Introduction
Prostate cancer is one of the most common cancers
among males (1,2). The development and progression of
prostate cancer are closely related to androgen-receptor activation
(3,4). Thus, prostate cancer is generally
dependent on androgen and responds to initial androgen-ablation
therapy (5,6). Although androgen ablation therapy
leads to remissions lasting 2 to 3 years, androgen-independent
cancer develops in the majority of patients. Regrettably, there was
only slightly effective therapy for such patients. In 2004,
however, two large randomized clinical trials showed that
docetaxel-based regimens palliate symptoms and prolong survival in
hormone-refractory prostate cancer (7,8).
Subsequently, docetaxel-based regimens have been used as the best
chemotherapy for prostate cancer (9,10).
Moreover, the recent E3805 CHAARTED trial showed that the addition
of docetaxel to standard androgen-ablation therapy extended the
survival of patients with newly diagnosed metastatic
hormone-sensitive prostate cancer by 13 months (11). Docetaxel is thus considered a very
useful anticancer agent for the management of prostate cancer.
Most anticancer chemotherapeutic drugs, including
docetaxel, primarily act by inhibiting proliferation and inducing
apoptosis of cancer cells (12,13).
Docetaxel binds to β-tubulin and stabilizes microtubules, resulting
in G2/M cell-cycle arrest with aberrant mitosis. Along with G2/M
arrest, taxanes induce apoptosis via activation of the extrinsic
and intrinsic death pathways (14–16).
Bcl-2 family proteins play crucial roles in the intrinsic pathway.
The members of the Bcl-2 family can be broadly divided into
anti-apoptotic and pro-apoptotic groups. Bcl-2 family members
regulate the release of cytochrome c from mitochondria,
resulting in activation of executioner caspases (17). Activation of transmembrane death
receptors initiates the extrinsic pathway, which is efficiently
amplified by the intrinsic pathway mediated by a pro-apoptotic
Bcl-2 family member, Bid (18–20).
Bid is cleaved to tBid by activated caspase-8; tBid is then
translocated to mitochondria, where it promotes cytochrome c
release by interacting with Bax and Bak (21,22).
Anti-apoptotic members, such as Bcl-2 and Bcl-xL, bind to
pro-apoptotic members, such as Bax, Bad, or Bid, to rescue cells
from apoptosis (23). Bcl-2 and
related anti-apoptotic proteins are frequently upregulated in many
types of cancer (24), including
prostate cancer (25,26). Because of their anti-apoptotic
potency, the relation between Bcl-2 overexpression and
chemoresistance of cancer cells has been argued frequently
(24,27,28).
However, in the case of taxanes such as docetaxel and paclitaxel,
the relation between Bcl-2 overexpression and chemoresistance
remains controversial. For instance, Inoue et al showed that
Bcl-2 overexpression enhances in vitro sensitivity to
docetaxel in non-small cell lung cancer (29).
The aim of this study was to elucidate the roles of
Bcl-2 status in the death of DU145 cells, an androgen-independent
human prostate cancer cell line, after treatment with a panel of
anticancer drugs, including docetaxel. For this purpose, we
established a panel of DU145 transfectants that express various
levels of Bcl-2 and examined their susceptibility to anticancer
drugs. We also determined whether pro-apoptotic caspases
participate in the docetaxel-induced death of DU145 cells.
Materials and methods
Cell lines and drugs
DU145 cells, an androgen-independent human prostate
cancer cell line (ATCC, Manassas, VA, USA), were cultured in
Dulbecco's modified Eagle's minimal essential medium with high
glucose (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10%
fetal bovine serum (FBS, Life Technologies™, Carlsbad, CA, USA).
LNCaP cells, clone FGC, an androgen-dependent human prostate cancer
cell line (RIKEN Cell Bank, Tsukuba, Japan), were cultured in
RPMI-1640 (Sigma-Aldrich) supplemented with 10% FBS. Cells were
incubated at 37°C in a humidified atmosphere containing 95% air and
5% CO2.
Docetaxel, paclitaxel, carboplatin, epirubicin, and
gemcitabine were purchased from Sawai Pharmaceutical (Osaka,
Japan). Doxorubicin was purchased from Kyowa Hakko Kirin (Tokyo,
Japan). Cisplatin and 5-fluorouracil were purchased from
Sigma-Aldrich. Human recombinant tumor necrosis factor-α (TNF-α)
was purchased from Wako Pure Chemicals (Osaka, Japan).
Establishment of Bcl-2-expressing stable
transfectants
OmicsLink™ Expression Clones for transcript variant
α of Bcl-2 (EX-H3307-M02) and control vector (EX-NEG-M02) were
purchased from GeneCopoeia (Germantown, MD, USA). DU145 cells were
transfected using a combination of Nupherin™ (Enzo Life Sciences,
Farmingdale, NY, USA) and Lipofectamine® 2000 (Life
Technologies) when the cells reached 70–80% confluence in 24-well
plates. First, 1 μg of plasmid DNA was mixed with 10 μg of Nupherin
in 150 μl of Opti-MEM® (Life Technologies) for 15 min
and then combined with 150 μl of Opti-MEM containing 1–4 μl of
Lipofectamine 2000 for another 40 min at room temperature. The
culture media were replaced with the transfection media, and the
cells were briefly centrifuged at 100 × g and incubated for 4 h.
The transfection media were then replaced with 1 ml of culture
media. After overnight culture, the cells were subcultured in F25
flasks with culture media containing 500 μg/ml of G418. The
transfected cells were then cloned by limiting dilution under G418
selection and were passaged many times (>20) to develop stable
cell lines.
Flow cytometry
Expression levels of Bcl-2 protein in transfectants
were examined by intracellular flow cytometry. Briefly, cells fixed
in 1.5% paraformaldehyde solution for 10 min were permeabilized by
BD FACS Permeabilizing Solution (BD Biosciences, San Jose, CA, USA)
for 10 min at room temperature. After washing with staining buffer
(phosphate-buffered saline containing 0.5% bovine serum albumin),
cells were stained with PE-conjugated anti-human Bcl-2 monoclonal
antibody (code no. 340576, BD Biosciences). Flow cytometric
analysis was performed using the Guava easyCyte™ 8HT system (Merck
Millipore, Darmstadt, Germany).
Chemosensitivity assay
Cells were seeded in 96-well plates at
2×103 cells/well and pre-cultured for 24 h. Cells were
treated with various concentrations of chemotherapeutic agents for
72 h. At the end of the culture period, the cells were trypsinized,
and absolute cell number was counted with Guava easyCyte™ 8HT.
Cell viability assay
Cell viability was evaluated using the [3-(4,
5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sul-fophenyl)-2H-tetrazolium,
inner salt (MTS)] (CellTiter 96® Aqueous One Solution
Cell Proliferation assay, Promega, Madison, WI, USA). Briefly,
cells were seeded in 96-well plates at 2×103 cells/well.
Twenty-four hours later, the cells were treated with the indicated
drugs. At the end of treatment, MTS solution was added to each
well, and the plates were incubated for 1–3 h. The plates were read
at a wavelength of 490 nm using a microplate reader (Infinite™
M1000, Tecan, Männedorf, Switzerland).
Fluorescence microscopy
Cells were seeded in 96-well plates at
2×103 cells/well after pre-culture for 24 h. Then, the
cells were treated with the indicated drugs. After treatment,
Hoechst 33342 (Dojindo, Kumamoto, Japan) and 7-amino-actinomycin D
(7-AAD, AnaSpec, Fremont, CA, USA) were added to the cells to a
final concentration of 5 and 2.5 μg/ml, respectively. Morphological
changes of nuclei and cell death were evaluated under fluorescence
microscopy after incubation at 37°C for 1 h.
Caspase-3/7 activity assay
The caspase-3 and -7 activities of cells were
measured using Amplite™ Fluorimetric Caspase-3/7 assay kits (AAT
Bioquest, Sunnyvale, CA, USA) according to the manufacturer's
protocol. Briefly, cells were seeded in 384-well plates at
2.5×103 cells/well after pre-culture for 24 h. After
treatment, caspase-3/7 assay solution was added, and the cells were
incubated for 1 h at room temperature. Cleavage substrate
fluorescence was measured by a microplate reader, Infinite M1000
(Ex/Em = 350/450 nm).
Xenograft mouse model
Male Balb/c nu/nu mice were obtained from CLEA Japan
(Tokyo, Japan). Bcl-2-overexpressing or control DU145 cells were
inoculated subcutaneously into the flank at 2×106 cells.
Docetaxel (12 mg/kg), cisplatin (5 mg/kg), or TNF-α (125 μg/kg) was
then administered intravenously once a week for 3 weeks, starting 3
weeks after inoculation. The tumor volume was estimated by using
the following equation: V = ab2/2, where a and b are the
tumor length and width, respectively. The present animal study was
approved by the Ethics Committee of Sawai Pharmaceuticals.
Statistical analysis
Statistical analysis was carried out using the
statistical program EXSUS (ver. 8.0.1, CAC EXICARE, Tokyo, Japan).
IC50 values (concentrations required to inhibit cell
viability by 50%) were calculated with a sigmoid
concentration-response model. The statistical significance of
differences between two groups was assessed with Student's t-test,
and Dunnett's test was used to compare multiple means with the
control group. The data for IC50 values and logarithmic
values of mean fluorescent intensity (MFI) of Bcl-2 expression
levels were fitted by linear regression analysis, and correlation
coefficients were calculated using Microsoft Excel 2010 (Seattle,
WA, USA).
Results
Varied sensitivity of Bcl-2-expressing
DU145 transfectants to anticancer drugs
Nine Bcl-2-expressing DU145 transfectants were
established (Table I). The
expression levels of Bcl-2 were measured by intracellular flow
cytometry. Although the expression levels varied among the nine
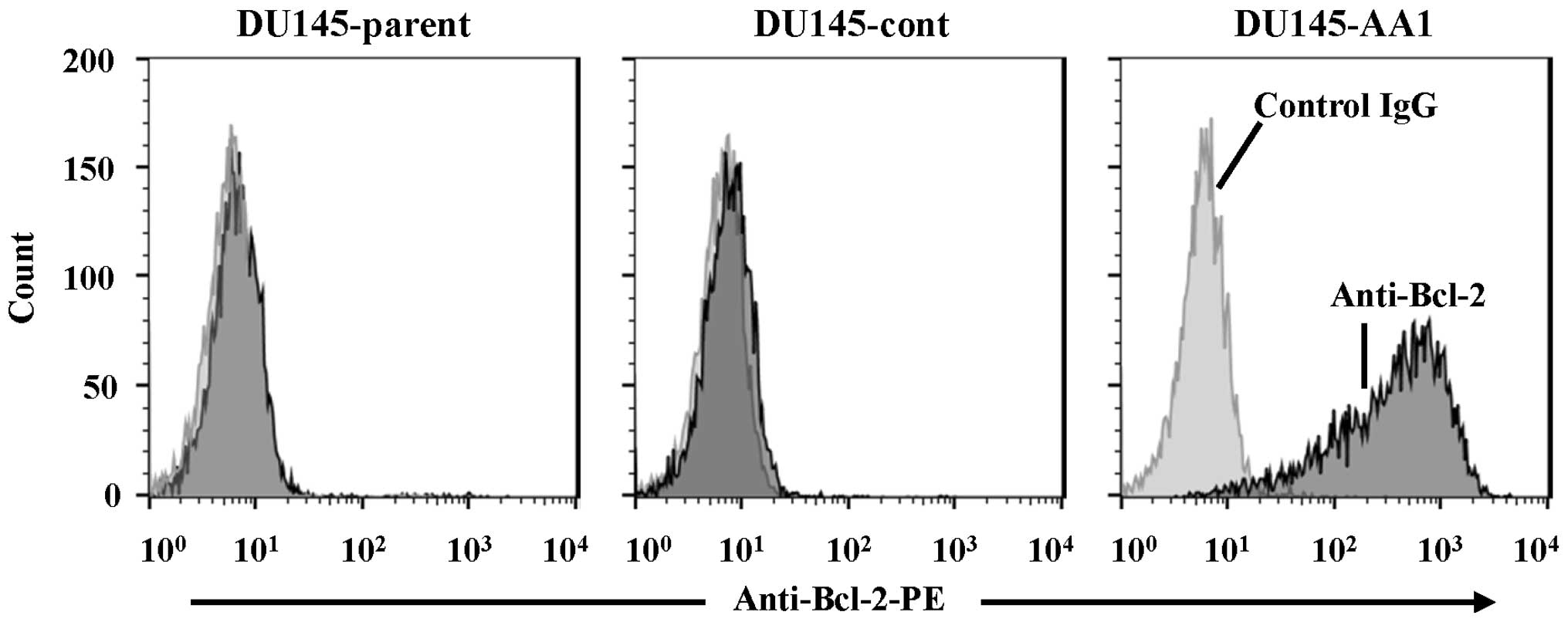
cell lines, line AA1 expressed Bcl-2 at the highest level.
Representative results of flow cytometry of parental DU145, control
DU145, and DU145-AA1 are shown in Fig.
1. Western blot analysis verified the expression of Bcl-2
protein as a molecular mass of 26 kDa, as predicted (data not
shown). Next, the sensitivities of these transfectants to a panel
of anticancer drugs were measured (Table I). The expression of Bcl-2 in DU145
cells did not correlate with the sensitivity to docetaxel or
paclitaxel. The Bcl-2-overexpressing AA1 line was more sensitive to
docetaxel than were control DU145, BA5, BB2, BD1, and G6 lines, all
of which expressed Bcl-2 at lower levels. As for the sensitivities
to the platinum-based anticancer agents cisplatin and carboplatin,
the relation was unique. Some Bcl-2 transfectants were more
sensitive to cisplatin and carboplatin than were the parental and
control lines. However, the Bcl-2 overexpressing lines, such as AA1
and AD2, were less sensitive. In addition, Bcl-2 overexpression
decreased their sensitivity to fluorouracil, but slightly increased
their sensitivity to anthracyclines such as epirubicin and
doxorubicin. Furthermore, the sensitivities of these transfectants
to TNF-α, which induces apoptosis via both intrinsic and extrinsic
pathways, were also analyzed. Because TNF-α alone did not induce
death in any cell line, TNF-α-induced activity was enhanced by the
addition of cycloheximide, a protein synthesis inhibitor. As a
result, the sensitivity of DU145 transfectants to TNF-α strongly
correlated with the Bcl-2 expression. Taken together, these results
indicate that although the sensitivity of DU145 transfectants to
TNF-α negatively correlated with Bcl-2 expression. A similar
tendency was not observed when the transfectants were treated with
docetaxel or other anticancer drugs.
 | Table ICytotoxic effects of chemotherapeutic
agents and TNF-α on parental, control, and Bcl-2-expressing DU145
transfectants. |
Table I
Cytotoxic effects of chemotherapeutic
agents and TNF-α on parental, control, and Bcl-2-expressing DU145
transfectants.
| |
IC50b (ng/ml) | |
|---|
| |
| |
|---|
| DU145 lines | Bcl-2
expressiona (MFI) | Docetaxel | Paclitaxel | Cisplatin | Carboplatin | Fluorouracil | Gemcitabine | Epirubicin | Doxorubicin | Viabilityc (%)TNF/CHX |
|---|
| Parent | 1.0 | 1.07 | 2.64 | 132.5 | 1,282 | 136.6 | 0.813 | 174.0 | 162.6 | 15.9 |
| Control | 0.3 | 0.80 | 2.72 | 146.8 | 1,604 | 137.7 | 0.588 | 169.4 | 167.6 | 13.0 |
| AA1 | 319.2 | 0.76 | 2.58 | 215.1 | 2,174 | 225.4 | 0.537 | 142.4 | 121.3 | 38.6 |
| AD2 | 173.4 | 0.77 | 2.15 | 197.2 | 2,274 | 146.5 | 0.766 | 140.5 | 137.5 | 47.0 |
| BA5 | 2.4 | 0.84 | 2.33 | 128.6 | 1,507 | 321.0 | 1.236 | 143.2 | 136.5 | 16.3 |
| BA6 | 184.1 | 0.88 | 2.34 | 11.4 | 169 | 2,229.2 | 1.203 | 94.7 | 96.9 | 50.9 |
| BB2 | 3.3 | 1.04 | 2.66 | 12.8 | 208 | 482.4 | 1.001 | 146.6 | 148.7 | 6.2 |
| BB6 | 0.8 | 0.65 | 1.95 | 75.6 | 963 | 774.2 | 0.621 | 133.9 | 129.4 | 7.0 |
| BD1 | 3.8 | 1.06 | 2.84 | 7.6 | 120 | 515.4 | 0.835 | 159.5 | 155.5 | 7.0 |
| DC4 | 96.0 | 0.70 | 2.34 | 77.5 | 955 | 798.0 | 0.675 | 121.3 | 116.3 | 42.9 |
| G6 | 3.0 | 1.21 | 3.15 | 163.6 | 1,192 | 267.3 | 0.531 | 152.0 | 158.4 | 27.6 |
| Correlation with
Bcl-2 expression | - | −0.232 | −0.236 | 0.186 | 0.213 | 0.336 | 0.088 | −0.625 | −0.727 | 0.882 |
Effects of Bcl-2 overexpression on in
vivo sensitivity to docetaxel, cisplatin, and TNF-α
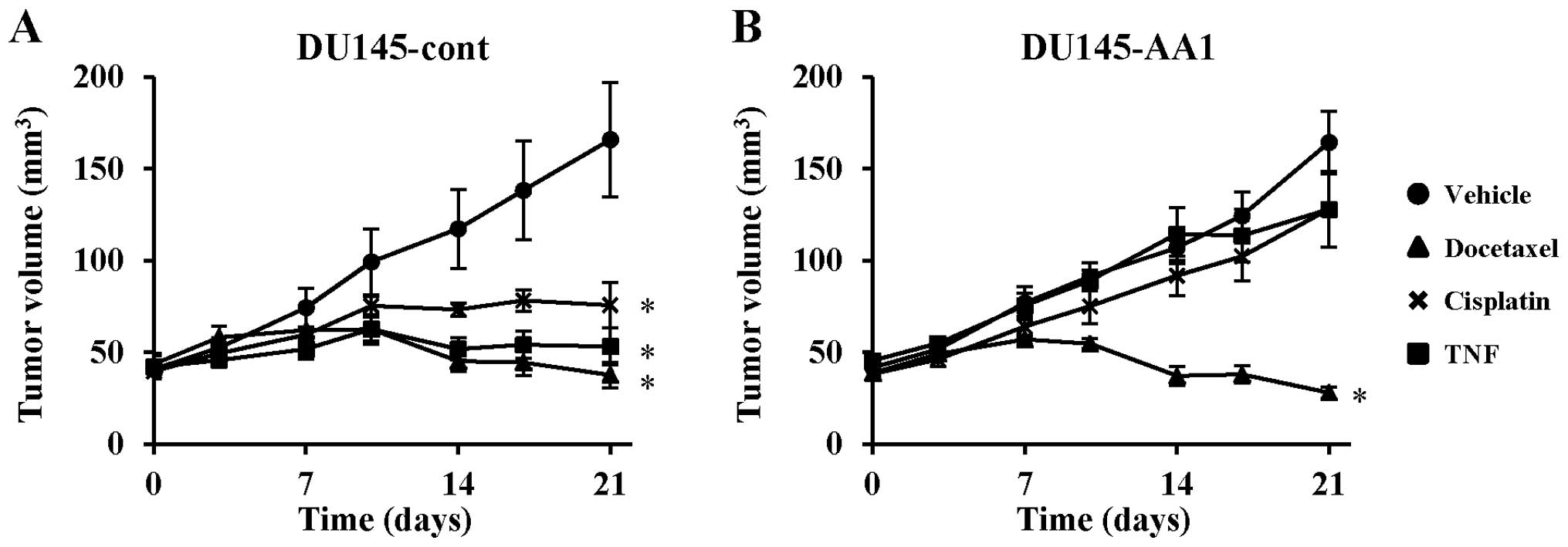
Next, we examined whether overexpression of Bcl-2
influenced the in vivo sensitivity of DU145 cells to
docetaxel, cisplatin, and TNF-α, using a xenograft mouse model.
DU145-cont and DU145-AA1, which showed the highest expression of
Bcl-2, were transplanted to nude mice, which were then treated with
the indicated drugs. In the vehicle-treated groups, there was no
difference in tumor growth between DU145-cont and DU145-AA1,
suggesting that the Bcl-2 expression level did not enhance the
in vivo growth of DU145 cells. Treatment with docetaxel,
cisplatin, or TNF-α significantly inhibited the growth of
DU145-cont cells as compared with the vehicle control (Fig. 2A). In contrast, docetaxel
suppressed the growth of DU145-AA1 cells, whereas neither cisplatin
nor TNF-α inhibited tumor growth (Fig.
2B). These results indicate that the Bcl-2 expression level
significantly influenced the in vivo sensitivity of DU145
cells to cisplatin and TNF-α, but was unrelated to the sensitivity
to docetaxel.
Cell death and morphological changes in
DU145 cells treated with docetaxel, cisplatin, or TNF-α
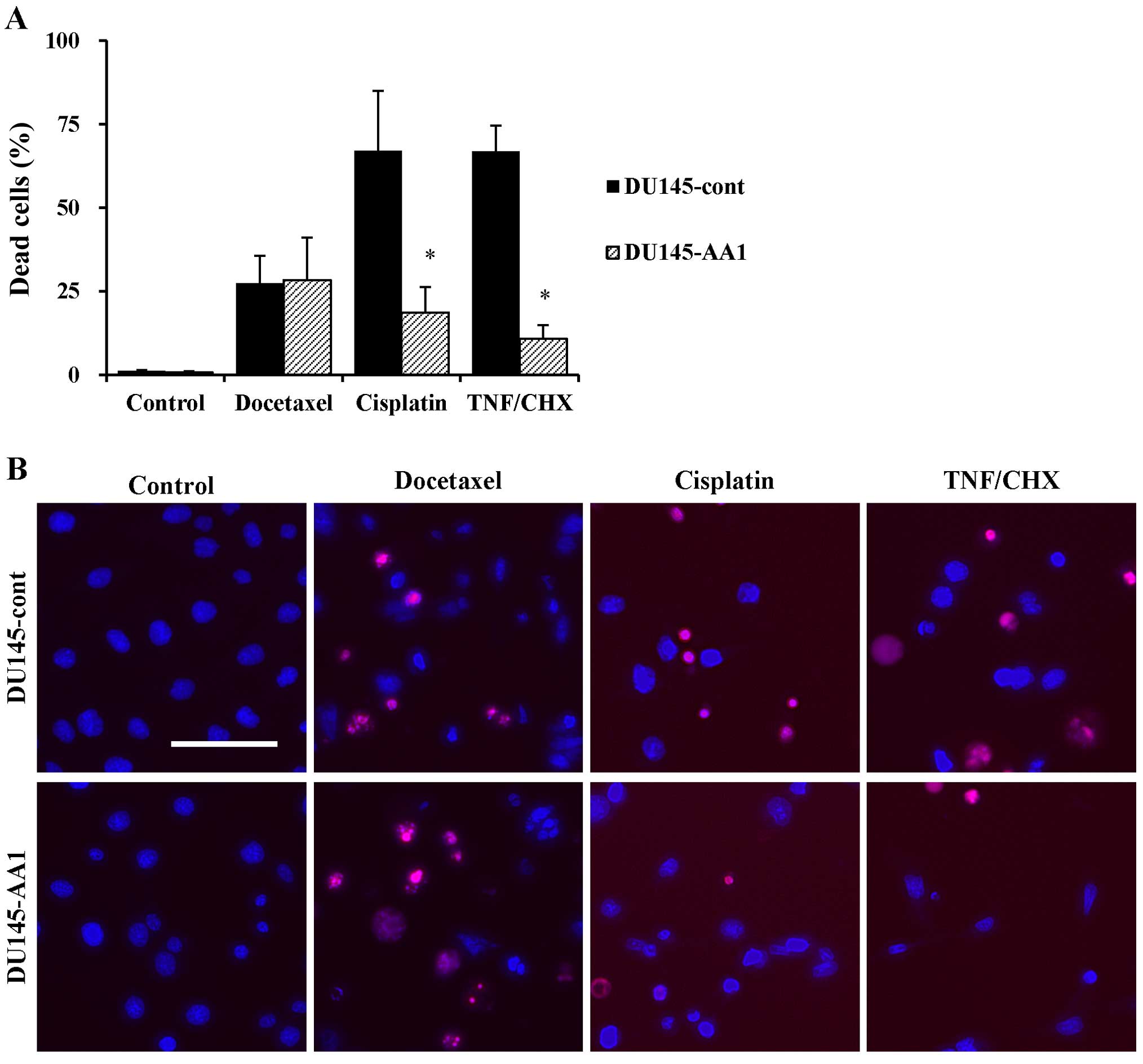
We next compared the sensitivity of DU145-cont and
DU145-AA1 cells to anticancer drugs by fluorescence microscopy. As
shown in Fig. 3A, although no
difference in their sensitivity to docetaxel was observed, the AA1
line was more resistant to cisplatin and TNF-α than was DU145-cont.
Morphological changes, such as nuclear shrinkage and condensation,
were observed in docetaxel-treated DU145-cont and DU145-AA1 cells
(Fig. 3B). DU145-cont cells
treated with cisplatin showed no typical apoptotic changes, but the
nucleus was partially condensed. These morphological changes were
not apparent in cisplatin-treated DU145-AA1 cells. In sharp
contrast, TNF-α strongly induced cell death and typical apoptotic
morphological changes of nuclei in DU145-cont cells, whereas
overexpression of Bcl-2 attenuated these changes. These results
indicate that overexpression of Bcl-2 had no effect on the
cytotoxicity of docetaxel in DU145 cells.
Caspase-3/7 activation in DU145 cells
treated with docetaxel, cisplatin, or TNF-α
It is well known that caspases play important roles
in apoptotic cell death. Therefore, we measured activities of
executioner caspase-3/7 in DU145 and LNCaP cells, the latter of
which are known to undergo apoptosis in response to docetaxel
(16). In the case of DU145-cont
cells, docetaxel and cisplatin slightly activated caspase-3/7,
whereas TNF-α rapidly activated caspase-3/7 (Fig. 4A). In contrast, no such activation
was observed in DU145-AA1 cells treated with docetaxel, cisplatin,
or TNF-α (Fig. 4B). In the case of
LNCaP cells, caspase-3/7 was rapidly and strongly activated by
either cisplatin or TNF-α, whereas the level of docetaxel-induced
activation of caspase-3/7 was low (Fig. 4C). These results indicate that
overexpression of Bcl-2 could inhibit TNF-α-induced caspase-3/7
activation in DU145 cells, whereas docetaxel could not trigger
caspase-3/7 activation.
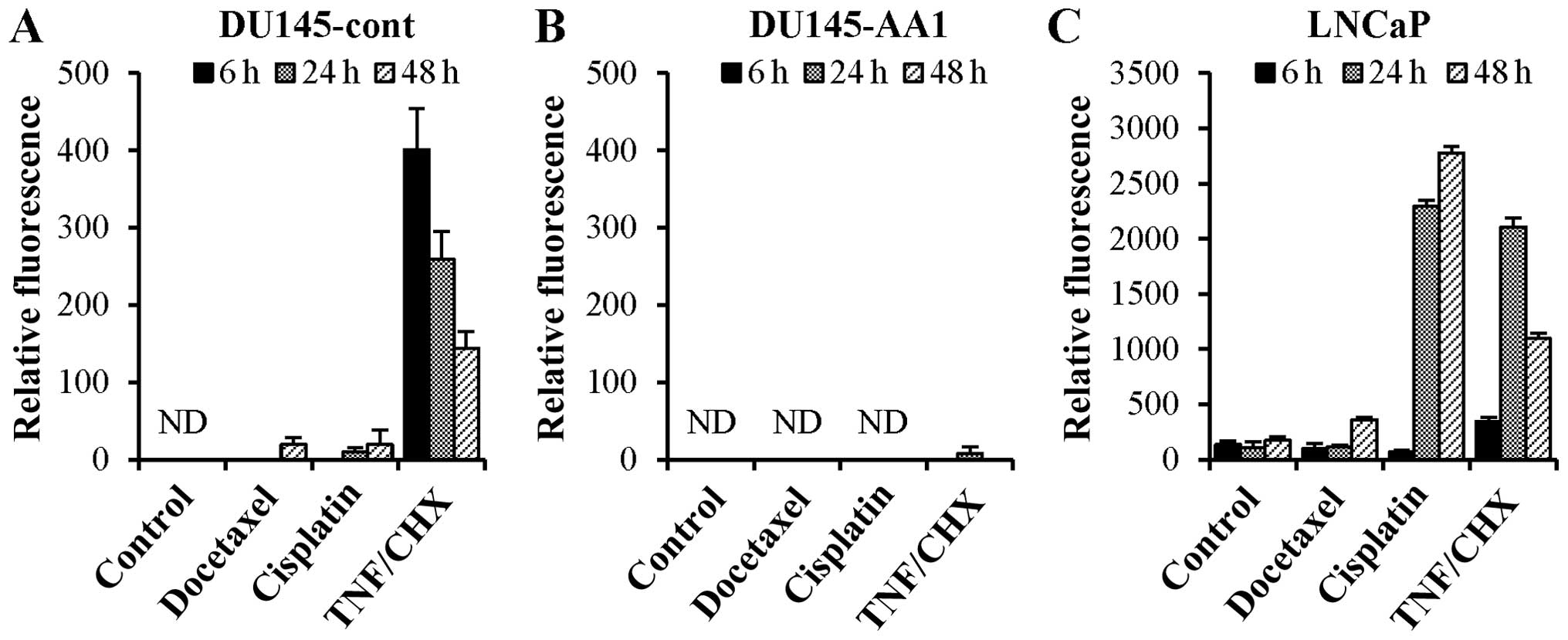 | Figure 4Caspase-3/7 activation in DU145-cont,
DU145-AA1, and LNCaP cells after treatment with docetaxel,
cisplatin, or TNF-α. DU145-cont (A), DU145-AA1 (B), and LNCaP (C)
cells were treated with docetaxel (4 ng/ml), cisplatin (4 μg/ml),
or TNF-α (10 ng/ml) and cycloheximide (10 μg/ml) (TNF/CHX) for 6,
24 and 48 h. Thereafter, the activation levels of caspase-3 and -7
were measured. Data (mean ± SD of triplicate determinations)
represent one of two separate experiments that had similar results.
ND, not detected. |
Roles of Bid and caspases in death of
DU145 cells
Bid mediates crosstalk between intrinsic and
extrinsic apoptotic pathways (19). Therefore, we determined whether
inhibition of Bid could rescue DU145 cells from death induced by
docetaxel, cisplatin, or TNF-α. The results showed that treatment
with BI6C9, a specific inhibitor of Bid, did not preserve the
viability of either DU145-cont or DU145-AA1 cells that were treated
with docetaxel, cisplatin, or TNF-α (Fig. 5A and B). In contrast, treatment
with BI6C9 significantly preserved the viability of LNCaP cells
treated with docetaxel, cisplatin, or TNF-α (Fig. 5C).
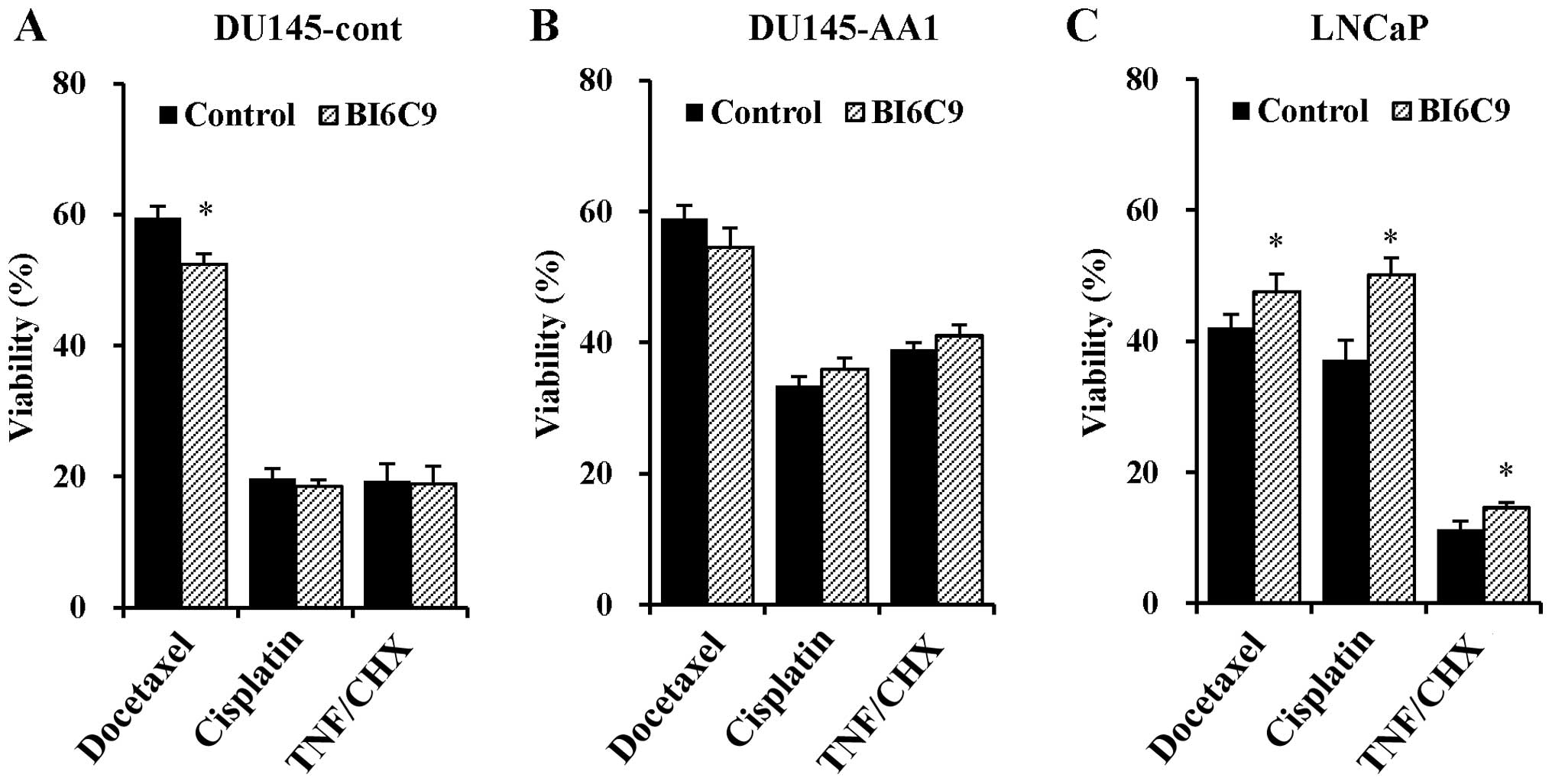 | Figure 5Effects of Bid inhibition on the
sensitivity of DU145-cont, DU145-AA1, and LNCaP cells to docetaxel,
cisplatin, or TNF-α. After pretreatment with BI6C9 (20 μM) for 30
min, DU145-cont (A), DU145-AA1 (B), and LNCaP (C) cells were
additionally treated with docetaxel (4 ng/ml) for 48 h, cisplatin
(4 μg/ml) for 48 h, or TNF-α (10 ng/ml) and cycloheximide (10
μg/ml) (TNF/CHX) for 24 h. Thereafter, the cell viability was
determined. Data are means ± SD of four independent experiments.
*P<0.05 by Student's t-test for the comparison
between with and without BI6C9 treatment. |
We further examined the contribution of caspase to
the death of docetaxel-treated DU145 cells, using a panel of
pro-apoptotic caspase inhibitors. Although none of the caspase
inhibitors preserved the viability of docetaxel- or
cisplatin-treated DU145-cont cells, all of the inhibitors
significantly preserved the viability of DU145-cont cells treated
with TNF-α (Fig. 6A). In the case
of DU145-AA1 cells, no such effect was observed (Fig. 6B). In contrast, only the
pan-caspase inhibitor preserved the viability of docetaxel-treated
LNCaP cells, and all inhibitors significantly preserved the
viability of LNCaP cells treated with cisplatin or TNF-α (Fig. 6C). Overall, these results indicate
that pro-apoptotic caspases were not involved in the death of
docetaxel-treated DU145 cells.
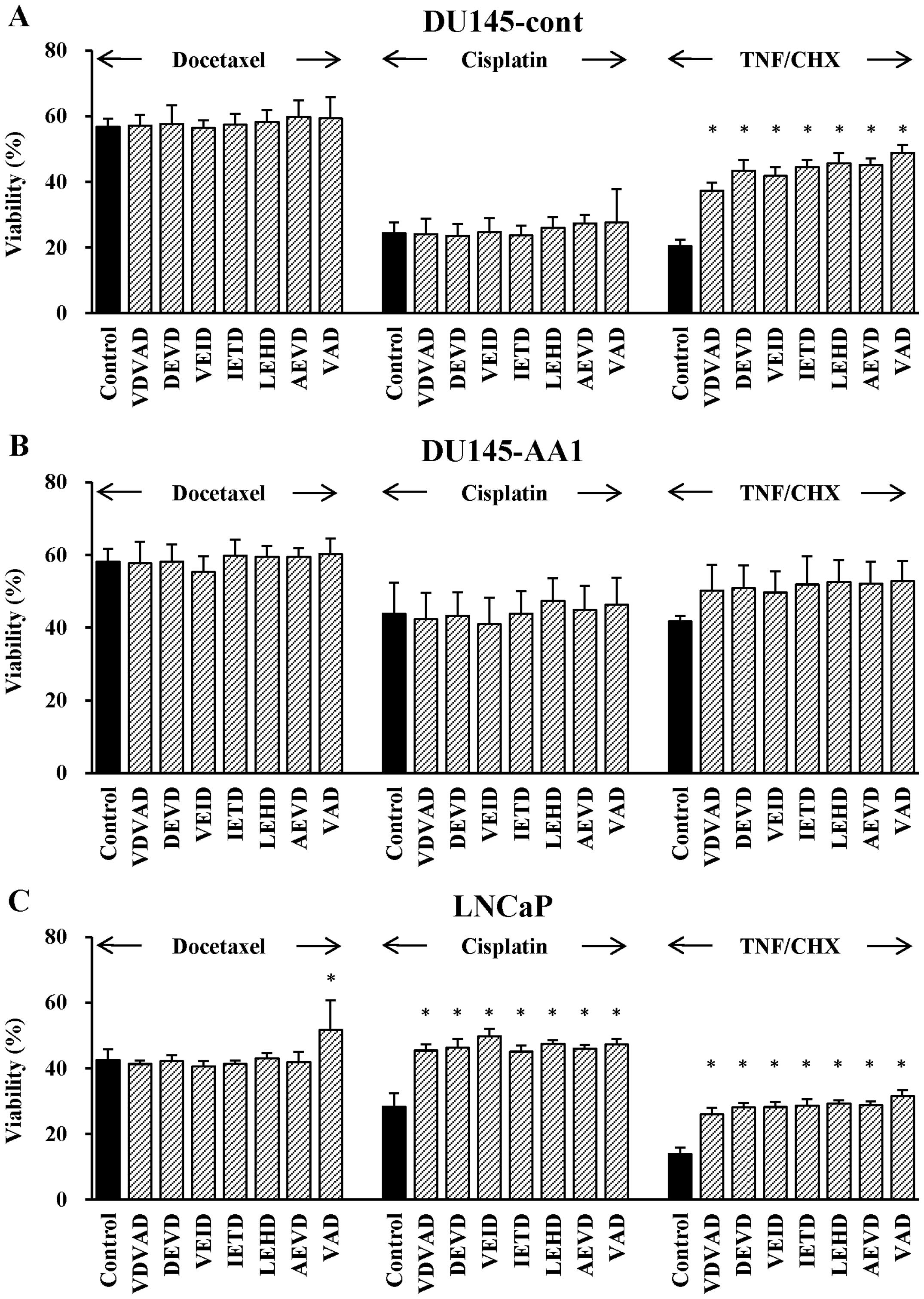 | Figure 6Effects of pro-apoptotic caspase
inhibitors on the sensitivity of DU145-cont, DU145-AA1, and LNCaP
cells to docetaxel, cisplatin, or TNF-α. After pretreatment with a
caspase inhibitor (20 μM) for 30 min, DU145-cont (A), DU145-AA1
(B), and LNCaP (C) cells were additionally treated with docetaxel
(4 ng/ml) for 48 h, cisplatin (4 μg/ml) for 48 h, or TNF-α (10
ng/ml) and cycloheximide (10 μg/ml) (TNF/CHX) for 24 h. The
following caspase inhibitors of fluoromethyl ketone
(FMK)-derivatized peptides were used: caspase-2, VDVAD;
caspase-3/7, DEVD; caspase-6, VEID; caspase-8, IETD; caspase-9,
LEHD; caspase-10, AEVD; and pan-caspase, VAD. Data are means ± SD
of four independent experiments. *P<0.05 by Dunnett's
test as compared with groups untreated with a caspase
inhibitor. |
Discussion
Although docetaxel has been the most useful
chemotherapeutic agent for patients with prostate cancer, recurrent
disease frequently becomes docetaxel-resistant. Therefore, in this
study we attempted to determine roles of the anti-apoptotic
molecule Bcl-2 in chemoresistance, using Bcl-2-expressing DU145
transfectants. Our results showed that Bcl-2 did not contribute to
the docetaxel-resistance of DU145 cells and that their
docetaxel-induced death is pro-apoptotic caspase-independent.
TNF-α treatment induced activation of caspase-3/7 in
control DU145 cells, whereas overexpression of Bcl-2 inhibited this
activation (Fig. 4), indicating
that TNF-α can induce caspase-dependent cell death in DU145 cells
and that Bcl-2 acts as an anti-apoptotic molecule in this process.
However, the relation between Bcl-2 expression and the
susceptibility of cancer cells to docetaxel remains controversial.
Previous studies have reported that abnormal Bcl-2 expression is
involved in malignant alteration and chemoresistance (24,27,28).
On the other hand, suppression of Bcl-2 gene expression by siRNA
has no effect on the anticancer activity of docetaxel in non-small
cell lung cancer cells (30).
Noguchi reported that clinical response of patients with breast
cancer to docetaxel is unrelated to the Bcl-2 expression level
(31). Therefore, we undertook
this study.
In our study, docetaxel-induced caspase-3/7
activation was lower in DU145 cells than in LNCaP cells (Fig. 4), as previously reported by Liu
et al (32). The underlying
mechanism can be explained by p53, which is a transcription factor
that responds to stress stimuli and induces pro-apoptotic Bcl-2
family members. DU145 cells and LNCaP cells carry mutated and
wild-type p53, respectively. P53 mutation results in inactivation
of caspases (33). In addition,
pro-apoptotic Bax expression is much lower in DU145 cells than in
LNCaP cells (34). Moreover, we
observed that Bid inhibition could not restore the
docetaxel-induced reduced viability of DU145 cells despite the fact
that such inhibition increased the viability of LNCaP cells that
were treated with docetaxel, cisplatin, or TNF-α (Fig. 5). Interestingly, a xenograft model
showed that overexpression of Bcl-2 exerted no effect on the in
vivo sensitivity of DU145 cells to docetaxel. On the basis of
these findings, we suppose that the crucial death pathway of
docetaxel-treated DU145 cells is independent of pro-apoptotic
caspases and p53. If so, what kinds of cell death were induced in
docetaxel-treated DU145 cells? Docetaxel induces activation of
cathepsins (35), which are
released from lysosome into the cytoplasm and subsequently induce a
cascade of intracellular events. Cathepsins mediate release of
cytochrome c from mitochondria and amplify the intrinsic
apoptosis cascade. Alternatively, cathepsins catalyze proteolytic
degradation of substrates that are crucial for cell survival,
leading to apoptosis (36).
Perhaps cathepsin-mediated apoptosis occurred in Bcl-2
overex-pressing DU145-AA1 cells. Furthermore, it has been shown
that cytotoxic agents can trigger key events contributing to cell
survival or death, including necroptosis (37), autophagy (38), endoplasmic reticulum stress
(39), and necrosis (40). Further studies are needed to
elucidate the precise mechanisms that mediate the death of
docetaxel-treated DU145 cells.
Interestingly, the sensitivities of control and
Bcl-2-expressing DU145 transfectants to chemotherapeutic agents
other than docetaxel varied. The sensitivity to cisplatin
fluctuated widely among the transfectants: some cell lines were
sensitized, whereas other cell lines with higher levels of Bcl-2
were desensitized. Downregulation of Bcl-2 is associated with
resistance to cisplatin in human small-cell lung cancer, but the
underlying mechanisms remain unclear (41). Previous studies have reported that
Bcl-2 transfection modulates the expression and activity of Bim, a
pro-apoptotic member (42). These
results suggest that the sensitivity of cancer cells to cisplatin
depends on the balance between the activities of anti- and
pro-apoptotic Bcl-2 family proteins. On the other hand, Bcl-2
transfection desensitized DU145 transfectants to fluorouracil, but
slightly sensitized them to anthracyclines such as epirubicin and
doxorubicin. It is very peculiar that the sensitivity to
fluorouracil was inversely related to that of anthracyclines
(correlation coefficient to epirubicin: −0.863 and doxorubicin:
−0.735). In addition, Bcl-2 transfection slightly delayed
proliferation (correlation coefficient: 0.314), and the relations
of the doubling time to the sensitivities to fluorouracil and to
anthracyclines were highly positive and moderately negative,
respectively (correlation coefficient to fluorouracil: 0.918,
epirubicin: −0.762 and doxorubicin: −0.690). These results suggest
that there may be a cross-correlation between the Bcl-2 expression
level and the growth rate with respect to the sensitivity to
fluorouracil and anthracyclines.
In conclusion, we investigated the effects of Bcl-2
expression in androgen-independent human prostate cancer DU145
cells on the susceptibility to docetaxel, as well as to cisplatin
and TNF-α. We also examined the effects of a panel of pro-apoptotic
caspase inhibitors on the sensitivity of DU145 cells to docetaxel.
Apoptosis is a hallmark of the anticancer activity of anticancer
agents, and caspase-3/7 and Bcl-2 family proteins play central
roles in the apoptosis pathway. However, our results indicate that
Bcl-2 expression does not necessarily contribute to the resistance
of DU145 cells to docetaxel and that the docetaxel-induced death of
DU145 cells is pro-apoptotic, and caspase-independent. Our data
also indicate that the expression level of Bcl-2, which is a potent
inhibitor of TNF-α-induced apoptosis, may have no influence on the
outcomes of patients with prostate cancer who receive
docetaxel.
Acknowledgements
We thank Mr. Peter Star of Medical Network K.K.
(Tokyo, Japan) for medical writing in the preparation of this
manuscript.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Ward E, Brawley O and Jemal A:
Cancer statistics, 2011: The impact of eliminating socioeconomic
and racial disparities on premature cancer deaths. CA Cancer J
Clin. 61:212–236. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Morgentaler A: Testosterone and prostate
cancer: An historical perspective on a modern myth. Eur Urol.
50:935–939. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Huggins C and Hodges CV: Studies on
prostatic cancer: I. The effect of castration, of estrogen and of
androgen injection on serum phosphatases in metastatic carcinoma of
the prostate. 1941. CA Cancer J Clin. 22:232–240. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Miyamoto H, Messing EM and Chang C:
Androgen deprivation therapy for prostate cancer: Current status
and future prospects. Prostate. 61:332–353. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hammerer P and Madersbacher S: Landmarks
in hormonal therapy for prostate cancer. BJU Int. 110(Suppl 1):
23–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tannock IF, de Wit R, Berry WR, Horti J,
Pluzanska A, Chi KN, Oudard S, Théodore C, James ND, Turesson I, et
al: Docetaxel plus prednisone or mitoxantrone plus prednisone for
advanced prostate cancer. N Engl J Med. 351:1502–1512. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Petrylak DP, Tangen CM, Hussain MHA, Lara
PNJ Jr, Jones JA, Taplin ME, Burch PA, Berry D, Moinpour C, Kohli
M, et al: Docetaxel and estramustine compared with mitoxantrone and
prednisone for advanced refractory prostate cancer. N Engl J Med.
351:1513–1520. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sinibaldi VJ: Docetaxel treatment in the
elderly patient with hormone refractory prostate cancer. Clin
Interv Aging. 2:555–560. 2007.
|
|
10
|
Merseburger AS, Bellmunt J, Jenkins C,
Parker C and Fitzpatrick JM; European Treatment Practices Group.
Perspectives on treatment of metastatic castration-resistant
prostate cancer. Oncologist. 18:558–567. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sweeney CJ and Chamberlain D: Insights
into E3805: The CHAARTED trial. Future Oncol. 11:897–899. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fulda S and Debatin K-M: Extrinsic versus
intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene.
25:4798–4811. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pienta KJ: Preclinical mechanisms of
action of docetaxel and docetaxel combinations in prostate cancer.
Semin Oncol. 28(Suppl 15): 3–7. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Stein CA: Mechanisms of action of taxanes
in prostate cancer. Semin Oncol. 26(Suppl 17): 3–7. 1999.PubMed/NCBI
|
|
15
|
Ganansia-Leymarie V, Bischoff P, Bergerat
J-P and Holl V: Signal transduction pathways of taxanes-induced
apoptosis. Curr Med Chem Anticancer Agents. 3:291–306. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Noguchi K, Shakuto S, Sakairi T and
Yoshida Y: Decrease in prostate specific antigen secretion
correlated with docetaxel-induced growth inhibition and apoptosis
in human prostate tumor cells. Gan To Kagaku Ryoho. 36:1863–1870.
2009.PubMed/NCBI
|
|
17
|
Green DR: Apoptotic pathways: Paper wraps
stone blunts scissors. Cell. 102:1–4. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kuwana T, Smith JJ, Muzio M, Dixit V,
Newmeyer DD and Kornbluth S: Apoptosis induction by caspase-8 is
amplified through the mitochondrial release of cytochrome c. J Biol
Chem. 273:16589–16594. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Green DR: Apoptotic pathways: The roads to
ruin. Cell. 94:695–698. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Scaffidi C, Fulda S, Srinivasan A, Friesen
C, Li F, Tomaselli KJ, Debatin KM, Krammer PH and Peter ME: Two
CD95 (APO-1/ Fas) signaling pathways. EMBO J. 17:1675–1687. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Luo X, Budihardjo I, Zou H, Slaughter C
and Wang X: Bid, a Bcl2 interacting protein, mediates cytochrome c
release from mitochondria in response to activation of cell surface
death receptors. Cell. 94:481–490. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li H, Zhu H, Xu CJ and Yuan J: Cleavage of
BID by caspase 8 mediates the mitochondrial damage in the Fas
pathway of apoptosis. Cell. 94:491–501. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gross A, McDonnell JM and Korsmeyer SJ:
BCL-2 family members and the mitochondria in apoptosis. Genes Dev.
13:1899–1911. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yip KW and Reed JC: Bcl-2 family proteins
and cancer. Oncogene. 27:6398–6406. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Johnson MI, Robinson MC, Marsh C, Robson
CN, Neal DE and Hamdy FC: Expression of Bcl-2, Bax, and p53 in
high-grade prostatic intraepithelial neoplasia and localized
prostate cancer: Relationship with apoptosis and proliferation.
Prostate. 37:223–229. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bubendorf L, Sauter G, Moch H, Jordan P,
Blöchlinger A, Gasser TC and Mihatsch MJ: Prognostic significance
of Bcl-2 in clinically localized prostate cancer. Am J Pathol.
148:1557–1565. 1996.PubMed/NCBI
|
|
27
|
Brown JM and Wilson G: Apoptosis genes and
resistance to cancer therapy: What does the experimental and
clinical data tell us? Cancer Biol Ther. 2:477–490. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Srivastava RK, Sasaki CY, Hardwick JM and
Longo DL: Bcl-2-mediated drug resistance: Inhibition of apoptosis
by blocking nuclear factor of activated T lymphocytes
(NFAT)-induced Fas ligand transcription. J Exp Med. 190:253–265.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Inoue Y, Gika M, Abiko T, Oyama T, Saitoh
Y, Yamazaki H, Nakamura M, Abe Y, Kawamura M and Kobayashi K: Bcl-2
overexpression enhances in vitro sensitivity against docetaxel in
non-small cell lung cancer. Oncol Rep. 13:259–264. 2005.PubMed/NCBI
|
|
30
|
Losert D, Pratscher B, Soutschek J, Geick
A, Vornlocher H-P, Müller M and Wacheck V: Bcl-2 downregulation
sensitizes nonsmall cell lung cancer cells to cisplatin, but not to
docetaxel. Anticancer Drugs. 18:755–761. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Noguchi S: Predictive factors for response
to docetaxel in human breast cancers. Cancer Sci. 97:813–820. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu C, Zhu Y, Lou W, Nadiminty N, Chen X,
Zhou Q, Shi XB, deVere White RW and Gao AC: Functional p53
determines docetaxel sensitivity in prostate cancer cells.
Prostate. 73:418–427. 2013. View Article : Google Scholar
|
|
33
|
Gurova KV, Rokhlin OW, Budanov AV,
Burdelya LG, Chumakov PM, Cohen MB and Gudkov AV: Cooperation of
two mutant p53 alleles contributes to Fas resistance of prostate
carcinoma cells. Cancer Res. 63:2905–2912. 2003.PubMed/NCBI
|
|
34
|
Tamaki H, Harashima N, Hiraki M, Arichi N,
Nishimura N, Shiina H, Naora K and Harada M: Bcl-2 family
inhibition sensitizes human prostate cancer cells to docetaxel and
promotes unexpected apoptosis under caspase-9 inhibition.
Oncotarget. 5:11399–11412. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mediavilla-Varela M, Pacheco FJ, Almaguel
F, Perez J, Sahakian E, Daniels TR, Leoh LS, Padilla A, Wall NR,
Lilly MB, et al: Docetaxel-induced prostate cancer cell death
involves concomitant activation of caspase and lysosomal pathways
and is attenuated by LEDGF/p75. Mol Cancer. 8:682009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chwieralski CE, Welte T and Bühling F:
Cathepsin-regulated apoptosis. Apoptosis. 11:143–149. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Delavallée L, Cabon L, Galán-Malo P,
Lorenzo HK and Susin SA: AIF-mediated caspase-independent
necroptosis: A new chance for targeted therapeutics. IUBMB Life.
63:221–232. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang ZJ, Chee CE, Huang S and Sinicrope
FA: The role of autophagy in cancer: Therapeutic implications. Mol
Cancer Ther. 10:1533–1541. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liao P-C, Tan S-K, Lieu C-H and Jung H-K:
Involvement of endoplasmic reticulum in paclitaxel-induced
apoptosis. J Cell Biochem. 104:1509–1523. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jang M-S, Lee S-J, Kang NS and Kim E:
Cooperative phosphorylation of FADD by Aur-A and Plk1 in response
to taxol triggers both apoptotic and necrotic cell death. Cancer
Res. 71:7207–7215. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kumar Biswas S, Huang J, Persaud S and
Basu A: Down-regulation of Bcl-2 is associated with cisplatin
resistance in human small cell lung cancer H69 cells. Mol Cancer
Ther. 3:327–334. 2004.PubMed/NCBI
|
|
42
|
Savry A, Carre M, Berges R, Rovini A,
Pobel I, Chacon C, Braguer D and Bourgarel-Rey V: Bcl-2-enhanced
efficacy of microtubule-targeting chemotherapy through Bim
overexpression: Implications for cancer treatment. Neoplasia.
15:49–60. 2013. View Article : Google Scholar : PubMed/NCBI
|