Introduction
Ovarian cancer is one of the most common types of
gynecological cancers and the 5th leading cause of cancer-related
mortality in females (1,2). Despite enormous improvements over the
past years, the prognosis of patients with ovarian cancer remains
dismal, and this is largely attributed to chemotherapeutic
resistance following several cycles of treatment. Hence, a better
understanding of the underlying mechanisms responsible for
chemotherapeutic resistance in ovarian cancer may aid in the
development of novel targets for therapeutic intervention in
ovarian cancer.
The Hippo pathway represents a novel tumor
suppressor pathway and the dysregulation of Hippo signaling has
been demonstrated to be involved in several biological process and
tumor development (3–5). From a molecular perspective, 4 core
kinase components have been reported to be essential for the tumor
suppressive function of Hippo signaling, including mammalian
STE20-like kinase (MST)1/2, protein salvador homolog 1 (SAV1),
large tumor-suppressor kinase (LATS)1/2 and Mps one binder kinase
activator-like 1 (MOB1) (5).
Physiologically, the Hippo signaling pathway is active, which
tightly controls the activity of 2 critical downstream effectors of
the Hippo pathway, YAP and tafazzin (TAZ), both spatially and
temporally via the phosphorylation of YAP and TAZ by these 4 core
kinase cassettes, leading to the degradation of YAP and TAZ
(6,7). Conversely, when the activity of Hippo
signaling is inhibited by unknown mechanisms, phosphorylated YAP
and TAZ translocate to the nucleus and elicit the transcriptional
activity of TEA domain (TEAD) family members, including TEAD1-4
(8–10), which further transcriptionally
activates downstream effectors to exert its versatile functions
(11–13). Recently, accumulating evidence has
indicated that the inactivation of Hippo promotes the development
of chemotherapeutic resistance in numerous human cancers. For
example, the study by Lai et al reported that upregulation
of TAZ induced Taxol resistance in breast cancer cells via
transcriptionally activating Cyr61 and CTGF (14); moreover, the hyperactivation of YAP
has been reported to be positively associated with a poor
chemotherapeutic response to taxanes and a poor prognosis in
patients with ovarian cancer (15), indicating that the inactivation of
the Hippo pathway is crucial for the development of cancer
chemoresistance. Therefore, a better understanding of the
mechanisms responsible for the inactivation of the Hippo pathway
may provide new insight and may aid in the development of more
effective treatment strategies for ovarian cancer.
MicroRNAs (miRNAs or miRs) are small endogenous
non-coding RNAs that are able to repress a variety of target genes
at the post-transcriptional level by binding with the 3′
untranslated region (3′UTR) of the target gene mRNAs (16). Numerous studies have reported that
miRNAs play crucial roles in tumorigenesis, malignant progression
and the metastasis of cancers through various mechanisms (17–22).
Furthermore, studies have also supported that the up- or
downregulation of a certain miRNA can be directly tied to the
response to chemotherapeutic agents (23–26).
In this study, from a publically available miRNA dataset from The
Cancer Genome Atlas (TCGA) and our experiments, we found that
miR-149-5p expression was significantly elevated in chemo-resistant
ovarian cancer tissues and cell lines compared with chemosensitive
ovarian cancer ones. Furthermore, the silencing of miR-149-5p
increased the apoptotic ratio and decreased the mitochondrial
potential of ovarian cancer cells in response to cisplatin (CDDP)
in vitro, and also enhanced the chemotherapeutic sensitivity
of ovarian cancer cells to CDDP in vivo. Conversely, the
upregulation of miR-149-5p decreased the apoptotic ratio and
increased the mitochondrial potential of ovarian cancer cells in
response to to CDDP in vivo. Our results further
demonstrated that miR-149-5p directly targeted MST1 and SAV1,
leading to the inactivation of TEAD transcription. Importantly, the
effects of miR-149-5p silencing on the apoptotic ratio,
mitochondrial potential and caspase-3 activity in ovarian cancer
cells were attenuated by the individual knockdown of MST1 and SAV1.
Therefore, our findings reveal a novel mechanism through which
miR-149-5p induces chemotherapeutic resistance in ovarian cancer,
suggesting that miR-149-5p may be used as a chemotherapeutic
response indicator in ovarian cancer.
Materials and methods
Cell culture and agents
The ovarian cancer cell lines, TOV-21G, A2780,
OVCAR-3, Caov-3, ES-2, HO-8910 and SK-OV-3, were obtained from the
Shanghai Chinese Academy of Sciences cell bank (Shanghai, China),
and all human ovarian cancer cell lines were maintained in
RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with
10% FBS (HyClone, Logan, UT, USA). CDDP (Cat. no. P4394;
Sigma-Aldrich, St. Louis, MO, USA), which undergoes relatively slow
solvolysis in DMSO, was dissolved in 0.9% NaCl at 4 mM, immediately
aliquoted and stored at −70°C. Thawed aliquots were used only once.
The stock solutions were further diluted with DMSO and added to the
cell cultures with the final DMSO concentrations being <0.05%.
The ovarian cancer cells were treated with cisplatin (5 µM,
48 h) before further functional assays.
Patients and tumor tissues
A total of 20 ovarian cancer tissues and 10 benign
ovarian lesion tissues, including 4 simple ovarian cyst and 6
simple ovarian serous cystadenoma were obtained during surgery at
the Sun Yat-sen University Cancer Center (Guangzhou, China)
(Tables I and II). Patients were diagnosed based on
clinical and pathological evidence, and the specimens were
immediately snap-frozen and stored in liquid nitrogen tanks. For
the use of these clinical samples for research purposes, prior
patient consents and approval from the Institutional Research
Ethics Committee of Sun Yat-sen University Cancer Center were
obtained (ethics approval no. 308-2016-03-01).
 | Table IThe basic information of 20 patients
with ovarian serous cystadenocarcinoma for miR-149-5p expression
analysis. |
Table I
The basic information of 20 patients
with ovarian serous cystadenocarcinoma for miR-149-5p expression
analysis.
| Characteristic | Cases (n) | Percentage |
|---|
| Age (years) | | |
| <55 | 6 | 30 |
| ≥55 | 14 | 70 |
| Stage | | |
| Stage I | 1 | 5 |
| Stage II | 1 | 5 |
| Stage III | 14 | 70 |
| Stage IV | 4 | 20 |
| Grade | | |
| G1 | 0 | 0 |
| G2 | 3 | 15 |
| G3 | 17 | 85 |
| Treatment | | |
| Complete
response | 10 | 50 |
| Partial
response | 1 | 5 |
| Stable
disease | 3 | 15 |
| Progressive
disease | 6 | 30 |
| Table IIThe basic information of 10 patients
with benign ovarian lesions for miR-149-5p expression analysis. |
Table II
The basic information of 10 patients
with benign ovarian lesions for miR-149-5p expression analysis.
| Characteristic | Cases (n) | Percentage |
|---|
| Age (years) | | |
| <55 | 4 | 40.0 |
| ≥55 | 6 | 60.0 |
| Type of
disease | | |
| Simple cyst | 4 | 40.0 |
| Simple serous
cystadenoma | 6 | 60.0 |
RNA extraction, reverse transcription and
real-time PCR
Total RNA from the tissues or cells was extracted
using TRIzol reagent (Life Technologies/Thermo Fisher Scientific,
Waltham, MA, USA) according to the manufacturer's instructions.
Messenger RNAs (mRNAs) were polyadenylated using a poly-A
polymerase-based First-Strand Synthesis kit (Takara, Dalian, China)
and the reverse transcription (RT) from total mRNA was performed
using a PrimeScript RT Reagent kit (Takara) according to the
manufacturer's instructions. miRNAs were reverse transcribed from
total mRNA using the RevertAid First Strand cDNA Synthesis kit
(Thermo Fisher Scientific) according to the manufacturer's
instructions. Complementary DNA (cDNA) was amplified and quantified
on an ABI 7500HT system (Applied Biosystems, Foster City, CA, USA)
using SYBR-Green I (Applied Biosystems). The RT-PCR reaction was
performed with the following parameter values: 15 min at 37°C, 10
min at 65°C, 5 min at 85°C and −20°C until use. RT-PCR analysis was
performed on an iQ5 Real Time PCR Detection System (Bio-Rad,
Hercules, CA, USA) with a 20 µl volume reaction containing 2
µl reverse transcription product, 10 µl 2All-in-One™
Q-PCR Mix, 2 µl PCR Forward Primer (2 µM), 2
µl Universal Adaptor PCR Primer (2 µM) and 4
µl ddH2O. The reactions were incubated in 96-well
plates at 95°C for 10 min, following by 40 cycles, and then ramped
from 66°C to 95°C to obtain the melting curve. Each sample was
analyzed in triplicate. The primers sequence and the product size
were listed in Table III.
Real-time PCR was performed according to a standard method, as
previously described (27).
Primers for U6 and miR-149-5p (miRQ0000450-1-1) were synthesized
and purified by RiboBio (Guangzhou, China). U6 or
glycer-aldehyde-3-phosphate dehydrogenase (GAPDH) was used as
endogenous controls for miRNA or mRNA respectively. Relative fold
expression was calculated with the comparative threshold cycle
(2−ΔΔCq) method as previously described (28).
| Table IIIList of primers used in the reactions
for RT-qPCR. |
Table III
List of primers used in the reactions
for RT-qPCR.
| Gene | Sequence
(5′-3′) | Product size
(bp) |
|---|
| BCL2-up |
GGTGGGGTCATGTGTGTGG | 89 |
| BCL2-dn |
CGGTTCAGGTACTCAGTCATCC | |
| BCL2L1-up |
CTGCTGCATTGTTCCCATAG | 289 |
| BCL2L1-dn |
TTCAGTGACCTGACATCCCA | |
| CTGF-up |
TGGAGATTTTGGGAGTACGG | 107 |
| CTGF-dn |
CAGGCTAGAGAAGCAGAGCC | |
| CYR61-up |
GGTCAAAGTTACCGGGCAGT | 115 |
| CYR61-dn |
GGAGGCATCGAATCCCAGC | |
| HOXA1-up |
TCCTGGAATACCCCATACTTAGC | 95 |
| HOXA1-dn |
GCACGACTGGAAAGTTGTAATCC | |
| SOX9-up |
AGCGAACGCACATCAAGAC | 85 |
| SOX9-dn |
CTGTAGGCGATCTGTTGGGG | |
| GAPDH-up |
TCCTCTGACTTCAACAGCGACAC | 126 |
| GAPDH-dn |
CACCCTGTTGCTGTAGCCAAATTC | |
Plasmids, small interfering RNA and
transfection
The miR-149-5p expression plasmid was generated by
cloning the genomic pre-miR-149-5p gene into the retroviral
transfer plasmid pMSCV-puro (Clontech Laboratories Inc., Tokyo,
Japan) to generate plasmid pMSCV-miR-149-5p. Subsequently,
pMSCV-miR-149-5p was co-transfected with the pIK packaging plasmid
(Cat. no. 75483; ATCC, VA, USA) into 293FT (CRL-1573) cells using
the standard calcium phosphate transfection method, as previously
described (29). At 36 h after the
co-transfection, the supernatants were collected and incubated with
the cells to be infected for 24 h in the presence of polybrene (2.5
µg/ml). Following infection, puromycin (1.5 µg/ml)
was used to select stably transduced cells over a 10-day period.
The luciferase reporter system of pTEAD-luc (Cat. no. 83467;
Addgene, Cambridge, MA, USA) was used to examine the
transcriptional activity of TEAD. The 3′-untranslated region
(3′UTR) regions of human MST1 and SAV1 were PCR-amplified from
genomic DNA and cloned into the pmirGLO luciferase reporter vector
(Promega, Madison, WI, USA) and the list of primers used in clone
reactions are presented in Table
IV. The miArrest plasmids for anti-miR-149-5p and negative
control plasmids were constructed and cloned into phU6 plasmids by
GeneChem (Shanghai, China). The sequence of anti-miR-149-5p was
GGGAGTGAAGACACGGAGCCAGA. Small interfering RNA (siRNA) for YAP,
TAZ, MST1 and SAV1 knockdown were obtained from Ribobio. The
transfection of siRNAs and the plasmids was performed using
Lipofectamine 3000 (Life Technologies/Thermo Fisher Scientific)
according to the manufacturer's instructions.
| Table IVList of primers used in the reactions
for clone PCR. |
Table IV
List of primers used in the reactions
for clone PCR.
| Gene | Sequence
(5′-3′) |
|---|
| miR-149-up |
GGGCACATTGCTCTCCATTCAG |
| miR-149-dn |
AGGCCCGAAACACCCGTAAG |
|
MST1-3′UTR-34nt-up |
CAGGCTTTGGGTGAATTCTGGATG |
|
MST1-3′UTR-1577nt-dn |
GTATCCTGAGGTGCAACCCAGTC |
|
SAV1-3′UTR-74nt-up |
CTCTGGTTAATAAATGCTGCTTCCTTTGTG |
|
SAV1-3′UTR-1449nt-dn |
CTGGGAGTAGTTTGTATTCTGGAAGAC |
Western blot analysis
Nuclear/cytoplasmic fractionation was carried out
using the Cell Fractionation kit (Cell Signaling Technology,
Danvers, MA, USA) according to the manufacturer's instructions, and
the whole cell lysates were extracted using RIPA Buffer (Cell
Signaling Technology). Western blot analysis was performed
according to a standard method, as previously described (30). Antibodies against YAP (Cat. no.
ab81183), TAZ (Cat. no. ab93362), MST1 (Cat. no ab76822), SAV1
(Cat. no. ab105105), Bcl-2 (Cat. no. ab194583) and Bcl-xL (Cat. no.
ab32370) were purchased from Abcam (Cambridge, MA, USA), and
antibodies against caspase-3 (Cat. no. 9662) and caspase-9 (Cat.
no. 9501) were from Cell Signaling Technology. The membranes were
stripped and reprobed with an anti-α-tubulin antibody
(Sigma-Aldrich) as the loading control.
Flow cytometric analysis
Flow cytometric analysis of apoptosis was carried
out using the FITC Annexin V Apoptosis Detection kit I (BD
Biosciences, San Jose, CA, USA), and was presented as the protocol
described below. Briefly, the cells were dissociated with trypsin
and resuspended at 1×106 cells/ml in binding buffer with
50 µl/ml FITC Annexin V and 50 µl/ml PI. The cells
were subsequently incubated for 15 min at room temperature, and
were then were analyzed using a Gallios flow cytometer (Beckman
Coulter, Atlanta, GA, USA). The cell inner mitochondrial membrane
potential (Δψm) was detected by flow cytometry using the MitoScreen
JC-1 staining kit (BD Biosciences), and was presented as the
protocol described below. Briefly, the cells were dissociated with
trypsin and resuspended at 1×106 cells/ml in Assay
Buffer, and then incubated at 37°C for 15 min with 10 µl/ml
JC-1. Before analyzed by flow cytometer, cells were washed twice by
Assay Buffer. Flow cytometry data were analyzed using FlowJo 7.6
software (TreeStar Inc., Ashland, OR, USA).
Caspase-9 or caspase-3 activity
assays
The activity of caspase-9 or caspase-3 was analyzed
by spectrophotometry using the Caspase-9 Colorimetric Assay kit or
Caspase-3 Colorimetric Assay kit (KeyGen BioTech, Nanjing, China),
and was presented as the protocol described below. Briefly,
5×106 cells or 100 mg fresh tumor tissues were washed
with cold PBS and re-suspended in lysis buffer and incubated on ice
for 30 min. Subsequently, 50 µl cell suspension was mixed
with 50 µl reaction buffer, and 5 µl caspase-3/-9
substrate, followed by incubation at 37°C for 4 h. The absorbance
was measured at 405 nm, and BCA protein quantitative analysis was
used as the reference to normalize each experimental group.
Establishment of tumor xenograft
model
For the establishment of a tumor xenograft model, 20
BALB/c-nu mice (6 weeks old, weighing 10–12 g) were purchased from
Hunan Silaikejingda Laboratory Animal Technology Co. (Hunan, China)
and randomly divided into 4 groups (n=5 per group) and the
indicated cells (1×106) were inoculated subcutaneously
into the inguinal folds of the nude mice. At 7 days after the cell
inoculation, the mice were injected intraperitoneally 15 mg/kg CDDP
twice per week for 5 weeks in one group of mice injected with the
vector cells and one group of mice injected with the
anti-miR-149-5p cells respectively. Tumor volume was determined
using an external caliper and calculated using the equation (L ×
W2)/2. On day 40, the animals were euthanized, and the
tumors were excised, weighed and stored in liquid nitrogen tanks.
For survival analyses, the mice were monitored daily for signs of
discomfort, and were either euthanized all at one time or
individually when presenting signs of distress, such as a 10% loss
of body weight, or head tilting. Overall survival was assessed by
the Kaplan-Meier method, using an exact log-rank test. Ethics
approval for the animal experiments was provided by The
Institutional Animal Care and Use Committee of Sun Yat-sen
University Cancer Center (ethics approval no. 102012016008I).
Luciferase assay
The cells (4×104) were seeded in
triplicate in 24-well plates and luciferase assay was carried out
as previously described (31). The
cells were transfected with 100 ng of HOP-flash reporter luciferase
plasmid, HIP-flash (HOP-flash mutant), pmirGLO-MST1-3′UTR or
-SAV1-3′UTR, luciferase plasmid, plus 5 ng pRL-TK Renilla
plasmid (Promega) using Lipofectamine 3000 (Invitrogen) according
to the manufacturer's instructions. Luciferase and Renilla
signals were measured at 36 h following transfection using a Dual
Luciferase Reporter Assay kit (Promega) according to the
manufacturer's instructions.
miRNA immunoprecipitation
The cells were co-transfected with the HA-Ago2
plasmid (10822; Addgene), followed by HA-Ago2 immunoprecipitation
using HA-antibody (Cat. no. H3663, Sigma-Aldrich) as previously
described (32). Real-time PCR
analysis of the IP material was used to test the association of the
mRNA of YAP1 and TAZ with the RISC complex.
Gene set enrichment analysis (GSEA)
The miRNA dataset of ovarian cancer from TCGA was
downloaded and we then procured the expression value of the
corresponding genes from the Level 3 data of each sample (the unit
was RNA-Seq by expectation maximization, RKPM). We then analyzed
the log2 value of each sample using Excel 2010 and GraphPad 5
software, as well as statistically analyzed the miRNA expression
levels of all ovarian cancer tissues using a paired t-test or
unpaired t-test. GSEA was performed with RNAseqV2 dataset of
ovarian cancer from TCGA as the MSigDB dataset. The high and low
expression level of miR-149-5p was stratified by the medium
expression level of miR-149-5p in ovarian cancer tissues. Gene set
analysis was performed by Molecular Signatures Database version
5.2.
Identification of potential targets of
miR-149-5p
As previously described (33,34),
the TargetScan (http://www.targetscan.org/vert_71/) and miRanda
(http://34.236.212.39/microrna/home.do) databases were used to
identify possible targets of miR-149-5p.
Statistical analysis
All values are presented as the means ± standard
deviation (SD). Significant differences were determined using
GraphPad 5.0 software (GraphPad Software, Inc., La Jolla, CA, USA).
One-way ANOVA with Tukey's post hoc test was used to determine
statistical differences between multiple groups. An unpaired or
paired t-test was used to determine statistical differences between
2 groups and confidence intervals, 95%. A value of p<0.05 was
considered to indicate a statistically significant difference. All
the experiments were repeated 3 times.
Results
miR-149-5p is upregulated in
chemoresistant ovarian cancer tissues
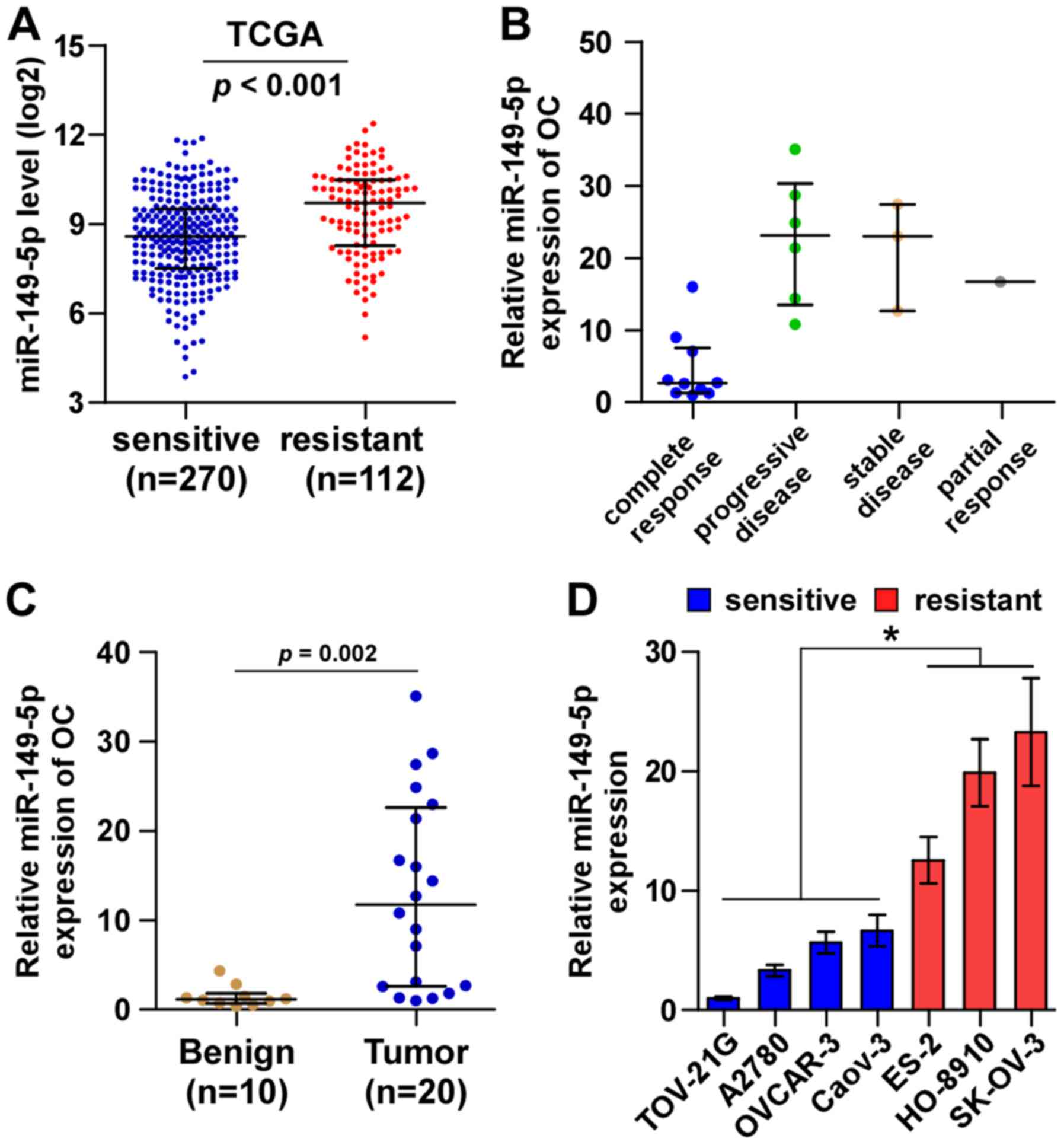
By analyzing TCGA ovarian cancer miRNA sequencing
datasets, we found that miR-149-5p expression was elevated in
chemoresistant ovarian cancer tissues compared with chemosensitive
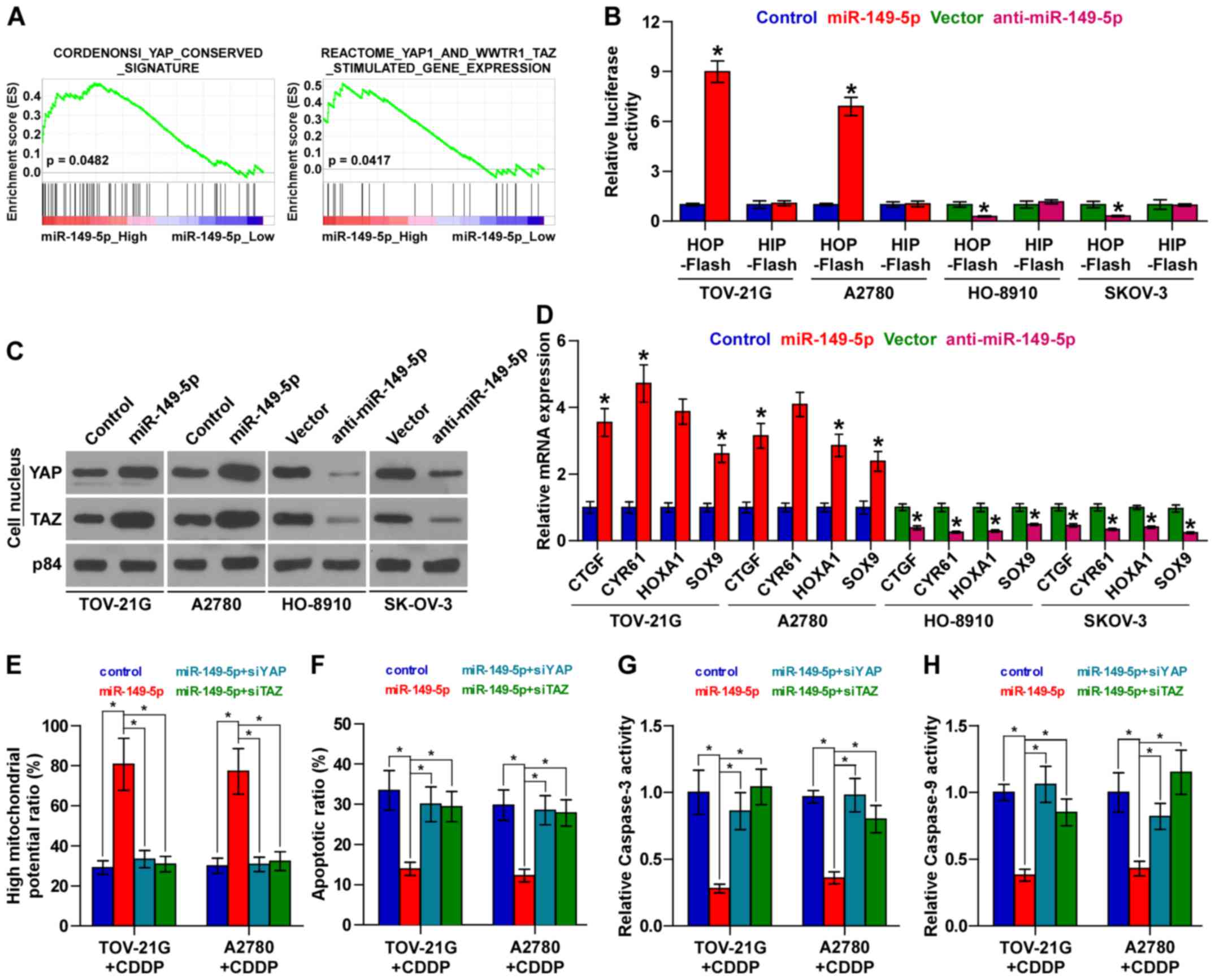
ovarian cancer tissues (Fig. 1A).
We further examined miR-149-5p expression in our own 20 ovarian
cancer tissues, including 10 complete response, 1 partial response,
3 stable disease and 6 progressive disease ovarian cancer tissues
perspectively. Consistently, we found that miR-149-5p expression in
the ovarian cancer tissues from patients with stable and
progressive disease was higher than that in ovarian cancer tissues
from patients with complete response (Fig. 1B). Furthermore, miR-149-5p
expression was markedly upregulated in ovarian cancer tissues
compared with benign ovarian disease tissues (Fig. 1C). The expression levels of
miR-149-5p in the ovarian cancer cell lines were evaluated and the
results revealed that compared with the chemosensitive ovarian
cancer cell lines (TOV-21G, A2780, OVCAR-3 and Caov-3), miR-149-5p
expression was differentially upregulated in the chemoresistant
ovarian cancer cell lines (ES-2, HO-8910, SK-OV-3) (Fig. 1C). Thus, these results suggest that
a high expression of miR-149-5p positively correlates with a poor
chemotherapeutic response in patients with ovarian cancer.
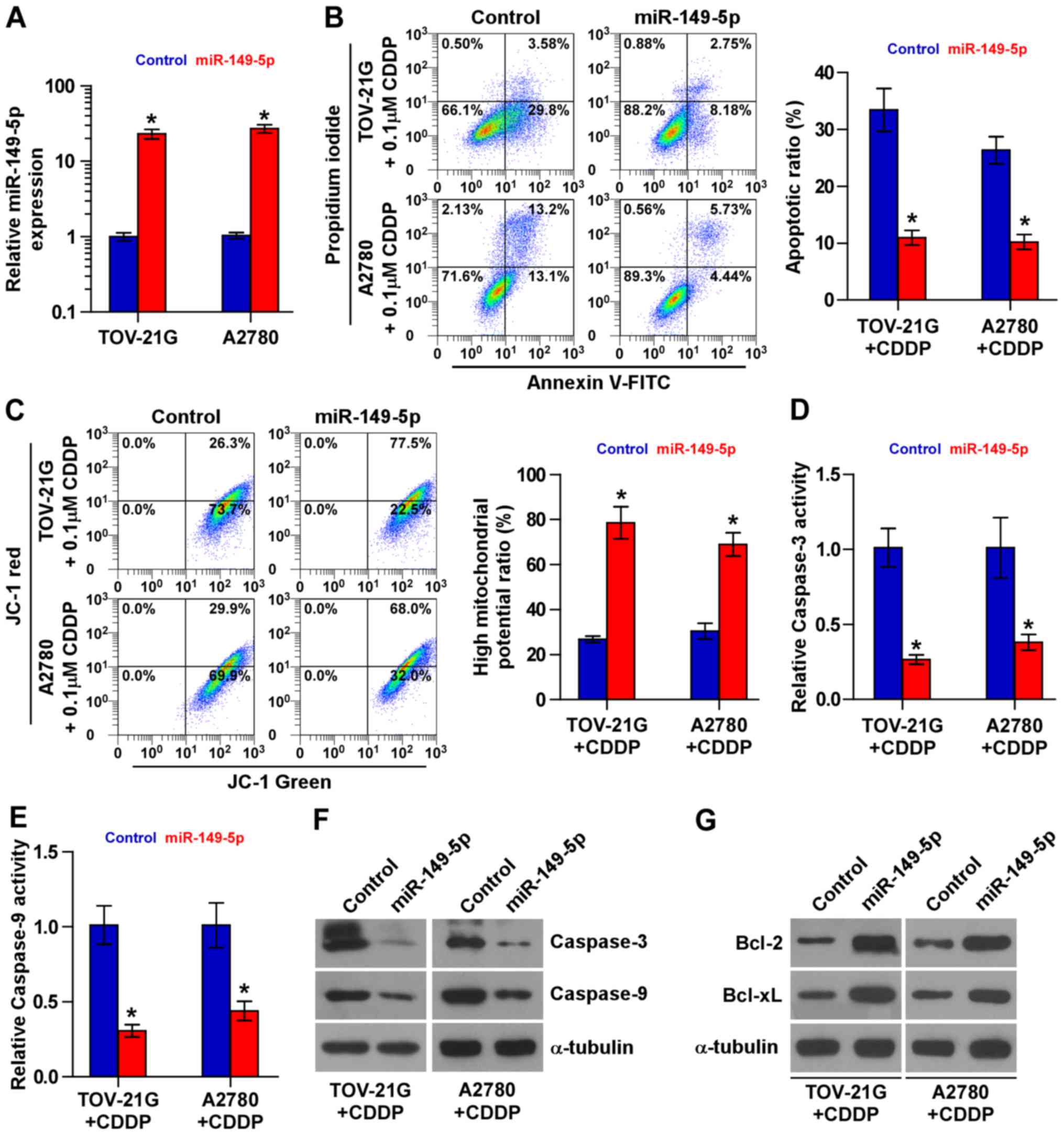
Upregulation of miR-149-5p enhances the
chemoresistance of ovarian cancer cells to CDDP
The role of miR-149-5p in the chemoresistance of
ovarian cancer was further examined. Based on the results of the
expression level of miR-149-5p shown in Fig. 1C, we decided to exogenously
overexpress miR-149-5p via viral transduction in the chemosensitive
TOV-21G and A2780 cells (Fig. 2A).
As shown in Fig. 2B, miR-149-5p
overexpression decreased the apoptotic rates of the TOV-21G and
A2780 cells treated with CDDP. The effects of miR-149-5p
overexpression on mitochondrial potential were further investigated
and the results revealed that miR-149-5p overexpression increased
the mitochondrial potential of the TOV-21G and A2780 cells treated
with CDDP (Fig. 2C). The effects
of the upregulation of miR-149-5p on apoptosis and caspase protein
expression were further confirmed by examining the activity of
caspase-9 and caspase-3, as well as the expression levels of the
anti-apoptotic proteins, Bcl-2 and Bcl-xL. The results revealed
that the overexpression of miR-149-5p reduced the activity and
expression of caspase-3 and caspase-9, and increased the expression
Bcl-2 and Bcl-xL (Fig. 2D–G).
Therefore, these results demonstrate that miR-149-5p promotes the
resistance of ovarian cancer cells to CDDP in vivo.
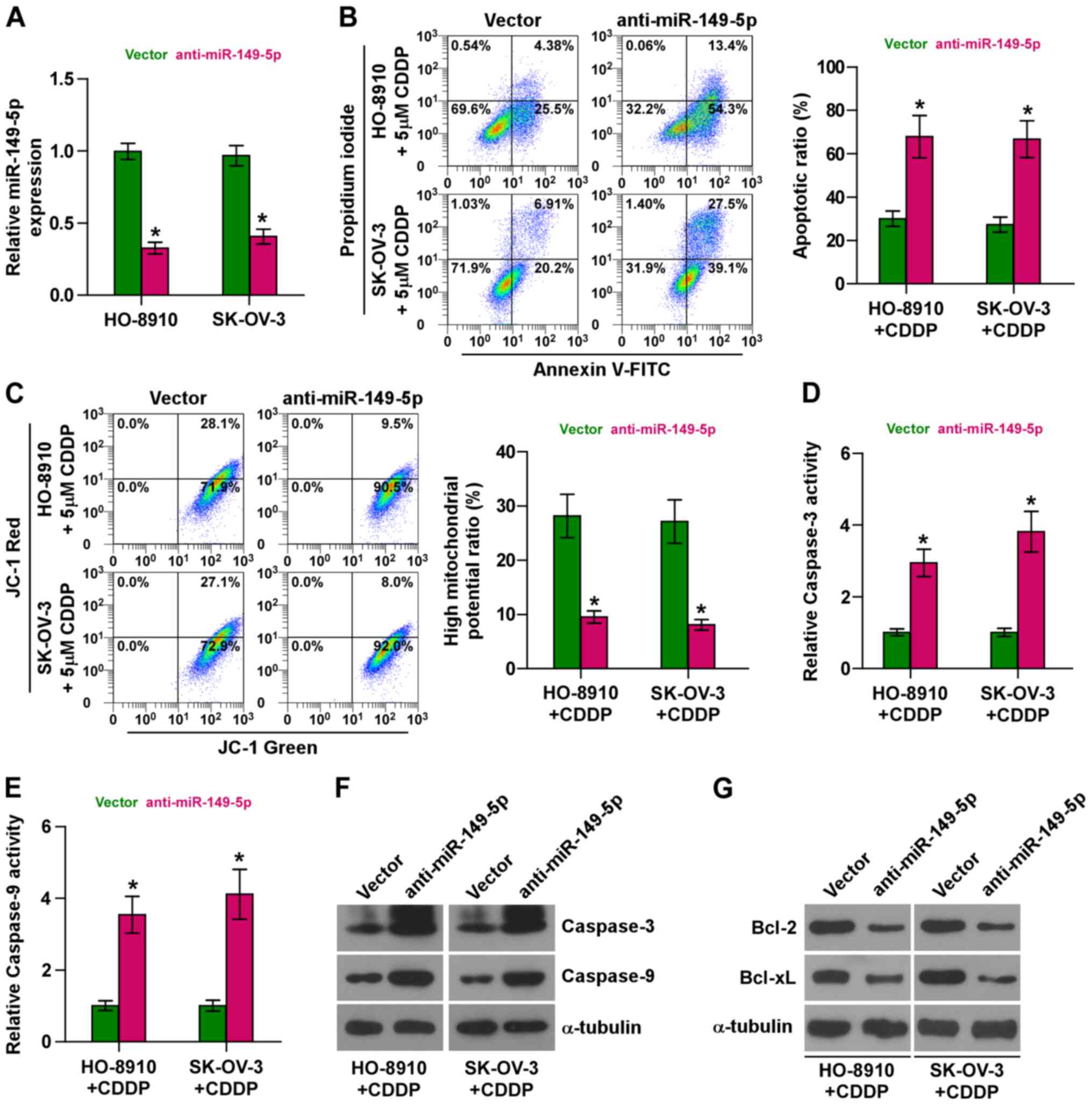
Silencing of miR-149-5p sensitizes
ovarian cancer cells to CDDP
We further endogenously silenced miR-149-5p by
transfecting anti-miR-149-5p into the chemoresistant HO-8910 and
SK-OV-3 cells (Fig. 3A) due to the
higher expression levels of miR-149-5p in the HO-8910 and SK-OV-3
cells (as shown in Fig. 1D). As
shown in Fig. 3B, the silencing of
miR-149-5p markedly increased the apoptotic rate of the HO-8910 and
SK-OV-3 cells treated with CDDP. Furthermore, the silencing of
miR-149-5p decreased the mitochondrial potential of the HO-8910 and
SK-OV-3 cells treated with CDDP (Fig.
3C). In addition, the silencing of miR-149-5p enhanced the
activity and expression of caspase-3 and caspase-9, and decreased
the expression levels of Bcl-2 and Bcl-xL (Fig. 3D–G). Collectively, these results
demonstrate that the silencing of miR-149-5p promotes the
resistance of ovarian cancer cells to CDDP.
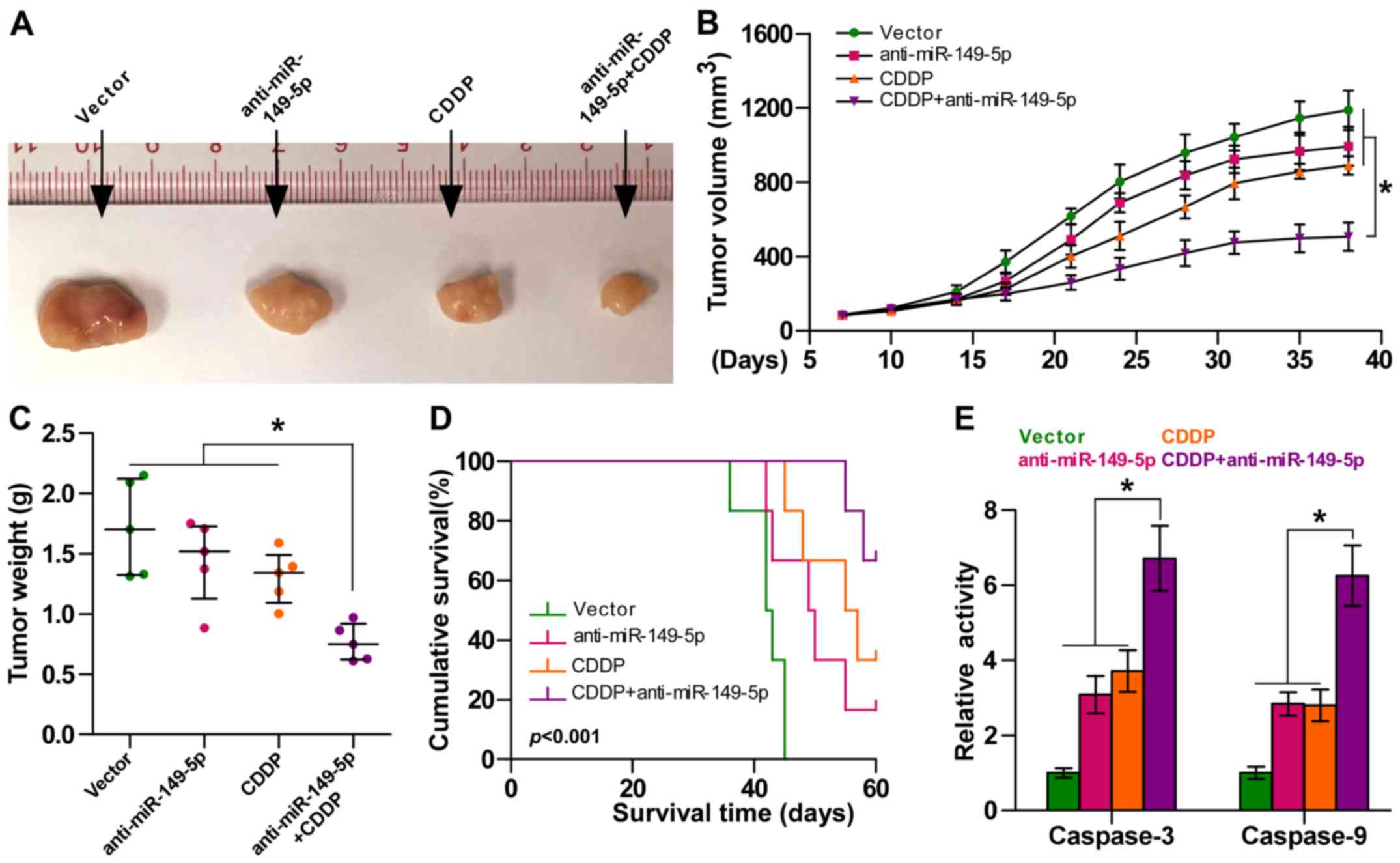
Silencing of miR-149-5p enhances the
chemosensitivity of ovarian cancer cells to CDDP in vivo
We further examined the effects of miR-149-5p on the
chemoresistance of ovarian cancer cells in vivo. Mice were
randomly divided into 4 groups (n=5/group). The vector cells were
inoculated subcutaneously into 2 groups of mice in the left dorsal
flank, and the other 2 groups of mice were inoculated with
anti-miR-149-5p cells (1×106 SK-OV-3 cells per mouse).
After 7 days, one group of mice injected with the vector cells and
one group of mice injected with the anti-miR-149-5p cells were
intraperitoneally injected with CDDP twice per week for 5 weeks
respectively [15 mg/kg (Fig. 4A)].
As shown in Fig. 4B and C, the
tumor volume and tumor weight were differentially decreased in the
anti-miR-149-5p group, CDDP treatment alone group and
anti-miR-149-5p plus CDDP group compared to the controls,
particularly in the anti-miR-149-5p plus CDDP group. Moreover, the
mice injected with anti-miR-149-5p-transfected cells in combination
with CDDP displayed a longer survival time (Fig. 4D). Caspase-3 and caspase-9
activities were markedly increased in the mice injected with
anti-miR-149-5p-transfected cells plus CDDP (Fig. 4E). Taken together, these findings
demonstrated that the silencing of miR-149-5p sensitized ovarian
cancer cells to CDDP in vivo.
miR-149-5p inactivates the Hippo
signaling pathway
To investigate the mechanisms underlying the
promoting effect of miR-149-5p on chemoresistance in ovarian
cancer, we performed a Gene set enrichment analysis (GSEA) of
miR-149-5p expression against the oncogenic signatures collection
of the MSigDB and found that miR-149-5p overexpression positively
correlated with the YAP and TAZ stimulated signatures (35) (Fig.
5A), suggesting that miR-149-5p regulates the Hippo signaling
pathway, which have been reported to play important roles in the
development of cancer chemoresistance (14,15).
As shown in Fig. 5B, we found that
the upregulation of miR-149-5p increased, while the silencing of
miR-149-5p decreased TEAD-dependent luciferase activity. Moreover,
the results of cellular fractionation and western blot analysis
revealed that the overexpression of miR-149-5p enhanced the nuclear
accumulation of YAP and TAZ, while the silencing of miR-149-5p
suppressed their nuclear expression (Fig. 5C). Real-time PCR analysis also
revealed that miR-149-5p overexpression significantly increased,
while the silencing of miR-149-5p inhibited the expression levels
of multiple downstream genes, including connective tissue growth
factor (CTGF), cysteine rich angiogenic inducer 61 (CYR61),
homeobox A1 (HOXA1) and SRY-box 9 (SOX9) in ovarian cancer cells
(Fig. 5D). Thus, these results
demonstrate that the downregulation of miR-149-5p inactivates the
Hippo signaling pathway in ovarian cancer cells.
YAP and TZA are essential for miR-149-5p
upregulation-induced chemoresistance in ovarian cancer cells
We then explored the functional significance of YAP
and TAZ in the chemoresistance of ovarian cancer cells. As shown in
Fig. 5E, the individual inhibition
of YAP or TAZ in miR-149-5p-overex-pressing cells abrogated the
stimulatory effects of miR-149-5p overexpression on mitochondrial
potential. Conversely, the inhibitory effects of miR-149-5p
upregulation on the apoptotic ratio, and caspase-3 and caspase-9
activities were reversed by the individual silencing of YAP and TAZ
in ovarian cancer cells (Fig.
5F–H). Taken together, our results indicate that YAP and TAZ
are functionally relevant effectors of miR-149-5p in the
chemotherapeutic response of ovarian cancer cells.
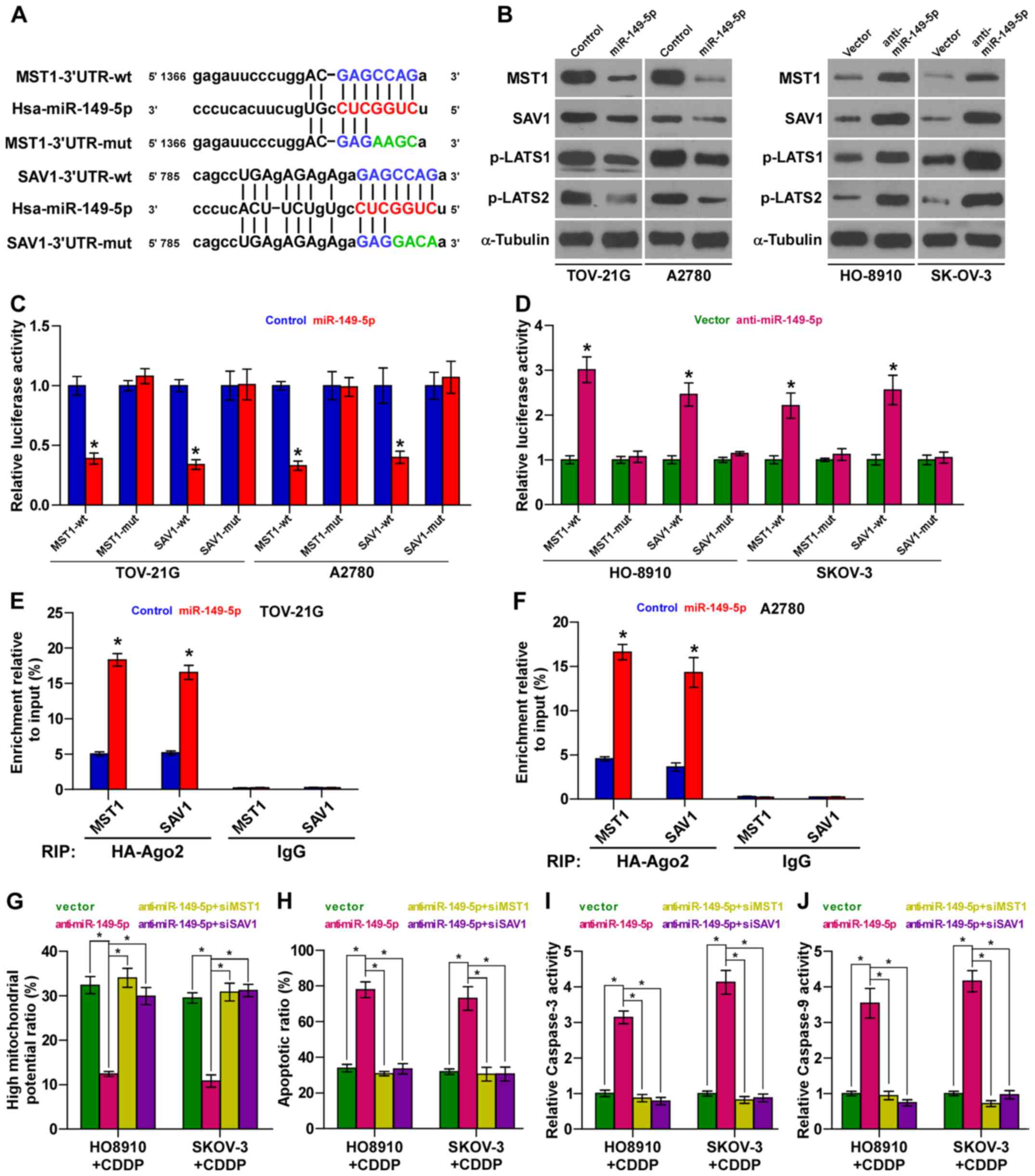
MST1 and SAV1 are authentic targets of
miR-149-5p in ovarian cancer cells
Using the publically available algorithms,
TargetScan and miRanda, we found that kinase components of Hippo
signaling, including MST1 and SAV1, may be potential targets of
miR-149-5p (Fig. 6A). Western blot
analysis revealed that miR-149-5p overexpression decreased the
expression levels of MST1 and SAV1; conversely, the silencing of
miR-149-5p increased their expression levels in the ovarian cancer
cells (Fig. 6B). Furthermore,
luciferase assay revealed that the upregulation of miR-149-5p
suppressed, while the silencing of miR-149-5p enhanced the reporter
activity driven by the 3′UTRs of the MST1 and SAV1 transcripts, but
not by the mutant 3′UTRs of these transcripts within
miR-149-5p-binding seed regions in ovarian cancer cells (Fig. 6C and D). Moreover,
microribonucleoprotein (miRNP) immunoprecipitation (IP) assay
revealed an association of miR-149-5p with MST1 and SAV1
transcripts (Fig. 6E and F),
further indicating the direct repressive effects of miR-149-5p on
MST1 and SAV1. Importantly, the stimulatory effects of
anti-miR-149-5p on the apoptotic ratio, and caspase-3 and caspase-9
activities were abrogated by the individual silencing of MST1 and
SAV1; however, the inhibitory effects of miR-149-5p silencing were
reversed by the individual silencing of MST1 and SAV1 in ovarian
cancer cells (Fig. 6G–J).
Collectively, our results indicate that miR-149-5p promotes the
chemoresistance of ovarian cancer cells by directly targeting MST1
and SAV1, leading to the inactivation of Hippo signaling.
Discussion
In the present study, our results revealed that
miR-149-5p expression was upregulated in chemoresistant ovarian
cancer tissues and cell lines compared with chemosensitive ovarian
cancer tissues and cell lines. The upregulation of miR-149-5p
enhanced, while the silencing of miR-149-5p inhibited the
chemoresistance of ovarian cancer cells to CDDP in vivo and
in vivo. Our results further revealed that miR-149-5p
directly suppressed MST1 and SAV1 expression, leading to the
inactivation of TEAD transcription and the downregulation of Hippo
downstream target genes. Therefore, our findings reveal a novel
biological role of miR-149-5p in the chemotherapeutic resistance of
ovarian cancer to CDDP.
The dysregulation of the Hippo signaling pathway has
been demonstrated to play an important role in the chemotherapeutic
resistance of cancer. YAP and TAZ are important downstream
effectors of the Hippo pathway and are suppressed by active Hippo
signaling (36,37). Studies have shown that the
upregulation of YAP and TAZ positively correlates with the
chemoresistance of several human cancer types (38,39).
Furthermore, the downregulation of the Hippo pathway core kinase
components has been reported to be implicated in chemotherapeutic
resistance of cancer cells to diverse chemotherapeutic drugs. For
example, the downregulation of MST1, which is mediated by heat
shock protein 70 (Hsp70) in a proteasomal-dependent manner,
promotes the CDDP resistance of prostate cancer cells (40). Additionally, the overexpression of
miR-181c contributes to chemoresistance in pancreatic cancer by
directly targeting MST1 and SAV1 (41). These studies indicate that the
inactivation of the Hippo signaling pathway, either the
upregulation of the downstream effectors, YAP and TAZ, or the
downregulation of core components of the Hippo signaling pathway,
confers chemotherapeutic resistance in human cancers. In this
study, our results demonstrated that a high expression of
miR-149-5p inactivated Hippo signaling by directly suppressing MST1
and SAV1, which further promoted the chemotherapeutic resistance of
ovarian cancer cells to CDDP. Importantly, the inhibitory effects
of anti-miR-149-5p on the chemoresistance of ovarian cancer cells
to CDDP were attenuated by the individual knockdown of MST1 and
SAV1. There finding indicate that miR-149-5p renders CDDP
chemoresistance in ovarian cancer cells by targeting MST1 and SAV1,
leading to the inactivation of Hippo signaling pathway.
The inactivation of Hippo signaling has been
extensively reported to be implicated in the initiation and
progression of ovarian cancer through various mechanisms. Fan et
al have reported that the disruption of Hippo signaling by
Sphingosine 1-phosphate (S1P) contributed to the development of
ovarian cancer by promoting unlimited cell proliferation (42). YAP, a major effector of the Hippo
signaling pathway, promotes the initiation and progression of
ovarian cancer by forming autocrine loops with ERBB signaling
(43). Furthermore, YAP confers
the resistance of ovarian cancer cells to chemotherapeutic agents,
and a high nuclear YAP expression strongly correlates with a poor
prognosis in patients with ovarian cancer (44,45).
These findings indicate that the disruption of Hippo signaling
plays an oncogenic role in ovarian cancer. In this study, our
results revealed that miR-149-5p inactivated Hippo signaling by
targeting MST1 and SAV1, which further rendered chemotherapeutic
resistance in ovarian cancer. Thus, our results present a novel
role of miR-149-5p in promoting the chemoresistance of ovarian
cancer cells to CDDP via the direct suppression of MST1 and SAV1,
leading to the inactivation of Hippo signaling.
There is evidence to indicate that miRNAs plays an
important role in regulating the activation of Hippo signaling via
the post-transcriptional suppression of multiple target mRNAs of
Hippo signaling components (46,47).
For example, miR-129-5p has been shown to inhibit proliferation and
survival by targeting the downstream co-activators, YAP and TAZ, of
Hippo signaling in ovarian cancer (48); moreover, the upregulation of
miR-130b has been shown to promote cancer stem cell characteristics
in glioblastoma cells by directly suppressing MST1 and SAV1
(49). These studies demonstrate
that the aberrant expression of miRNAs is involved in the
development and progression of cancer via the Hippo signaling
pathway. In this study, through bioinformatics analysis, we found
that the core kinase components of Hippo signaling are potential
targets of miR-149-5p. Furthermore, the upregulation of miR-149-5p
enhanced the tanscriptional activity of TEAD, nuclear translocation
of TAP and TAZ and expression levels of multiple downstream genes
of Hippo signaling pathway; conversely, silencing miR-149-5p
yielded an opposite effect. Collectively, our results uncover a
novel mechanism of the inactivation of the Hippo signaling pathway,
which further promotes the chemoresistance of ovarian cells to
CDDP.
The upregulation of miR-149-5p has been reported to
be implicated in the progression and metastasis of several
malignancies, including non-small cell lung cancer, squamous cell
carcinomas of the nasopharynx and prostate cancer, and a high
expression of miR-149-5p predicts a poor prognosis (50–52).
Furthermore, other studies have indicated that miR-149-5p
expression is decreased in hepatocellular carcinoma, breast cancer
and colorectal cancer (53–55).
These studies suggested that the pro- or anticancer roles of
miR-149-5p were tumor-type dependent. However, little is known
about the biological roles of miR-149-5p in ovarian cancer. In this
study, we found that miR-149-5p was markedly upregulated in
chemoresistant ovarian cancer tissues and cell lines compared with
chemo-sensitive ovarian cancer tissues and cell lines. Moreover,
the upregulation of miR-149-5p enhanced, while the silencing of
miR-149-5p inhibited the chemoresistance of ovarian cancer cells to
CDDP in vivo and in vivo. Our results further
revealed that miR-149-5p directly suppressed MST1 and SAV1
expression, leading to the inactivation of TEAD transcription and
the downregulation of nuclear YAP and TAZ and Hippo signaling
downstream target genes. Therefore, our results demonstrate that
miR-149-5p promotes the chemoresistance of ovarian cancer cells to
CDDP by inactivating the Hippo signaling pathway.
In conclusion, our study reveals that the
upregulation of miR-149-5p contributes to the chemoresistance of
ovarian cancer cells to CDDP as a pivotal suppressor of Hippo
signaling pathway. Thus, an in-depth understanding of the precise
role of miR-149-5p in the chemoresistance of ovarian cancer
faciliates to increase our knowledge of the development of
chemoresistance in ovarian cancer, which may aid in the development
of noveltherapeutic strategies against ovarian cancer.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (no. 81672561).
Notes
[1] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics, 2009. CA Cancer J Clin. 59:225–249.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cho KR and Shih IeM: Ovarian cancer. Annu
Rev Pathol. 4:287–313. 2009. View Article : Google Scholar :
|
|
3
|
Pan D: The hippo signaling pathway in
development and cancer. Dev Cell. 19:491–505. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Halder G and Johnson RL: Hippo signaling:
Growth control and beyond. Development. 138:9–22. 2011. View Article : Google Scholar :
|
|
5
|
Harvey KF, Zhang X and Thomas DM: The
Hippo pathway and human cancer. Nat Rev Cancer. 13:246–257. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lei QY, Zhang H, Zhao B, Zha ZY, Bai F,
Pei XH, Zhao S, Xiong Y and Guan KL: TAZ promotes cell
proliferation and epithelial-mesenchymal transition and is
inhibited by the hippo pathway. Mol Cell Biol. 28:2426–2436. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Oka T, Mazack V and Sudol M: Mst2 and Lats
kinases regulate apoptotic function of Yes kinase-associated
protein (YAP). J Biol Chem. 283:27534–27546. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao B, Ye X, Yu J, Li L, Li W, Li S, Yu
J, Lin JD, Wang CY, Chinnaiyan AM, et al: TEAD mediates
YAP-dependent gene induction and growth control. Genes Dev.
22:1962–1971. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang H, Liu CY, Zha ZY, Zhao B, Yao J,
Zhao S, Xiong Y, Lei QY and Guan KL: TEAD transcription factors
mediate the function of TAZ in cell growth and
epithelial-mesenchymal transition. J Biol Chem. 284:13355–13362.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chan SW, Lim CJ, Loo LS, Chong YF, Huang C
and Hong W: TEADs mediate nuclear retention of TAZ to promote
oncogenic transformation. J Biol Chem. 284:14347–14358. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shimomura T, Miyamura N, Hata S, Miura R,
Hirayama J and Nishina H: The PDZ-binding motif of Yes-associated
protein is required for its co-activation of TEAD-mediated CTGF
transcription and oncogenic cell transforming activity. Biochem
Biophys Res Commun. 443:917–923. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lu L, Li Y, Kim SM, Bossuyt W, Liu P, Qiu
Q, Wang Y, Halder G, Finegold MJ, Lee JS, et al: Hippo signaling is
a potent in vivo growth and tumor suppressor pathway in the
mammalian liver. Proc Natl Acad Sci USA. 107:1437–1442. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Imajo M, Miyatake K, Iimura A, Miyamoto A
and Nishida E: A molecular mechanism that links Hippo signalling to
the inhibition of Wnt/β-catenin signalling. EMBO J. 31:1109–1122.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lai D, Ho KC, Hao Y and Yang X: Taxol
resistance in breast cancer cells is mediated by the hippo pathway
component TAZ and its downstream transcriptional targets Cyr61 and
CTGF. Cancer Res. 71:2728–2738. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jeong W, Kim SB, Sohn BH, Park YY, Park
ES, Kim SC, Kim SS, Johnson RL, Birrer M, Bowtell DSL, et al:
Activation of YAP1 is associated with poor prognosis and response
to taxanes in ovarian cancer. Anticancer Res. 34:811–817.
2014.PubMed/NCBI
|
|
16
|
Bartel DP: Bartel. Cell. 136:215–233.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ren D, Wang M, Guo W, Huang S, Wang Z,
Zhao X, Du H, Song L and Peng X: Double-negative feedback loop
between ZEB2 and miR-145 regulates epithelial-mesenchymal
transition and stem cell properties in prostate cancer cells. Cell
Tissue Res. 358:763–778. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ren D, Wang M, Guo W, Zhao X, Tu X, Huang
S, Zou X and Peng X: Wild-type p53 suppresses the
epithelial-mesenchymal transition and stemness in PC-3 prostate
cancer cells by modulating miR-145. Int J Oncol. 42:1473–1481.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang X, Liu J, Zang D, Wu S, Liu A, Zhu
J, Wu G, Li J and Jiang L: Upregulation of miR-572
transcriptionally suppresses SOCS1 and p21 and contributes to human
ovarian cancer progression. Oncotarget. 6:15180–15193.
2015.PubMed/NCBI
|
|
20
|
Ren D, Yang Q, Dai Y, Guo W, Du H, Song L
and Peng X: Oncogenic miR-210 3p promotes prostate cancer cell EMT
and bone metastasis via NF-κB signaling pathway. Mol Cancer.
16:1172017. View Article : Google Scholar
|
|
21
|
Guo W, Ren D, Chen X, Tu X, Huang S, Wang
M, Song L, Zou X and Peng X: HEF1 promotes epithelial mesenchymal
transition and bone invasion in prostate cancer under the
regulation of microRNA-145. J Cell Biochem. 114:1606–1615. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dai Y, Ren D, Yang Q, Cui Y, Guo W, Lai Y,
Du H, Lin C, Li J, Song L, et al: The TGF-β signalling negative
regulator PICK1 represses prostate cancer metastasis to bone. Br J
Cancer. 117:685–694. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen X, Ba Y, Ma L, Cai X, Yin Y, Wang K,
Guo J, Zhang Y, Chen J, Guo X, et al: Characterization of microRNAs
in serum: A novel class of biomarkers for diagnosis of cancer and
other diseases. Cell Res. 18:997–1006. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bitarte N, Bandres E, Boni V, Zarate R,
Rodriguez J, Gonzalez-Huarriz M, Lopez I, Javier Sola J, Alonso MM,
Fortes P, et al: MicroRNA-451 is involved in the self-renewal,
tumorigenicity, and chemoresistance of colorectal cancer stem
cells. Stem Cells. 29:1661–1671. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang Y, Talmon G and Wang J: MicroRNA-587
antagonizes 5-FU-induced apoptosis and confers drug resistance by
regulating PPP2R1B expression in colorectal cancer. Cell Death Dis.
6:e18452015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ren D, Lin B, Zhang X, Peng Y, Ye Z, Ma Y,
Liang Y, Cao L, Li X, Li R, et al: Maintenance of cancer stemness
by miR-196b-5p contributes to chemoresistance of colorectal cancer
cells via activating STAT3 signaling pathway. Oncotarget.
8:49807–49823. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang M, Ren D, Guo W, Huang S, Wang Z, Li
Q, Du H, Song L and Peng X: N-cadherin promotes
epithelial-mesenchymal transition and cancer stem cell-like traits
via ErbB signaling in prostate cancer cells. Int J Oncol.
48:595–606. 2016. View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
29
|
Hahn WC, Dessain SK, Brooks MW, King JE,
Elenbaas B, Sabatini DM, DeCaprio JA and Weinberg RA: Enumeration
of the simian virus 40 early region elements necessary for human
cell transformation. Mol Cell Biol. 22:2111–2123. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang X, Ren D, Guo L, Wang L, Wu S, Lin
C, Ye L, Zhu J, Li J, Song L, et al: Thymosin beta 10 is a key
regulator of tumorigenesis and metastasis and a novel serum marker
in breast cancer. Breast Cancer Res. 19:152017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang X, Zhang L, Lin B, Chai X, Li R,
Liao Y, Deng X, Liu Q, Yang W, Cai Y, et al: Phospholipid
Phosphatase 4 promotes proliferation and tumorigenesis, and
activates Ca2+-permeable Cationic Channel in lung carcinoma cells.
Mol Cancer. 16:1472017. View Article : Google Scholar :
|
|
32
|
Li X, Liu F, Lin B, Luo H, Liu M, Wu J, Li
C, Li R, Zhang X, Zhou K, et al: miR-150 inhibits proliferation and
tumorigenicity via retarding G1/S phase transition in
nasopharyngeal carcinoma. Int J Oncol. Mar 10–2017.Epub ahead of
print. View Article : Google Scholar
|
|
33
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
eLife. 4:42015. View Article : Google Scholar
|
|
34
|
Betel D, Wilson M, Gabow A, Marks DS and
Sander C: The microRNA.org resource: Targets and expression.
Nucleic Acids Res. 36:Database. D149–D153. 2008. View Article : Google Scholar :
|
|
35
|
Cordenonsi M, Zanconato F, Azzolin L,
Forcato M, Rosato A, Frasson C, Inui M, Montagner M, Parenti AR,
Poletti A, et al: The Hippo transducer TAZ confers cancer stem
cell-related traits on breast cancer cells. Cell. 147:759–772.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Huang J, Wu S, Barrera J, Matthews K and
Pan D: The Hippo signaling pathway coordinately regulates cell
proliferation and apoptosis by inactivating Yorkie, the Drosophila
homolog of YAP. Cell. 122:421–434. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu CY, Zha ZY, Zhou X, Zhang H, Huang W,
Zhao D, Li T, Chan SW, Lim CJ, Hong W, et al: The hippo tumor
pathway promotes TAZ degradation by phosphorylating a phosphodegron
and recruiting the SCF{beta}-TrCP E3 ligase. J Biol Chem.
285:37159–37169. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huo X, Zhang Q, Liu AM, Tang C, Gong Y,
Bian J, Luk JM, Xu Z and Chen J: Overexpression of Yes-associated
protein confers doxorubicin resistance in hepatocellullar
carcinoma. Oncol Rep. 29:840–846. 2013. View Article : Google Scholar
|
|
39
|
Touil Y, Igoudjil W, Corvaisier M, Dessein
AF, Vandomme J, Monté D, Stechly L, Skrypek N, Langlois C, Grard G,
et al: Colon cancer cells escape 5FU chemotherapy-induced cell
death by entering stemness and quiescence associated with the
c-Yes/YAP axis. Clin Cancer Res. 20:837–846. 2014. View Article : Google Scholar
|
|
40
|
Ren A, Yan G, You B and Sun J:
Down-regulation of mammalian sterile 20-like kinase 1 by heat shock
protein 70 mediates cisplatin resistance in prostate cancer cells.
Cancer Res. 68:2266–2274. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen M, Wang M, Xu S, Guo X and Jiang J:
Upregulation of miR-181c contributes to chemoresistance in
pancreatic cancer by inactivating the Hippo signaling pathway.
Oncotarget. 6:44466–44479. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fan Q, Cheng Y, Chang HM, Deguchi M, Hsueh
AJ and Leung PCK: Sphingosine-1-phosphate promotes ovarian cancer
cell proliferation by disrupting Hippo signaling. Oncotarget.
8:27166–27176. 2017.PubMed/NCBI
|
|
43
|
He C, Lv X, Hua G, Lele SM, Remmenga S,
Dong J, Davis JS and Wang C: YAP forms autocrine loops with the
ERBB pathway to regulate ovarian cancer initiation and progression.
Oncogene. 34:6040–6054. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang X, George J, Deb S, Degoutin JL,
Takano EA, Fox SB and Bowtell DD: The Hippo pathway transcriptional
co-activator, YAP, is an ovarian cancer oncogene. Oncogene.
30:2810–2822. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hall CA, Wang R, Miao J, Oliva E, Shen X,
Wheeler T, Hilsenbeck SG, Orsulic S and Goode S: Hippo pathway
effector Yap is an ovarian cancer oncogene. Cancer Res.
70:8517–8525. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhang Y and Lai ZC: Mob as tumor
suppressor is regulated by bantam microRNA through a feedback loop
for tissue growth control. Biochem Biophys Res Commun. 439:438–442.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lin CW, Chang YL, Chang YC, Lin JC, Chen
CC, Pan SH, Wu CT, Chen HY, Yang SC, Hong TM, et al: MicroRNA-135b
promotes lung cancer metastasis by regulating multiple targets in
the Hippo pathway and LZTS1. Nat Commun. 4:18772013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tan G, Cao X, Dai Q, Zhang B, Huang J,
Xiong S, Zhang Y, Chen W, Yang J and Li H: A novel role for
microRNA-129-5p in inhibiting ovarian cancer cell proliferation and
survival via direct suppression of transcriptional co-activators
YAP and TAZ. Oncotarget. 6:8676–8686. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhu G, Wang Y, Mijiti M, Wang Z, Wu PF and
Jiafu D: Upregulation of miR-130b enhances stem cell-like phenotype
in glioblastoma by inactivating the Hippo signaling pathway.
Biochem Biophys Res Commun. 465:194–199. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lingzi X, Zhihua Y, Xuelian L, Yangwu R,
Haibo Z, Yuxia Z and Baosen Z: Genetic variants in microRNAs
predict non-small cell lung cancer prognosis in Chinese female
population in a prospective cohort study. Oncotarget.
7:83101–83114. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang C, Sturgis EM, Chen X, Zheng H, Wei Q
and Li G: Pre-miRNA variants as predictors of clinical outcome in
patients with squamous cell carcinomas of the nonoropharynx.
Oncotarget. 7:26444–26453. 2016.PubMed/NCBI
|
|
52
|
Zhu J, Wang S, Zhang W, Qiu J, Shan Y,
Yang D and Shen B: Screening key microRNAs for castration-resistant
prostate cancer based on miRNA/mRNA functional synergistic network.
Oncotarget. 6:43819–43830. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Luo G, Chao YL, Tang B, Li BS, Xiao YF,
Xie R, Wang SM, Wu YY, Dong H, Liu XD, et al: miR-149 represses
metastasis of hepatocellular carcinoma by targeting
actin-regulatory proteins PPM1F. Oncotarget. 6:37808–37823. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chan SH, Huang WC, Chang JW, Chang KJ, Kuo
WH, Wang MY, Lin KY, Uen YH, Hou MF, Lin CM, et al: MicroRNA-149
targets GIT1 to suppress integrin signaling and breast cancer
metastasis. Oncogene. 33:4496–4507. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Øster B, Linnet L, Christensen LL, Thorsen
K, Ongen H, Dermitzakis ET, Sandoval J, Moran S, Esteller M, Hansen
TF, et al COLOFOL steering group: Non-CpG island promoter
hypomethylation and miR-149 regulate the expression of SRPX2 in
colorectal cancer. Int J Cancer. 132:2303–2315. 2013. View Article : Google Scholar
|