Introduction
The heat shock proteins (HSPs) are ubiquitously
expressed in almost all cells and across species. The protein
family was so named owing to their initial discovery, namely,
identified from cells under heat shock and is now known to be
involved in a number of other cellular stresses including heat,
carcinogens, mechanical and chemical stress (1,2).
These proteins serve as molecular chaperones and act as a mechanism
to assist protein folding, repair damaged proteins, or assist in
degrading the unwanted proteins after stress and injury, thus by
doing so the proteins protect the injured cells from potentially
lethal damage (3,4). The HSP proteins are thus important
regulators in normal physiology, but are also involved in diseases
including cardiovascular, wound healing and cancer.
Similarly discovered from cells under heat shock,
HSP27 has been found to be widely involved in other types of cell
stress, including oxidative response (5–8). The
HSP27 is a protein of approximately 27 kDa in size and belongs to
the HSP family in which sizes vary between 8–150 kDa. The main
cellular function of HSP27 is protecting cells from becoming
apoptotic, or anti-apoptotic. Although this role is critically
important in homeostasis, it has serious implications in clinical
cancer. Firstly, HSP27 has been found to be overexpressed in a
variety of human cancers and linked to a poor outcome for those
cancer patients (9–13); secondly, high levels of HSP27 in
cancer cells have been shown to promote cancer cell growth
(14–16); and thirdly, HSP27 was found to be
extremely elevated following chemotherapy, most probably as a
protective mechanism to chemical stress (17–19).
In the latter case, HSP27 and HSP70 were two of the gene products
that were upregulated in a cisplatin resistant ovarian cancer cell
line (19). Thus, this has caused
an enormous challenge to cancer treatment and is frequently linked
to chemotherapy resistance in cancer.
The key mode of activation of HSP27 is by
phosphorylation. Phosphorylation of HSP27 allows itself to form
oligomers which may facilitate the chaperone process under cellular
stress (20,21). Phospho-HSP27 interacts with other
pro-apoptotic proteins such as DAXX and ASK1 and therefore by
blocking pro-apoptotic protein interaction blocks the apoptotic
pathway (22,23). Other known pathways of HSP27
include interaction with AKT1 and the oestrogen receptor. By
interacting with these signalling proteins, phospho-HSP27 blocks
some of their actions in cancer cells. The most commonly reported
sites of phosphorylation with HSP27 are S15, S78, S82 and S86.
Consequently, phosphorylated HSP27 protein levels have been found
to be frequently raised in clinical cancer tissues and
chemoresistant cancer cells (24,25).
Together, it is suggested that HSP27 is a valid
target for cancer treatment. Methods to either reduce the protein
levels of HSP27 or the phosphorylation of HSP27, would be desirable
in devising cancer treatment (26). A new anti-HSP27 agent, known as
OGX427 (Apatorsen) has entered into a late phase clinical trial in
patients with various malignant conditions including pancreatic,
lung, bladder or prostate cancer, alone or in combination with
other chemotherapies (27–30). Early studies have shown a
favourable outcome from the new treatment.
There have been clinical trials which have shown
that YangZheng XiaoJi (YZXJ), a combination of traditional
Chinese medicinal herbs benefit cancer patients; however, how this
mechanism of action is achieved remains unknown (31–34).
The present study reports an accidental and surprising discovery
that a traditional Chinese herbal medicine, known as YangZheng
XiaoJi (YZXJ) used in treating patients with cancer, can
suppress the phosphorylation of HSP27 and cell functions related to
this protein.
Materials and methods
Materials
Antibodies to human HSP27 (sc-13132), caspase-3
(sc-7148), caspase-8 (sc-70501), caspase-9 (sc-17784), phospho-FAK
(sc-81493), and GAPDH (sc-32233) were purchased from Santa Cruz
Biotechnologies Inc. (Santa Cruz, CA, USA). Therapeutic agents
including cisplatin, topotecan, pacilitaxol and 5-FU were purchased
from Sigma-Aldrich (Poole, Dorset, UK). Antibodies to FAK
(ab131435) and phospho-HSP-27 (S86) (ab17938) were purchased from
Abcam (Cambridge, UK). FITC/TRITC-conjugated Phalloidin and FITC-
and TRITC conjugated secondary antibodies were from Sigma-Aldrich.
Secondary antibodies (fluorescence- and HRT-conjugated) were also
from Sigma-Aldrich. Anti-HSP27 siRNA, control siRNA and
transfection reagents were also obtained from Santa Cruz
Biotechnologies Inc.
Cells
Human gastric cancer (AGS and HGC27), pancreatic
cancer (PANC1), ovarian cancer (SKOV3 and COV504), lung cancer
(A549 and SKMES1), breast cancer (MDA MB-231), prostate cancer
(PC-3) cells were purchased from LGC Standard/ATCC (Southampton,
UK). Ovarian cancer cells A2780 and its cisplatin resistant strain
A2780/CP70 were gifts from Imperial College London (Dr Euan
Stronach).
YangZheng XiaoJi extracts
An extract from YZXJ, named DME25 was
prepared using a DMSO based method that has been described in full
(35,36). The herbal medicinal formula,
YangZheng XiaoJi was obtained from Yiling Pharmaceuticals
(Shijiazhuang, HeBei, China). The formula contained the following
16 ingredients: Panax ginseng C.A. Mey, Astragalus
memebranaceus (Fisch.) Bge.var. mongholicus (Bge.) Hsiao,
Ligustrum lucidum Ait, Curcuma phaeocaulis Val, Ganodema lucidum,
Gynostemma pentaphylla (Thunb) Mak, Atractylodes macrocephala
Koidz, Scutellaria barbata D.Don, Oldenlandia diffusa (willd.)
Roxb, Poria cocos, Duchesnea indica Focke, Solanum lyratum Thunb,
Artemisia scoparia (Bge.) Ki, Cynanchum paniculatum Kitag,
Eupolyphaga sinensis Walker, and Gallus domesticus
Brisson. For experimental use, the extract was diluted in the
respective cell culture media and the dilution range was between
1:100 to 1:2,000.
Immunofluorescent staining
Cells, plated with test agents in 8-well chamber
slides, were first fixed using 4% formalin, lightly permeabilised
with 0.1% Triton X100 for 5 min and blocked with a Tris buffer (25
mM, pH 8.4) that contained 7.5% pre-immuned goat serum for 1 h.
Primary antibodies (including anti-HSP27, anti-phospho-HSP27 (S86),
anti-caspases), diluted in the blocking buffer was added to the
respective slides which were kept in the dark at full humidity on a
slow moving platform for 1 h, and then washed thoroughly.
FITC-tagged secondary antibodies were then added. After 1 h, the
slides were washed thoroughly and mounted using FluorSave™
(Calbiochem, Nottingham, UK). TRITC-conjugated phalloidin was
diluted at final concentration of 10 μg/ml and added to the cells
together with the secondary antibodies. The slides were examined on
an Olympus microscope and photographed using a Hamamatsu digital
camera. The staining intensity was determined using ImageJ
software.
Knockdown of HSP27
Cells at approximate 70% confluence were rinsed with
BSS buffer. Anti-HSP27 siRNA, diluted in RNA free water was first
mixed with a transfection reagent and left at room temperature for
30 min before being added to the cells. After 7 h medium was
replaced and cells were left to incubate for another 24 h.
Expression of HSP27 was tested using RT-PCR.
Cell growth assay
Cancer cells were first seeded in 96-well tissue
culture plates and allowed to adhere. The test materials and their
combinations were then added to the cells, which were subsequently
incubated for a period of 72 h. After removing the culture media,
cells were fixed with 4% formalin and stained with 0.5% Crystal
violet. After extensive washing, the staining was extracted with
10% acetic acid and plates were read on a multiple channel plate
reader at 540 nm (Elx800; Bio-Tek, Swindon, UK). The test regimens
included chemotherapeutic agent alone, DME25 alone, DME25 and
chemotherapeutic agent combination and DME25 pre-treatment followed
by chemotherapeutic agent.
Protein arrays for detecting
phosphorylation changes
Cancer cells at 90% confluence in two T75 tissue
culture flasks were washed with BSS buffer and then placed into a
fresh batch of DMEM supplemented with 5% FCS. After 5 h, treatment
was added, again in 5% FCS. After a 5-h period, cells were removed
from the flasks with a cell scraper. The cells were pelleted using
a centrifuge at 2,500 rpm for 5 min. Lysis buffer was added to the
cell pellets and placed on a spinning wheel for 1 h at 4°C. The
lysates were then spun at 12,000 × g for 10 min at 4°C. The
supernatants were carefully collected and the insolubles discarded.
Based on absorbance the protein concentration in the cell lysates
were quantified using the scatter line chart of Microsoft Excel and
then adjusted to 2 mg/ml. The samples were stored at −20°C until
use. Antibody based protein arrays, namely KAM850, which has 854
capture antibodies spotted on to each array slides (Kinexus
Bioinformatics Ltd., Vancouver, Canada) were used in the present
study. The following are the key parameters collected and used for
the data analysis: Globally Normalized Signal Intensity, Background
corrected intensity values are globally normalized. The Globally
Normalized Signal Intensity was calculated by summing the
intensities of all the net signal median values for a sample. Z
Scores, Z score transformation corrects data internally within a
single sample. Z Score Difference, The difference between the
observed protein Z scores in samples in comparison. Z Ratios,
Divide the Z Score Differences by the SD of all the differences for
the comparison.
Electric cell-substrate impedance sensing
(ECIS) based analyses on cell adhesion and cell migration
Briefly, 96-well W96E1 microarrays were used on the
ECIS Ztheta instrument (Applied Biophysics Ltd., Troy, NJ, USA).
Lung cancer cells were added to the wells of the array, followed by
immediate tracking of cell adhesion over a range of frequencies
(1,000 to 64,000 Hz) using automated modules. The adhesion was
analysed using the mathematical modelling methods as previously
described. For cellular migration, confluent lung cancer monolayers
in the arrays were electrically wounded (2,000 mA for 20 sec each),
after which the migration of the cells was immediately tracked,
again over a range of frequencies. All the experiments were
conducted in triplicate.
Western blotting analysis of HSP27
proteins
Proteins, prepared in a similar fashion to that for
protein analysis, were separated on a SDS-PAGE genes, following
transfer of proteins to nitrocellulose membranes, the membranes
were probed with the respective antibodies in a set protocol that
including extensive washings in between the antibody probing and
finally detecting the antibody signals using chemilluminescence
reagents. The membranes were visualised using an imager (G-Box;
Syngene, Cambridge, UK).
Results
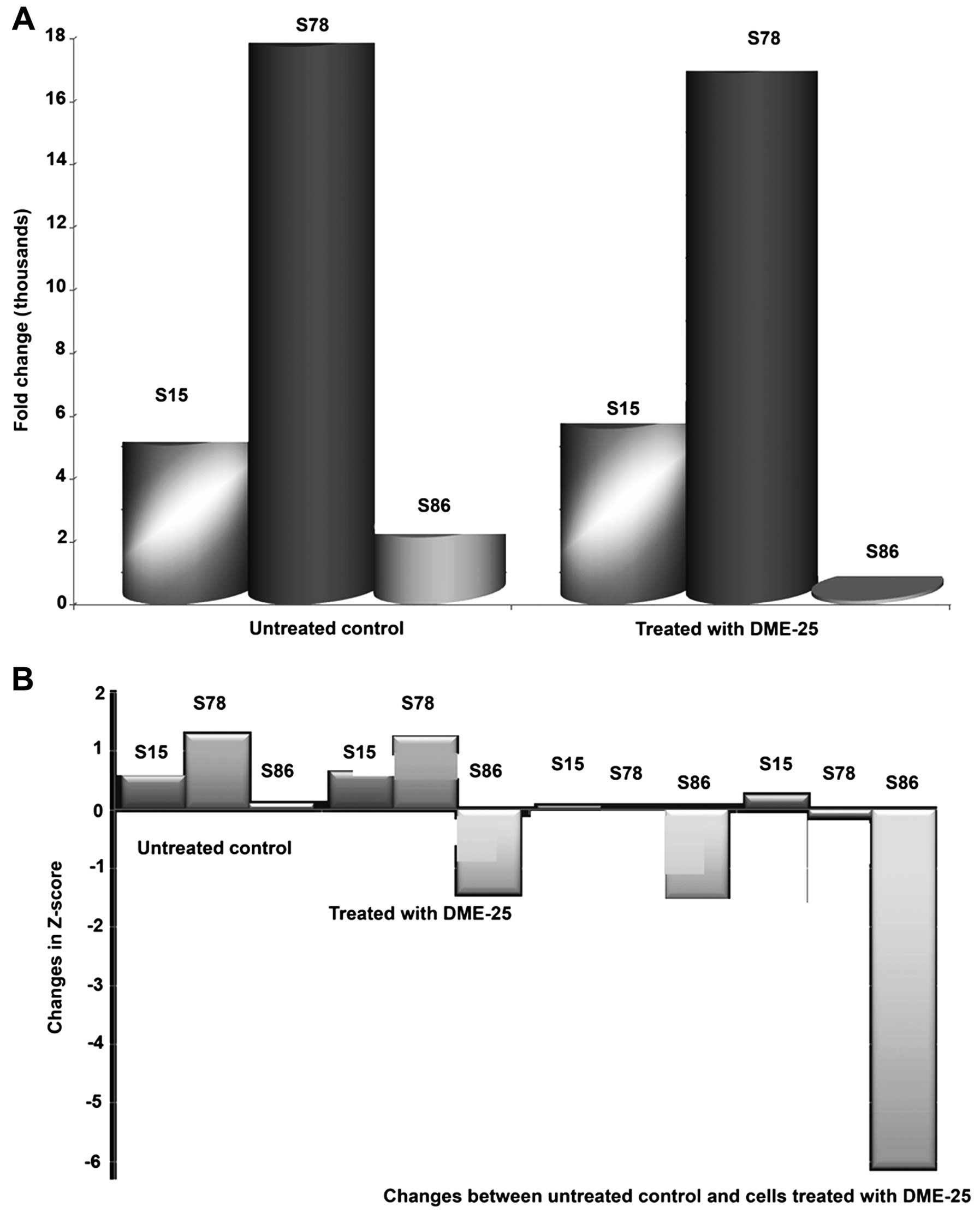
DME25 has a marked effect on the
phosphorylation of HSP27
Using a protein array, we found that the YZXJ
extract DME25 resulted in a marked reduction of phosphorylation of
HSP27, particularly on Serine86 (S86) phosphorylation, in SKMES-1
lung cancer and PANC-1 pancreatic cancer cells (Fig. 1A and B, respectively).
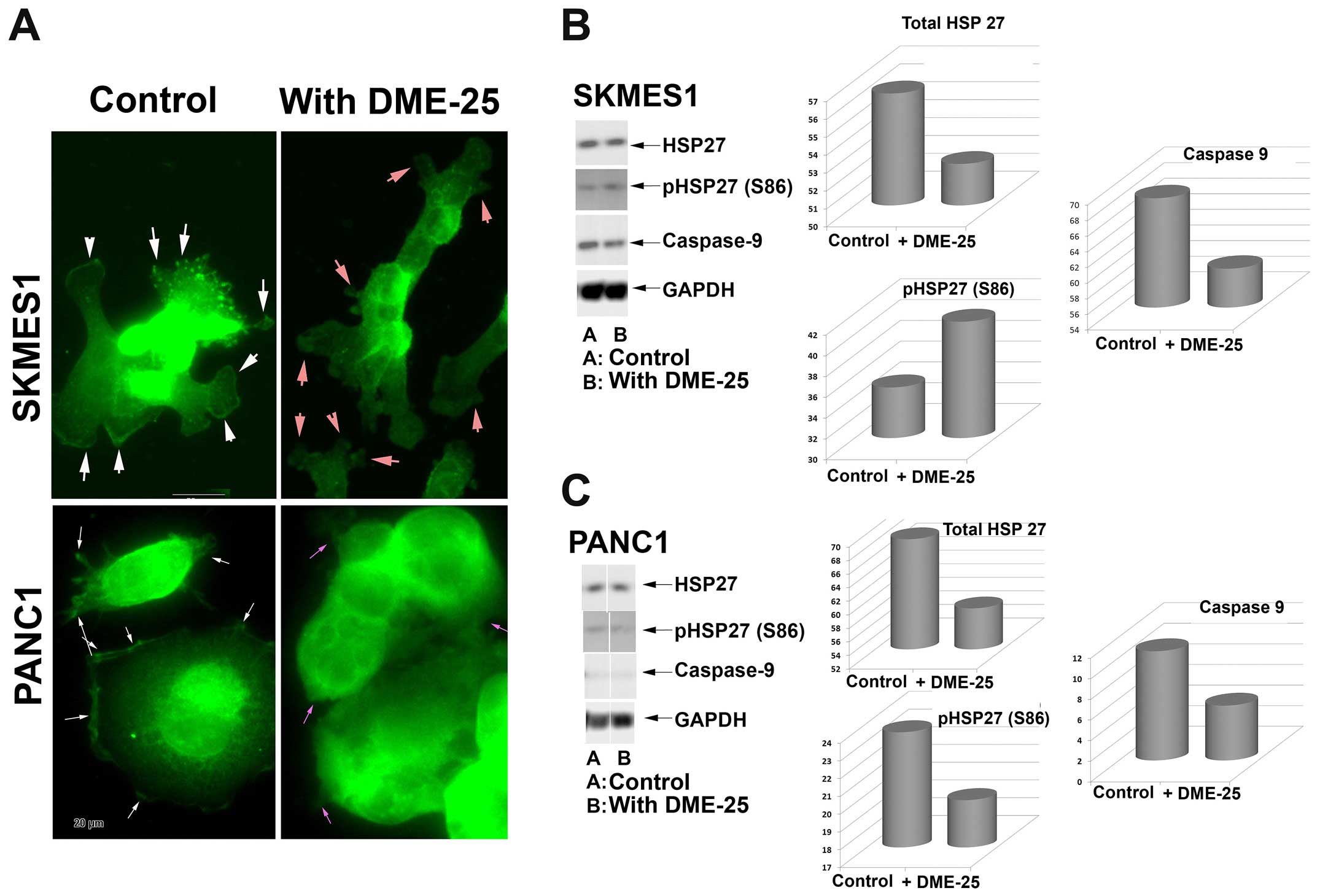
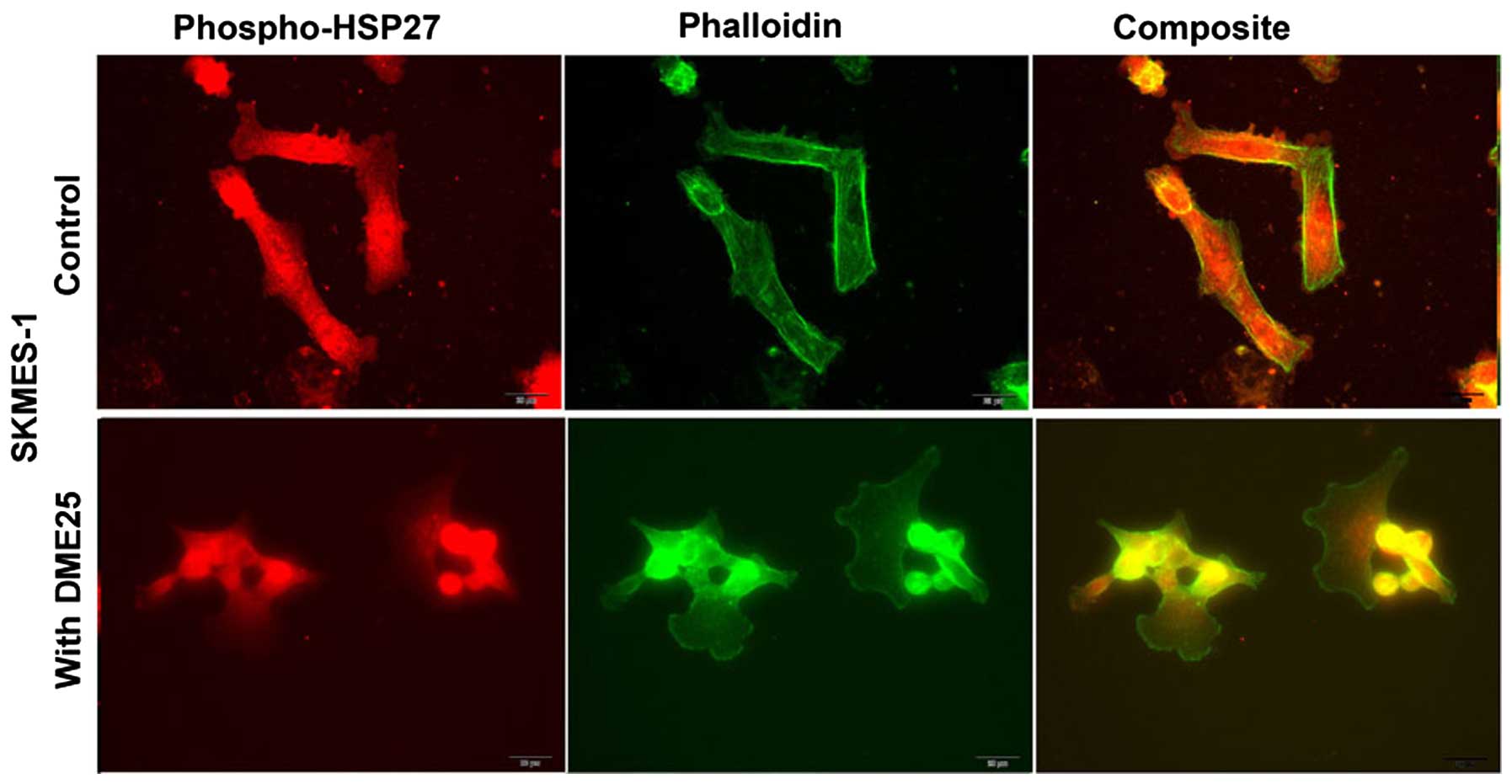
Localisation of HSP27 and phospho-HSP27
(S86) in cancer cells, assessed by immunofluorescence
HSP27 was seen broadly in the nucleus and in
cytoplasmic region of lung cancer cells (Fig. 2A, top panel). It is very
interesting to note that both total HSP27 and phospho-HSP27 was
localised in focal adhesion and pseudopodia regions of the cells.
Treatment of the lung cancer cells, SKMES1 with DME25 resulted in
loss of phospho-HSP27 from the focal adhesion and pseudopodia
regions of the cells, although the changes of total HSP27 did not
appear to differ (Fig. 2). The
same changes of phospho-HSP27 were seen in pancreatic cancer cells,
PANC1 (Fig. 2A, bottom panel) as
well as in ovarian cancer cell SKOV3 and gastric cancer AGS cells
(data not shown). The inhibition on levels and activation of HSP27
was also demonstrated by western blotting analysis in both the lung
and pancreatic cells (Fig. 2B and
C, respectively).
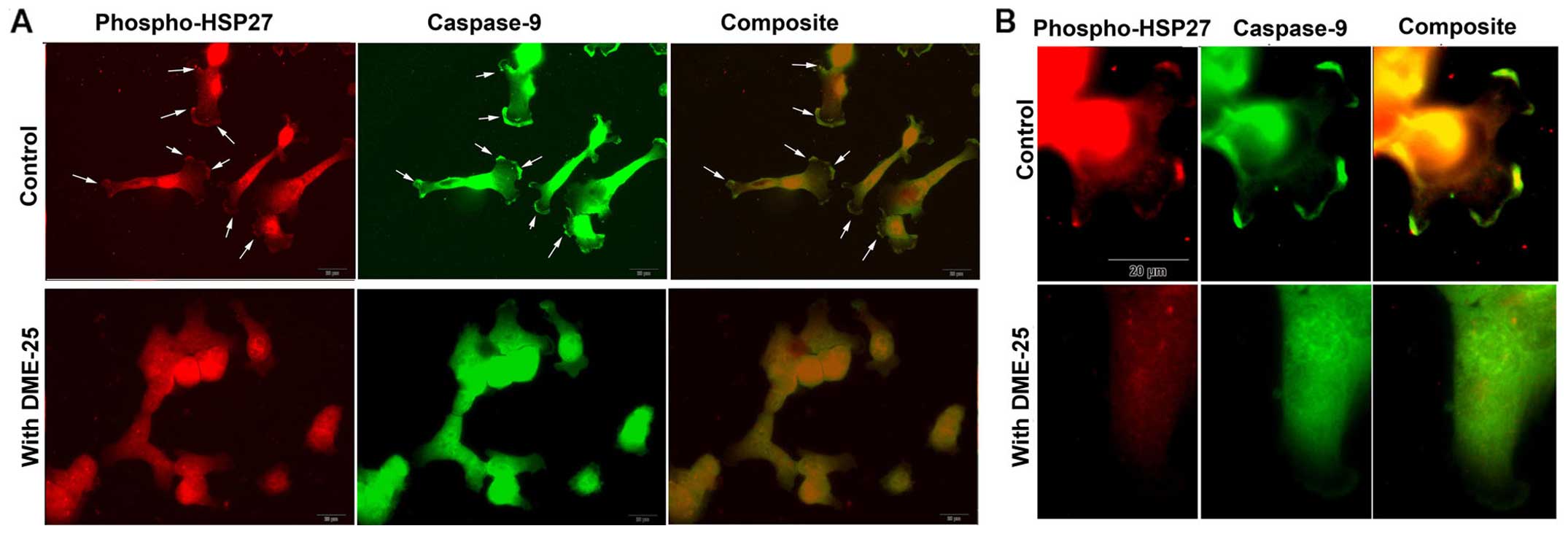
Activated HSP27 is co-localised with
caspase-9 in cancer cells, which is prevented when cells are
treated with DME25
When co-stained for phospho-HSP27 (S86) and
caspase-9, it was found that both molecules co-localised in regions
of focal adhesion (FAC) and pseudopodia (Fig. 3A and B). It was very interesting to
note that when cells were treated with DME25, this pattern of
co-localisation appears to break (Fig.
3B).
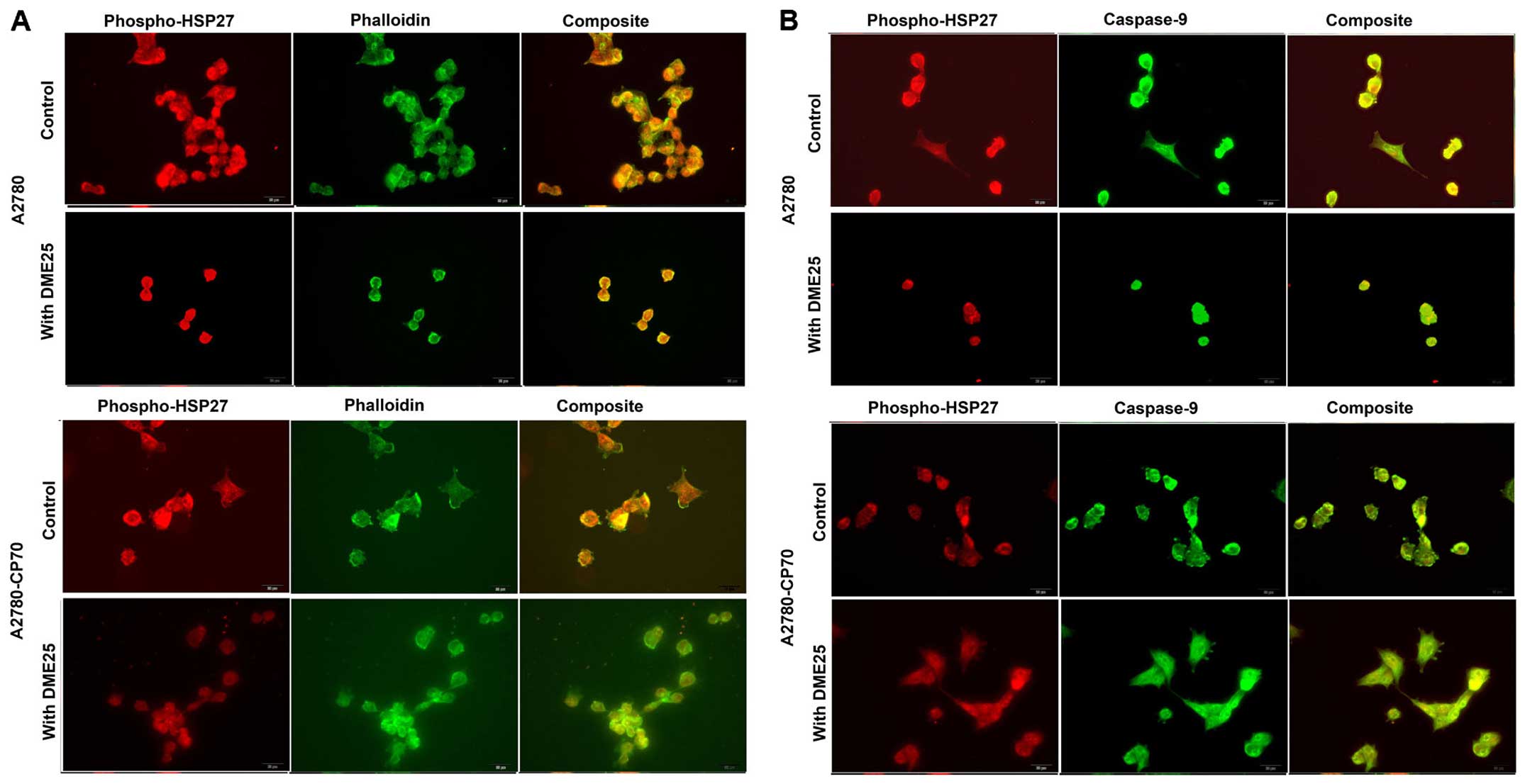
Phospho-HSP27 and its co-localisation
with filamentous actin and caspase-9 in cancer cells and in
chemoresistant cancer cells
Using dual fluorescence staining, we have shown that
phospho-HSP27 and F-actin did co-localise in FAC regions in both
wild-type ovarian cancer A2780 (Fig.
4A, top panels) and in cisplatin resistant A2780/CP70 cells
(Fig. 4 bottom panels). The same
was seen with lung cancer cells (Fig.
5). In all the cells tested, treatment with DME25 reduced the
location of phospho-HSP27 to the FAC region. The co-localisation of
phospho-HSP27 and caspase-9 in wild-type A2780 and cisplatin
resistant A2780/CP70 were also reduced in the presence of DME25
(Fig. 4B).
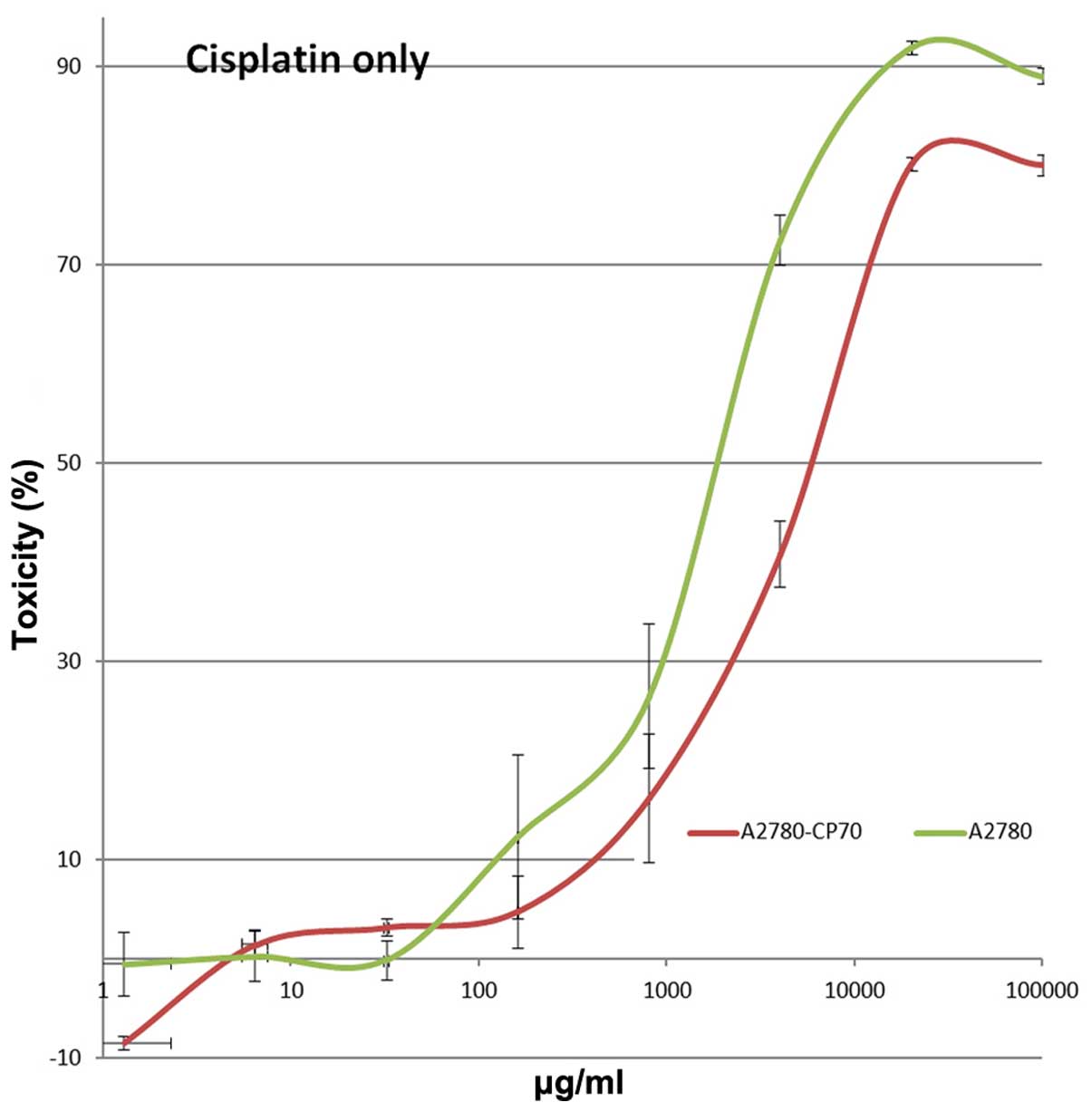
Effect of DME25 on the growth of ovarian
cancer cells
A2780/CP70 showed approximately 8 times less
sensitivity to cisplatin compared with the wild-type parental cells
(Fig. 6). Co-culture of both
cisplatin and DME25 resulted in the cells becoming more sensitive
to cisplatin, as seen for lung, gastric, pancreatic and ovarian
cancer cells (Tables I–V). However, knocking down HSP27 by siRNA
did not significantly influence the sensitivity of the cells to the
chemotherapeutic agents (data not shown).
 | Table IEffects of cisplatin on the growth of
cancer cells in combination with DME25. |
Table I
Effects of cisplatin on the growth of
cancer cells in combination with DME25.
| 16 ng/ml | 400 ng/ml | 10 μg/ml |
|---|
|
|
|
|
|---|
| Cisplatin | Control | YZXJ | Control | YZXJ | Control | YZXJ |
|---|
| AGS | 7.9±2.5 | 9.1±0.7 | 20.8±2.0 | 14.5±2.6 | 93.1±0.5 | 91.5±0.5 |
| MIAPACA2 | 50.1±4.5 | 92.1±1.5 | 77.5±1.5 | 93.1±4.4 | 77.8±2.5 | 93.2±2.6 |
| A549 | 20.1±1.7 | 12.2±2.7 | 50.4±1.3 | 44.1±1.7 | 90.4±0.4 | 89.9±0.2 |
| SKMES1 | 1.7±0.3 | 34.6±1.7 | 23.0±1.4 | 46.3±2.0 | 80.5±3.5 | 86.9±2.6 |
| COV504 | NC | 3.8±6.2 | 27.2±4.7 | 43.4±2.7 | 74.6±2.8 | 85.1±1.3 |
| SKOV3 | NC | 17.2±1.9 | 0.5±1.5 | 9.8±2.6 | 46.7±1.5 | 66.7±2.7 |
| Table VEffects of cisplatin on the growth of
wild-type and cisplatin-resistant ovarian cancer cells in
combination with DME25. |
Table V
Effects of cisplatin on the growth of
wild-type and cisplatin-resistant ovarian cancer cells in
combination with DME25.
| A2780 | A2780/CP70 |
|---|
|
|
|
|---|
| Treatment | 160 ng/ml | 4 μg/ml | 160 ng/ml | 4 μg/ml |
|---|
| Cisplatin only | 12.3±8.2 | 72.5±2.5 | 4.7±4.0 | 40.8±8.6 |
| Cisplatin +
YZXJ | 72.4±3.6 | 80.5±3.3 | 26.8±0.4 | 46.3±1.3 |
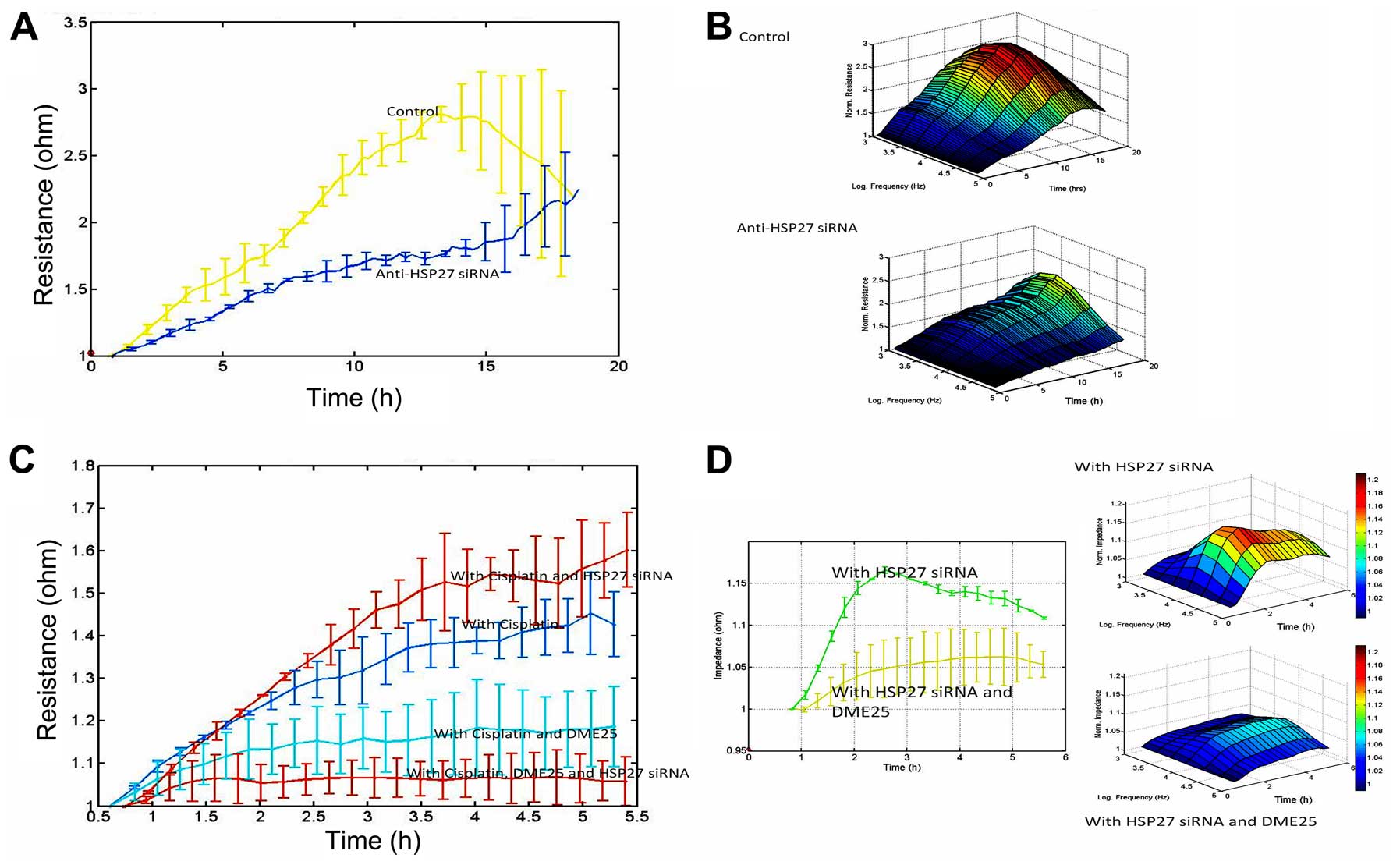
HSP27 shows marked influence on cell
migration and acts synergistically with YangZheng XiaoJi on cancer
cell migration
In light of the co-localisation of HSP27 and
phalloidin at the focal adhesion regions of cancer cells and our
previous reports on the impact of DME25 on the migration of various
cells including cancer cells, we tested how HSP27 might affect cell
migration. It was readily demonstrable that knocking down HSP27
resulted in a sustained reduction of cell migration (Fig. 7A and B). It was further confirmed
that knocking down HSP27 increased sensitivity of cancer cells to
low dosage chemotherapeutic agents and in particular when DME25 was
present (Fig. 7C and D).
Discussion
The current study stemmed from a surprising finding
in our search for the potential pathways that are influenced by
DME25, a traditional Chinese medicinal formula used in cancer
treatment.
HSP27, YangZheng XiaoJi and sensitivity
to chemotherapeutic agents
The role of HSP27 in the sensitivity to chemotherapy
agents is not without controversy. For example, it has been
reported recently that overexpression of HSP27 in pancreatic cancer
cells would increase the sensitivity of the cancer cells to
gemcitabine (37). Although
reasons for this are not clear, the type of drugs used in the study
may be an explanation, as these drugs are transported and act in
cancer cells via different mechanisms.
HSP27 acts as a cyto-protective molecule during
androgen independence in prostate cancer patients and in prostate
cancer cell models (38).
Likewise, levels of HSP27 correlated with resistance to irinotecan
in colorectal cancer cells (39).
HSP27, together with HSP70 is linked to the sensitivity of
colorectal cancer to the naturally-occurring anticancer agent
curcumin (40).
Knocking down HSP27 sensitises glioblastoma
multiforme tumour cells to the HSP90 inhibitor,
17-N-allylamino-17-demethoxygeldanamycin (17-AAG) and staurosporine
by changes in apoptosis (41). The
same was seen in glioma cells, in that silencing HSP27 resulted in
an increase in apoptosis in response to temozolomide and quercetin
(42).
HSP27, YangZheng XiaoJi and cell
migration
The finding that HSP27 is co-localised with
phalloidin to the focal adhesion regions of cancer cells is
interesting. From the immunofluorescence staining, HSP27
particularly the phosphorylated form of the protein, is frequently
co-localised with Phallodin at the focal adhesion regions of cancer
cells, where caspase-9 is also seen. It is of interest to note that
inclusion of YZXJ extract DME25 in the study markedly reduce
the presence of phospho-HSP27 (S86) at the focal adhesions. Given
the key role of focal adhesion in cell-matrix adhesion and cellular
migration, it is plausible to suggest that activation of HSP27 (by
way of phosphorylation) coordinates with other proteins in the
focal adhesion area, such as integrins and the focal adhesion
kinase (FAK) to regulate the adhesion and migration of cancer
cells. We have recently reported that YZXJ was able to inhibit the
phosphorylation of FAK in cancer cells and in endothelial cells
(35,43).
Together with the finding that knocking down HSP27
resulted in an increase in cellular migration in cancer cells, it
is suggested that HSP27, in particular when combined with YZXJ, is
an important regulator of EMT reversal or MET. It has been reported
recently that HSP27 is key to IL-6 dependent and IL-6 independent
EMT in prostate cancer cells, by acting on IL-6 induced activation
of STAT3/Twist and Snail (44,45).
It is also involved in EGF induced EMT in prostate cancer cells by
influencing the migration of these cells (46,47).
Other than the reported effects on cancer cells, HSP27 has also
been shown to be a mediator for angiogenesis (48), suggesting that HSP27 is involved in
a wider range of processes in cancer development and
progression.
Taken together, it is clear that HSP27 is a viable
target in cancer treatment and that a combined approach, with
either YZXJ or another method, would be a valid
consideration when devising a therapy in either targeting HSP27 or
in certain chemoresistance cases.
Acknowledgements
The authors wish to thank Cancer Research Wales,
National Research Network for Wales-Ser Cymru and the Albert Hung
Foundation for supporting their work.
References
|
1
|
Bonnycastle LL, Yu CE, Hunt CR, Trask BJ,
Clancy KP, Weber JL, Patterson D and Schellenberg GD: Cloning,
sequencing, and mapping of the human chromosome 14 heat shock
protein gene (HSPA2). Genomics. 23:85–93. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dix DJ, Allen JW, Collins BW, Mori C,
Nakamura N, Poorman-Allen P, Goulding EH and Eddy EM: Targeted gene
disruption of Hsp70-2 results in failed meiosis, germ cell
apoptosis, and male infertility. Proc Natl Acad Sci USA.
93:3264–3268. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bowen-Jones D, Fantes J and Gupta R:
Diabetes and heat shock protein. Nature. 355:119–120. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Keyse SM and Emslie EA: Oxidative stress
and heat shock induce a human gene encoding a protein-tyrosine
phosphatase. Nature. 359:644–647. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ackerley S, James PA, Kalli A, French S,
Davies KE and Talbot K: A mutation in the small heat-shock protein
HSPB1 leading to distal hereditary motor neuronopathy disrupts
neurofilament assembly and the axonal transport of specific
cellular cargoes. Hum Mol Genet. 15:347–354. 2006. View Article : Google Scholar
|
|
6
|
Evgrafov OV, Mersiyanova I, Irobi J, Van
Den Bosch L, Dierick I, Leung CL, Schagina O, Verpoorten N, Van
Impe K, Fedotov V, et al: Mutant small heat-shock protein 27 causes
axonal Charcot-Marie-Tooth disease and distal hereditary motor
neuropathy. Nat Genet. 36:602–606. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hickey E, Brandon SE, Potter R, Stein G,
Stein J and Weber LA: Sequence and organization of genes encoding
the human 27 kDa heat shock protein. Nucleic Acids Res.
14:4127–4145. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wyttenbach A, Sauvageot O, Carmichael J,
Diaz-Latoud C, Arrigo AP and Rubinsztein DC: Heat shock protein 27
prevents cellular polyglutamine toxicity and suppresses the
increase of reactive oxygen species caused by huntingtin. Hum Mol
Genet. 11:1137–1151. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Erkizan O, Kirkali G, Yörükoğlu K and
Kirkali Z: Significance of heat shock protein-27 expression in
patients with renal cell carcinoma. Urology. 64:474–478. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Langdon SP, Rabiasz GJ, Hirst GL, King RJ,
Hawkins RA, Smyth JF and Miller WR: Expression of the heat shock
protein HSP27 in human ovarian cancer. Clin Cancer Res.
1:1603–1609. 1995.PubMed/NCBI
|
|
11
|
Whelan RD and Hill BT: Differential
expression of steroid receptors, hsp27, and pS2 in a series of drug
resistant human breast tumor cell lines derived following exposure
to antitumor drugs or to fractionated X-irradiation. Breast Cancer
Res Treat. 26:23–39. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zimmermann M, Nickl S, Lambers C, Hacker
S, Mitterbauer A, Hoetzenecker K, Rozsas A, Ostoros G, Laszlo V,
Hofbauer H, et al: Discrimination of clinical stages in non-small
cell lung cancer patients by serum HSP27 and HSP70: A
multi-institutional case-control study. Clin Chim Acta.
413:1115–1120. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Giaginis C, Daskalopoulou SS, Vgenopoulou
S, Sfiniadakis I, Kouraklis G and Theocharis SE: Heat Shock
Protein-27, -60 and -90 expression in gastric cancer: Association
with clinicopathological variables and patient survival. BMC
Gastroenterol. 9:142009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Garrido C, Brunet M, Didelot C, Zermati Y,
Schmitt E and Kroemer G: Heat shock proteins 27 and 70:
Anti-apoptotic proteins with tumorigenic properties. Cell Cycle.
5:2592–2601. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gibert B, Eckel B, Gonin V, Goldschneider
D, Fombonne J, Deux B, Mehlen P, Arrigo AP, Clézardin P and
Diaz-Latoud C: Targeting heat shock protein 27 (HspB1) interferes
with bone metastasis and tumour formation in vivo. Br J Cancer.
107:63–70. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schweiger T, Nikolowsky C, Starlinger P,
Traxler D, Zimmermann M, Birner P, Hegedüs B, Dome B, Bergmann M,
Mildner M, et al: Stromal expression of heat-shock protein 27 is
associated with worse clinical outcome in patients with colorectal
cancer lung metastases. PLoS One. 10:e01207242015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Andrieu C, Taieb D, Baylot V, Ettinger S,
Soubeyran P, De-Thonel A, Nelson C, Garrido C, So A, Fazli L, et
al: Heat shock protein 27 confers resistance to androgen ablation
and chemotherapy in prostate cancer cells through eIF4E. Oncogene.
29:1883–1896. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Musiani D, Konda JD, Pavan S, Torchiaro E,
Sassi F, Noghero A, Erriquez J, Perera T, Olivero M and Di Renzo
MF: Heat-shock protein 27 (HSP27, HSPB1) is up-regulated by MET
kinase inhibitors and confers resistance to MET-targeted therapy.
FASEB J. 28:4055–4067. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yamamoto K, Okamoto A, Isonishi S, Ochiai
K and Ohtake Y: Heat shock protein 27 was up-regulated in cisplatin
resistant human ovarian tumor cell line and associated with the
cisplatin resistance. Cancer Lett. 168:173–181. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kuramitsu Y, Wang Y, Taba K, Suenaga S,
Ryozawa S, Kaino S, Sakaida I and Nakamura K: Heat-shock protein 27
plays the key role in gemcitabine-resistance of pancreatic cancer
cells. Anticancer Res. 32:2295–2299. 2012.PubMed/NCBI
|
|
21
|
Rogalla T, Ehrnsperger M, Preville X,
Kotlyarov A, Lutsch G, Ducasse C, Paul C, Wieske M, Arrigo AP,
Buchner J, et al: Regulation of Hsp27 oligomerization, chaperone
function, and protective activity against oxidative stress/tumor
necrosis factor alpha by phosphorylation. J Biol Chem.
274:18947–18956. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Charette SJ and Landry J: The interaction
of HSP27 with Daxx identifies a potential regulatory role of HSP27
in Fas-induced apoptosis. Ann N Y Acad Sci. 926:126–131. 2000.
View Article : Google Scholar
|
|
23
|
Charette SJ, Lavoie JN, Lambert H and
Landry J: Inhibition of Daxx-mediated apoptosis by heat shock
protein 27. Mol Cell Biol. 20:7602–7612. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kang D, Choi HJ, Kang S, Kim SY, Hwang YS,
Je S, Han Z, Kim JH and Song JJ: Ratio of phosphorylated HSP27 to
nonphosphorylated HSP27 biphasically acts as a determinant of
cellular fate in gemcitabine-resistant pancreatic cancer cells.
Cell Signal. 27:807–817. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nakashima M, Adachi S, Yasuda I, Yamauchi
T, Kawaguchi J, Itani M, Yoshioka T, Matsushima-Nishiwaki R, Hirose
Y, Kozawa O, et al: Phosphorylation status of heat shock protein 27
plays a key role in gemcitabine-induced apoptosis of pancreatic
cancer cells. Cancer Lett. 313:218–225. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ciocca DR and Calderwood SK: Heat shock
proteins in cancer: Diagnostic, prognostic, predictive, and
treatment implications. Cell Stress Chaperones. 10:86–103. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Baylot V, Andrieu C, Katsogiannou M, Taieb
D, Garcia S, Giusiano S, Acunzo J, Iovanna J, Gleave M, Garrido C,
et al: OGX-427 inhibits tumor progression and enhances gemcitabine
chemotherapy in pancreatic cancer. Cell Death Dis. 2:e2212011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kamada M, So A, Muramaki M, Rocchi P,
Beraldi E and Gleave M: Hsp27 knockdown using nucleotide-based
therapies inhibit tumor growth and enhance chemotherapy in human
bladder cancer cells. Mol Cancer Ther. 6:299–308. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lelj-Garolla B, Kumano M, Beraldi E, Nappi
L, Rocchi P, Ionescu DN, Fazli L, Zoubeidi A and Gleave ME: Hsp27
Inhibition with OGX-427 sensitizes non-small cell lung cancer cells
to erlotinib and chemotherapy. Mol Cancer Ther. 14:1107–1116. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Matsui Y, Hadaschik BA, Fazli L, Andersen
RJ, Gleave ME and So AI: Intravesical combination treatment with
antisense oligonucleotides targeting heat shock protein-27 and
HTI-286 as a novel strategy for high-grade bladder cancer. Mol
Cancer Ther. 8:2402–2411. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li Sun JR: Yei Tan, Xuedong Gao: Yang
capsule in treatment of hepatocellular carcinoma with
interventional chemotherapy random double-blind, multicenter
clinical study. World Chin J. 8:688–691. 2013.
|
|
32
|
Ligan Xing XZ, Li G, Zhang X, Sun X, Guo Q
and Yu J: Controlled clinical studies in the effects of Yang
capsule in advanced lung cancer in combination with chemotherapy.
China J Cancer Prev Treat. 21:384–386. 2014.
|
|
33
|
Xingjun Cui WM and Bi X: Effect of
Yangzheng Xiaoji capsule on cellular immune function in patients
with advanced gastric cancer chemotherapy. Chin J Difficult
Complicated Cases. 10:703–704. 2011.
|
|
34
|
Xue Kan SF and Ji J: Meta-anlaysis of the
safety of Yangzheng Xiaoji capsule for the treatment of cancer and
precancerosis. Chin J Clin Oncol. 40:1318–1323. 2013.
|
|
35
|
Jiang WG, Ye L, Ji K, Frewer N, Ji J and
Mason MD: Inhibitory effects of Yangzheng Xiaoji on angiogenesis
and the role of the focal adhesion kinase pathway. Int J Oncol.
41:1635–1642. 2012.PubMed/NCBI
|
|
36
|
Ye L, Ji K, Frewer N, Ji J and Jiang WG:
Impact of Yangzheng Xiaoji on the adhesion and migration of human
cancer cells: The role of the AKT signalling pathway. Anticancer
Res. 32:2537–2543. 2012.PubMed/NCBI
|
|
37
|
Guo Y, Ziesch A, Hocke S, Kampmann E, Ochs
S, De Toni EN, Göke B and Gallmeier E: Overexpression of heat shock
protein 27 (HSP27) increases gemcitabine sensitivity in pancreatic
cancer cells through S-phase arrest and apoptosis. J Cell Mol Med.
19:340–350. 2015. View Article : Google Scholar :
|
|
38
|
Rocchi P, So A, Kojima S, Signaevsky M,
Beraldi E, Fazli L, Hurtado-Coll A, Yamanaka K and Gleave M: Heat
shock protein 27 increases after androgen ablation and plays a
cytoprotective role in hormone-refractory prostate cancer. Cancer
Res. 64:6595–6602. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Choi DH, Ha JS, Lee WH, Song JK, Kim GY,
Park JH, Cha HJ, Lee BJ and Park JW: Heat shock protein 27 is
associated with irinotecan resistance in human colorectal cancer
cells. FEBS Lett. 581:1649–1656. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rashmi R, Santhosh Kumar TR and
Karunagaran D: Human colon cancer cells differ in their sensitivity
to curcumin-induced apoptosis and heat shock protects them by
inhibiting the release of apoptosis-inducing factor and caspases.
FEBS Lett. 538:19–24. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Belkacemi L and Hebb MO: HSP27 knockdown
produces synergistic induction of apoptosis by HSP90 and kinase
inhibitors in glioblastoma multiforme. Anticancer Res.
34:4915–4927. 2014.PubMed/NCBI
|
|
42
|
Jakubowicz-Gil J, Langner E, Bądziul D,
Wertel I and Rzeski W: Silencing of Hsp27 and Hsp72 in glioma cells
as a tool for programmed cell death induction upon temozolomide and
quercetin treatment. Toxicol Appl Pharmacol. 273:580–589. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jiang WG, Ye L, Ruge F, Owen S, Martin T,
Sun PH, Sanders AJ, Lane J, Satherley L, Weeks HP, et al: Yangzheng
Xiaoji exerts anti-tumour growth effects by antagonising the
effects of HGF and its receptor, cMET, in human lung cancer cells.
J Transl Med. 13:2802015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Shiota M, Bishop JL, Nip KM, Zardan A,
Takeuchi A, Cordonnier T, Beraldi E, Bazov J, Fazli L, Chi K, et
al: Hsp27 regulates epithelial mesenchymal transition, metastasis,
and circulating tumor cells in prostate cancer. Cancer Res.
73:3109–3119. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wettstein G, Bellaye PS, Kolb M, Hammann
A, Crestani B, Soler P, Marchal-Somme J, Hazoume A, Gauldie J,
Gunther A, et al: Inhibition of HSP27 blocks fibrosis development
and EMT features by promoting Snail degradation. FASEB J.
27:1549–1560. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cordonnier T, Bishop JL, Shiota M, Nip KM,
Thaper D, Vahid S, Heroux D, Gleave M and Zoubeidi A: Hsp27
regulates EGF/β-catenin mediated epithelial to mesenchymal
transition in prostate cancer. Int J Cancer. 136:E496–E507. 2015.
View Article : Google Scholar
|
|
47
|
Voll EA, Ogden IM, Pavese JM, Huang X, Xu
L, Jovanovic BD and Bergan RC: Heat shock protein 27 regulates
human prostate cancer cell motility and metastatic progression.
Oncotarget. 5:2648–2663. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Thuringer D, Jego G, Wettstein G, Terrier
O, Cronier L, Yousfi N, Hébrard S, Bouchot A, Hazoumé A, Joly AL,
et al: Extracellular HSP27 mediates angiogenesis through Toll-like
receptor 3. FASEB J. 27:4169–4183. 2013. View Article : Google Scholar : PubMed/NCBI
|