Introduction
Cancer is known to have multi-factorial etiology.
Malignant transformation of terminally differentiated cells is
characterized by several key hallmarks that directly or indirectly
lead to uncontrolled cell proliferation, reduced cell death,
enhanced angiogenesis and metastatic potential (1,2).
Along with these more recently chronic inflammation is a
well-recognized hallmark of cancer which is interestingly both a
cause and effect of malignant transformation (3). The genetic events causing malignant
transformation also initiate the expression of inflammation-related
machinery that leads to development of a tumor inflammatory mileu
(4). Oncogenes such as RAS,
MYC, and tyrosine kinases are known to induce proinflammatory
cytokines and angiogenic signals (5). Conversely, chronic inflammation
caused by carcinogens and infections is also known to induce the
malignant genetic changes.
Epidemiologic studies increasingly support the
notion that a strong positive correlation exits between chronic
inflammatory diseases and risk of cancer development. Several
critical molecular targets such as inducible nitric oxide synthase
(iNOS), nuclear factor κB (NF-κB) and hypoxia-inducible factor-1α
(HIF-1α), have been identified to mediate the secretion of
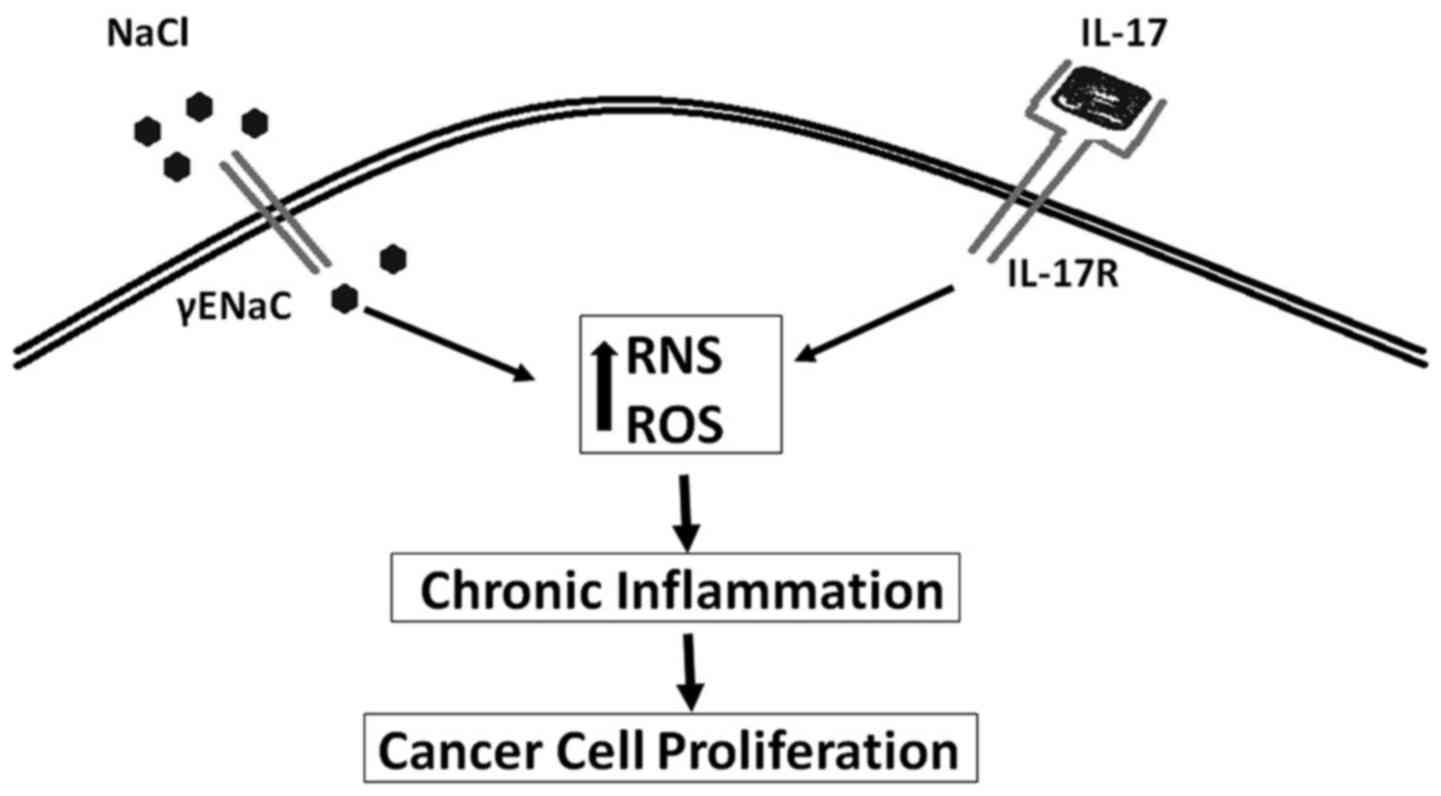
inflammatory cytokines (Fig. 1)
that either directly or indirectly induce carcinogenesis (6). Although several inflammatory causes
for cancers have been suggested environmental and dietary link is
well established (7). High salt
diet is well known to induce a pro-inflammatory cascade leading to
chronic inflammatory diseases such as hypertension, myocardial
infarction, neurological transient ischemic attack and cancer
(8,9). While the exact magnitude of
salt-intake needed to induce inflammatory diseases is yet to be
defined, however a cellular and immune dysregulation mediated by
salt and potentially leading to cancer is well established and will
be discussed in this review.
2. Impact of salt on cancer metabolism
Role of inflammatory sodium ion
channels
An intratumor high salt concentration was
demonstrated in early 1980s, when researchers demonstrated that the
sodium content of breast tumors is significantly higher compared to
a normal lactating breast epithelium (10). In these original studies it is
unclear if the tumor activity is directly correlated with high
intratumor salt concentration or if the high sodium concentration
is extracellular or intracellular. Regardless, salt is known to
induce inflammation. One of the well realized inflammatory markers,
inducible nitric oxide synthases (iNOS) is shown to be upregulated
in several cancers including breast cancers (11). Our cellular studies on breast
cancer cell lines MDA-MB-231 and MCF7 have shown that high sodium
chloride concentration (0.05 M above basal level) was able to
induce the production of reactive nitrogen and oxygen species
(RNS/ROS) (12). Importantly an
equimolar addition of mannitol to the media did not induce similar
effect, thus suggesting sodium play a direct pro-inflammatory
effect on breast cancer cell lines. Furthermore, earlier studies in
1980s in brain cancers have shown that an intracellular influx of
sodium ions induced cancerous cell proliferation (13,14).
An increasing number of studies have shown that epithelial sodium
channel (ENaC) which controls the intracellular entry of sodium
ions is correlated with cancerous cell changes including,
uncontrolled cell proliferation, anti-apoptosis and cell migration.
Studies by Bondarava et al have demonstrated that ENaC
played a critical role in the sodium induced cell proliferation of
HepG2 cells (15). In line with
this evidence, recent studies from our laboratory (Fig. 1) have demonstrated increased sodium
concentration specifically induced upregulation of γENaC in the
breast cancer cell lines and enhanced cellular proliferation
(12). It is of interest to note
that, the ENaC expression levels in breast cancer cell lines were
upregulated by the mineralo-corticoid hormone aldosterone, known to
control ionic equilibrium and not glucocorticoid steroid hormones
(16). All these data suggest a
direct functional role of ENaC in high-sodium induced cancerous
development.
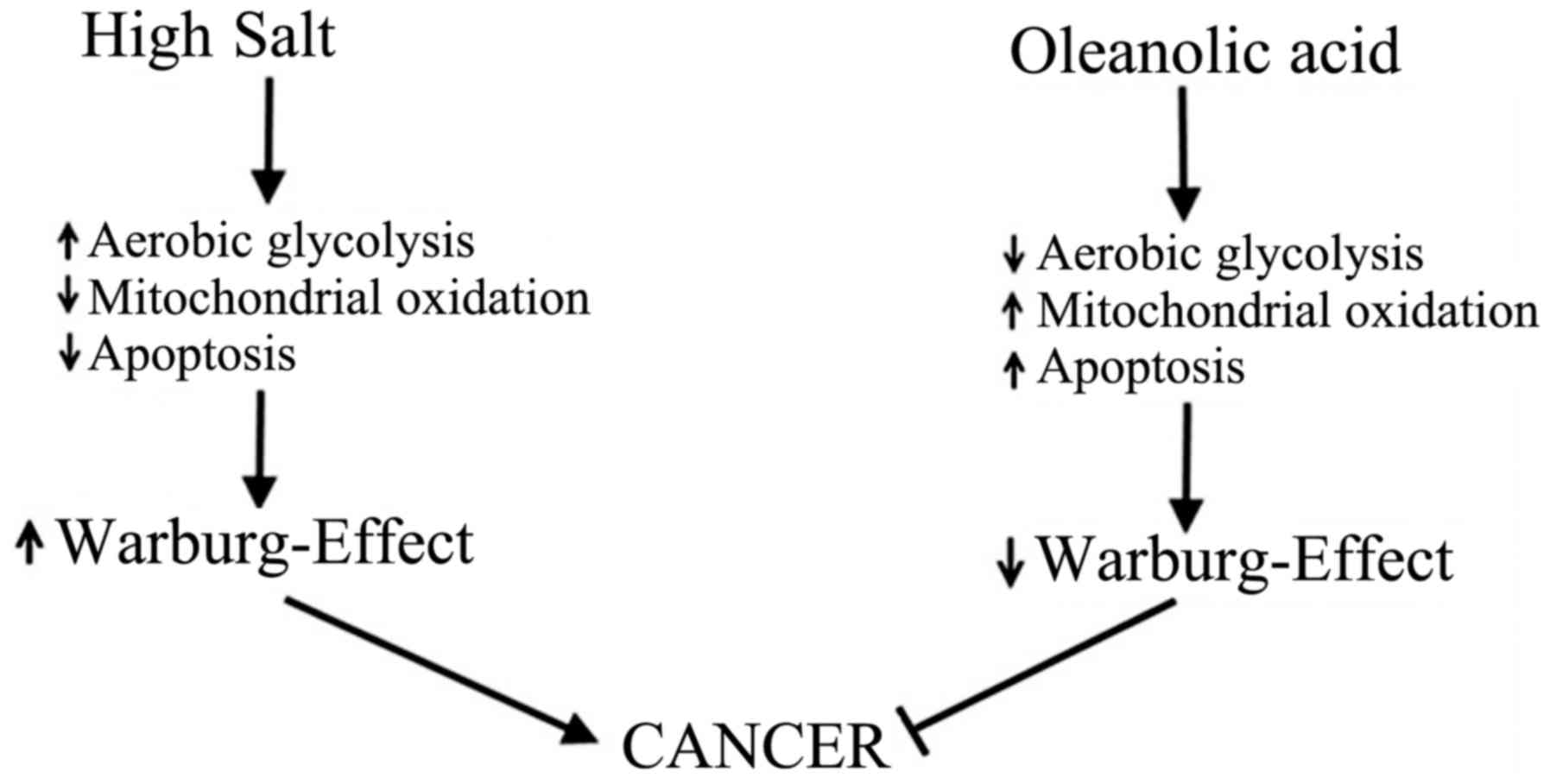
Role of salt in Warburg-metabolism
In a terminally differentiated cell, under normoxic
conditions, glucose metabolism by mitochondrial oxidation yields
36–38 ATP. However, a cancerous cell under similar normoxic
conditions, yields 2 ATP with subsequent metabolism of glucose to
lactate. This aberrant behavior of cancerous cells is called
aerobic glycolysis or Warburg effect named after the scientist Otto
Warburg who originally discovered this phenomena in 1923 (17). This glycolytic switch in cancer
cells has been used to explain their extraordinary ability to
tolerate hypoxia and inflammatory-stress. Furthermore, this
glycolytic switch converts a catabolic glucose metabolism producing
carbon dioxide to anabolic glucose metabolism resulting in higher
lipid biosynthesis needed for cancerous cell proliferation
(18). Osmotic stress induced by
hyertonic saline has been suggested to induce Warburg-like effect
by enhancing glucose transport and lactic acidosis in cancer cells
(19). In line with these
findings, our recent studies have demonstrated that high salt in
the external environment induced Warburg-like effect of increased
glucose consumption and lactic acid production in breast cancer
cells (Fig. 2), which was
abrogated by anti-inflammatory compound oleanolic acid (20). Taken together, these data suggest
that high sodium plays a direct role in inducing cancerous
metabolic phenotype.
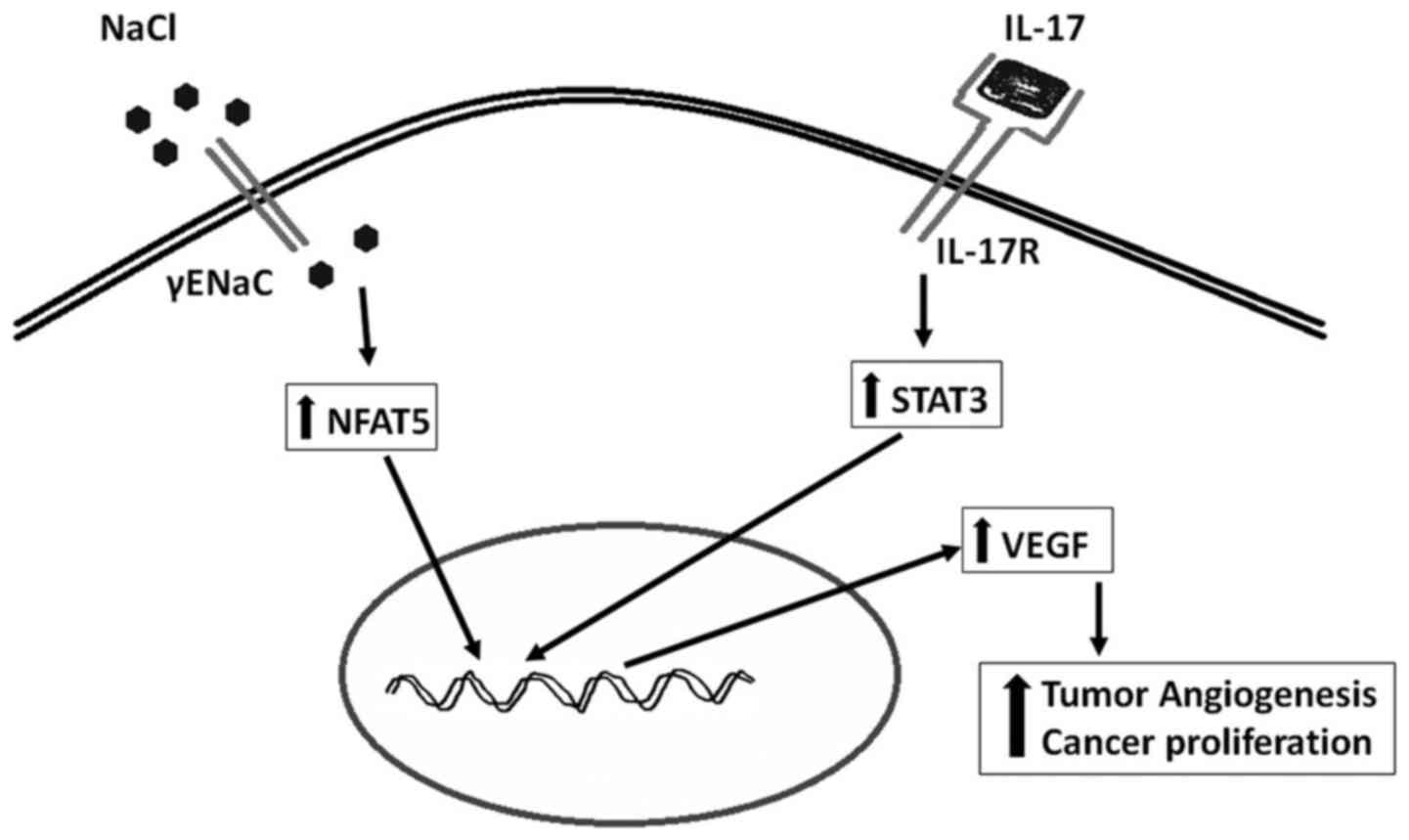
Role of high salt in angiogenesis
Increased formation of new blood vessels to the
newly formed tumor to support its growth and metabolism is
considered as one of the important hallmarks of cancer development.
Vascular endothelial growth factor (VEGF) is known to induce
angiogenesis to tumors through activation of cancer-specific
AkT/PI3k signaling mechanism (21). Recently strategies targeted against
VEGF have been utilized in anticancer therapies (22). Our recent studies have demonstrated
that high salt directly induces the expression of VEGF in breast
cancer cells through the transcription factor (Fig. 3), nuclear factors of activated T
cells or NFAT5 signaling (23).
The NFAT5 also known as tonicity-responsive enhancer binding
protein (TonEBP) is a well-established osmotic response element
binding protein known to regulate intracellular osmotic tonicity
(24). The NFAT5 is shown to exert
a critical role in immune functioning and lymphocyte
differentiation (25). Gene
expression studies by Remo et al (26) on breast cancer patients (n=197)
have suggested a strong positive correlation between the expression
of NFAT5 and inflammatory breast cancer. These accumulating data
clearly suggest that high salt mediates a direct pro-angiogenic
response in the tumor mileu.
3. Impact of salt on macrophages
The innate myeloid lineage macrophages play a
critical role of antigen presentation and immune homeostasis. These
terminally differentiated cells uniquely demonstrate a high degree
of plasticity in response to changes in microenvironment. Two well
recognized phenotypes of tissue-resident macrophages are
classically activated pro-inflammatory type 1 macrophages (M1) and
the alternatively-activated immunosuppressive type 2 macrophages
(M2) (27). Various factors have
been implicated in the phenotype switch of macrophages. The
anti-inflammatory M2 phenotype is well established to be a resident
phenotype in tumor TAMs (tumor associated macrophages) are shown to
be negatively correlated with cancer prognosis. One of the
signature cytokines released from TAMs is interleukin-10.
Studies in our laboratory have demonstrated that
treatment with high salt in the culture media induced specific
differentiation of peripheral blood mononuclear cells to M2
phenotype (CD11b+CD14lowCD16+). As
tumors are known to have high salt microenvironment (28), our data strongly suggest that high
salt induces anti-inflammatory M2 phenotype which are predominantly
accumulated in tumor microenvironment (29). The cytokine array expressed in the
tumor microenvironment is known to plays a critical role in the
activation and differentiation of newly migrated mononuclear
phagocytes (30). While, the M1
phenotype is induced by the cytokines interferon γ (IFNγ) and tumor
necrosis factor α (TNFα) (31),
the alternatively activated immune-suppressive TAM phenotype is
induced by transforming growth factor β (TGFβ) in the cancer
microenvironment (32). An
important dissimilarity between MΦ1 and M2 phenotypes is their
cytokine pattern. The M1 phenotype secrets inflammatory cytokines
(IL6, IL-1β and TNFα) that are antitumor, while, M2 phenotype
secretes pro-tumor IL-10, and VEGF (9). However, it seems the differentiation
of tissue resident macrophages is also site-specific. Studies by
Jantsch et al have demonstrated that, high salt activation
macrophage induced an M1 phenotype switch offering protection
against protozoan Leishmania skin infection (33). These data suggest that the high
salt mediated activation of macrophage is tissue-specific and
dependent on resident microenvironment.
4. Impact of salt on CD4+ T
cells
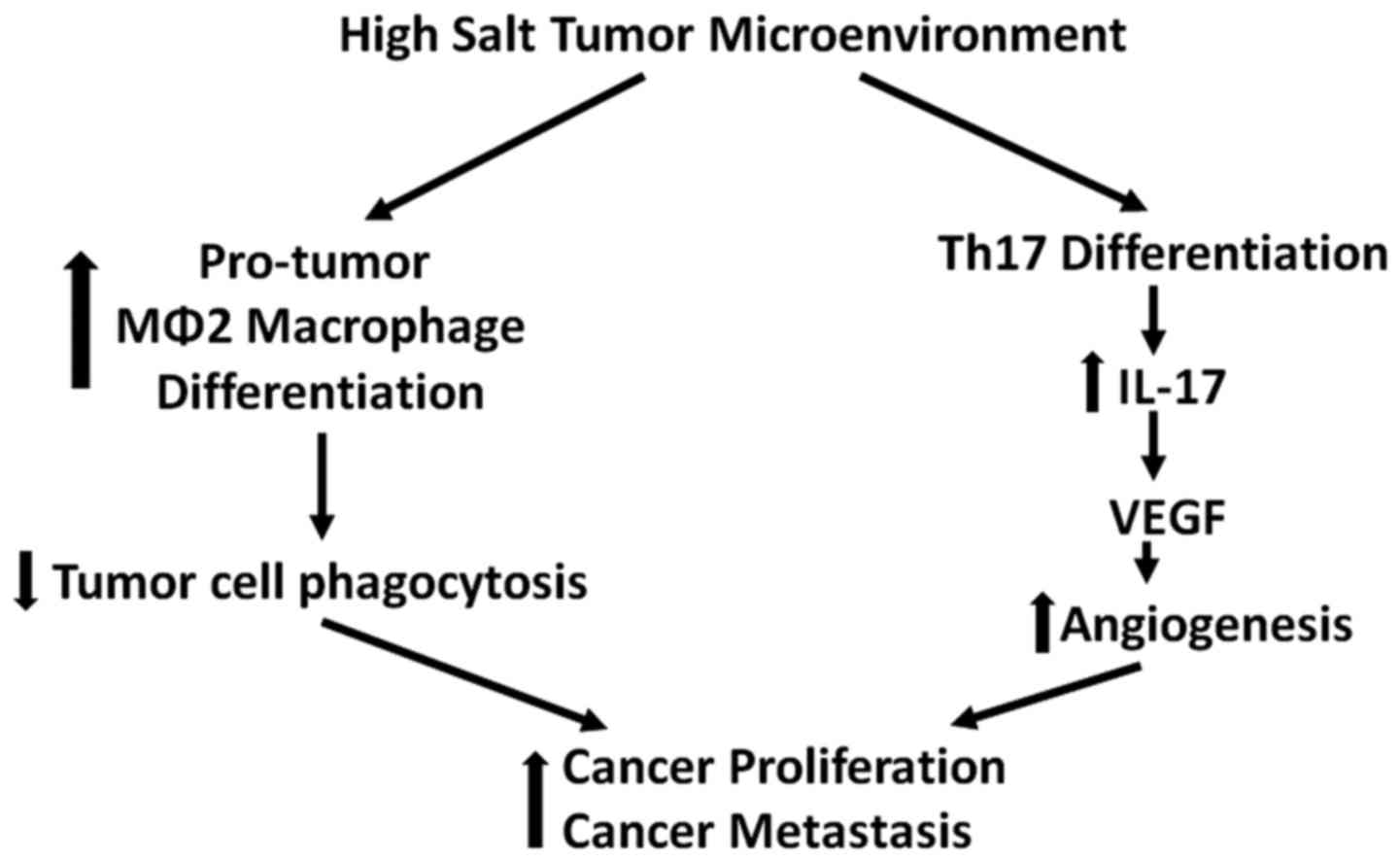
Local tumor inflammatory mileu is known to recruit
several immune cells including CD4+ T helper (Th)
lymphocytes. A subset of Th-cells which secrets a pro-inflammatory
cytokine IL-17 is well established and is called Th17 phenotype
(34). Several pre-clinical human
cancer studies have demonstrated increased frequency of Th17 cells
in the tumor infiltrating lymphocytes in several solid organ tumors
(35,36). The secretion of chemokines, MCP-1
and RANTES, by tumor resident fibroblasts have been suggested to
play a key role in the recruitment of Th17 cells into the tumor
(37). Along with these chemokines
several other factors released by tumor cells such as Aryl
hydrocarbon receptor (AhR) ligands, adenosine, and metabolites from
hypoxia have been suggested to induce Th17 differentiation of naïve
CD4+ Th cells in the tumor microenvironment (38,39).
Furthermore, the newly recruited Th17 cells have been suggested to
further induce tumor growth and metastasis (Fig. 4). Specifically, IL-17 has been
demonstrated to induce pro-angiogenic VEGF in the tumor mileu
leading to enhanced tumorigenicity and metastasis (40).
While, the precise mechanisms causing the Th17
differentiation in the tumor microenvironment are unclear, one
possible hypothesis would be the possibility of high salt in the
tumor microenvironment might influence differentiation of naïve
CD4+ T cells to Th17 phenotype. A very important
indirect evidence to this effect, has been provided by Wu et
al, where under in vitro conditions the researchers
demonstrated that high salt in the culture media induced Th17
differentiation of naïve CD4+ T cells (41). In this study the authors have
identified a salt-sensing transcription factor,
serum-glucocorticoid kinase (SGk1), as the principal molecule it
induces salt-specific differentiation to Th17 phenotype. The SGk1
knockdown in these experiments failed to induce salt-mediated Th17
differentiation.
Immunologically regulatory T cells (Tregs) have been
shown to exert an antagonistic effect on Th17 differentiation and
its molecular downstream effector function. The Treg cells are
known to exert anti-inflammatory effects through secretion of
immune-suppressive IL-10 cytokine. To further consolidate the
notion that salt induces Th17 differentiation, studies by Hernandez
et al, demonstrated that high salt exerts inhibitory effect
on the suppressive activity of Treg (42). In these studies, researchers
demonstrated that the exposure of Treg to high salt inhibited the
suppressive function along with increase in the pro-inflammatory
factors such as IFNG, TBX21 and CXCR3. Furthermore,
under these conditions there was an increase in the inflammatory
IL-17 transcript confirming the studies by Wu et al
(41) mentioned above.
Furthermore, utilizing a humanized mouse model of graft versus host
disease (GvHD) and a murine model of experimental colitis,
Hernandez et al have shown that high salt induced an
inflammatory CD4+ T cell immunophenotype and worsening
of the disease (42). However, a
direct study on the role of salt on the tumor will be needed to
confirm the effect of salt on tumors.
5. Future perspectives
High salt diet is known to be the key culprit in
several chronic inflammatory diseases including cardiovascular
disease, cancer and autoimmune diseases. A typical adult Western
diet contains 10–12 g sodium per day which is 2–3 times above the
WHO recommended 4.8 g of sodium per day (43). Several epidemiological studies have
conclusively supported a positive correlation between salt and
cancer. A wide-spread awareness on salt-restricted diet might be
able to reduce the community disease load. Recent pre-clinical and
clinical studies have identified several molecular targets which
mediate salt-induced damage. These molecular targets could offer
novel futuristic intervention treatment strategies for personalized
precision medicine.
References
|
1
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Balkwill F and Mantovani A: Inflammation
and cancer: Back to Virchow? Lancet. 357:539–545. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mantovani A, Allavena P, Sica A and
Balkwill F: Cancer-related inflammation. Nature. 454:436–444. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Borrello MG, Alberti L, Fischer A,
Degl'innocenti D, Ferrario C, Gariboldi M, Marchesi F, Allavena P,
Greco A, Collini P, et al: Induction of a proinflammatory program
in normal human thyrocytes by the RET/PTC1 oncogene. Proc Natl Acad
Sci USA. 102:14825–14830. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ischenko I, Zhi J, Moll UM, Nemajerova A
and Petrenko O: Direct reprogramming by oncogenic Ras and Myc. Proc
Natl Acad Sci USA. 110:3937–3942. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lu H, Ouyang W and Huang C: Inflammation,
a key event in cancer development. Mol Cancer Res. 4:221–233. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang XQ, Terry PD and Yan H: Review of
salt consumption and stomach cancer risk: Epidemiological and
biological evidence. World J Gastroenterol. 15:2204–2213. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aburto NJ, Ziolkovska A, Hooper L, Elliott
P, Cappuccio FP and Meerpohl JJ: Effect of lower sodium intake on
health: Systematic review and meta-analyses. BMJ. 346(apr03 3):
f13262013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ge S, Feng X, Shen L, Wei Z, Zhu Q and Sun
J: Association between habitual dietary salt intake and risk of
gastric cancer: A systematic review of observational studies.
Gastroenterol Res Pract. 2012:8081202012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sparks RL, Pool TB, Smith NK and Cameron
IL: Effects of amiloride on tumor growth and intracellular element
content of tumor cells in vivo. Cancer Res. 43:73–77.
1983.PubMed/NCBI
|
|
11
|
Lechner M, Lirk P and Rieder J: Inducible
nitric oxide synthase (iNOS) in tumor biology: The two sides of the
same coin. Semin Cancer Biol. 15:277–289. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Amara S, Ivy MT, Myles EL and Tiriveedhi
V: Sodium channel γENaC mediates IL-17 synergized high salt induced
inflammatory stress in breast cancer cells. Cell Immunol. 302:1–10.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
O'Donnell ME, Cragoe E Jr and Villereal
ML: Inhibition of Na+ influx and DNA synthesis in human
fibroblasts and neuroblastoma-glioma hybrid cells by amiloride
analogs. J Pharmacol Exp Ther. 226:368–372. 1983.PubMed/NCBI
|
|
14
|
O'Donnell ME and Villereal ML: Membrane
potential and sodium flux in neuroblastoma X glioma hybrid cells:
Effects of amiloride and serum. J Cell Physiol. 113:405–412. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bondarava M, Li T, Endl E and Wehner F:
alpha-ENaC is a functional element of the hypertonicity-induced
cation channel in HepG2 cells and it mediates proliferation.
Pflugers Arch. 458:675–687. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Masilamani S, Kim GH, Mitchell C, Wade JB
and Knepper MA: Aldosterone-mediated regulation of ENaC alpha,
beta, and gamma subunit proteins in rat kidney. J Clin Invest.
104:R19–R23. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Maldonado EN and Lemasters JJ: ATP/ADP
ratio, the missed connection between mitochondria and the Warburg
effect. Mitochondrion. 19(Pt A): 78–84. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Palsson-McDermott EM and O'Neill LA: The
Warburg effect then and now: From cancer to inflammatory diseases.
BioEssays. 35:965–973. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Epstein T, Xu L, Gillies RJ and Gatenby
RA: Separation of metabolic supply and demand: Aerobic glycolysis
as a normal physiological response to fluctuating energetic demands
in the membrane. Cancer Metab. 2:72014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Amara S, Zheng M and Tiriveedhi V:
Oleanolic acid inhibits high salt-induced exaggeration of
Warburg-like metabolism in breast cancer cells. Cell Biochem
Biophys. 74:427–434. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pidgeon GP, Barr MP, Harmey JH, Foley DA
and Bouchier-Hayes DJ: Vascular endothelial growth factor (VEGF)
upregulates BCL-2 and inhibits apoptosis in human and murine
mammary adenocarcinoma cells. Br J Cancer. 85:273–278. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Harmey JH and Bouchier-Hayes D: Vascular
endothelial growth factor (VEGF), a survival factor for tumour
cells: Implications for anti-angiogenic therapy. BioEssays.
24:280–283. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Amara S, Alotaibi D and Tiriveedhi V:
NFAT5/STAT3 interaction mediates synergism of high salt with IL-17
towards induction of VEGF-A expression in breast cancer cells.
Oncol Lett. 12:933–943. 2016.PubMed/NCBI
|
|
24
|
Neuhofer W: Role of NFAT5 in inflammatory
disorders associated with osmotic stress. Curr Genomics.
11:584–590. 2010. View Article : Google Scholar
|
|
25
|
Berga-Bolaños R, Drews-Elger K, Aramburu J
and López-Rodríguez C: NFAT5 regulates T lymphocyte homeostasis and
CD24-dependent T cell expansion under pathologic hypernatremia. J
Immunol. 185:6624–6635. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Remo A, Simeone I, Pancione M, Parcesepe
P, Finetti P, Cerulo L, Bensmail H, Birnbaum D, Van Laere SJ,
Colantuoni V, et al: Systems biology analysis reveals NFAT5 as a
novel biomarker and master regulator of inflammatory breast cancer.
J Transl Med. 13:1382015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Quatromoni JG and Eruslanov E:
Tumor-associated macrophages: Function, phenotype, and link to
prognosis in human lung cancer. Am J Transl Res. 4:376–389.
2012.PubMed/NCBI
|
|
28
|
Roger S, Besson P and Le Guennec JY:
Involvement of a novel fast inward sodium current in the invasion
capacity of a breast cancer cell line. Biochim Biophys Acta.
1616:107–111. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Amara S, Whalen M and Tiriveedhi V: High
salt induces anti-inflammatory MΦ2-like phenotype in peripheral
macrophages. Biochem Biophys Rep. 7:1–9. 2016.PubMed/NCBI
|
|
30
|
Solinas G, Germano G, Mantovani A and
Allavena P: Tumor-associated macrophages (TAM) as major players of
the cancer-related inflammation. J Leukoc Biol. 86:1065–1073. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lewis CE and Pollard JW: Distinct role of
macrophages in different tumor microenvironments. Cancer Res.
66:605–612. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Colotta F, Allavena P, Sica A, Garlanda C
and Mantovani A: Cancer-related inflammation, the seventh hallmark
of cancer: Links to genetic instability. Carcinogenesis.
30:1073–1081. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jantsch J, Schatz V, Friedrich D, Schröder
A, Kopp C, Siegert I, Maronna A, Wendelborn D, Linz P, Binger KJ,
et al: Cutaneous Na+ storage strengthens the
antimicrobial barrier function of the skin and boosts
macrophage-driven host defense. Cell Metab. 21:493–501. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Guéry L and Hugues S: Th17 cell plasticity
and functions in cancer immunity. BioMed Res Int. 2015:3146202015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang L, Yi T, Kortylewski M, Pardoll DM,
Zeng D and Yu H: IL-17 can promote tumor growth through an
IL-6-Stat3 signaling pathway. J Exp Med. 206:1457–1464. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bailey SR, Nelson MH, Himes RA, Li Z,
Mehrotra S and Paulos CM: Th17 cells in cancer: The ultimate
identity crisis. Front Immunol. 5:2762014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Su X, Ye J, Hsueh EC, Zhang Y, Hoft DF and
Peng G: Tumor microenvironments direct the recruitment and
expansion of human Th17 cells. J Immunol. 184:1630–1641. 2010.
View Article : Google Scholar
|
|
38
|
Kimura A, Naka T, Nohara K, Fujii-Kuriyama
Y and Kishimoto T: Aryl hydrocarbon receptor regulates Stat1
activation and participates in the development of Th17 cells. Proc
Natl Acad Sci USA. 105:9721–9726. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wilson JM, Kurtz CC, Black SG, Ross WG,
Alam MS, Linden J and Ernst PB: The A2B adenosine receptor promotes
Th17 differentiation via stimulation of dendritic cell IL-6. J
Immunol. 186:6746–6752. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu X, Yang T, Liu X, Guo JN, Xie T, Ding
Y, Lin M and Yang H: IL-17 promotes tumor angiogenesis through
Stat3 pathway mediated upregulation of VEGF in gastric cancer.
Tumour Biol. 37:5493–5501. 2016. View Article : Google Scholar
|
|
41
|
Wu C, Yosef N, Thalhamer T, Zhu C, Xiao S,
Kishi Y, Regev A and Kuchroo VK: Induction of pathogenic TH17 cells
by inducible salt-sensing kinase SGK1. Nature. 496:513–517. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hernandez AL, Kitz A, Wu C, Lowther DE,
Rodriguez DM, Vudattu N, Deng S, Herold KC, Kuchroo VK,
Kleinewietfeld M, et al: Sodium chloride inhibits the suppressive
function of FOXP3+ regulatory T cells. J Clin Invest.
125:4212–4222. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cordain L, Eaton SB, Sebastian A, Mann N,
Lindeberg S, Watkins BA, O'keefe JH and Brand-Miller J: Origins and
evolution of the Western diet: Health implications for the 21st
century. Am J Clin Nutr. 81:341–354. 2005.PubMed/NCBI
|