysate was
electrophoresed in a 10% SDS-PAGE gel and transferred to a
polyvinylidene fluoride membrane (Beijing Solarbio Science &
Technology Co., Ltd.). The membranes were blocked in 10% non-fat
milk for 2 h and incubated with the GFAP, NF-H and NSE primary
antibodies at 4°C overnight, After rinsing with PBS for 5 min, the
membranes were incubated with anti-rabbit IgG (1:2,000; Affinity
Biosciences) for 2 h at room temperature and visualized by
SuperSignal West Pico Chemiluminescent Substrate (Thermo Fisher
Scientific, Waltham, MA, USA) using the Image AlphaEaseFC system
(Alpha Innotech Co., San Leandro, CA, USA).
Calculation of frequency of neuron-like
cells
A total of 10 non-overlapping representative fields
under an inverted microscope were selected in each group, in which
the total number of cells and neuron-like cells were counted. The
proportion of neuron-like cells was calculated and averaged from
the 10 fields for each group. Results were presented as mean ±
standard deviation.
Statistical analysis
Statistical analysis was performed by using SPSS
19.0 software (SPSS, Inc., Chicago, IL, USA). Comparison on the
experimental data with a completely randomized design was tested by
analysis of variance among groups and the Student-Newman-Keuls
method (q-test). Difference in the proportion of neuron-like cells
among groups was analyzed by the Chi-square test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Growth and morphological changes of
hUMSCs
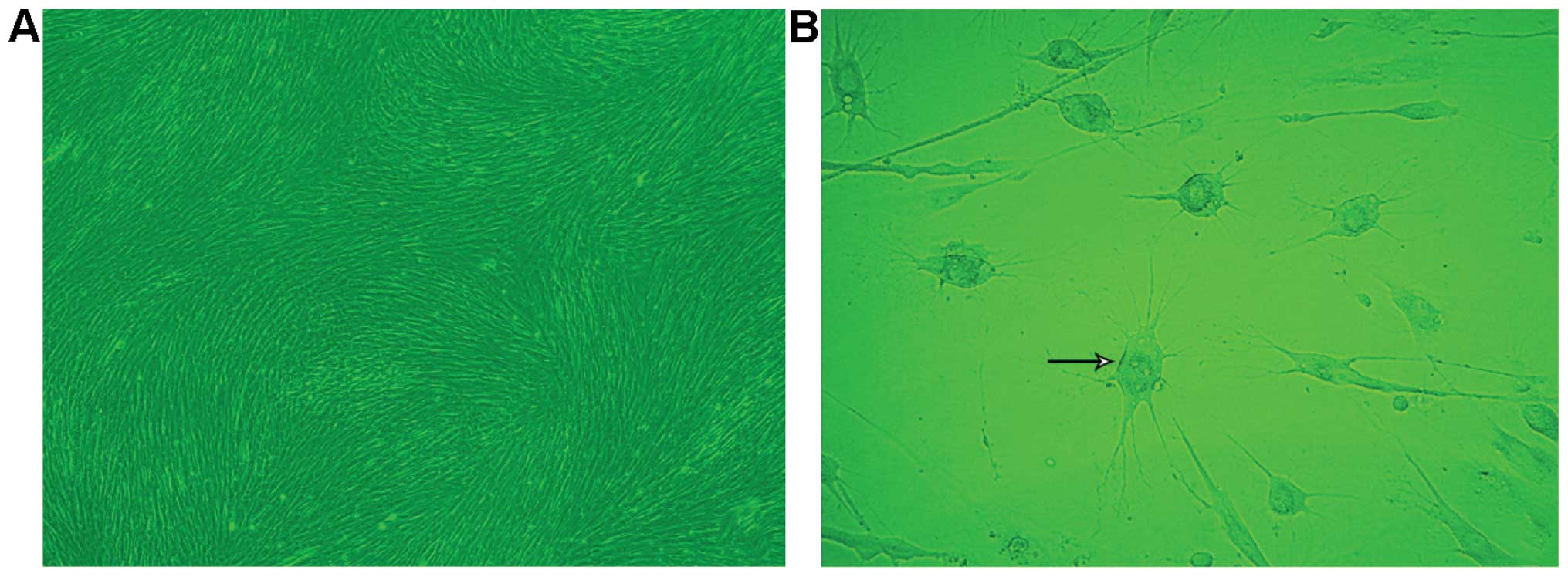
Primary cultured cells were passaged at the ratio of
1:1, and most cells were adherent after 12 h. However, the adherent
cells were not outstretched, but appeared triangular or diamond in
shape. The cell culture medium was replaced to remove non-adherent
cells after 24 h. The adherent cells proliferated rapidly and
became significantly larger and more uniform, long fusiform cells
after 48 h of culture. They achieved 80–90% confluency on the 7th
day as a monolayer arranged in a radial or spiral shaped pattern.
After digestion, the hUMSCs cells floated as spherical single cells
in the culture medium and were passaged at the ratio of 1:3 after
centrifugation. Most cells were adherent 24 h after passaging and
achieved 80–90% confluency on the 7th day with a radial or spiral
shaped arrangement (Fig. 1A). The
hUMSCs of the 10th passaged maintained a strong proliferative
capacity.
Cellular phenotype of hUMSCs
We detected the cellular phenotype of the 2nd, 5th
and 10th passages of hUMSCs by flow cytometry and found that all
generations of the cells tested co-expressed CD105, CD90 and CD73,
but not CD11b, CD34, CD19, CD45 and histocompatibility antigen
HLA-DR (major histocompatibility complex class II) (Fig. 2).
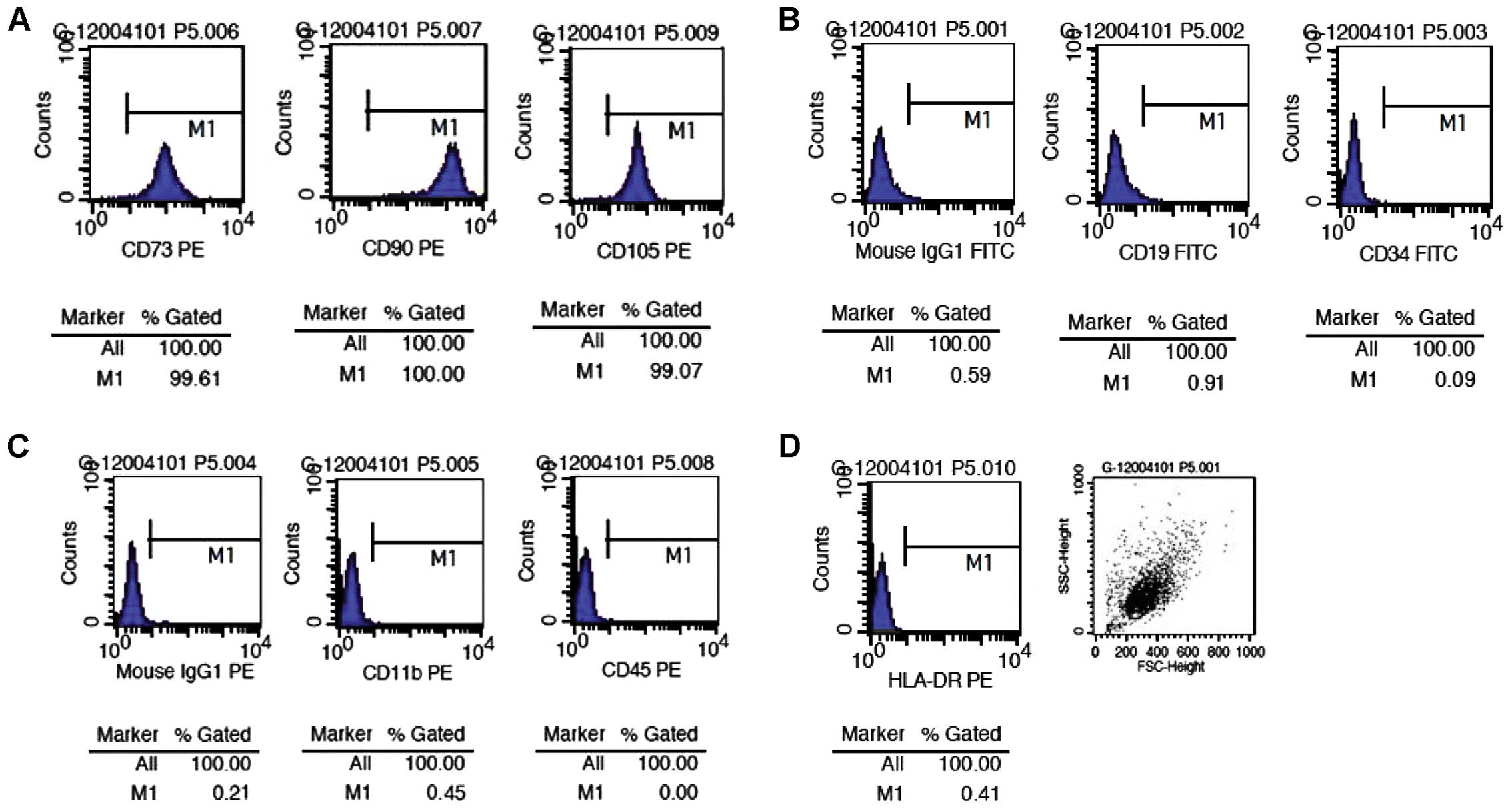 | Figure 2Surface marker expression in human
umbilical cord-derived mesenchymal stem cells (hUMSCs) detected by
flow cytometry. The percentages of (A) hUMSCs expressing CD73, CD90
and CD105 were 99.61, 100 and 99.07%, respectively. However, the
percentages of (B and C) hUMSCs expressing CD19, CD34, CD11b and
CD45, and (B and D) the expression of isotype controls and human
leukocyte antigen (HLA)-DR were very low. These results confirmed
that the cells we cultured were mesenchymal stem cells, and not
hematopoietic stem cells. PE, phycoerythrin; IgG, immunoglobulin G;
FITC, fluorescein isothiocyanate. |
Morphological and quantitative changes of
neuron-like cells in each group
After treatment with TMP for 0.5 h, 40% of cells in
group A contracted into an oval shape and extended protuberances
from their cell bodies (Fig. 1B).
After treatment for 1 h, the ratio of neuron-like cells reached
55%, with the cell bodies contracted and the protuberances
lengthened further. At this time, the cells appeared connected in a
net-like pattern. After treatment for 1.5 h, ~85% of the cells
appeared as neuron-like cells. After treatment for 4 h, some of the
neuron-like cells began to detach, and the ratio was reduced to
~80% in the 6th h. In group B, ~45% of the cells were neuron-like
cells after treated with TMP for 1.5 h, and the ratio increased to
80% until the 2nd h. Thereafter, some of the neuron-like cells
started to become non-adherent, and the ratio was reduced to 70% in
the 6th h. In group C, ~30% of the cells were neuron-like cells
after treatment with TMP for 4.5 h; however, some became detached,
and the ratio was ~55% at the 6th h. The control group D had no
significant changes before or after treated with medium alone. As
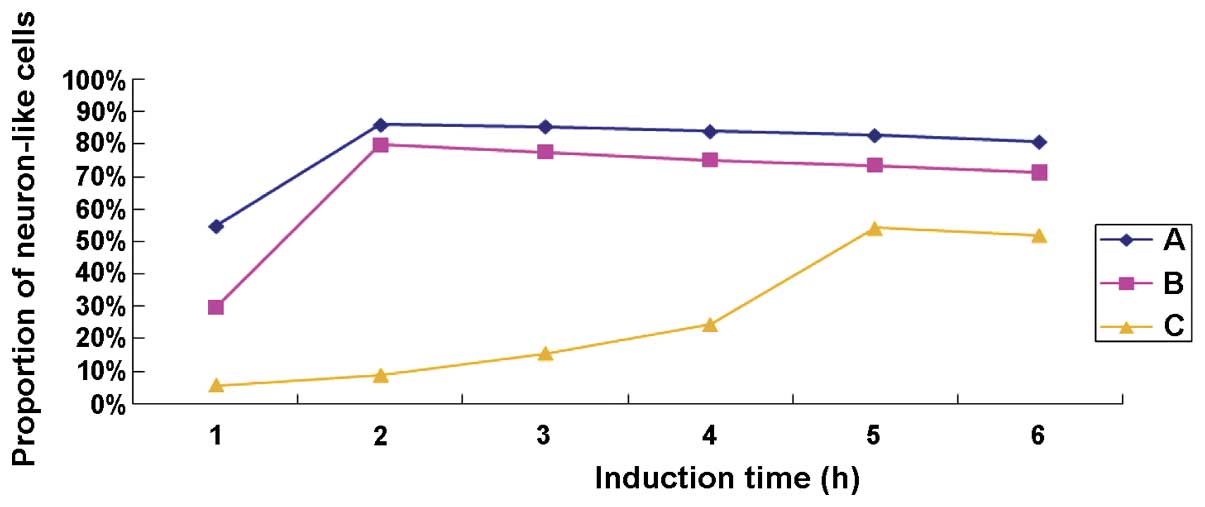
shown in Fig. 3, the proportion of
TMP-induced neuron-like cells was, from high to low, group A
>group B >group C at each time point, and it declined
gradually over the induction time indicating that the neuron-like
cells were continually becoming detached. The neuron-like cells in
group C required more time to reach the highest ratio, and the
detaching cells could also be seen during the process of
differentiation. Treatment of TMP at the concentration of 4.67
mg/ml in group A was employed in the subsequent experiment to
detect the neuronal-specific markers by western blotting.
Immunocytochemical detection of
neuronal-specific markers
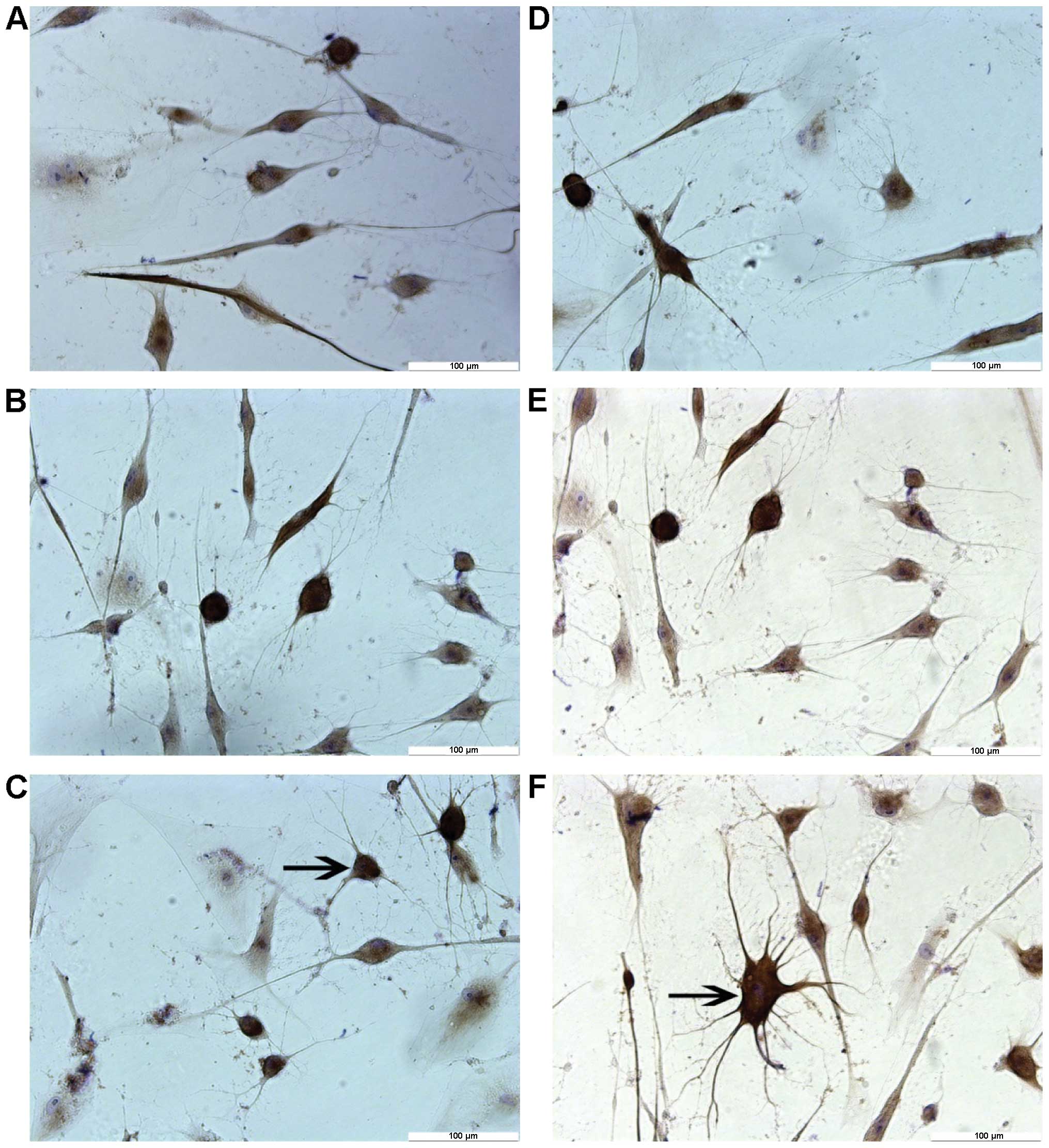
Results from immunocytochemical staining showed that
most neuron-like cells treated by different concentrations of TMP
for 6 h were positive for NF-H and NSE, but negative for GFAP
(Fig. 4). By contrast, no cells
were found positively expressing these markers in the control
group. Negative staining was evidenced when PBS was used to replace
the primary antibody (data not shown). As shown in Table II, differences in the expression
levels of NF-H and NSE among these groups were significant
(P<0.01).
| Table IIComparison of antigen expression in
neuron-like cells induced from hUMSCs after 6 h of TMP treatment
(%, n=3). |
Table II
Comparison of antigen expression in
neuron-like cells induced from hUMSCs after 6 h of TMP treatment
(%, n=3).
| Group | NF-H | NSE |
|---|
| A-TMP 4.67 mg/ml | 80.79±4.36 | 79.76±2.46 |
| B-TMP 2.34 mg/ml | 71.30±1.94a,b | 69.83±4.42a,c |
| C-TMP 1.17 mg/ml | 52.01±3.66a,c | 50.18±4.07a,c |
| D-Control | 0 | 0 |
Western blot detection of
neuronal-specific markers
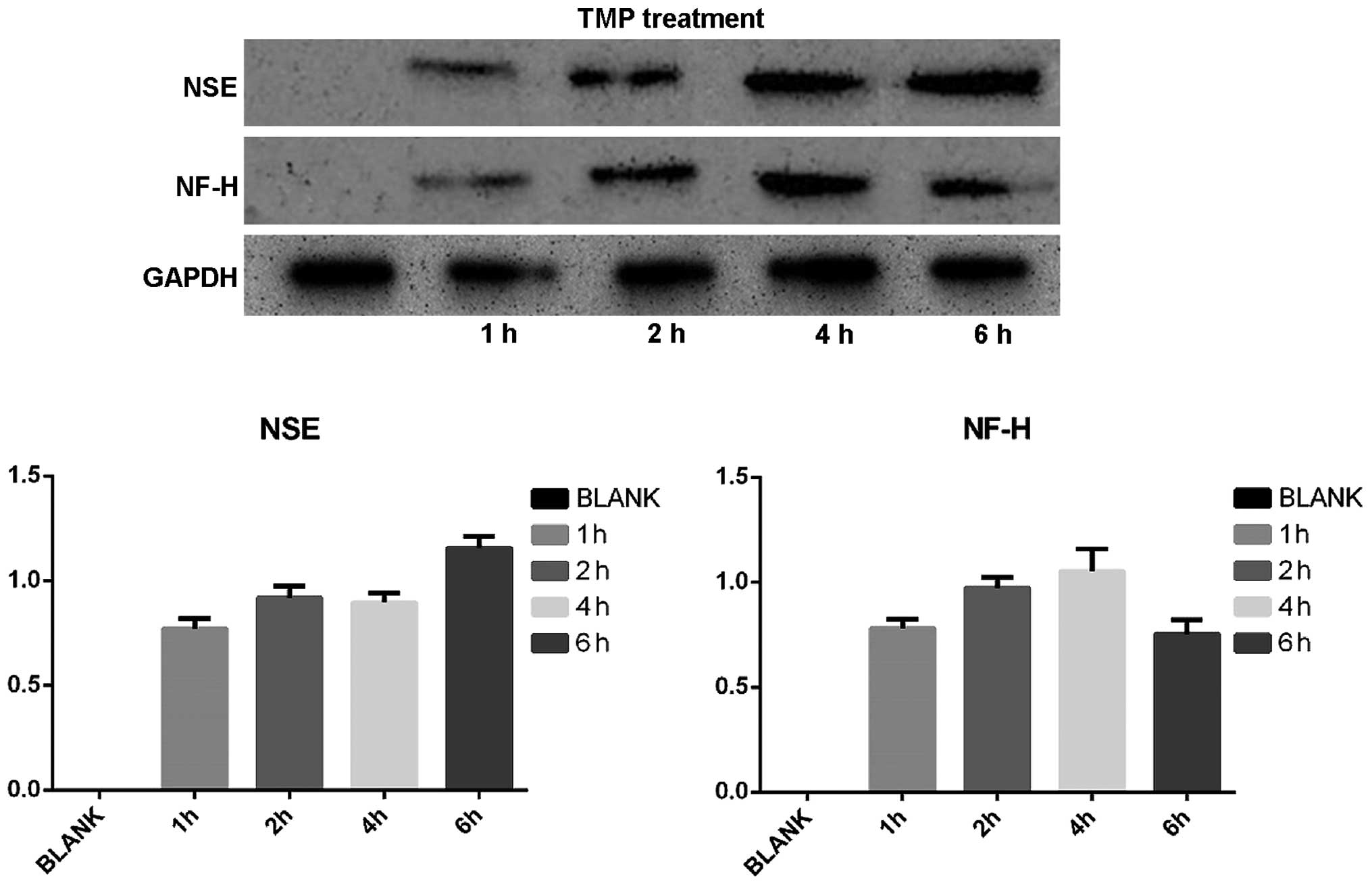
On the basis of neuron-like cell positive rate, we
selected the TMP treatment at a concentration of 4.67 mg/ml to
detect the expression of NSE, NF-H, GFAP using western blotting.
Protein expression of cells expressing neuronal markers as well as
GAPDG was observed after TMP treatment. The calculated relative
transcript level for protein expression of NSE was 0, 0.717±0.097,
0.919±0.056, 1.097±0.143 and 1.157±0.055 before treatment, for 1,
2, 4 and 6 h, respectively; whereas, the relative level for NH-F
expression was 0, 0.780±0.10, 0.973±0.150, 1.053±0.107 and
0.753±0.094, respectively. No expression of GFAP was detected
before or after induction, whereas all tested samples showed high
expression of GAPDH after TMP treatment (Fig. 5).
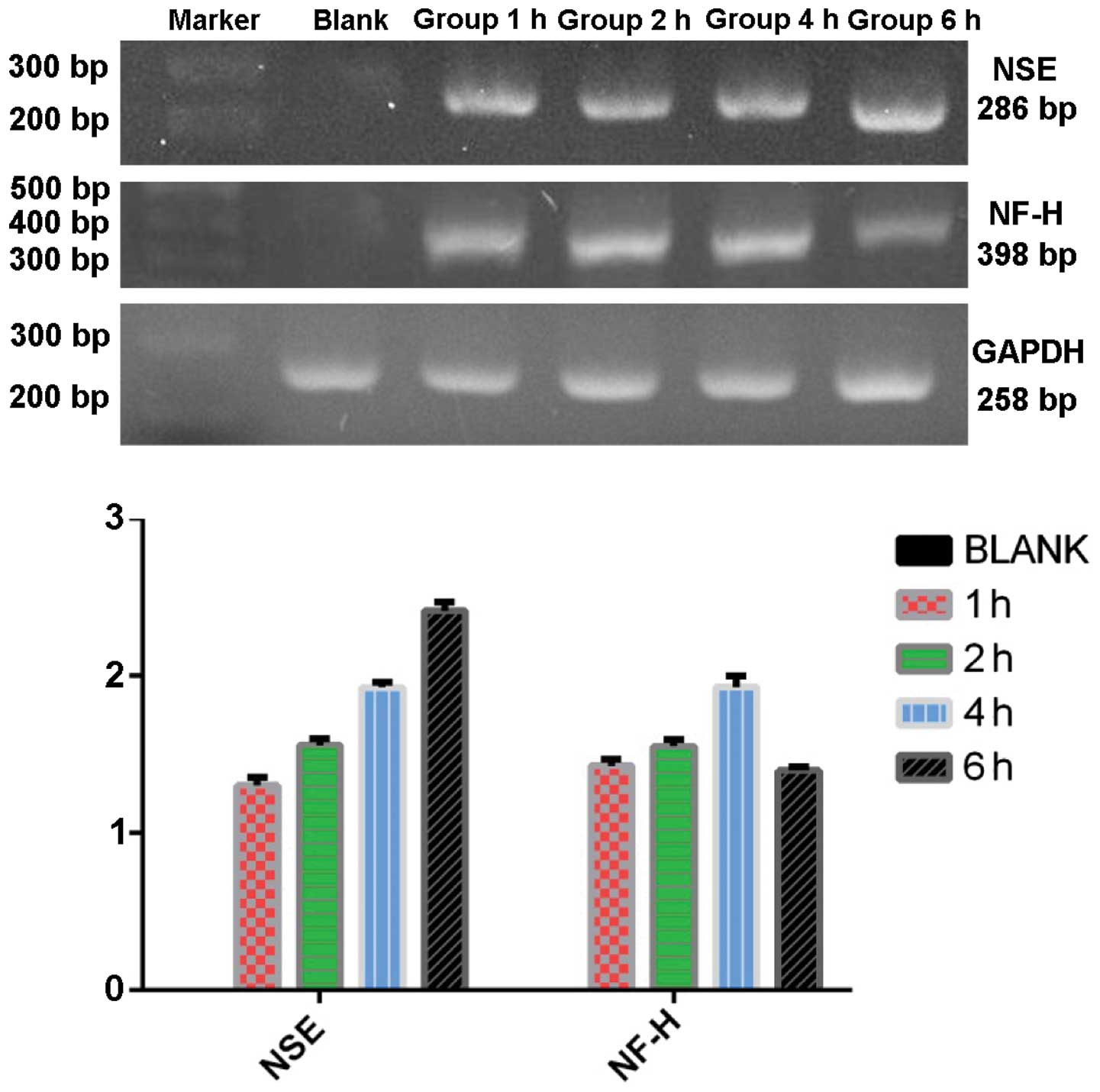
RT-PCR detection of neuronal-specific
markers
RT-PCR was used to detect the expression of NSE,
NF-H, and GFAP of the group C in each time point after TMP
treatment. The mRNA expression of NSE was 0, 1.303±0.031,
1.558±0.025, 1.927±0.019 and 2.415±0.033 in proper order. The mRNA
expression of NF-H was 0, 1.429±0.025, 1.551±0.024, 1.930±0.042 and
1.398±0.014 in proper order. There was no expression of GFAP before
or after induction and all the groups showed high expression of
GAPDH at each time point (Fig.
6).
Discussion
The plasticity of stem cells refers to the ability
of adult stem cells to lose their phenotype of a specific tissue or
germ layer and differentiate into other types of cells (6–8).
Typically, MSCs exhibit cell plasticity by being able to
differentiate into bone, cartilage, smooth muscle, skeletal muscle
and cardiac muscle, as well as cells from other germ layers such as
skin and liver (9–19). Woodbury et al (2) first reported in 2000 that BMSCs can
differentiate into neuron-like cells under certain conditions, a
finding that has attracted significant attention. Soon afterwards,
numerous domestic and foreign laboratories carried out in
vitro and in vivo studies on the neural differentiation
of MSCs from different species and sources. These studies have
shown that MSCs of rats, mice, humans, rabbits and other mammals
can be induced to differentiate into neuron-like cells under
certain conditions. In this context, newborn umbilical cords, as a
reliable source of MSCs that can be obtained non-invasively and
without ethical constraints, have been widely used in stem cell
transplantation therapy and experiments of neural
differentiation.
Some inducers, including chemical inducer,
neurotrophic factor, and Chinese medicine active ingredients and
their preparations, have been shown to be able to differentiate
MSCs into neuron-like cells expressing surface antigen markers of
neural cells. Our study confirmed that the TMP monomer
(2,3,5,6-tetramethylpyrazine), an active ingredient in Chinese
medicine, could effectively induce hUMSCs to differentiate into
neuron-like cells in vitro and express NSE and NF-H, but not
GFAP. Moreover, the optimal concentration of TMP for obtaining
these inductive effects was determined to be 4.67 mg/ml, which is
the saturation concentration of TMP in aqueous solution at 37°C
(20).
Different inducers have different mechanisms of
facilitating neuronal cell differentiation. The common feature of
chemical inducers is their ability to increase the intracellular
concentration of cAMP, suggesting that the second messenger is
involved in the induction of MSCs to differentiate into neural
precursor cells (21). Butylated
hydroxyanisole, β-mercaptoethanol and other antioxidants promote an
increase of intracellular cAMP in different ways and then activate
the PKA pathway and phosphorylation of downstream target proteins.
Moreover, PKC has an important role in the induction process to
maintain cell survival. The MEK-ERK signaling pathway also plays an
important role in the process of neural cell induction from MSCs.
Neurotrophic factor inducers include basic fibroblast growth factor
(bFGF), EGF, retinoic acid (RA), nerve growth factor (NGF) and
brain-derived neurotrophic factor (BDNF). In the neuronal
differentiation of mouse MSCs, the medium used by Kohyama et
al (22) included a
demethylation agent (5-azaC), NGF, NT-3 and BDNF, while Jin et
al (23) successfully used
EGF, bFGF, RA and NGF. The mechanism by which neurotrophic factors
promote neural differentiation of MSCs may involve their high
concentrations which can potentially simulate the microenvironment
of embryonic developmental stages of neurogenesis, thereby
promoting the differentiation of MSCs into neural cells. Previous
studies have shown that neurotrophic factors increase the
expression of MSC membrane proteins TrkA, TrkB and TkrC, which are
neurotrophin receptors. The binding of neurotrophin and its
receptor initiates changes in some gene expression (24). Traditional Chinese medicines may
have antioxidant and anti-ischemic properties and other effects, as
well as improve microcirculation. Previously, we also found that
they have protective effects against nerve cell injury (25). TMP may play a role as antioxidant
in promoting the increase in the intracellular second messenger
cAMP, which subsequently activates the PKA pathway and the MEK-ERK
signaling pathway, and thus plays a role in the neural induction
process. Liu et al (26)
and others have pointed out that TMP as a Ca2+ chelator,
via the inhibition of the intracellular Ca2+ signal, can
upregulate the expression of NSE and Nurrl, thereby accelerating
the differentiation of hUMSCs into nerve cells. Zhao et al
(27) indicated that
sub-totipotent stem cells still express sub-totipotent genes after
the embryo has developed into adulthood, but they gradually lose
part of the original stem cell phenotype. If the tissue-specific
gene expression programs of such cells were activated in an
appropriate microenvironment, they can differentiate into various
histocytes. hUMSCs are sub-totipotent stem cells, but whether the
microenvironment provided by TMP can activate the specific gene
expression program of nerve cells to further differentiate into
neural cells will require further study.
The ultimate goal of inducing MSCs to differentiate
into nerve cells is to use them in vivo as a replacement
therapy. However, electrophysiological evidence of whether the
induced cells can function as nerve cells is still lacking. Some
researchers have claimed that the morphological change of cells was
initiated by the toxicity of the inducers, and the long
protuberances were caused by the reduction of cytoplasm due to
destruction of the actin cytoskeleton. According to this argument,
the expression of nerve cell antigen markers is more likely to be
result of an abnormal combination of protein interactions rather
than a series of genetic events (28). Moreover, most induced nerve cells
live for only a short period of time in vitro, which creates
a barrier to their effective use in transplantation therapies.
In conclusion, our study demonstrated that TMP can
induce hUMSCs to effectively differentiate into neuron-like cells
with the optimal concentration of 4.67 mg/ml. After induction, the
NSE and NF-H of the neuron-like cells were positive but the GFAP-2
was negative. Future studies will verify whether the differentiated
cells are bona fide nerve cells, and the extra-cellular environment
required to maintain neuron-like cells after induction in
vitro also need further investigation, which may provide
additional evidence and rationale for using in vitro induced
and differentiated hUMSCs in potential nerve cell
transplantation.
Acknowledgements
This study was supported by the Topic Outstanding
Youth Science Foundation of Natural Science Fund of Hebei (no.
C2009001547), the Natural Science Fund of Hebei (no. H2013206399),
and the Medical Science Research Key Project of Hebei (no.
20130240).
References
|
1
|
McElreavey KD, Irvine AI, Ennis KT and
McLean WH: Isolation, culture and characterisation of
fibroblast-like cells derived from the Wharton's jelly portion of
humanumbilical cord. Biochem Soc Trans. 19:29S1991. View Article : Google Scholar
|
|
2
|
Woodbury D, Schwarz EJ, Prockop DJ and
Black IB: Adult rat and human bone marrow stromal cells
differentiate into neurons. J Neurosci Res. 61:364–370. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li SH and Guo PD: Research progress about
traditional Chinese medicine (TCM) inducing bone marrow mesenchymal
stem cells to differentiate into neural cell. Gansu J Tradit Chin
Med. 22:68–69. 2009.
|
|
4
|
Liu YX and Gao JZ: Clinical observation of
tetramethylpyrazine early treatment of acute craniocerebral injury.
Chin J Clin Neurosurg. 12:754–755. 2007.(In Chinese).
|
|
5
|
Li YQ and Lan TM: Research progress about
tetramethylpyrazine in the pharmacokinetics of the nervous system
and clinical application. Seek Med Ask The Med. 9:3342011.(In
Chinese).
|
|
6
|
Xin ZC, Wang L, Lin GT, et al: The markers
and tracer marker of mesenchymal stem cells. J Peking Univ.
45:514–517. 2013.(In Chinese).
|
|
7
|
Verfaillie C: Stem cell plasticity.
Hematology. 10(Suppl 1): 293–296. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Filip S, Mokrý J, English D and Vojácek J:
Stem cell plasticity and issues of stem cell therapy. Folia Biol
(Praha). 51:180–187. 2005.
|
|
9
|
Friedman MS, Long MW and Hankenson KD:
Osteogenic differentiation of human mesenchymal stem cells is
regulated by bone morphogenetic protein-6. J Cell Biochem.
98:538–554. 2006. View Article : Google Scholar
|
|
10
|
Wang T, Xu Z, Jiang W and Ma A:
Cell-to-cell contact induces mesenchymal stem cell to differentiate
into cardiomyocyte and smooth muscle cell. Int J Cardiol.
109:74–81. 2006. View Article : Google Scholar
|
|
11
|
Moscoso I, Centeno A, López E,
Rodriguez-Barbosa JI, Santamarina I, Filgueira P, Sánchez MJ,
Domínguez-Perles R, Peñuelas-Rivas G and Domenech N:
Differentiation ‘in vitro’ of primary and immortalized porcine
mesenchymal stem cells into cardiomyocytes for cell
transplantation. Transplant Proc. 37:481–482. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen LB, Jiang XB and Yang L:
Differentiation of rat marrow mesenchymal stem cells into
pancreatic islet beta-cells. World J Gastroenterol. 10:3016–3020.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gang EJ, Jeong JA, Hong SH, Hwang SH, Kim
SW, Yang IH, Ahn C, Han H and Kim H: Skeletal myogenic
differentiation of mesenchymal stem cells isolated from human
umbilical cord blood. Stem Cells. 22:617–624. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu W, Zhang X, Qian H, Zhu W, Sun X, Hu J,
Zhou H and Chen Y: Mesenchymal stem cells from adult human bone
marrow differentiate into a cardiomyocyte phenotype in vitro. Exp
Biol Med (Maywood). 229:623–631. 2004.
|
|
15
|
de la Fuente R, Abad JL, García-Castro J,
Fernández-Miguel G, Petriz J, Rubio D, Vicario-Abejón C, Guillén P,
González MA and Bernad A: Dedifferentiated adult articular
chondrocytes: a population of human multipotent primitive cells.
Exp Cell Res. 297:313–328. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bosnakovski D, Mizuno M, Kim G, Ishiguro
T, Okumura M, Iwanaga T, Kadosawa T and Fujinaga T: Chondrogenic
differentiation of bovine bone marrow mesenchymal stem cells in
pellet cultural system. Exp Hematol. 32:502–509. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bhagavati S and Xu W: Isolation and
enrichment of skeletal muscle progenitor cells from mouse bone
marrow. Biochem Biophys Res Commun. 318:119–124. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pittenger M, Vanguri P, Simonetti D and
Young R: Adult mesenchymal stem cells: Potential for muscle and
tendon regeneration and use in gene therapy. J Musculoskelet
Neuronal Interact. 2:309–320. 2002.
|
|
19
|
Sato Y, Araki H, Kato J, Nakamura K,
Kawano Y, Kobune M, Sato T, Miyanishi K, Takayama T, Takahashi M,
et al: Human mesenchymal stem cells xenografted directly to rat
liver are differentiated into human hepatocytes without fusion.
Blood. 106:756–763. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao LP, Cai JH, He HH, Huang LS and Fu ZH:
Determination of the equilibrium solubility and oil/water apparent
partition coefficient (P) of tetramethylpyrazine by HPLC. Strait
Pharm J. 25:24–26. 2013.
|
|
21
|
Deng W, Obrocka M, Fischer I and Prockop
DJ: In vitro differentiation of human marrow stromal cells into
early progenitors of neural cells by conditions that increase
intracellular cyclic AMP. Biochem Biophys Res Commun. 282:148–152.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kohyama J, Abe H, Shimazaki T, Koizumi A,
Nakashima K, Gojo S, Taga T, Okano H, Hata J and Umezawa A: Brain
from bone: Efficient ‘meta-differentiation’ of marrow
stroma-derived mature osteoblasts to neurons with Noggin or a
demethylating agent. Differentiation. 68:235–244. 2001. View Article : Google Scholar
|
|
23
|
Jin K, Mao XO, Batteur S, Sun Y and
Greenberg DA: Induction of neuronal markers in bone marrow cells:
Differential effects of growth factors and patterns of
intracellular expression. Exp Neurol. 184:78–89. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yuan Y, Yang SY, Han ZC, et al:
Amplification and differentiation towards neuron-like cells of
human umbilical cord derived mesenchymal stem cells. Chin J
Neuromed. 5:230–236. 2006.
|
|
25
|
Sun WX and An HM: The mechanism research
concerning protective effects of traditional chinese medicine on
nerve system injury caused by oxidative stress. Chin J Chin Med.
4:561–564. 2015.(In Chinese).
|
|
26
|
Liu YY, Zhao XX, Zhao HB, Ge BF, Liu XY
and Chen KM: Tetramethylpyrazine induces the differentiation of
mouse bone marrow-derived mesenchymal stem cells into nerve cells
mediated by Ca2+ signaling. J Gansu Agric Univ. 45:1–5.
2010.
|
|
27
|
Zhao CH, Fang BJ, Han Q, et al: Study
about biological property of pluripotent stem cells and
transplantation application. J Chin Microcirc. 8:3452004.
|
|
28
|
Lu P, Blesch A and Tuszynski MH: Induction
of bone marrow stromal cells to neurons: Differentiation,
transdifferentiation, or artifact? J Neurosci Res. 77:174–191.
2004. View Article : Google Scholar : PubMed/NCBI
|