Introduction
Bladder cancer is the most common cancer of urinary
tract with an estimation of approximately 80,500 new cases and
32,900 deaths and non-uniform distribution among urban and rural
areas in China in 2015 (1).
Epidemiologic studies revealed that cigarette smoking, exposure to
arsenic and occupational exposure to aromatic amines are risk
factors of bladder cancer (2).
Benzidine, also called 4,4′-diaminobiphenyl, is an
aromatic amine, which has been classified as definite human
carcinogen by the International Agency for Research on Cancer
(IARC) with urinary bladder being a major target (3–5). Due
to its wide distribution in the chemical, dye and rubber
industries, cigarette smoke, automobile exhaust and industrial
pollutant emissions, benzidine still is a great hazard to human
health (6–8). Notwithstanding, there have been
epidemiological research revealing the correlation between
benzidine exposure and bladder oncogenesis, the molecular
pathogenesis is still largely unknown.
Epithelial-mesenchymal transition (EMT) is a
pathophysiological process that involves in multiple processes
including embryogenesis, tissue reconstruction, organ fibrosis and
cancer development. Epithelia lose epithelial characteristics and
obtain mesenchymal traits via EMT, such as loss of cell polarity,
enhancement of cell invasive and metastatic ability (9–11).
Abundant evidence demonstrates the crucial role of EMT in the
invasion and metastasis of cancer cells, including bladder cancer.
Benzidine exposure has been manifested to induce EMT in urinary
bladder (12). However, the
molecular mechanism of benzidine-induced EMT remains insufficiently
illuminated.
The mitogen-activated protein kinases (MAPKs)
include four major subfamilies: extracellular signal-related
kinases (ERK 1/2), Jun amino-terminal kinases (JNK1/2/3), p38
proteins (p38α/β/γ/δ) and ERK5 (13). ERK5 (also called big
mitogen-activated protein kinase 1) (BMK1), is the least studied
member of MAPK family. The MEK/ERK5 is involved in multiple
important processes including cell differentiation and survival,
cell proliferation, angiogenesis and tumorigenesis (14–17).
Activated protein 1 (AP-1) is a heterodimer composed of Jun and Fos
proteins. As a transcription activator, AP-1 acts in various cell
activities. Compelling evidence shows that the mitogen activated
protein kinase (MAPK)/AP-1 pathway plays an important role in
cancer initiation and progression (18,19).
In a previous study, our groups revealed that activation of
ERK5/AP-1 upregulates cigarette smoke induced EMT of normal
urothelial cells (20), but
investigations have not been made to detect the correlation between
ERK5/AP-1 and benzidine exposure, and the role of ERK5 in
benzidine-provoked EMT remains to be elucidated.
Curcumin is a polyphenol
(1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione) with
an outstanding safety profile and is the major bioactive component
of turmeric, a spice commonly used as a food additive and a
traditional herbal medicine with a long history throughout Asia
(21). Former research
demonstrated the anticancer activities of curcumin derivatives on a
variety of cancers including lung, breast, colorectal and thyroid
cancer in vitro and in vivo. However, few studies
have been conducted to detect the interventional activities of
curcumin on benzidine-related bladder EMT.
The aim of present study was to investigate the
mechanism of benzidine induced malignancy in human normal
urothelial cells as well as chemical preventive effect of curcumin
on this process, which may provide new insight into
benzidine-related carcinogenesis and cancer intervention.
Materials and methods
Chemicals and reagents
SV-40 immortalized human urothelial cell line
(SV-HUC-1) was obtained from the Chinese Academy of Type Culture
Collection Cell Bank (Shanghai, China). Curcumin was purchased from
Sigma-Aldrich (St. Louis, MO, USA; purity, 99.0%). Benzidine
(4,4′-diaminobiphenyl; ≥98.0%, RT), dimethyl sulfoxide (DMSO) and
methanol were obtained from Merck (Reading Township, NJ, USA).
Growth media (F-12K medium, Kaighn's Medium; Wisent Inc., Montreal,
QC, Canada). Fetal bovine serum (FBS), phosphate-buffered saline
(PBS), antibiotics and trypsin were obtained from HyClone
Laboratories (Logan, UT, USA).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
was purchased from Sigma-Aldrich. Phosphorylated ERK5 (p-ERK5),
phosphorylated c-jun (p-c- jun), phosphorylated c-fos (p-c-fos),
E-cadherin, N-cadherin and vimentin were obtained from Cell
Signaling Technology (Beverly, MA, USA). The antibody for ZO-1 was
obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The
GAPDH antibody was obtained from Biogot Technology (Nanjing,
China). XMD8-92 was purchased from Selleck Chemicals (Huston, TX,
USA). Sources of other materials are noted accordingly in the
text.
Cell culture and treatment
SV-HUC-1 cells were cultivated with F-12K growth
medium with addition of 10% FBS, 100 U/ml penicillin and 100
μg/ml streptomycin under the atmosphere of 5% CO2
at 37°C in an incubator. Cells were seeded in 10-cm2
culture plates. The medium was changed every day until cells
reached 80–90% confluence, and then was treated with various
concentrations of benzidine, XMD8-92 or curcumin.
Cell toxicity assay
SV-HUC-1 cells were seeded in 96-well plate at the
density of 2×103 cells/well. Then, the medium mixed with
benzidine or curcumin at different concentrations was used to
cultivate cells and changed every day for 4 days (12,22,23).
Ten microliters of methylthiazoletetrazolium assay solution (5
mg/ml) was added to each well and the plates were further incubated
for 4 h at 37°C. Medium containing MTT was removed and the
precipitants were solubilized in DMSO. Absorbance was measured at
490 nm using a microplate reader (Titertek Instruments, Incorp.,
Huntsville, AL, USA). All measurements were performed in
triplicate.
Migration assay
Wound healing assay was applied to detect change of
migratory ability. Medium without benzidine or within benzidine at
concentration of 0.005 or 0.05 μM was used for pretreating
SV-HUC-1 cells for 4 days. Then, cells were seeded in 6-well plate
at 4×105/well and cultivated for 24 h with serum-free
medium before wounding. Afterwards, manual scrape was made to the
cell monolayer to create a wound with a pipette tip. After washing
the plate with PBS, each well was covered with medium to acquire
images at the time-points of 0, 12, 24 and 36 h.
Invasion assay
The invasion assays were performed in a 24-well
Boyden chamber with an 8-μm pore size polycarbonate membrane
(Millipore, Billerica, MA, USA) coated with Matrigel to form a
matrix barrier. Pretreated cells without or with benzidine (0.005
and 0.05 μM) for 4 days, were suspended and 1×104
cells in 150 μl serum-free medium were added to the upper
compartment of the chamber. Moreover, 800 μl F-12K medium
supplemented with 10% FBS was added into the lower compartment.
After culturing in the incubator for 48 h, cells remaining on the
membranes inside the chamber were wiped off with a cotton-tipped
swab, while cells adhered to lower membranes were fixed with
methanol stained with crystal violet and finally photographed under
a microscope. Then the number of invaded cells per microscope field
was counted and analyzed.
Western blot analysis
SV-HUC-1 cells were harvest after a 4-day culture.
Cells were washed with cryopreserved PBS and lysed with RIPA buffer
(Thermo Fisher Scientific, Waltham, MA, USA). Concentrations of
extracted proteins were measured with BCA protein assay (Pierce,
Rockford, IL, USA). Then, the proteins were diluted to equal
concentrations, boiled for 5 min and separated by 7.5–10% SDS-PAGE,
transferred onto polyvinylidene difluoride membranes (Millipore).
Afterwards, membranes were blocked in 5% fat-free dry milk in
Tween-20 Tris-buffered saline (TBST) and then incubated in primary
antibodies and secondary antibodies. Protein bands were
subsequently developed using High-sig ECL western blotting
substrate kit (Tanon Science and Technology, Co., Ltd., Shanghai,
China). GAPDH served as loading control.
Quantitative real-time PCR
Total RNA was isolated by RNAiso Plus according to
the manufacturer's instructions. (Takara Bio, Shiga, Japan). Then,
total RNA was transcribed into cDNA using the AMV reverse
transcriptase (Takara) following the manufacturer's protocol.
qRT-PCR was performed using the Power SYBR-Green Master Mix
(Takara) and an ABI 7300 Real-Time PCR Detection system (Applied
Biosystems, Foster City, CA, USA). The primers used were as
follows: E-cadherin, forward, 5′-TCGACACCCGATTCAAAGTGG-3′ and
reverse, 5′-TTCCAGAAACGGAGGCCTGAT-3′; ZO-1, forward,
5′-GCAGCCACAACCAATTCATAG-3′ and reverse,
5′-GCAGACGATGTTCATAGTTTC-3′; vimentin, forward,
5′-CCTTGACATTGAGATTGCCA-3′ and reverse, 5′-GTATCAACCAGAGGGAGTGA-3′;
N-cadherin forward, 5′-ATCAAGTGCCATTAGCCAAG-3′ and reverse,
5′-CTGAGCAGTGAATGTTGTCA-3′; and GAPDH, forward,
5′-GCTGCCCAACGCACCGAATA-3′ and reverse, 5′-GAGTCAACGGATTTGGTCGT-3′.
All of the primers were synthesized by Invitrogen (Carlsbad, CA,
USA). The levels of mRNA expression for each gene were normalized
by its respective GAPDH. Fold changes in gene expression were
calculated by a comparative threshold cycle (Ct) method using the
formula 2−(ΔΔCt).
Immunofluorescence
Pretreated SV-HUC-1 cells were transferred into
35-mm Glass bottom dishes (Cellvis, Mountain View, CA, USA) at the
density of 5×104/dish. After fixing and washing, dishes
were stained with rabbit E-cadherin (1:150 dilution) and vimentin
(1:100 dilution) antibodies at 4°C overnight and then incubated
with fluorescein isothiocyanate (FITC)-conjugated goat anti-rabbit
secondary antibody for 1 h. To stain the nuclei,
4′,6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich) was added for
10 min. Images were captured using fluorescence microscopy (Zeiss,
LSM700B; Carl Zeiss AG, Germany).
Statistical analysis
Statistical analyses were performed with the SPSS
16.0. All data were expressed as mean ± standard deviation (SD).
One-way ANOVA was used for comparison of statistical differences
among multiple groups, followed by the LSD significant difference
test. Unpaired Student's t-test was also used for the comparison
between the two groups. P-values of P<0.05 or P<0.01 were
considered significantly different.
Results
Benzidine induces EMT in SV cells
Occupational exposure to aromatic amine is a major
cause of bladder cancer, and benzidine-induced EMT is important in
benzidine-associated malignant generation. To investigate the cell
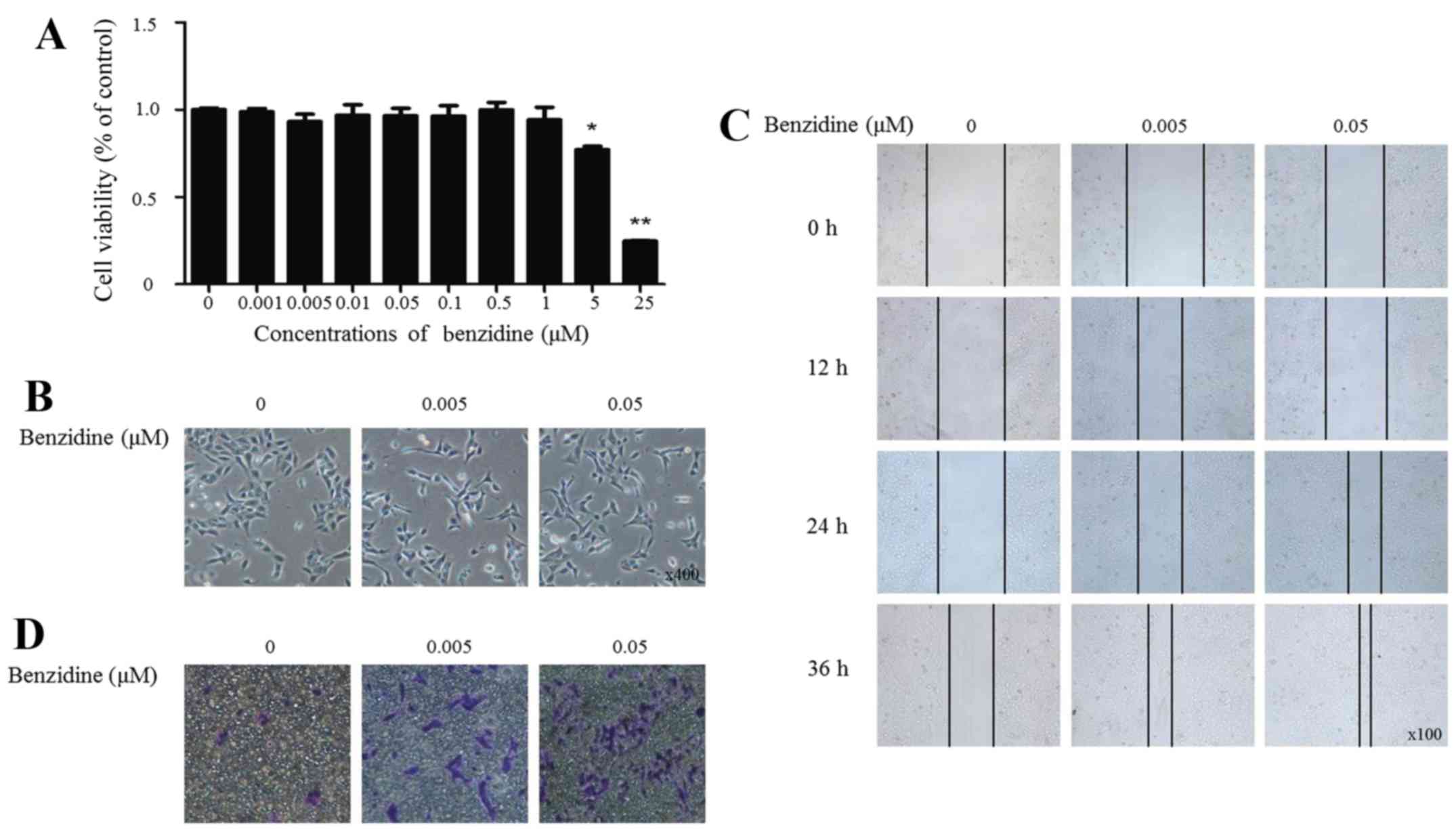
viability change in this process, cells were treated with benzidine
(0.001, 0.005, 0.01, 0.05, 0.1, 1, 5, 25 and 100 μM) for 4
days and were detected by MTT assay. The results showed that the
cell viability decreased <80% when the cells were exposed to 25
μM benzidine or to higher concentrations (Fig. 1A). Therefore, concentrations range
from 0.001 to 0.1 μM were selected for the following
experiments.
EMT process was assessed by alterations in the cell
morphology, migratory and invasive capacity, and expression levels
of epithelial and mesenchymal markers. Benzidine induced
morphological change from epithelial to spindle-like mesenchymal
shape, as shown by morphological examination of SV-HUC-1 cells
following benzidine treatment for 4 days (Fig. 1B). Migratory ability was enhanced
as wound healed in the treated groups compared with the control
group (Fig. 1C). In invasion
assay, cells transferred through the Matrigel at about the number
of 10 per field in the control group. While in treated groups,
cells increased to about 12 and 30 times, respectively, indicating
bezidine induced elevated invasive capacity (Fig. 1D).
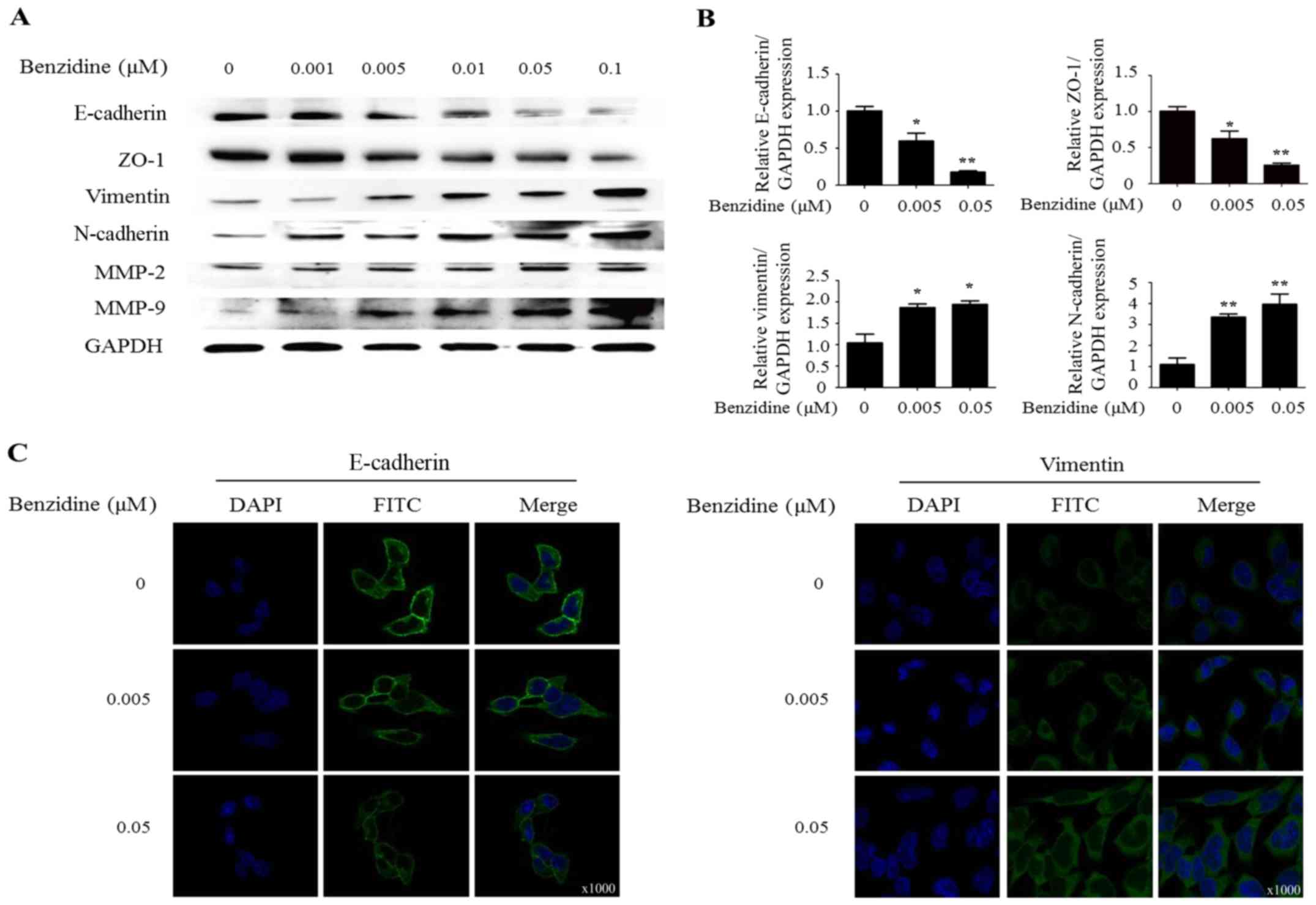
Occurrence of molecular changes
Western blot analysis exhibited the decrease of
protein levels of epithelial markers E-cadherin and ZO-1. On the
contrary, benzidine exposure enhanced the protein expression of
mesenchymal markers vimentin, N-cadherin, MMP-2 and MMP-9 (Fig. 2A). Subsequent RT-PCT of mRNA levels
of epithelial and mesenchymal markers showed similar changes.
Expression levels of E-cadherin mRNA at 0.005 and 0.05 were 55.3%
(P<0.05), 24.1% (P<0.01). In addition, vimentin mRNA levels
at 0.005 and 0.05 were 186.3% (P<0.05) and 194.2% (P<0.05)
(Fig. 2B). Results of
immunofluorescent staining manifested changes not only in
morphology but also in protein levels of epithelial and mesenchymal
markers as described (Fig. 2C).
Taken together, our results suggested that benzidine could induce
EMT in SV-HUC-1 cells.
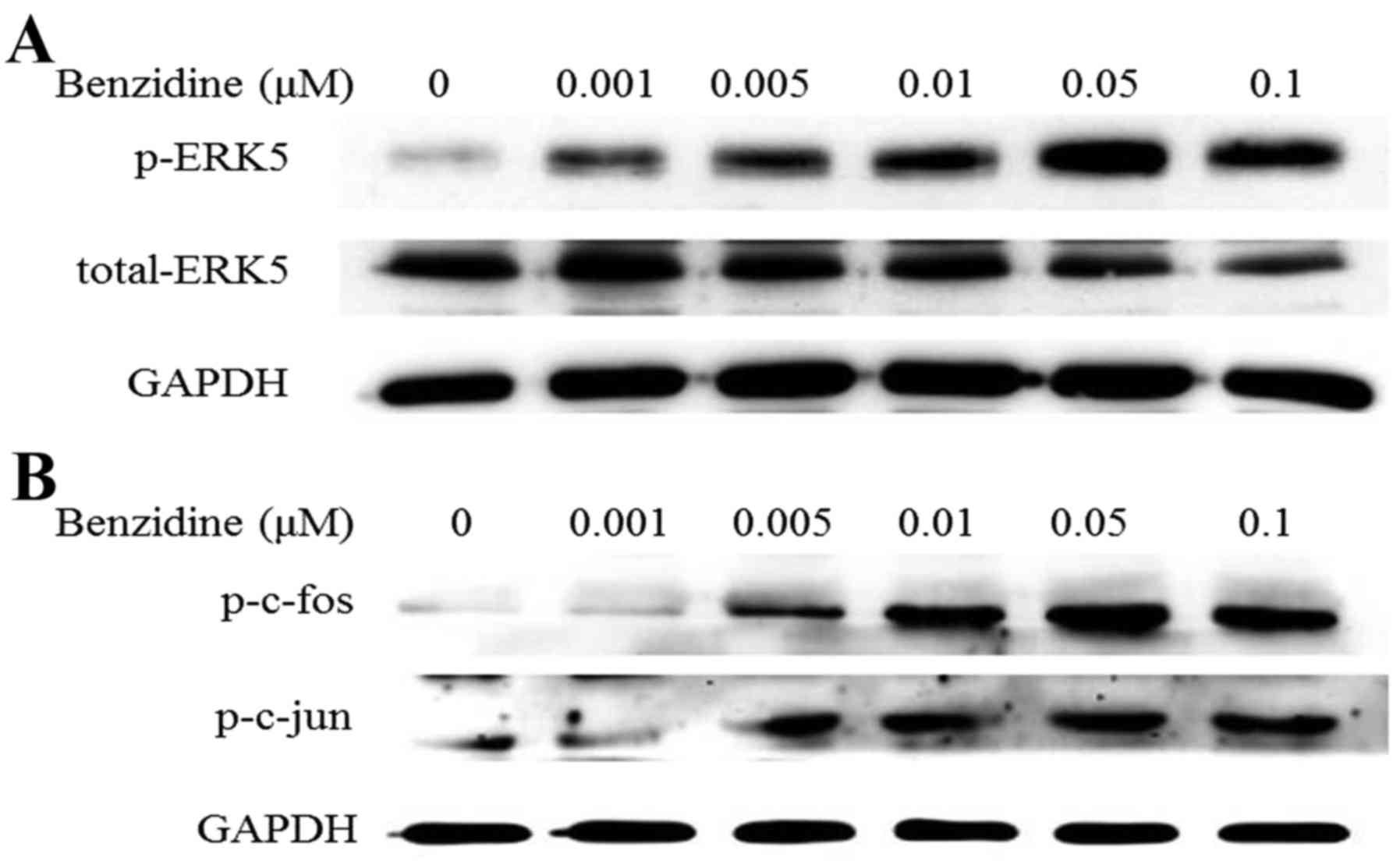
Benzidine activates ERK5/AP-1 signal
pathway
ERK5 is a member of MAPKs family, which was thought
to be onco- signaling. AP-1 is a downstream transcriptional factor
of ERK5. To detect whether ERK5/AP-1 pathway was activated after
SV-HUC-1 cells were exposed to benzidine for 4 days, protein levels
of ERK5/AP-1 markers, p-ERK5, p-c-jun and p-c-fos were measured.
The results showed notable increasing of p-ERK5 with dose-dependent
manner and simultaneously restrained the level of total ERK5,
indicating the upregulation effect of benzidine on ERK5 activity
(Fig. 3A). Moreover, benzidine
increased protein levels of p-c-jun and p-c-fos (Fig. 3B).
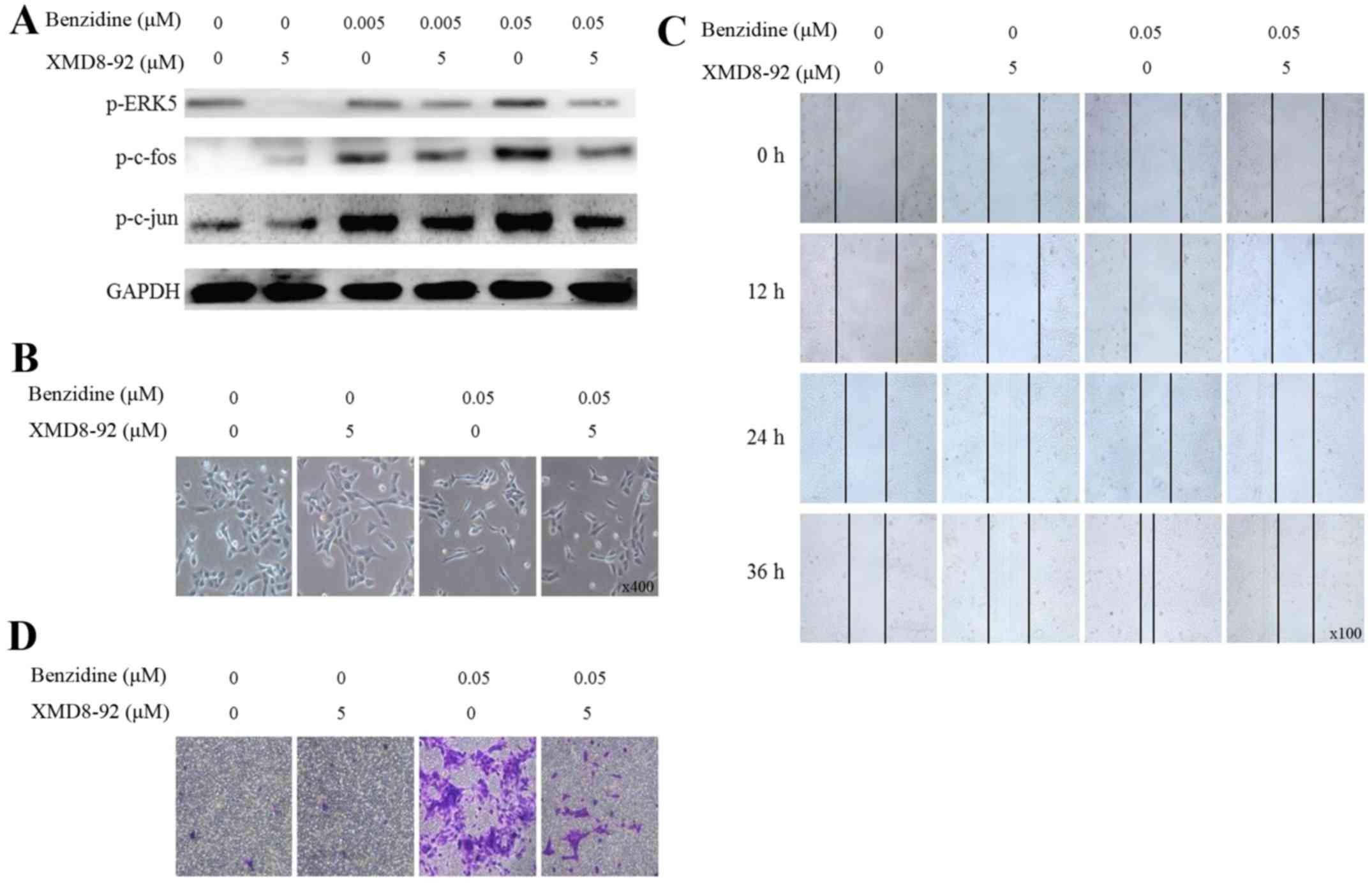
ERK5/AP-1 positively regulates
benzidine-induced EMT
To further explore the correlation between
benzidine-induced EMT and activation of ERK5/AP-1, XMD8-92 (5
μM), a highly specific ERK5 inhibitor was used. Consistent
with previous research, we found the inhibitor significantly
suppressed the ERK5/AP-1 activity (Fig. 4A). Then we observed effects of
XMD8-92 on benzidine-induced EMT process. Results showed that
inhibitor alone had no effect on cell phenotype, while
benzidine-induced SV-HUC-1 morphological changes were reversed
after cells were exposed to benzidine combined with the inhibitor
(Fig. 4B). Inhibitor resulted in
attenuation of benzidine-enhanced cell migration and invasion
ability (Fig. 4C and D).
Moreover, molecular changes were detected. Results
indicated that suppression of ERK5 activity leads to downregulation
of epithelial markers E-cadherin, ZO-1 as well as upregulation of
mesenchymal markers vimentin and N-cadherin in protein and mRNA
levels (Fig. 4E and F).
Collectively, our results indicated that benzidine promoted
SV-HUC-1 EMT via upregulating the ERK5/AP-1 signal pathway.
Curcumin attenuates benzidine-induced EMT
via suppression of ERK5/AP-1
To determine whether curcumin could interfere with
benzidine-induced EMT, we first conducted MTT assay to screen
appropriate concentration for subsequent experiments. The data
showed that curcumin at doses <1 μM had no influence on
cell viability, and cell viability was decreased <85% at
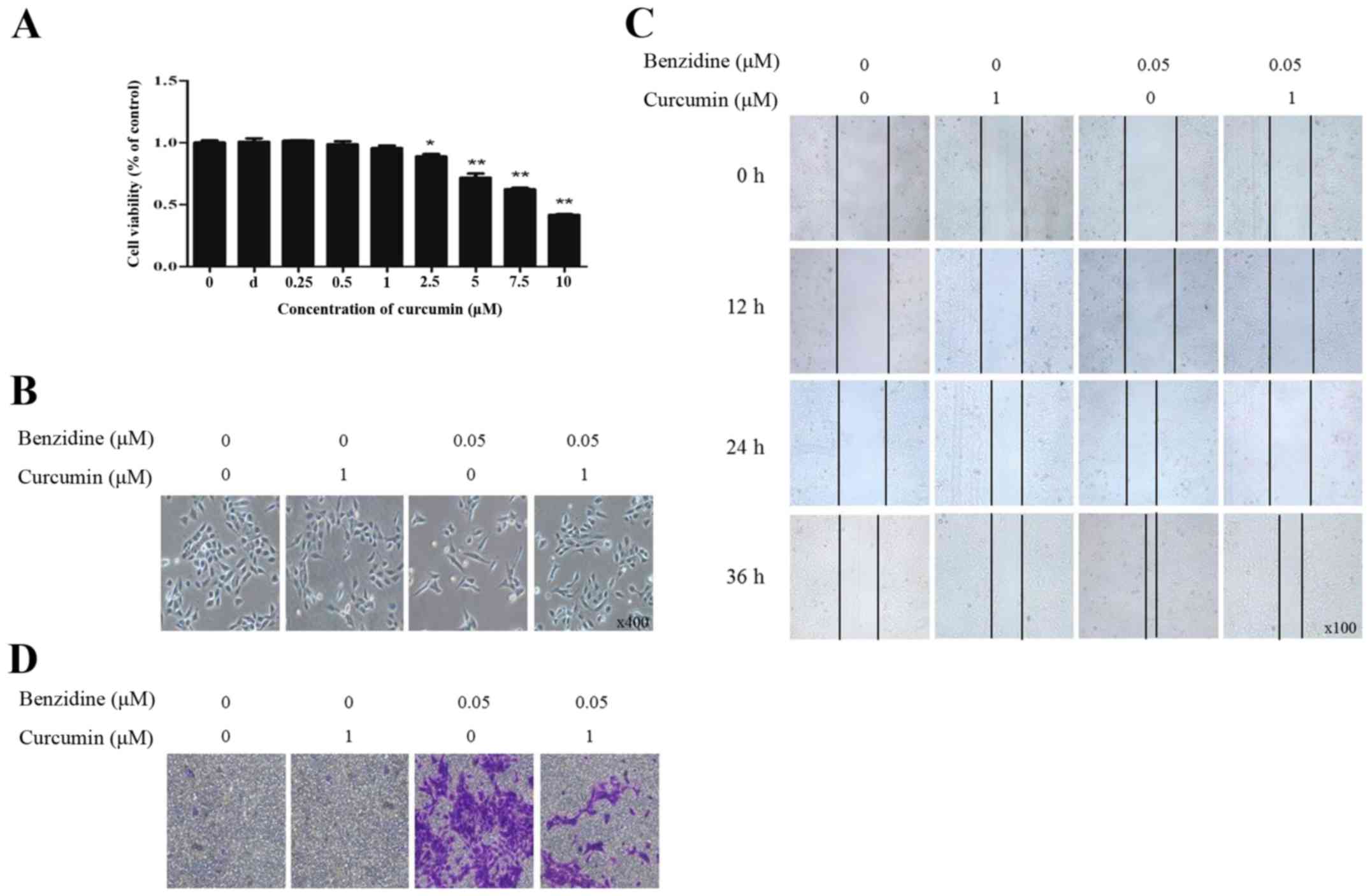
concentrations >2.5 μM and had significance (Fig. 5A). Therefore, curcumin at 1
μM was chosen for the following experiments. Results showed
that curcumin alone had no effect on cell morphology. The
benzidine-mediated cell morphological changes were reversed by
curcumin (Fig. 5B).
Benzidine-enhanced capacity of migration and invasion were also
detected to be weakened by curcumin (Fig. 5C and D).
Subsequently, we measured epithelium and mesenchymal
markers at molecular levels. Results indicated that treatment with
curcumin elevated protein and mRNA levels of E-cadherin and ZO-1,
reduced vimentin and N-cadherin, which demonstrated the suppression
of curcumin in benzidine-induced EMT (Fig. 5E and F). Furthermore, we detected
ERK5/AP-1 activity as cells exposed to curcumin and found that
benzidine- activated ERK5/AP-1 was inhibited by curcumin (Fig. 5G). In summary, these findings
confirmed that curcumin was able to reverse benzidine-triggered
SV-HUC-1 cell EMT via suppression of the ERK5/AP-1 signal
pathway.
Discussion
Occupational exposure to aromatic amine including
benzidine has been manifested as a major cause of bladder cancer.
However, the underlying mechanisms by which benzidine results in
tumorigenesis remain to be elucidated. Results of a previous study
revealed the important role of ERK1/2 in benzidine-triggered EMT
(12), whereas the role of ERK5 in
benzidine-promoted bladder cancer development has not been
investigated. In addition, the chemical prevention of
benzidine-induced malignancy has not been explored. In the present
study, we demonstrated the regulation role of ERK5/AP-1 in
benzidine-induced EMT in SV-HUC-1 cells. Moreover, we identified
the prevention effect of curcumin on benzidine-mediated EMT. These
finding may provide new insight into benzidine-related tumor
formation, as well as chemical intervention for bladder cancer.
Characteristics by alteration in cell morphology,
migratory and invasive capacity, as well as expression of
epithelial and mesenchymal markers, EMT is considered as a critical
step for the initiation and development of epithelium-originated
malignancy. It has been documented that many molecular changes
occur during EMT process, including downregulation of E-cadherin
and upregulation of N-cadherin. E-cadherin has been established as
a cell adhesion molecule, which plays an essential role in
epithelial cell-to-cell interactions since it mediates the
connections between adjacent epithelial cells and maintains the
phenotype and apical-base polarity of epithelial cells. N-cadherin
(neural cadherin), another adhesion molecule, is associated with a
heightened migratory and invasive potential in cancer (24,25).
Consistent with former studies, the results of the present study
showed that benzidine exposure induces EMT in SV-HUV-1 cell line
manifested by the cell phenotype and molecular changes after
benzidine exposure. Taken together, we identified benzidine induced
EMT in vitro.
Several signal pathways have been implicated in EMT,
such as PI3K, NF-κB and MAPK (26–28).
To date, scarce research has been conducted to explore the function
of ERK5 in benzidine-triggered EMT. In the present study, we found
that ERK5 was activated during benzidine exposure. We further
confirmed the regulation of ERK5 in urocystic EMT. ERK5 inhibitor
significantly reversed benzidine-induced EMT, as showed by repaired
cell shape, attenuated migratory and invasive capacity and
molecular changes. In summary, these results indicated the
upregulation role of ERK5 in benzidine-induced EMT.
ERK5 is twice the size of the other members. Similar
to ERK1/2, the N-terminal of ERK5 contains a kinase domain and has
the Thr-Glu-Tyr (TEY) activation motif. ERK5 differ in its large
C-terminal-half that contains a transcriptional activation domain.
Upon activation, ERK5 could not only phosphorylate and activate
downstream target molecules, including transcription factors such
as members of the AP-1 proteins, but also auto-phosphorylate
C-terminus, which alone has the ability to increase transcriptional
activity (29–31). Therefore, ERK5 is different from
other ERKs for possessing transcriptional activation activity.
Given its unique structure, ERK5 regulates the transcription of
downstream molecules in two ways i.e., through either the
phosphorylation or the enhancement of the transcription activity of
target molecules (14,15). It has been found that ERK5
transcriptionally regulates gene expression in a tissue-specific
manner (32). In breast cancer
cells, activation of ERK5 significantly enhance cyclin D1
expression and promotes neoplastic transformation (33). In lung microvascular endothelial
cells, activation of ERK5 suppresses HIF-1α expression and inhibits
angiogenesis (34). In this study,
we showed that benzidine-meditated activation of ERK5 triggered the
activation of AP-1 proteins c-Jun and c-Fos; on the contrary,
inhibition of ERK5 suppressed the activation of c-Jun and c-Fos,
indicating the positive modulation role of ERK5 on AP-1. Research
has proved the important role of AP-1 proteins in EMT modulation by
elevating EMT-related molecules and transcriptional factors such as
Zeb1/2, TGFβ, Twist1, integrin α5 and epithelial cell adhesion
molecule (EpCAM), which in turn transcriptionally regulate
expression of mesenchymal markers vimentin and N-cadherin, and
epithelial markers E-cadherin and ZO-1 (35–38).
In the present study, we found that benzidine activated ERK5,
leading to activation of AP-1, resulting in promoted migratory and
invasive abilities, as well as decreased levels of epithelial
markers and increased levels of mesenchymal markers. In summary,
the present study demonstrated the positive role of ERK5/AP-1 in
benzidine-induced urocystic EMT.
As a promising natural compound, curcumin has been
applied to cancer therapy. Curcumin pleiotropic activities emanate
from its ability to modulate numerous signaling, transcriptional
molecules, signal transducer and adhesion molecules such as MMP-9,
intracellular adhesion molecule-1 (ICAM-1) and vascular cell
adhesion molecule-1 (VCAM-1) (39,40).
Evidence has confirmed that curcumin regulates different signal
pathways or cytokines to prevent cancer initiation and promotion.
It has been acknowledged that curcumin can be used as an efficient
adjuvant to cisplatin cancer therapy by modulating STAT3 and Nrf2
signaling in human squamous carcinoma (41). Woo et al (42) demonstrated that curcumin suppresses
MAPK signaling pathways in human astroglioma cells. As for the
present study, we proved that curcumin repairs benzidine-induced
EMT in both phenotype and genotype levels. Moreover,
benzidine-triggered activation of ERK5/AP-1 signal pathway was
attenuated by curcumin, indicating the interventional role of
curcumin in benzidine- meditated urocystic malignancies.
In conclusion, this study identified the positive
role of ERK5 in benzidine-promoted EMT, as well as the suppression
role of curcumin in benzidine-related bladder malignancy. These
findings indicate new mechanism of benzidine-induced bladder
malignancy and provide a potential strategy for prevention and
intervention of bladder cancer.
Acknowledgments
The present study was supported by grants from the
National Natural Science Foundation of China (nos. 81373005,
81072330 and 81202194), the Priority Academic Program Development
of Jiangsu Higher Education Institutions (Public Health and
Preventive Medicine), the Anhui Medical University Scientific
Research Funds (Hefei, China; grant no. H0514) and the Anhui Public
Welfare Research Linkage Plan (Hefei, China; grant no.
1501ld04045).
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Letašiová S, Medve'ová A, Šovčíková A,
Dusinská M, Volkovová K, Mosoiu C and Bartonová A: Bladder cancer,
a review of the environmental risk factors. Environ Health.
11(Suppl 1): S112012. View Article : Google Scholar
|
|
3
|
Choudhary G: Human health perspectives on
environmental exposure to benzidine: A review. Chemosphere.
32:267–291. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Whysner J, Verna L and Williams GM:
Benzidine mechanistic data and risk assessment: Species- and
organ-specific metabolic activation. Pharmacol Ther. 71:107–126.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baan R, Grosse Y, Straif K, Secretan B, El
Ghissassi F, Bouvard V, Benbrahim-Tallaa L, Guha N, Freeman C,
Galichet L, et al WHO International Agency for Research on Cancer:
Monograph Working Group: A review of human carcinogens–Part F:
Chemical agents and related occupations. Lancet Oncol.
10:1143–1144. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Snyderwine EG, Sinha R, Felton JS and
Ferguson LR: Highlights of the eighth international conference on
carcinogenic/mutagenic N-substituted aryl compounds. Mutat Res.
506–507:1–8. 2002. View Article : Google Scholar
|
|
7
|
Yu MC, Skipper PL, Tannenbaum SR, Chan KK
and Ross RK: Arylamine exposures and bladder cancer risk. Mutat
Res. 506–507:21–28. 2002. View Article : Google Scholar
|
|
8
|
Chen HI, Liou SH, Loh CH, Uang SN, Yu YC
and Shih TS: Bladder cancer screening and monitoring of
4,4′-methylenebis(2-chloroaniline) exposure among workers in
Taiwan. Urology. 66:305–310. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kalluri R: EMT: When epithelial cells
decide to become mesenchymal-like cells. J Clin Invest.
119:1417–1419. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li L and Li W: Epithelial-mesenchymal
transition in human cancer: Comprehensive reprogramming of
metabolism, epigenetics, and differentiation. Pharmacol Ther.
150:33–46. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Voulgari A and Pintzas A:
Epithelial-mesenchymal transition in cancer metastasis: Mechanisms,
markers and strategies to overcome drug resistance in the clinic.
Biochim Biophys Acta. 1796:75–90. 2009.PubMed/NCBI
|
|
12
|
Zhao L, Geng H, Liang ZF, Zhang ZQ, Zhang
T, Yu DX and Zhong CY: Benzidine induces epithelial-mesenchymal
transition in human uroepithelial cells through ERK1/2 pathway.
Biochem Biophys Res Commun. 459:643–649. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chang L and Karin M: Mammalian MAP kinase
signalling cascades. Nature. 410:37–40. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Drew BA, Burow ME and Beckman BS:
MEK5/ERK5 pathway: The first fifteen years. Biochim Biophys Acta.
1825:37–48. 2012.
|
|
15
|
Nishimoto S and Nishida E: MAPK
signalling: ERK5 versus ERK1/2. EMBO Rep. 7:782–786. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hayashi M, Fearns C, Eliceiri B, Yang Y
and Lee JD: Big mitogen-activated protein kinase 1/extracellular
signal-regulated kinase 5 signaling pathway is essential for
tumor-associated angiogenesis. Cancer Res. 65:7699–7706.
2005.PubMed/NCBI
|
|
17
|
Amano S, Chang YT and Fukui Y: ERK5
activation is essential for osteoclast differentiation. PLoS One.
10:e01250542015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhong CY, Zhou YM, Douglas GC, Witschi H
and Pinkerton KE: MAPK/AP-1 signal pathway in tobacco smoke-induced
cell proliferation and squamous metaplasia in the lungs of rats.
Carcinogenesis. 26:2187–2195. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao J, Harper R, Barchowsky A and Di YPP:
Identification of multiple MAPK-mediated transcription factors
regulated by tobacco smoke in airway epithelial cells. Am J Physiol
Lung Cell Mol Physiol. 293:L480–L490. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Geng H, Zhao L, Liang Z, Zhang Z, Xie D,
Bi L, Wang Y, Zhang T, Cheng L, Yu D, et al: ERK5 positively
regulates cigarette smoke- induced urocystic epithelial-mesenchymal
transition in SV-40 immortalized human urothelial cells. Oncol Rep.
34:1581–1588. 2015.PubMed/NCBI
|
|
21
|
Park W, Amin ARMR, Chen ZG and Shin DM:
New perspectives of curcumin in cancer prevention. Cancer Prev Res
(Phila). 6:387–400. 2013. View Article : Google Scholar
|
|
22
|
Kawami M, Harabayashi R, Miyamoto M,
Harada R, Yumoto R and Takano M: Methotrexate-induced
epithelial-mesenchymal transition in the alveolar epithelial cell
line A549. Lung. 194:923–930. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang P, Dong Z, Cai J, Zhang C, Shen Z,
Ke A, Gao D, Fan J and Shi G: BRD4 promotes tumor growth and
epithelialmesenchymal transition in hepatocellular carcinoma. Int J
Immunopathol Pharmacol. 28:36–44. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Peralta Soler A, Knudsen KA, Jaurand MC,
Johnson KR, Wheelock MJ, Klein-Szanto AJ and Salazar H: The
differential expression of N-cadherin and E-cadherin distinguishes
pleural mesotheliomas from lung adenocarcinomas. Hum Pathol.
26:1363–1369. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nakajima S, Doi R, Toyoda E, Tsuji S, Wada
M, Koizumi M, Tulachan SS, Ito D, Kami K, Mori T, et al: N-cadherin
expression and epithelial-mesenchymal transition in pancreatic
carcinoma. Clin Cancer Res. 10:4125–4133. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang X, Song Q, Wei C and Qu J: LRIG1
inhibits hypoxia- induced vasculogenic mimicry formation via
suppression of the EGFR/PI3K/AKT pathway and
epithelial-to-mesenchymal transition in human glioma SHG-44 cells.
Cell Stress Chaperones. 20:631–641. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li J, Deng Z, Wang Z, Wang D, Zhang L, Su
Q, Lai Y, Li B, Luo Z, Chen X, et al: Zipper-interacting protein
kinase promotes epithelial-mesenchymal transition, invasion and
metastasis through AKT and NF-κB signaling and is associated with
metastasis and poor prognosis in gastric cancer patients.
Oncotarget. 6:8323–8338. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Park JH, Yoon J, Lee KY and Park B:
Effects of geniposide on hepatocytes undergoing
epithelial-mesenchymal transition in hepatic fibrosis by targeting
TGFβ/Smad and ERK-MAPK signaling pathways. Biochimie. 113:26–34.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kasler HG, Victoria J, Duramad O and
Winoto A: ERK5 is a novel type of mitogen-activated protein kinase
containing a transcriptional activation domain. Mol Cell biol.
20:8382–8389. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Morimoto H, Kondoh K, Nishimoto S,
Terasawa K and Nishida E: Activation of a C-terminal
transcriptional activation domain of ERK5 by autophosphorylation. J
biol Chem. 282:35449–35456. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nithianandarajah-Jones GN, Wilm B,
Goldring CE, Müller J and Cross MJ: ERK5: Structure, regulation and
function. Cell Signal. 24:2187–2196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sohn SJ, Li D, Lee LK and Winoto A:
Transcriptional regulation of tissue-specific genes by the ERK5
mitogen-activated protein kinase. Mol Cell Biol. 25:8553–8566.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mulloy R, Salinas S, Philips A and
Hipskind RA: Activation of cyclin D1 expression by the ERK5
cascade. Oncogene. 22:5387–5398. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pi X, Garin G, Xie L, Zheng Q, Wei H, Abe
J, Yan C and Berk BC: BMK1/ERK5 is a novel regulator of
angiogenesis by destabilizing hypoxia inducible factor 1alpha. Circ
Res. 96:1145–1151. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bakiri L, Macho-Maschler S, Custic I,
Niemiec J, Guío- Carrión A, Hasenfuss SC, Eger A, Müller M, Beug H
and Wagner EF: Fra-1/AP-1 induces EMT in mammary epithelial cells
by modulating Zeb1/2 and TGFβ expression. Cell Death Differ.
22:336–350. 2015. View Article : Google Scholar
|
|
36
|
Qiao Y, Shiue CN, Zhu J, Zhuang T, Jonsson
P, Wright AP, Zhao C and Dahlman-Wright K: AP-1–mediated chromatin
looping regulates ZEB2 transcription: New insights into TNFα-
induced epithelial-mesenchymal transition in triple-negative breast
cancer. Oncotarget. 6:7804–7814. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nam EH, Lee Y, Moon B, Lee JW and Kim S:
Twist1 and AP-1 cooperatively upregulate integrin α5 expression to
induce invasion and the epithelial-mesenchymal transition.
Carcinogenesis. 36:327–337. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gao J, Yan Q, Wang J, Liu S and Yang X:
Epithelial-to- mesenchymal transition induced by TGF-β1 is mediated
by AP1–dependent EpCAM expression in MCF-7 cells. J Cell Physiol.
230:775–782. 2015. View Article : Google Scholar
|
|
39
|
Gupta SC, Patchva S and Aggarwal BB:
Therapeutic roles of curcumin: Lessons learned from clinical
trials. AAPS J. 15:195–218. 2013. View Article : Google Scholar :
|
|
40
|
Shanmugam MK, Rane G, Kanchi MM, Arfuso F,
Chinnathambi A, Zayed ME, Alharbi SA, Tan BK, Kumar AP and Sethi G:
The multifaceted role of curcumin in cancer prevention and
treatment. Molecules. 20:2728–2769. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fetoni AR, Paciello F, Mezzogori D, Rolesi
R, Eramo SL, Paludetti G and Troiani D: Molecular targets for
anticancer redox chemotherapy and cisplatin-induced ototoxicity:
The role of curcumin on pSTAT3 and Nrf-2 signalling. Br J Cancer.
113:1434–1444. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Woo MS, Jung SH, Kim SY, Hyun JW, Ko KH,
Kim WK and Kim HS: Curcumin suppresses phorbol ester-induced matrix
metalloproteinase-9 expression by inhibiting the PKC to MAPK
signaling pathways in human astroglioma cells. Biochem Biophys Res
Commun. 335:1017–1025. 2005. View Article : Google Scholar : PubMed/NCBI
|