Introduction
Ovarian carcinoma is the gynaecologic malignancy
associated with the highest mortality in industrialised countries,
with a reported 5-year survival rate of <30% (1). The prognosis for patients with
ovarian cancer is determined by conventional factors such as
surgical stage and histological grade and type. Nevertheless, no
single molecular profile has helped identify the most aggressive
tumours or aided in the guidance of suitable therapeutic strategies
for specific patients. Due to the indefinite presentation of
ovarian cancer, it is often not diagnosed until it has reached
stage 3 or 4, at which point the 5-year survival rate drops
significantly; the mortality rate has remained unchanged for >30
years. Despite this, due to the lack of a known, reliable biomarker
for the disease, there is currently no ovarian cancer screening
programme used by the National Health Service. Many carcinomas
activate growth factor receptor signalling pathways that exhibit
genetic alterations involving either the receptor, or other factors
that drive the proliferation. Interestingly, several pathways
converge on the highly conserved serine/threonine kinase mTOR
(2).
The mTOR pathway is an integral hub of growth and
proliferation; gauging external energy, growth factor and stress
signals. It influences processes such as protein synthesis, lipid
biogenesis and autophagy to find the balance between anabolic and
catabolic mechanisms in relation to the whole-organism state. The
PI3K/Akt/mTOR pathway is activated in advanced stage disease and
inhibition of this pathway with inhibitors to Akt or its downstream
effector, mTOR, increases chemo-sensitivity to paclitaxel in
ovarian carcinoma cell lines (3).
Cell signalling by mTOR plays a critical role in protein synthesis
and proliferation of both normal and malignant cells. mTOR and one
of its substrates, S6 kinase, is activated in ovarian cancer cells
and inhibition of the mTOR pathways has anti-proliferative effects
(4). Rapamycin inhibits the growth
of a broad spectrum of malignancies including pancreatic cancer,
leukaemia and B-cell lymphoma (5).
Rapamycin (an mTOR inhibitor) may act as a substrate for the MDR
transporter P-glycoprotein and consequently limit its utility in
some tumours (6). In a transgenic
mouse model of ovarian cancer, treatment with the rapamycin
analogue everolimus (an inhibitor of mTOR) exhibited a delay in
cancer progression (7).
Deforolimus, another rapamycin analogue has been shown to inhibit
sarcoma and endometrial cancer growth both in vitro and
in vivo (8). In addition,
the rapalogue temsirolimus has exhibited therapeutic benefit when
administered to patients with clear cell carcinoma of the ovary
(9).
Moreover, a limitation to successful cancer
chemotherapy treatment is the acquisition of drug resistance. In
advanced-stage ovarian cancer, mTOR pathway is upregulated, and
inhibition of this pathway increases chemosensitivity in ovarian
carcinoma cell lines. Previous data from our laboratory has
revealed significant upregulation of DEPTOR in paclitaxel-resistant
(TaxR) SKOV-3TaxR and PEO1TaxR cell lines. SKOV-3TaxR exhibited
downregulation of RICTOR, RAPTOR and mTOR, whereas PEO1-TaxR showed
down-regulation of RAPTOR and upregulation of RICTOR and mTOR
(10).
In this study, we investigated the effects of
rapalogues on ovarian cancer using two cell lines (SKOV3 and
MDAH-2774) as in vitro experimental models. We expanded on
these observations by mapping the expression of mTOR components
(including DEPTOR, rictor, raptor and S6K) in tissue and peripheral
blood of ovarian cancer patients.
Materials and methods
Ovarian cancer clinical samples
Gene expression of mTOR, Deptor, Rictor and Raptor
were mapped in 12 clinical samples from ovarian cancer patients
using qPCR. Clinical samples were of ovarian origin and obtained
from patients at the 1st Department of Obstetrics and Gynecology,
‘Papageorgiou’ General Hospital, Medical School, Aristotle
University, Thessaloniki, Greece. Ethical permission was obtained
locally. The majority of ovarian cancers were deemed to be third
grade (10 out of 12) and at stage 3 (11 out of 12).
RNA isolation, cDNA synthesis and
quantitative RT-PCR
Ovarian tissue (40 mg) was lysed in a Qiagen Tissue
Lyser II (Qiagen, Hilden, Germany) for 2 min with a 3-mm stainless
steel ball bearing. RNA was extracted from tissue lysate using the
GenElute™ mRNA MiniPrep kit (Sigma-Aldrich, MO, USA), a silica
membrane/spin column method, and stored at −80°C until further
use.
cDNA was synthesised from mRNA using Superscript II
(Invitrogen, MA, USA). cDNA concentration was normalised using RNA
concentrations determined by NanoDrop (Thermo Scientific, MA, USA)
and was synthesised to a concentration of either 500 or 1,000
ng.
Primers
Relative expression of mTOR, DEPTOR, rictor and
raptor (Table I) were assessed by
quantitative PCR (Q-PCR) on an xxpress® (BJS
Biotechnologies, Middlesex, UK) thermal cycler using Kapa SYBR Fast
Universal Mastermix (Kapa Biosystems, MA, USA). According to MIQE
(minimum information for publication of quantitative real-time PCR
experiments) guidelines (11), an
assessment of the most stably expressed reference genes specific to
the samples used must be carried out prior to any qPCR experiment.
In light of this, a selection of 8 ovarian clinical samples were
assessed using the geNorm human 12 gene kit (Primer Design,
Southampton, UK) according to the manufacturer's instructions.
Reference gene expression stability was analysed using
qbaseplus software (Biogazelle, Zwijnaarde, Belgium).
Primers for mTOR, Deptor, Rictor and Raptor were used as previously
described (10). qPCR data were
analysed using the ΔCq method whereby the Cq
of the endogenous control was subtracted from the Cq of
the gene of interest and an RQ (relative quantity) value was
calculated by finding 2−ΔCq (11,12).
Where more than one reference gene was used, the RQ values were
averaged. A Student's t-test was used to calculate statistical
significance.
 | Table IThe primer sequences for the mTOR,
Deptor, Rictor and Raptor genes used in qPCR experiments for the
clinical samples and the in vitro experiments. |
Table I
The primer sequences for the mTOR,
Deptor, Rictor and Raptor genes used in qPCR experiments for the
clinical samples and the in vitro experiments.
| Name | Product length
(bases) | Strand | Size | Sequence (bases) |
|---|
| mTOR | 135 | Forward | 20 |
tgccaactaccttcggaacc |
| | Reverse | 20 |
gctcgcttcacctcaaattc |
| Deptor | 202 | Forward | 20 |
caccatgtgtgtgatgagca |
| | Reverse | 20 |
tgaaggtgcgctcatacttg |
| Rictor | 117 | Forward | 20 |
ggaagcctgttgatggtgat |
| | Reverse | 20 |
ggcagcctgttttatggtgt |
| Raptor | 170 | Forward | 20 |
actgatggagtccgaaatgc |
| | Reverse | 20 |
tcatccgatccttcatcctc |
Cell lines and treatments
SKOV3 (human ovarian clear cell adenocarcinoma) and
MDAH-2774 (human ovarian endometrioid adenocarcinoma) cell lines
were purchased from American Type Culture Collection (ATCC,
Rockville, MD, USA) and were used as models for epithelial ovarian
cancer. Both cell lines were cultured at 37°C/5% CO2 in
DMEM supplemented with 10% FBS, 1% penicillin/streptomycin and 1%
L-glutamine (all Gibco, MA, USA). SKOV3 and MDAH-2774 cells were
treated with specific mTOR inhibitors (Table II) at two concentrations for three
lengths of time, 24, 48 and 72 h.
| Table IIDetails of the mTOR pathway
inhibitory agents used in this study. |
Table II
Details of the mTOR pathway
inhibitory agents used in this study.
| Inhibitor | Mode of action | Concentrations |
|---|
| Rap | Allosteric mTOR
inhibitor | 20 nM | 100 nM |
| Eve | | 20 nM | 100 nM |
| Def | | 100 nM | 1,000 nM |
| Tem | | 10 nM | 100 nM |
| Res | Dual mTOR (via
DEPTOR) and PI3 kinase (via ATP binding site) inhibitor | 25 μM | 50 μM |
| BEZ | Dual mTOR and PI3
kinase inhibitor via ATP binding sites of both | 10 nM | 100 nM |
Western blotting
Proteins extracted from cultured cells following
treatments were assessed for cleaved and total caspase 9 and 3
levels, hallmarks of apoptosis, and phospho-p70S6K. Proteins were
separated by mass by SDS-PAGE in a 12.5% resolving gel. The
separated proteins were then electrophoretically transferred onto a
nitrocellulose membrane (Thermo Scientific) in wet-transfer buffer.
The membrane was probed using primary antibody for caspases 3 and 9
and phospho (Thr389) p70S6K (Cell Signaling Technology,
MA, USA) and an anti-rabbit HRP-conjugated secondary antibody
diluted in 5% bovine serum albumin/TBS Tween. The developed western
blots were analysed densitometrically using ImageJ software
(National Institutes of Health, MD, USA) and cleaved caspase 9 was
normalised internally against uncleaved caspase 9. The data were
analysed statistically using Student's t-test.
Ovarian tissue microarray
Unstained paraffin-embedded tissue micro-array
slides containing 70 ovarian cancers were obtained from US Biomax
(MD, USA). The paraffin-embedded slides were deparaffinised and
rehydrated by a series of washes in reducing concentrations of
ethanol (100, 95, 70 and 50%) followed by rinsing in tap water for
10 min. Antigen retrieval was accomplished by boiling slides in
sodium citrate (pH 6.0) for 20 min in a microwave. Slides were
washed in 0.4% of PBS Tween for 5 min and then incubated for 15 min
in PBS containing 0.3% H2O2 to eliminate
endogenous peroxidase activity. Blocking was carried out with 5%
goat serum, followed by 48-h incubation with primary antibody
(p70S6K). After several washes with PBS, slides were incubated with
HRP conjugated secondary antibody for 60 min. Further washing in
PBS Tween was carried out for 20 min before performing staining.
Slides were then subjected to DAB staining, counterstained with
haematoxylin and washed with 0.1% sodium bicarbonate. Slides were
analysed for immunoreactivity of p70S6K by a light microscope and
positive results were measured by the percentage of positive tumour
cells.
Proliferation assay
Proliferation, death and viability of the cells were
assessed after treatment using a Countess Automated Cell Counter
(Invitrogen) and trypan blue stain. Media was aspirated from the
cells which were then incubated with 200 μl of trypLE™ Express
(Thermo Scientific) per well and manually agitated to detach the
cells. The cells were resuspended in 800 μl of appropriate media to
make 1 ml of cell suspension in total. An equal volume of cell
suspension was mixed thoroughly with trypan blue stain (0.4%,
trypan blue is selectively absorbed by dead cells) and applied to a
Countess™ cell counting chamber slide (Invitrogen). Three readings
were taken per sample and an average value calculated.
Wound healing assay
Cells were assessed for their ability to close an
artificially created gap in cell growth area. One confluent T-75
cell culture flask was seeded per three 6-well plates with 2 ml
appropriate media per well and grown until there was a confluent
monolayer. A 20-μl pipette tip was used to create a ‘scratch’ in
growth area perpendicular to a line drawn on the underside of the
well in marker pen. The media was then aspirated and replaced with
treated media. Cells were placed under a Zeiss Axiovert 200M
microscope and images were recorded at 0, 6, 12 and 18 h. The
perpendicular line of marker was used as a landmark to ensure that
an image of the same area was taken at each time-point.
ImageStream
Protein expression and localisation of mTOR, DEPTOR,
rictor and raptor was investigated using ImageStream high
resolution flow cytometry (Merck Millipore, Darmstadt, Germany).
Media was aspirated from the cells which were then incubated with
TrypLE™ Express (Invitrogen). Washes in PBS were performed to
remove debris. The cells were resuspended and incubated in 4%
paraformaldehyde (PFA, Sigma, MO, USA) for 7 min and the PFA
removed. Further washed in PBS were performed and stored ice-cold
at −20°C until further use. The cells were incubated in blocking
buffer (5% bovine serum in PBS) for 30 min followed by the
appropriate primary antibody (diluted in blocking buffer) overnight
at 4°C. Washes were performed with PBS and incubated in secondary
antibody (diluted in blocking buffer) for 30 min. After secondary
antibody incubation the cells were washed again in PBS to remove
any remaining antibody. PBS was removed and the cells were
resuspended in 100 μl Accumax (Innovative Cell Technologies, CA,
USA) to dissociate any cellular aggregates. Draq5 nuclear stain (1
μl) was added before visualisation on ImageStream.
Statistical analysis
Changes observed in experiments were assessed for
statistical significance using the Student's t-test. An assessment
for homoscedasticity (variance) of data from each category was made
using the F-test. If homoscedasticity was proven, an unpaired,
two-tailed Student's t-test was performed to assess significance in
all cases as no matched pairs of samples were used. If data were
not homoscedastic, an unpaired, two-tailed Student's t-test with
Welch's correction was performed to account for variance. All
statistical tests were performed using GraphPad Prism®
software (GraphPad Software Inc., CA, USA). P-values were denoted
on graphs if significant (p<0.05).
Results
Expression of mTOR components in ovarian
cancer cell lines
Prior to studying the effects of mTOR inhibitors, we
assessed the basal expression of mTORC1 and mTORC2 components in
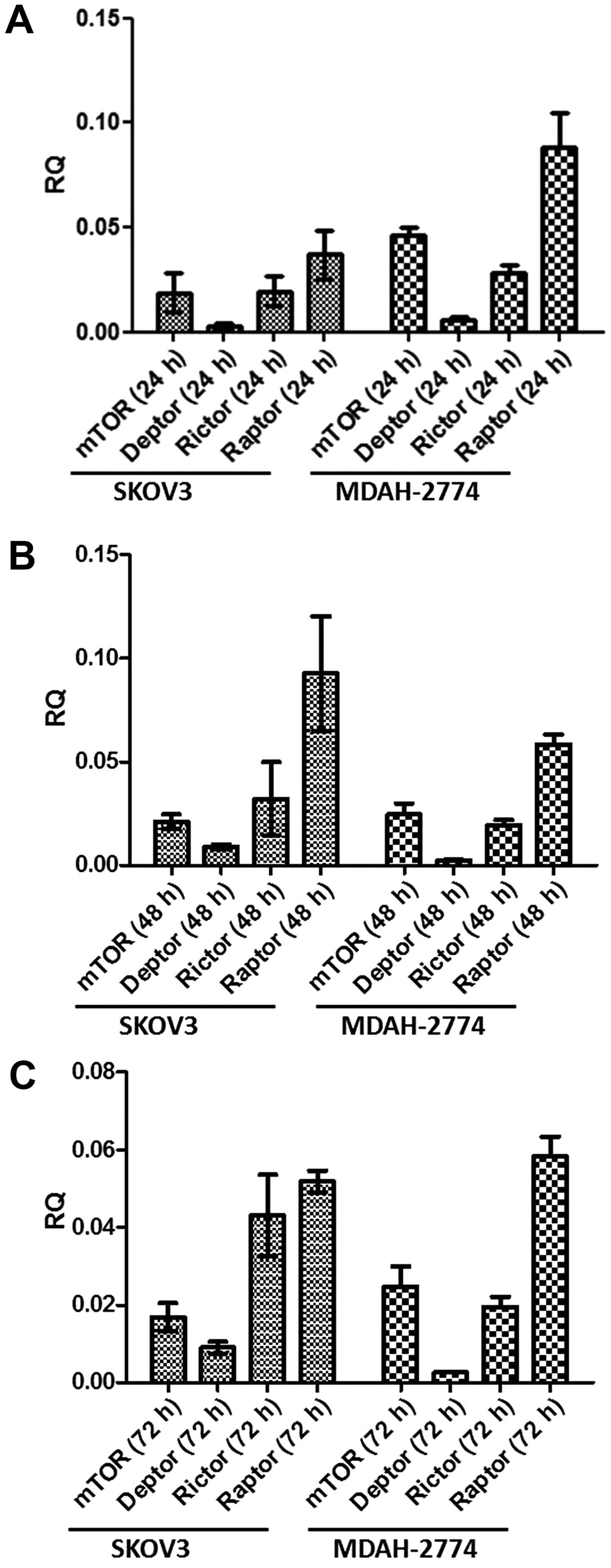
both cell lines over 72 h. All key components, i.e., mTOR, DEPTOR,
rictor and raptor are expressed in both cell lines in a similar
fashion over the course of 72 h (Fig.
1). The predominant component was raptor, with DEPTOR
demonstrating minimal expression in both cell lines under basal
conditions. Following qPCR analysis, we used the state-of-the-art
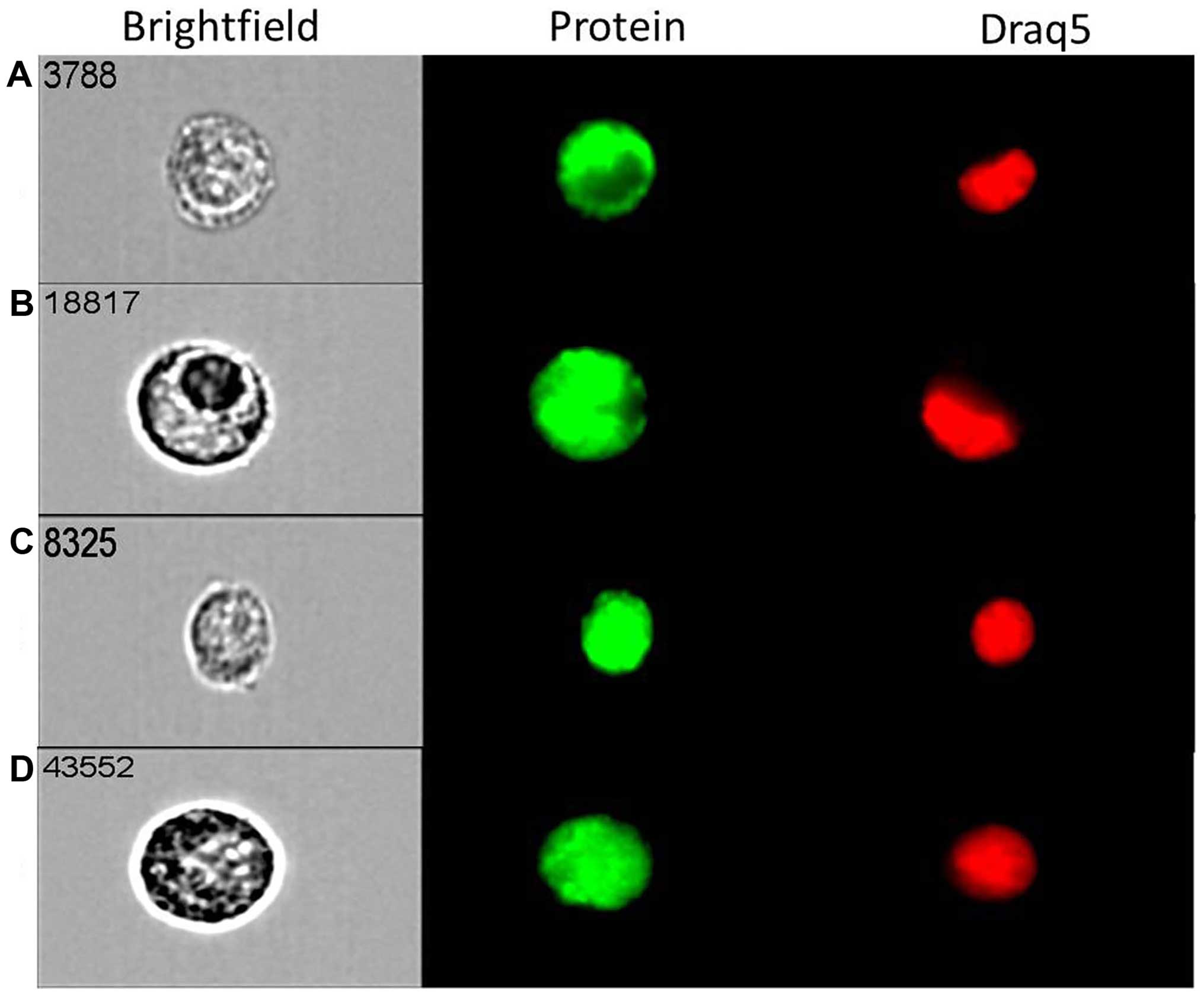
ImageStream technology that combines flow cytometry with cell
imaging for parallel quantification and visualisation of single
cells. Strong cytoplasmic and nuclear expression for both mTOR
(Fig. 2A) and DEPTOR (Fig. 2B) was evident in most of ovarian
cancer (MDAH-2774) single cells studied. The expression of rictor
(Fig. 2C) and raptor (Fig. 2D) was primarily
nuclear/cytoplasmic. Similar cellular distribution of these key
components of both mTORC1 and mTORC2 complexes was evident in SKOV3
cells (data not shown).
mTOR inhibitors compromise migratory
capacity and cell proliferation of ovarian cancer cells in
vitro
Wound healing assays can give a visual indication of
cell proliferative and migratory capacity spatially in
vitro. Rapamycin (20 and 100 nM) was selected from the
rapalogues (since they have a similar chemical structure) and
resveratrol (25 and 50 μM) and BEZ (10 and 100 nM) were used as
they are mechanistically different to the rapalogues and one
another. SKOV3 cells showed a marked growth into the wound which
was not shared by any of the inhibitor-treated cells (Fig. 3). In MDAH-2774 cells, a more modest
response was recorded using the same inhibitors (data not
shown).
Based on the wound healing data, we have decided to
expand on the repertoire of rapalogues; therefore, trypan blue
staining was used to make an assessment of the proliferative
capacity of SKOV3 and MDAH-2774 cells up to 72 h following
treatment with the following inhibitors: rapamycin (Rap, 20 and 100
nM), everolimus (Eve, 20 and 100 nM), deforolimus (Def, 100 and
1,000 nM), temsirolimus (Tem, 10 and 100 nM), resveratrol (Res, 25
and 50 μM), and NVP BEZ-235 (BEZ, 10 and 100 nM). Treatment of the
SKOV3 cell line with Rap and the rapalogues showed a
non-dose-dependent decrease in viable cells (Fig. 4, Rap 100 nM: p=0.0355 at 72 h, Eve
100 nM: p=0.0047 at 72 h, Tem 100 nM: p=0.0446 at 72 h, Def 1,000
nM did not reach statistical significance: p=0.0597). The decrease
in viable cells shown by Res and BEZ was dose-dependent (Fig. 4, Res 50 μM: p<0.0001 at 72 h,
BEZ 100 nM: p<0.0001 at 72 h).
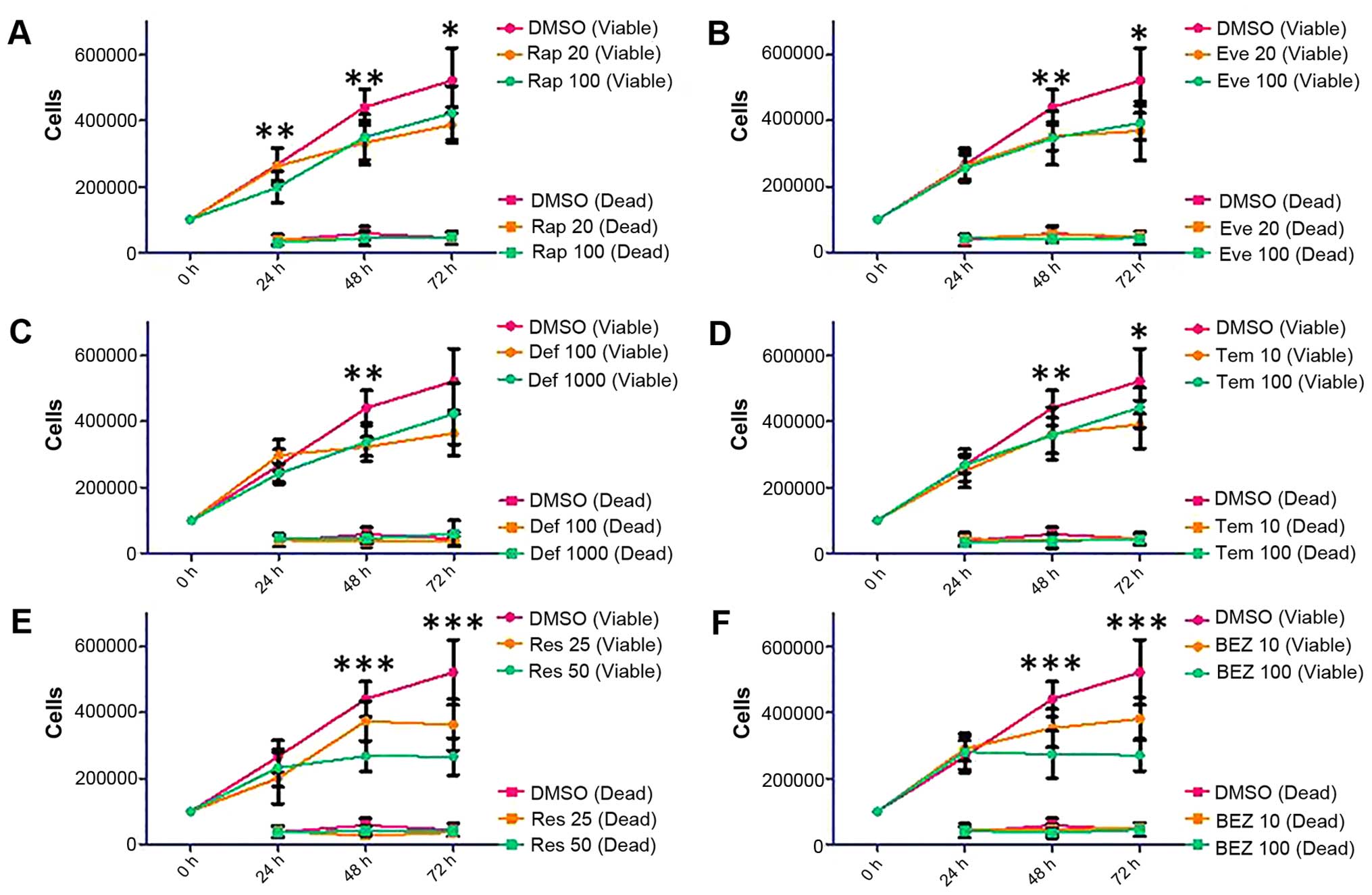 | Figure 4Proliferation and survival analysis of
SKOV3 cells following treatment with Rap [(A) 20 and 100 nM],
everolimus [(B) Eve, 20 and 100 nM], deforolimus [(C) Def, 100 and
1,000 nM], temsirolimus [(D) Tem, 10 and 100 nM], Res [(E) 25 and
50 μM] and BEZ [(F) 10 and 100 nM]. SKOV3 cells were seeded in
6-well plates at a density of 100,000 cells per well and allowed to
proliferate for 24 h before treatment. Cells were treated with mTOR
pathway inhibitors for 24, 48 and 72 h. At each time-point, cells
were detached and mixed with an equal volume of trypan blue. A
count of viable and dead cells was made using a Countess automated
cell counter (Invitrogen). Error bars depict standard deviation,
*p=0.01–0.05, **p=0.001–0.009,
***p<0.009. |
Viable cell count in MDAH-2774 cells was
differentially affected by Rap (Rap 100 nM: p=0.0013 at 48 h),
rapalogues (Def 1,000 nM: p=0.0004 at 48 h, Tem 100 nM: p=0.0065 at
72 h) or Res (Res 50 μM: p=0.0014 at 72 h) treatment (Fig. 5). BEZ treatment at 10 nM
concentration did not have an effect on viable cell count, but at
100 nM, proliferation was almost entirely eliminated (Fig. 5, p<0.0001 at 48 and 72 h).
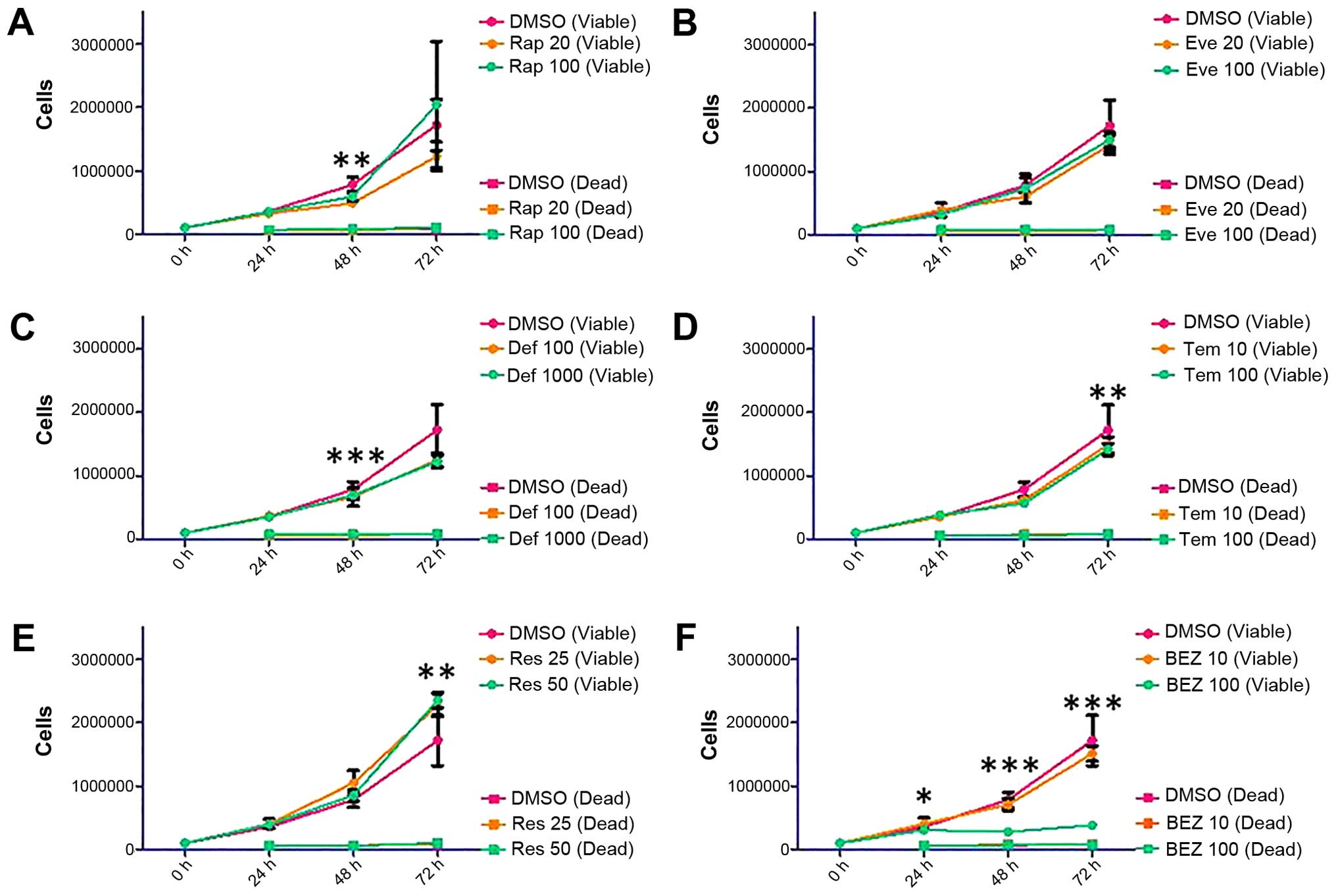 | Figure 5Proliferation and survival analysis of
MDAH-2774 cells following treatment with Rap [(A) 20 and 100 nM],
everolimus [(B) Eve, 20 and 100 nM], deforolimus [(C) Def, 100 and
1,000 nM], temsirolimus [(D Tem, 10 and 100 nM), Res [(E) 25 and 50
μM] and BEZ [(F) 10 and 100 nM)]. MDAH-2774 cells were seeded in
6-well plates at a density of 100,000 cells per well and allowed to
proliferate for 24 h before treatment. Cells were treated with mTOR
pathway inhibitors for 24, 48 and 72 h. At each time-point, cells
were detached and mixed with an equal volume of trypan blue. A
count of viable and dead cells was made using a Countess automated
cell counter (Invitrogen). Error bars depict standard deviation,
*p=0.01–0.05, **p=0.001–0.009,
***p<0.009. |
Effects of mTOR inhibitors on caspase 9
and phospho-p70S6K
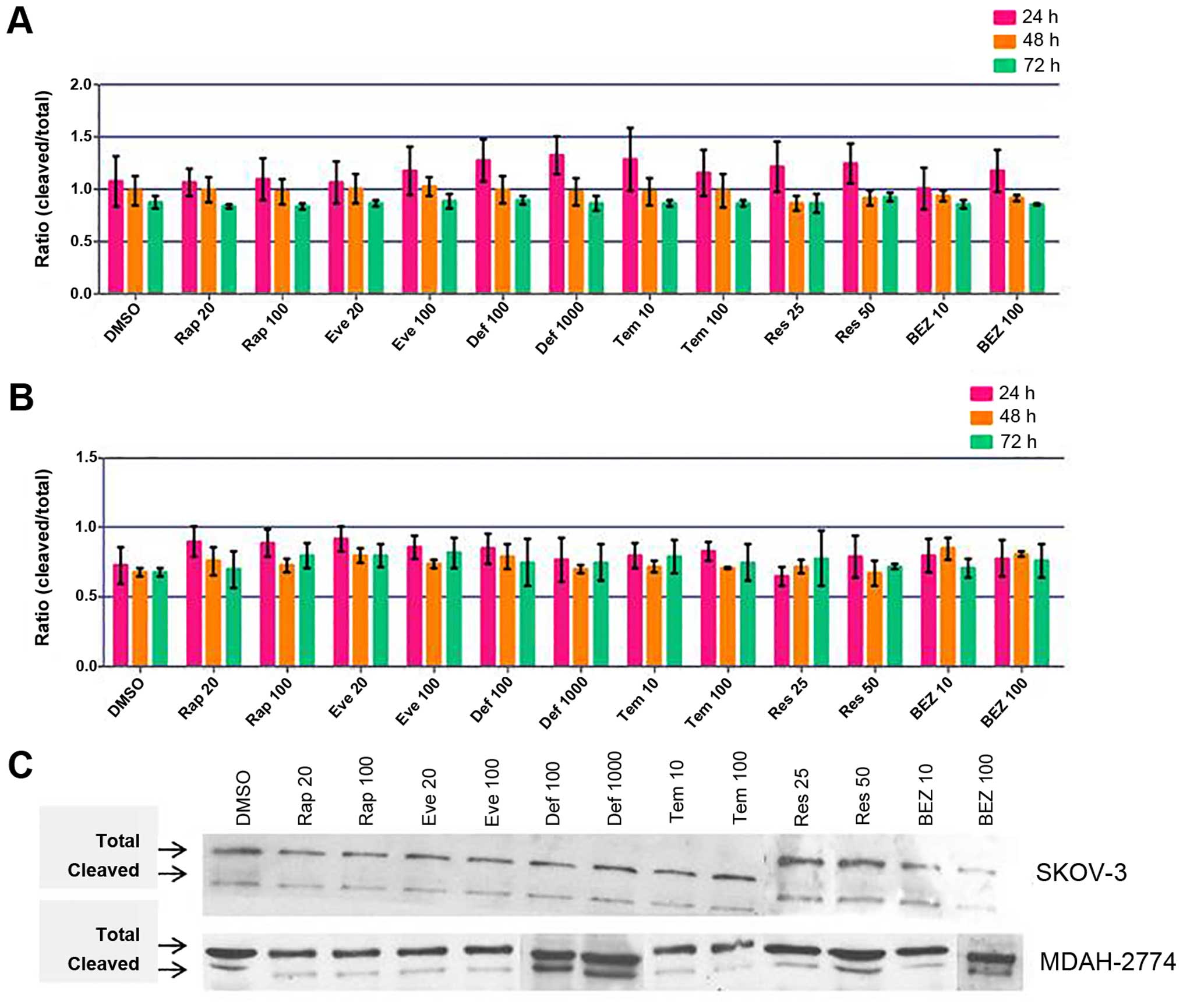
Cleavage of caspase 9 could be seen after all
treatments and time-points in both cell lines. When analysed
densitometrically, SKOV3 cells showed a clear trend of highest
caspase 9 cleavage at 24 h and lessening over time in each case
(Fig. 6A). These changes did not
reach statistical significance in comparison to controls; however,
significant changes were found between time-points in Rap 20 nM
(p=0.0069, 24–72 h), Rap 100 nM (p=0.00159, 24–72 h), Def 1,000 nM
(p=0.0496, 24–48 h), Res 25 μM (p=0.0367, 24–48 h and p=0.0353,
24–72 h) and Res 50 μM (p=0.0483, 24–72 h) treatments, indicating
that cleavage of caspase 9 does decrease significantly over time in
these cases. However, MDAH-2774 cells showed no increase in caspase
9 cleavage upon treatment with mTOR inhibitors (Fig. 6B and C).
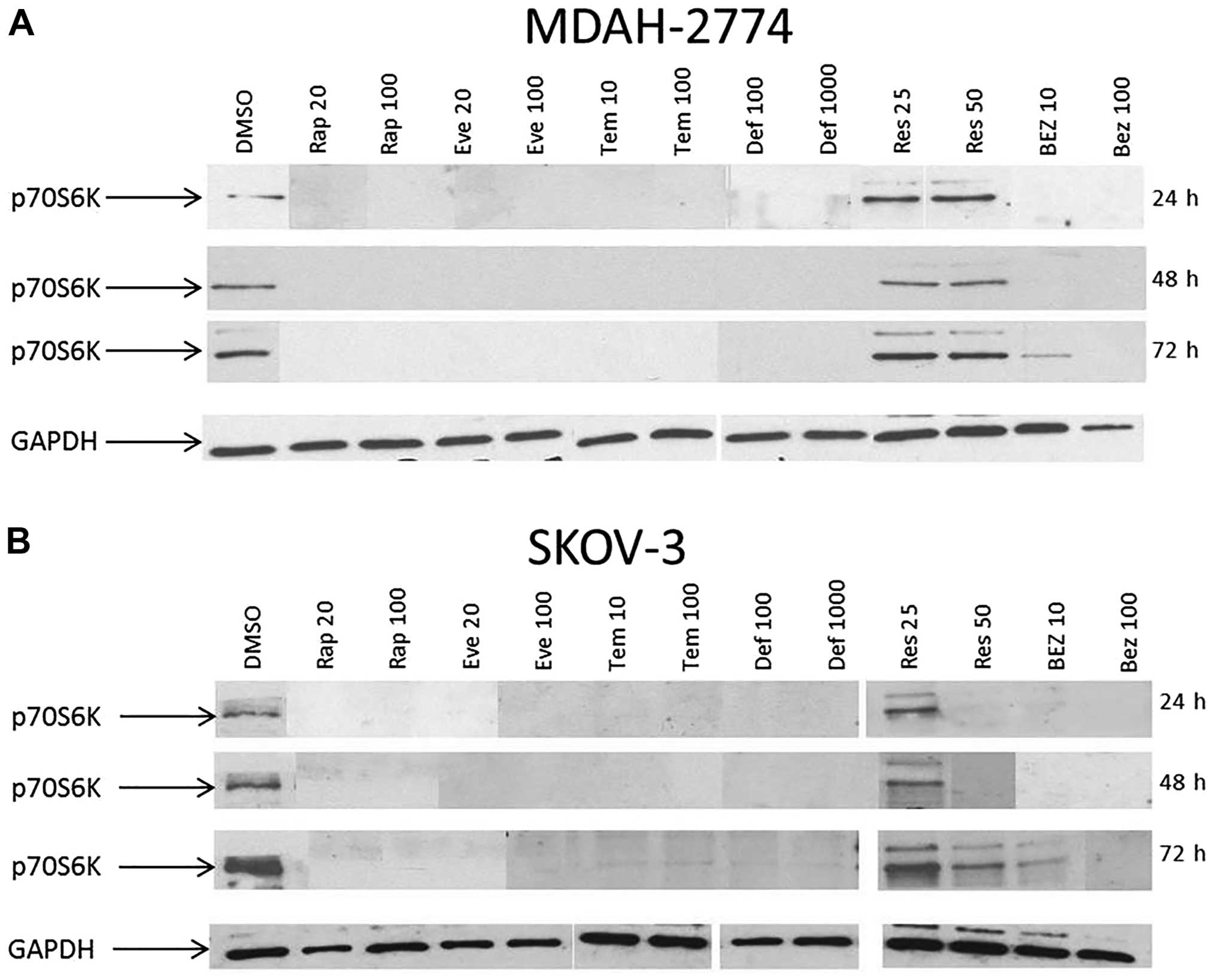
Phosphorylation status of p70S6K was examined to
better understand the activity of the mTOR pathway after
inhibition. At 24, 48 and 72 h, the basal phosphorylation of p70S6K
was completely abolished in both MDAH-2774 and SKOV3 (Fig. 7) cells treated with rapamycin, all
of the other rapalogues and at 100 nM of BEZ treatment. In SKOV3
cells treated with Res, dephosphorylation of p70S6K only occurred
at 50 μM of Res at 24 and 48 h. Res treatment did not
dephosphorylate p70S6K at 25 or 50 μM in MDAH-2774 cells.
Expression of S6K in ovarian tissue
array
Based on the previous data on p70S6K, it is evident
that the mTORC1 activity is compromised upon treatment with mTOR
inhibitors. Given the significance that p70S6K has in moderating
the activity of mTORC1, we studied the cellular distribution of
this kinase on an ovarian tissue array. Ovarian cancer (n=70)
tissue samples were probed with a p70S6K antibody by
immunohistochemistry using DAB staining. Three areas of each tissue
sample were selected at random and total cells and total positive
cells were counted in each area. An average percentage of positive
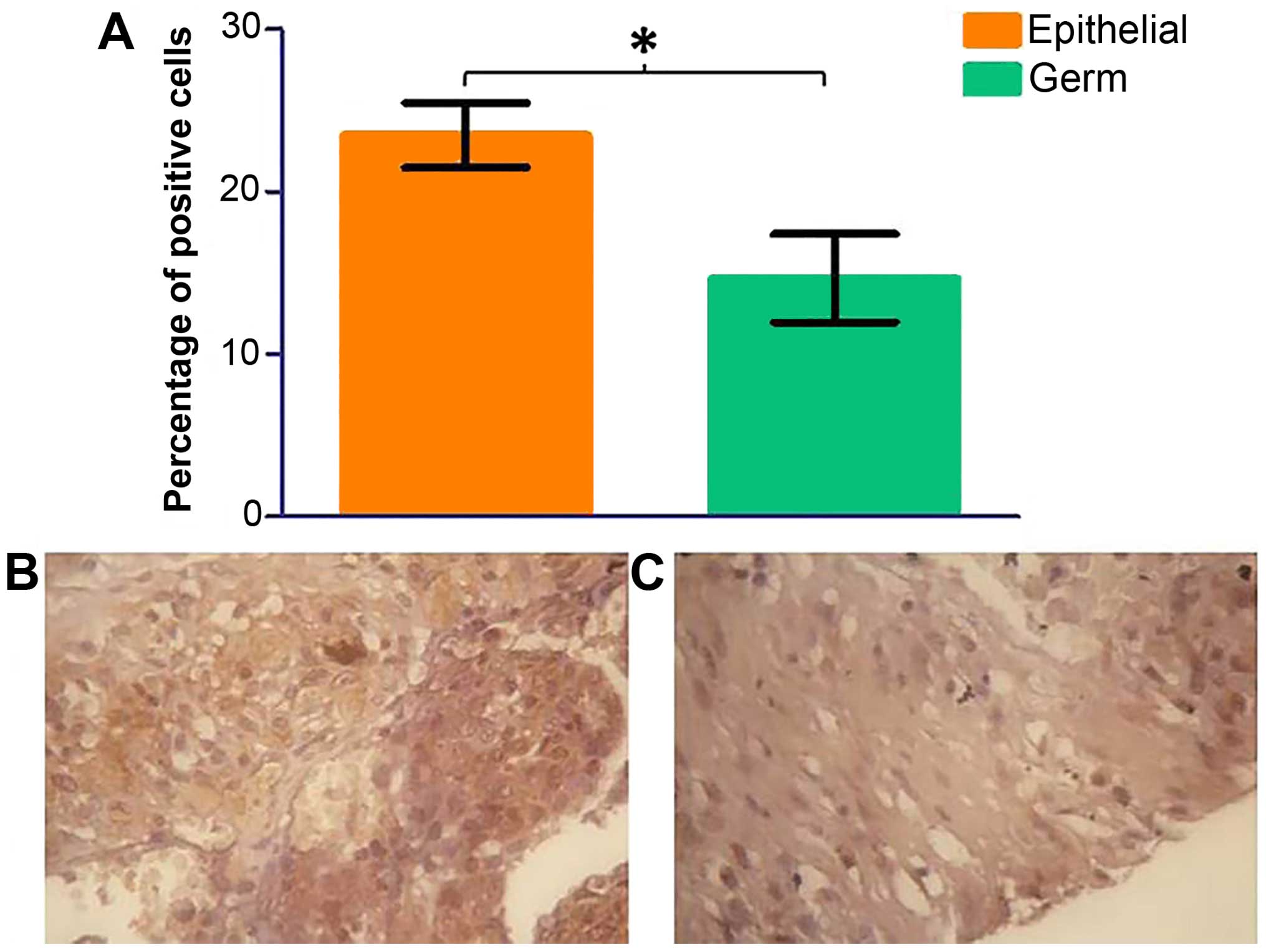
cells was calculated. There was a significant decrease in
phospho-p70S6K protein expression in germ cell tumours in
comparison to epithelial cell tumours (Fig. 8, p=0.0398). No significant change
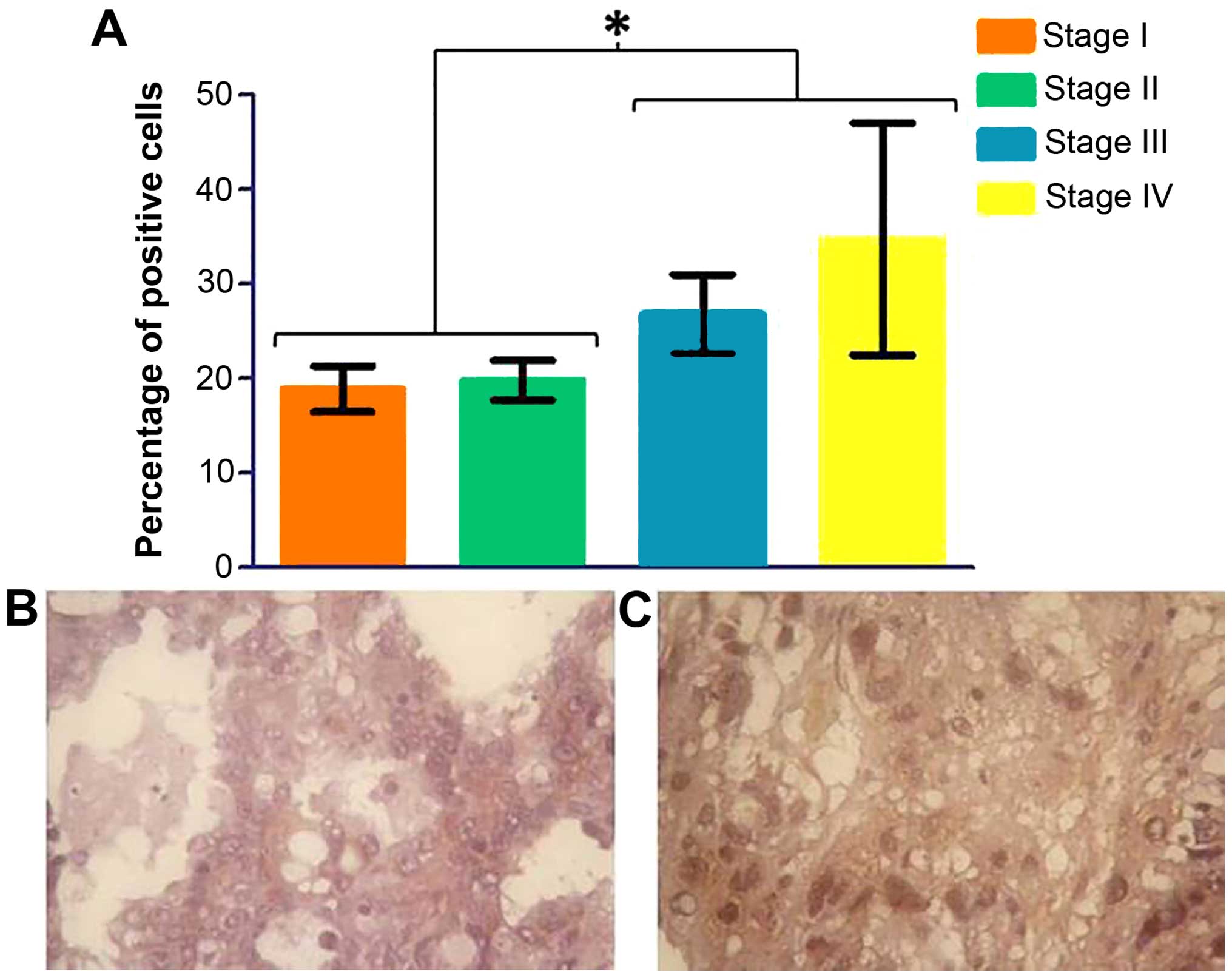
was seen in the grade of ovarian cancer (data not shown); however,
phospho-p70S6 kinase shows a significant increase in later stages
(Fig. 9).
Expression of mTOR components in ovarian
cancer patients
It is becoming more widely accepted that an
explorative test should be carried out for each set of samples to
determine a suitable reference gene in each case (11,13).
For this reason, we performed an analysis of reference gene
stability in ovarian clinical tissue samples using the geNorm™ 12
gene kit. Eight clinical samples were selected to represent the
whole cohort. The two most stable genes were RPL13A (ribosomal
protein L13A; a gene that encodes a component of the 60S ribosomal
subunit) and YWHAZ (a gene that encodes a 14-3-3 signal
transduction protein). These reference genes were used in qPCR
experiments involving clinical samples as they have been proven
stable enough to provide robust normalisation.
Therefore, qPCR for mTOR, DEPTOR, rictor and raptor
was carried out using the reference genes RPL13A and YWHAZ on cDNA
synthesised from the extracted RNA from ovarian tissue of ovarian
cancer patients (n=13) and non-affected controls (n=34). Data were
analysed using the ΔCq method and an RQ value calculated
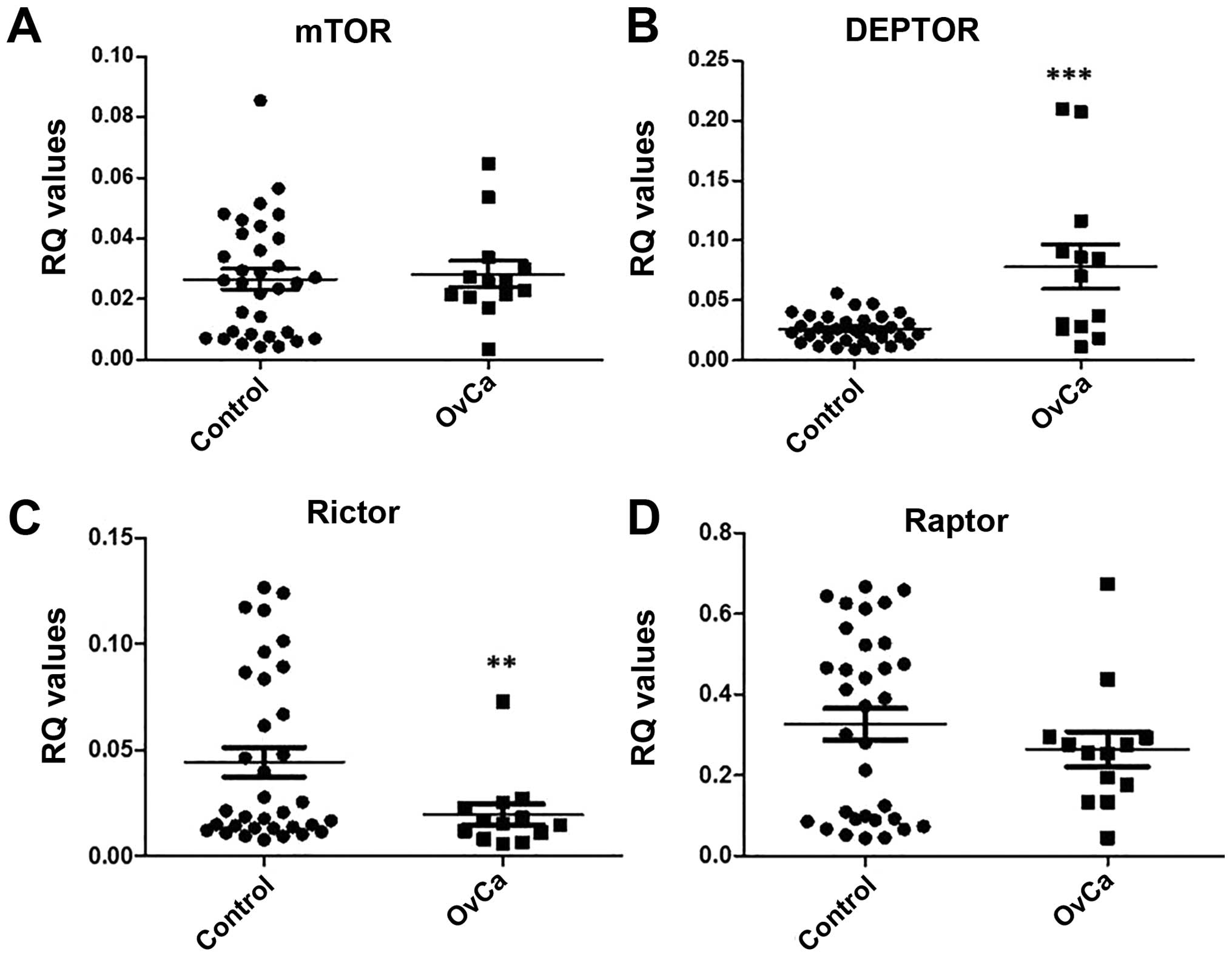
by 2−ΔCq. Results show a significant increase in DEPTOR
expression (p=0.0007) and a significant decrease in rictor
expression (p=0.0388) in comparison to control (Fig. 10).
Discussion
In this study, using two different ovarian cancer
cell lines and clinical samples, we investigated the effect of
rapalogues as well as dual kinase inhibitors in ovarian cancer. We
have used SKOV3 and MDAH-2774 cell lines as models for epithelial
ovarian cancer. The cell lines differed in their cells of origin,
SKOV3 cells are clear cell derived, and MDAH-2774 are endometrioid.
Using these cell lines we were able to study the two forms of
malignancy most associated with endometriosis (14–16).
In future, a useful addition to this study should be the inclusion
of primary malignant and non-malignant ovarian surface epithelial
cells. All key components of mTORC1 and mTORC2 complexes, i.e.,
mTOR, DEPTOR, rictor and raptor are expressed in both cell lines in
a similar manner. The predominant component was raptor, with DEPTOR
demonstrating minimal expression under basal (no treatment)
conditions. Using high-resolution flow cytometry we demonstrate for
the first time a cytoplasmic and nuclear expression of mTOR and
DEPTOR in MDAH-2774. Similar localisation was evident in SKOV3
corroborating previous data from our laboratory (10). Rictor and raptor appeared to be
expressed primarily in the cytoplasm (data not shown). It is
evident therefore, that under basal conditions both mTOR complexes
are expressed at mRNA and protein level.
Furthermore, we assessed the expression of these
genes in ovarian cancer patients (n=13) and controls (n=34). We
demonstrate a significant increase in DEPTOR expression and a
significant decrease in rictor expression in comparison to
controls. In order to assess mTOR pathway component gene expression
in a large cohort of patients (>50), we utilised the in
silico analysis method Oncomine™. mTOR, DEPTOR and raptor
expression was analysed, but due to the small sample size, rictor
data was not available. mTOR gene expression was significantly
higher (1.166-fold) in data from the Bonome dataset in ovarian
carcinoma (n=185) patients compared to controls (n=10). DEPTOR gene
expression was significantly higher (1.683-fold) in patients with
ovarian serous adenocarcinoma (n=43) compared to controls (n=10;
Yoshihara dataset). There were no significant changes in the
expression of raptor between ovarian serous adenocarcinoma patients
(n=43) and controls (n=10; Yoshihara dataset).
As mentioned, this study investigated the effects of
mTOR pathway inhibition in vitro. Western blot experiments
indicated cleavage of caspase 9 after 24-h treatment which was then
decreased by 48 h in SKOV3 cells. This indicates that initial
response to mTOR inhibitor treatment is to induce apoptosis but
this effect is rescued by 48 h of treatment. Caspase 9 is an
initiator caspase which cleaves the pro-form of caspase 3 to induce
apoptosis. However, there was no cleavage of caspase 3 in either of
the cell lines following treatment with mTOR inhibitors (data not
shown), suggesting that there is limited impact on apoptotic
events. This data was further supported by proliferation assays
showing no change in dead cell number over 72 h. Results showing no
significant change in dead cell number over the course of the
experiment in any case indicate that the effects of mTOR pathway
inhibition are cytostatic as opposed to cytotoxic. This data is
supported by previous research which shows that rapamycin and
temsirolimus cause cell cycle arrest at G0/G1 phase (17), whereas Eve treatment reduced viable
cell number, but was not able to induce apoptosis in SKOV3 cells
(18).
The inhibitory effects of Res and BEZ on SKOV3 cells
were significantly greater for these dual mTOR/PI3 kinase
inhibitors than for the rapalogues alone. This suggests that
additional targeting of PI3 kinase increases inhibitory efficacy in
these ovarian cancer cells. In the more proliferative MDAH-2774
cell line, 100 nM BEZ treatment profoundly decreased the number of
viable cells to almost basal level; however dual mTOR and PI3
kinase inhibitors were not more effective than inhibition by
rapamycin or rapalogue in these cells. The lower efficacy of dual
inhibitors observed in MDAH-2774 cells may be due to the negative
feedback loop directed to PI3 kinase that is relieved on mTOR
inhibition and can induce PI3 kinase signalling via Akt to cause
upregulation in mTORC2 signalling (19). Dual mTOR and PI3 kinase, or mTORC1
and mTORC2 inhibitors are able to target both mTORC1 signalling and
the after-effect of mTORC2 signalling and therefore provide better
repression of the mTOR pathway (20,21).
There is also evidence that a negative feedback loop exists from
mTOR to the MAP kinase pathway (22) and inhibition of both the MAP kinase
and PI3 kinase pathways has proven to be more effective than
targeting solely PI3 kinase (20).
Collectively, these data demonstrate that these inhibitors act in a
cell-specific manner within the microenvironment of ovarian
cancer.
Phosphorylation status of p70S6K was also assessed
to obtain a better understanding of the effects of these inhibitors
on mTORC1 activity. Rapamycin, rapalogues and BEZ (at high
concentration in MDAH-2774) completely dephosphorylated p70S6K in
both cell lines. In SKOV3 cells, treatment with 50 μM of Res also
dephosphorylated p70S6K. However, Res did not dephosphorylate
p70S6K in MDAH-2774 cells. These data corroborate previous findings
on p70S6K using BEZ in neuroblastoma cells (23), temsirolimus and everolimus in EGFR
mutant lung cancer cells, PC-9, and HCC827 (24). In one of our in vitro
models, Res had little effect on the phosphorylation status of
p70S6K. This again could be a cell-specific effect since Res
inhibits p70S6K in other cell lines such as the osteoblast-like
MC3T3-E1 cells (24), or the human
nasopharyngeal carcinoma (NPC) cells (26) to name a few.
Given the importance of p70S6K in ovarian cancer, we
mapped the protein expression of this phospho-p70S6K in 70 ovarian
cancer patients and controls. We demonstrate for the first time a
significant increase in staining for phospho-p70S6K with worsening
stage. Since p70S6K can be a prerequisite of tumour metastasis
(27), targeting this kinase
particularly in patients with high stage tumours could be of
therapeutic value. Recently it has been shown that phosphorylated
p70S6K predicted tamoxifen resistance in postmenopausal breast
cancer patients (28). Future
studies should concentrate on immunostaining for phosphorylated
p70S6K in a larger cohort of patients in order to define whether
this kinase is of a diagnostic or prognostic value in ovarian
cancer.
This study highlights the need for ‘tailor-made’
therapies against ovarian cancer depending on the gene expression
profile of the patient. The non-apoptotic effects of the mTOR
pathway inhibitors used in this study may exclude them from being
effective single agent therapies in the treatment of malignancy.
However, it has been shown that rapalogue treatment is able to
potentiate the apoptotic effects of tamoxifen, doxorubicin, UCN-01
and cisplatin treatment in vitro, indicating that mTOR
pathway inhibitors may be useful in combinatorial therapeutic
approaches to cancer treatment (18,29–31).
Using mTOR pathway inhibitors may therefore allow for lower doses
of cytotoxic chemotherapeutic agents in cancer treatment and
therefore a reduction in the harmful side-effects brought about by
these drugs (31).
References
|
1
|
De Cecco L, Marchionni L, Gariboldi M,
Reid JF, Lagonigro MS, Caramuta S, Ferrario C, Bussani E,
Mezzanzanica D, Turatti F, et al: Gene expression profiling of
advanced ovarian cancer: Characterization of a molecular signature
involving fibroblast growth factor 2. Oncogene. 23:8171–8183. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hay N and Sonenberg N: Upstream and
downstream of mTOR. Genes Dev. 18:1926–1945. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim SH, Juhnn YS and Song YS: Akt
involvement in paclitaxel chemoresistance of human ovarian cancer
cells. Ann NY Acad Sci. 1095:82–89. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schiff PB and Horwitz SB: Taxol stabilizes
microtubules in mouse fibroblast cells. Proc Natl Acad Sci USA.
77:1561–1565. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huang S and Houghton PJ: Inhibitors of
mammalian target of rapamycin as novel antitumor agents: From bench
to clinic. Curr Opin Investig Drugs. 3:295–304. 2002.PubMed/NCBI
|
|
6
|
Kurmasheva RT, Huang S and Houghton PJ:
Predicted mechanisms of resistance to mTOR inhibitors. Br J Cancer.
95:955–960. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mabuchi S, Altomare DA, Connolly DC,
Klein-Szanto A, Litwin S, Hoelzle MK, Hensley HH, Hamilton TC and
Testa JR: RAD001 (Everolimus) delays tumor onset and progression in
a transgenic mouse model of ovarian cancer. Cancer Res.
67:2408–2413. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Squillace RM, Miller D, Cookson M,
Wardwell SD, Moran L, Clapham D, Wang F, Clackson T and Rivera VM:
Antitumor activity of ridaforolimus and potential cell-cycle
determinants of sensitivity in sarcoma and endometrial cancer
models. Mol Cancer Ther. 10:1959–1968. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Takano M, Kikuchi Y, Kudoh K, Goto T,
Furuya K, Kikuchi R, Kita T, Fujiwara K, Shiozawa T and Aoki D:
Weekly administration of temsirolimus for heavily pretreated
patients with clear cell carcinoma of the ovary: A report of six
cases. Int J Clin Oncol. 16:605–609. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Foster H, Coley HM, Goumenou A, Pados G,
Harvey A and Karteris E: Differential expression of mTOR signalling
components in drug resistance in ovarian cancer. Anticancer Res.
30:3529–3534. 2010.PubMed/NCBI
|
|
11
|
Bustin SA, Benes V, Garson JA, Hellemans
J, Huggett J, Kubista M, Mueller R, Nolan T, Pfaffl MW, Shipley GL,
et al: The MIQE guidelines: Minimum information for publication of
quantitative real-time PCR experiments. Clin Chem. 55:611–622.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
13
|
Jacob F, Guertler R, Naim S, Nixdorf S,
Fedier A, Hacker NF and Heinzelmann-Schwarz V: Careful selection of
reference genes is required for reliable performance of RT-qPCR in
human normal and cancer cell lines. PLoS One. 8:e591802013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pearce CL, Templeman C, Rossing MA, Lee A,
Near AM, Webb PM, Nagle CM, Doherty JA, Cushing-Haugen KL, Wicklund
KG, et al; Ovarian Cancer Association Consortium. Association
between endometriosis and risk of histological subtypes of ovarian
cancer: A pooled analysis of case-control studies. Lancet Oncol.
13:385–394. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Brinton LA, Sakoda LC, Sherman ME,
Frederiksen K, Kjaer SK, Graubard BI, Olsen JH and Mellemkjaer L:
Relationship of benign gynecologic diseases to subsequent risk of
ovarian and uterine tumors. Cancer Epidemiol Biomarkers Prev.
14:2929–2935. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Merritt MA, Green AC, Nagle CM and Webb
PM; Australian Cancer Study (Ovarian Cancer); Australian Ovarian
Cancer Study Group. Talcum powder, chronic pelvic inflammation and
NSAIDs in relation to risk of epithelial ovarian cancer. Int J
Cancer. 122:170–176. 2008. View Article : Google Scholar
|
|
17
|
Fagone P, Donia M, Mangano K, Quattrocchi
C, Mammana S, Coco M, Libra M, McCubrey JA and Nicoletti F:
Comparative study of rapamycin and temsirolimus demonstrates
superimposable anti-tumour potency on prostate cancer cells. Basic
Clin Pharmacol Toxicol. 112:63–69. 2013. View Article : Google Scholar
|
|
18
|
Treeck O, Wackwitz B, Haus U and Ortmann
O: Effects of a combined treatment with mTOR inhibitor RAD001 and
tamoxifen in vitro on growth and apoptosis of human cancer cells.
Gynecol Oncol. 102:292–299. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Peterson TR, Laplante M, Thoreen CC,
Sancak Y, Kang SA, Kuehl WM, Gray NS and Sabatini DM: DEPTOR is an
mTOR inhibitor frequently overexpressed in multiple myeloma cells
and required for their survival. Cell. 137:873–886. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zitzmann K, Rüden J, Brand S, Göke B,
Lichtl J, Spöttl G and Auernhammer CJ: Compensatory activation of
Akt in response to mTOR and Raf inhibitors - a rationale for
dual-targeted therapy approaches in neuroendocrine tumor disease.
Cancer Lett. 295:100–109. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hisamatsu T, Mabuchi S, Matsumoto Y,
Kawano M, Sasano T, Takahashi R, Sawada K, Ito K, Kurachi H,
Schilder RJ, et al: Potential role of mTORC2 as a therapeutic
target in clear cell carcinoma of the ovary. Mol Cancer Ther.
12:1367–1377. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Carracedo A, Ma L, Teruya-Feldstein J,
Rojo F, Salmena L, Alimonti A, Egia A, Sasaki AT, Thomas G, Kozma
SC, et al: Inhibition of mTORC1 leads to MAPK pathway activation
through a PI3K-dependent feedback loop in human cancer. J Clin
Invest. 118:3065–3074. 2008.PubMed/NCBI
|
|
23
|
Durbas M, Horwacik I, Boratyn E, Kamycka E
and Rokita H: GD2 ganglioside specific antibody treatment
downregulates PI3K/Akt/mTOR signaling network in human
neuroblastoma cell lines. Int J Oncol. 47:1143–1159.
2015.PubMed/NCBI
|
|
24
|
Ishikawa D, Takeuchi S, Nakagawa T, Sano
T, Nakade J, Nanjo S, Yamada T, Ebi H, Zhao L, Yasumoto K, et al:
mTOR inhibitors control the growth of EGFR mutant lung cancer even
after acquiring resistance by HGF. PLoS One. 8:e621042013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kondo A, Otsuka T, Kuroyanagi G, Yamamoto
N, Matsushima-Nishiwaki R, Mizutani J, Kozawa O and Tokuda H:
Resveratrol inhibits BMP-4-stimulated VEGF synthesis in
osteoblasts: Suppression of S6 kinase. Int J Mol Med. 33:1013–1018.
2014.PubMed/NCBI
|
|
26
|
Zhang M, Zhou X and Zhou K: Resveratrol
inhibits human nasopharyngeal carcinoma cell growth via blocking
pAkt/p70S6K signaling pathways. Int J Mol Med. 31:621–627.
2013.PubMed/NCBI
|
|
27
|
Ip CK, Yung S, Chan TM, Tsao SW and Wong
AS: p70 S6 kinase drives ovarian cancer metastasis through
multicellular spheroid-peritoneum interaction and P-cadherin/b1
integrin signaling activation. Oncotarget. 5:9133–9149. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Beelen K, Opdam M, Severson TM, Koornstra
RH, Vincent AD, Wesseling J, Muris JJ, Berns EM, Vermorken JB, van
Diest PJ, et al: Phosphorylated p-70S6K predicts tamoxifen
resistance in postmenopausal breast cancer patients randomized
between adjuvant tamoxifen versus no systemic treatment. Breast
Cancer Res. 16:R62014. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Avellino R, Romano S, Parasole R, Bisogni
R, Lamberti A, Poggi V, Venuta S and Romano MF: Rapamycin
stimulates apoptosis of childhood acute lymphoblastic leukemia
cells. Blood. 106:1400–1406. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hahn M, Li W, Yu C, Rahmani M, Dent P and
Grant S: Rapamycin and UCN-01 synergistically induce apoptosis in
human leukemia cells through a process that is regulated by the
Raf-1/MEK/ERK, Akt, and JNK signal transduction pathways. Mol
Cancer Ther. 4:457–470. 2005.PubMed/NCBI
|
|
31
|
Beuvink I, Boulay A, Fumagalli S,
Zilbermann F, Ruetz S, O'Reilly T, Natt F, Hall J, Lane HA and
Thomas G: The mTOR inhibitor RAD001 sensitizes tumor cells to
DNA-damaged induced apoptosis through inhibition of p21
translation. Cell. 120:747–759. 2005. View Article : Google Scholar : PubMed/NCBI
|