Introduction
Diffuse large B cell lymphoma (DLBCL) is a common
and highly aggressive and heterogeneous subtype of non-Hodgkin's
lymphoma (NHL), which accounts for almost 40% of all NHL cases
(1). Despite combined
chemotherapy, patients with DLBCLs still have a 5-year mortality
rate of approximately 50% (2).
Rituximab is an anti-CD20 mouse-human chimeric monoclonal antibody,
which is known to exert a good curative effect in patients with
DLBCL. Rituximab combined with cyclophosphamide, adriamycin,
vincristine and prednisone (R-CHOP) is currently the standard
first-line treatment regimen for patients with DLBCL, resulting in
complete remission in approximately 80% of patients (3,4).
However, the widespread clinical use of rituximab has been
associated with increasing rituximab resistance (RR) in patients
with DLBCL, and previous studies have revealed that 30% of cases
are resistant to rituximab or rituximab-based chemotherapeutic
regimens (5,6). Furthermore, as previously
demonstrated, 60% of patients with lymphoma who previously
responded to rituximab failed to derive benefit when treated with
rituximab again, as a result of acquired drug resistance (7). Although some studies have
investigated the pathogenesis of RR (5-9), the
underlying mechanisms remain incompletely understood. It is
therefore crucial to explore the mechanisms responsible for RR in
order to improve the therapeutic outcomes of patients with
DLBCL.
Interleukin (IL)-17A, a member of the IL-17 family
(IL-17A-F), is an inflammatory cytokine that protects the body from
bacterial and fungal infections by recruiting neutrophils to the
site of inflammation (10). IL-17A
exerts its effects by binding to the IL-17 receptor (IL-17R)
(11). IL-17A is mainly secreted
by T helper (Th)17 cells, and the process is enhanced by IL-6
combined with transforming growth factor (TGF)-β (12). CD4+ T cells exhibit
considerable plasticity in terms of the differentiation and
development of Th1/Th2, Th1/Th17 and Th17/regulatory T (Treg) cells
(13).
IL-17+Foxp3+ Treg cells comprise a novel
subtype of CD4+ T cells that secrete IL-17A,
representing an intermediate differentiation stage between Th17 and
Treg cells (14).
IL-17+Foxp3+ Treg cells have recently been
shown to promote the growth of esophageal and colon cancers
(15,16).
DLBCL is known to be associated with cytokines in
the tumor microenvironment (17).
Th17 cells and IL-17A play important roles in tumor immune
regulation in NHL, and Yang et al found that the development
of Th17 cells and IL-17A was significantly decreased in the tumor
microenvironment in patients with NHL (18). Furthermore, a recent study
indicated that the frequency of Th17 cells and IL-17A levels in
peripheral blood (PB) were markedly lower in patients with DLBCL
compared with healthy individuals, and the differentiation of
circulating Th17 cells increased in relapsed patients with DLBCLs
(19). Another study verified that
IL-17A promoted the growth of human germinal center-derived NHL,
including DLBCL (20). We have
previously demonstrated that irradiated NHL cells (k1106 cells)
promoted Foxp3+ Treg cells to secrete IL-17 by
increasing the secretion of IL-6; secreted IL-17 then inhibited the
irradiated-induced apoptosis of NHL cells by suppressing p53
(21). IL-17A is thus a pro-tumor
factor in DLBCL.
Recently, published data have indicated that the
therapeutic use of kinase inhibitors targeting B-Raf
proto-oncogene, serine/threonine kinase (BRAF), ALK receptor
tyrosine kinase (ALK) or epidermal growth factor receptor (EGFR)
induces secretomes, which contribute to drug resistance (22). Some studies have also shown that
serum IL-6 levels in patients with NHL are elevated in vivo
by rituximab-based chemotherapeutic regimens (23,24).
IL-6 is known to promote the differentiation of Th17 cells, which
secrete IL-17A. Thus, we hypothesized that rituximab may induce
secretomes, such as IL-17A and IL-6, to promote RR in patients with
DLBCL, although the mechanisms through which rituximab affects
IL-17A secretion remain to be elucidated.
In the present study, our aim was to examine the
effects of rituximab on IL-17A and to investigate the role of
IL-17A in RR and its prognostic value in patients with DLBCL. We
retrospectively analyzed the effects of rituximab on Th17 and
IL-17+Foxp3+ Treg cell differentiation, and
IL-17A and IL-6 secretion in patients with DLBCL and in SU-DHL-4
cell co-cultures in vitro. IL-17 receptor knockdown and
wild-type SU-DHL4 cells were cultured with rituximab and IL-17A,
respectively, to determine the effects of IL-17A on apoptosis,
proliferation and p53 expression. We also analyzed the survival of
73 patients with DLBCL in order to evaluate the prognostic value of
IL-17A.
Materials and methods
Reagents
Antibodies against IL-6, p53 and IL-17R were
purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Human recombinant IL-6 (Cat. no. 206-IL) and IL-17A (Cat. no.
7955-IL), and human neutralizing antibodies to IL-6 (aIL-6) (Cat.
no. AF-206-NA) and IL-17A (aIL-17A) (Cat. no. AF-317-NA) were
obtained from R&D Systems (Minneapolis, MN, USA). Enzyme-linked
immunosorbent assay (ELISA) kits for IL-17A, IL-6, IL-21 and TGF-β
were purchased from BioLegend (San Diego, CA, USA).
Fluorescence-activated cell sorting (FACS) human antibodies,
including anti-CD3-phycoerythrin (PE)-Cy7 (Cat. no. 557851),
anti-CD8-allophycocyanin (Cat. no. 561952), anti-CD4-fluorescein
isothiocyanate (Cat. no. 561005), anti-Foxp3-PE (Cat. no. 560852),
anti-IL-17A-PE (Cat. no. 560436), anti-IL-17A-PE-Cy7 (Cat. no.
560799) and their matched anti-mouse IgG1 K-PE (Cat. no.
551436)/PE-Cy7 (Cat. no. 552811) were all purchased from BD
Biosciences (San Jose, CA, USA). Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) reagents were obtained from
Takara (Beijing, China). Rituximab was purchased from Novartis
(Basel, Switzerland).
Patients and sample collection
This study included 113 patients newly diagnosed
with DLBCL from January 2010 to December 2016 at Guangzhou First
People's Hospital, Guangzhou Medical University, and Sun Yat-Sen
University Cancer Center (Guangzhou, China). All patients with
DLBCL were diagnosed by experienced pathologists and the diagnosis
was consistent with DLBCL diagnostic criteria. In addition, 20
healthy individuals were selected as the controls. We collected PB
samples from all 113 patients and the 20 controls prior to
treatment, which were designated as the DLBCL group (untreated
group, n=113) and the Ctrl group (n=20). All the patients were then
treated with an R-CHOP or R-CHOP-like regimen (rituximab,
cyclophosphamide, doxorubicin, vincristine and prednisone), or with
a CHOP or CHOP-like regimen. Further PB samples were collected
after 2-4 cycles and the curative effects were evaluated as
complete remission (CR) or no complete remission (NCR). The 113
patients were then divided into 4 groups according to their
chemotherapeutic regimens and curative effects: The CHOP-CR group
(n=25), CHOP-NCR group (n=15), R-CHOP-CR group (n=56) and
R-CHOP-NCR group (n=17). With the inclusion of the Ctrl group and
the DLBCL group (untreated group, before treatment), in total,
there were 6 groups in this study. The baseline clinical
characteristics of all the groups are listed in Table I. The survival data of the 73
patients treated with the R-CHOP regimens were collected through
phone calls or clinical visits. Survival time was defined as the
period from diagnosis to the last visit, relapse or death. This
study was approved by the Ethics Committee of Guangzhou First
People's Hospital (2014-SYL-034). Written informed consent was
obtained from all participants or their families prior to obtaining
the samples.
 | Table IBaseline clinical characteristics of
the patients with DLBCL and the healthy individuals. |
Table I
Baseline clinical characteristics of
the patients with DLBCL and the healthy individuals.
| Variable | Patients with DLBCL
| Healthy individuals
|
|---|
| DLBCL | CHOP-CR | CHOP-NCR | R-CHOP-CR | R-CHOP-NCR | Ctrl |
|---|
| Age (years) | | | | | | |
| <50 | 43 | 10 | 7 | 19 | 7 | 7 |
| ≥50 | 70 | 15 | 8 | 37 | 10 | 13 |
| P-valuea | 0.957 | | | | | |
| Sex | | | | | | |
| Male | 66 | 14 | 10 | 31 | 11 | 14 |
| Female | 47 | 11 | 5 | 25 | 6 | 6 |
| P-valuea | 0.849 | | | | | |
| Ann Arbor
stageb | | | | | | |
| I–II | 45 | 9 | 6 | 22 | 8 | / |
| III–IV | 68 | 16 | 9 | 34 | 9 | / |
| P-valuea | 0.970 | | | | | |
| IPI scorec | | | | | | |
| 1–3 | 64 | 13 | 10 | 29 | 12 | / |
| 4–5 | 49 | 12 | 5 | 27 | 5 | / |
| P-valuea | 0.606 | | | | | |
Cell lines and cell culture
The SU-DHL-4 cell line was purchased from ATCC
(Manassas, VA, USA) and cultured in RPMI-1640 medium (HyClone,
South Logan, UT, USA) containing 10% fetal bovine serum (FBS)
(HyClone), 4 mM L-glutamine (Sigma-Aldrich, St. Louis, MO, USA),
100 U/ml of penicillin (HyClone), and 100 U/ml of streptomycin
(HyClone). 293T cells were purchased from ATCC and cultured in
Dulbecco's modified Eagle's medium (HyClone) supplemented with 10%
FBS. All the cells were cultured in a humidified chamber at 37°C
with an atmosphere of 5% CO2.
Cell cultures in vitro
We conducted 4 cell culture experiments in
vitro. All the experiments were carried out using SU-DHL-4
cells and PB mononuclear cells (PBMCs) at 2×106
cells/well. PBMCs were isolated using the Ficoll–Hypaque method.
Experiment 1. The SU-DHL-4 cells were cultured with various
concentrations of rituximab (0.0, 0.1, 1.0, 10.0 and 100.0
μg/ml) in a 6-well plate, and then incubated for 72 h. The
IL-6 mRNA and protein levels were analyzed by RT-qPCR), and by
ELISA and western blot analysis, respectively.
Experiment 2
The PBMCs from the healthy donors were cultured in T
cell medium containing 10% FBS and 100 IU/ml human IL-2 (HyClone)
in a 48-well plate, and stimulated with OKT3 (1 μg/ml,
HyClone). The cells were then divided into 7 groups as follows:
PBMCs alone; PBMCs + 100 μg/ml rituximab; PBMCs + rituximab
+ 1 μg/ml IL-6; PBMCs + SU-DHL-4 cells; PBMCs + rituximab +
SU-DHL-4 cells; PBMCs + rituximab + SU-DHL-4 + IL-6; and PBMCs +
rituximab + SU-DHL-4 + 1 μg/ml aIL-6. All the groups of
cells were incubated for 72 h and then detected by ELISA and FACS
analysis.
Experiment 3
This included 6 groups of cells cultured for 72 h as
follows: SU-DHL-4 cells alone; SU-DHL-4 + 100 μg/ml
rituximab; SU-DHL-4 + rituximab + PBMCs; SU-DHL-4 cells + rituximab
+ PBMCs + 1 μg/ml aIL-17A; SU-DHL-4 cells + rituximab + 1
μg/ml IL-17A; and SU-DHL-4 cells + IL-17A.
Experiment 4
IL-17 receptor knockdown (IL-17R-KD) and wild-type
SU-DHL4 cells were cultured with 100 μg/ml anti-CD20
monoclonal antibody in the presence of various concentrations of
IL-17A (0-10 ng/ml). The cells were then collected and analyzed by
FACS analysis and Cell Counting kit-8 (CCK-8) assay.
Cell viability assay
Cell viability was assessed by CCK-8 assay (Dojindo
Laboratories, Kumamoto, Japan). Each sample was allocated in 6
wells and CCK-8 was added 4 h before the end of the culture time.
Wells without cells were set as the blank controls. The absorbance
at 450 nm was measured using a Universal Microplate
Spectrophotometer (Thermo Fisher Scientific, Inc., Waltham, MA,
USA). The relative proliferation rate (%) of the SU-DHL-4 cells was
calculated as follows: [optical density of cancer cell lines at 450
(OD450) with IL-17A/OD450 of cancer cell lines without IL-17A]
×100%.
RT-qPCR
Total RNA was isolated from the cells using TRIzol
reagent (Takara) according to the manufacturer's instructions. RNA
was reverse transcribed into cDNA using PrimeScript RT Master Mix
(Takara) according to the manufacturer's instructions. qPCR was
performed using SYBR Premix Ex Taq II (Tli RNase H Plus) (Takara)
on a LightCycler 480II system (Roche, Mannheim, Germany). The
levels of IL-6 and p53 were normalized to those of
glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The primers
(5′→3′) used for qPCR were as follows: IL-6 forward,
ACTTCGTGCATGACTTCAGC and reverse, TCTTTGTTGGAGGGTGAGGG; p53
forward, TGGCCATCTACAAGCAGTCA and reverse, GGTACAGTCAGAGCCAACCT;
and GAPDH forward, GCACCGTCAAGGCTGAGAAC and reverse,
TGGTGAAGACGCCAGTGGA. The thermocycling conditions were as follows:
Initial denaturation 94°C for 5 min, denaturation 94°C for 15 sec,
annealing 59°C for 30 sec, extension 72°C for 2 min 55 sec, and
then run for 35 cycles.
Western blot analysis
The cells were lysed using sodium dodecyl sulfate
buffer containing proteinase inhibitors (Roche). Equal amounts of
protein (50 μg) were separated by 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred onto
polyvinylidene difluoride membranes (Bio-Rad Laboratories,
Shanghai, China). The membranes were blocked and incubated with
antibodies against GAPDH (dilution 1:200, Cat. no. sc-47724), IL-6
(1:200, Cat. no. sc-65327), p53 (1:200, Cat. no. sc-47698) and
IL-17R (1:100, Cat. no. sc-376374) overnight at 4°C. Antibodies
against GAPDH, IL-6, p53 and IL-17R were purchased from Santa Cruz
Biotechnology. The membranes were then incubated with horseradish
peroxidase-labeled secondary antibody (1:500, Cat. no. sc-2031)
(Santa Cruz Biotechnology). The protein bands were visualized using
an enhanced chemiluminescence reagent.
Flow cytometry
The frequencies of Th17 cells and
IL-17+Foxp3+ Treg cells were detected by flow
cytometry. Cell density was adjusted to 2×106/ml. For
the Th17 cells, the cells were stimulated by the addition of 50
ng/ml phorbol myristate acetate, 1 μg/ml ionomycin and 10
μg/ml Brefeldin A to the medium for 5 h at 37°C and 5%
CO2. The cells were then stained with
fluorescence-labeled anti-CD3, anti-CD8 and anti-IL-17A antibodies
for 1 h at room temperature. For the
IL-17+Foxp3+ Treg cells, the cells were
stained with fluorescence-labeled anti-IL-17A, anti-CD4, and
anti-Foxp3 antibodies for 1 h at room temperature. The cells were
then analyzed using a flow cytometer (FACSCanto II; BD Biosciences)
and the data were analyzed with FlowJo software.
CD3+CD8−IL-17A+ cells were defined
as Th17 cells, and
CD4+IL-17A+Foxp3+ cells were
defined as IL-17+Foxp3+ Tregs.
ELISA
Serum from patient PB or the supernatants from cell
cultures were assessed for IL-17A, IL-21, IL-6 and TGF-β levels by
ELISA, following the manufacturer's instructions.
Knockdown of IL-17R expression in
SU-DHL-4 cells
IL-17R-specific short hairpin RNAs (shRNAs) were
delivered by lentiviral infection to knock down IL-17R expression
in the SU-DHL cells. The RNAi Consortium human IL-17R shRNA and
Trans-Lentiviral shRNA Packaging kit with Calcium Phosphate
Transfection Reagent (including pGIPZ™ Non-silencing Control Vector
DNA) were purchased from Dharmacon (Lafayette, CO, USA). The IL-17R
shRNA sequence was (5′→3′) GGCTAAACTGCACGGTCAAGAC. The
IL-17R-specific shRNA was cloned into the pLKO.1 puro plasmids.
Briefly, the 293T cells were seeded at a density of
5.5×106 cells/14 ml in a 100-mm plate, as described in
our previous study (25). The
following day, 42 μg shRNA plasmid was transfected into the
293T cells, together with 30 μl of the Trans-Lentiviral
Packaging kit (including 5 plasmids: pTLA1-PAK, pTLA1-ENZ,
pTLA1-ENV, pTLA1-TOFF and pTLA1-TAT/REV) using calcium phosphate
(105 μl). At 48 h after transfection, the supernatant
containing lentiviral particles was harvested and filtered through
a 0.45 μm-diameter filter, and used to infect the SU-DHL-4
cells. The cells were selected using puromycin. The protein levels
of IL-17R were evaluated by western blot analysis.
Assessment of apoptosis
The SU-DHL-4 cells from experiment 4 describe above
were stained with propidium iodide (PI) and Annexin V-FITC reagent
(BD Biosciences) following the manufacturer's instructions. The
cells were then analyzed using flow cytometry. Annexin
V+PI− cells were determined as apoptotic
cells.
Statistical analysis
All analyses were performed using SPSS 17.0
software. Numerical data are presented as the means ± standard
deviation. Single-factor analysis of variance (one-way ANOVA),
Student-Newman-Keuls/Dunnett's T3 tests (between each 2 set of
groups) were used for comparisons among multiple groups. The
Chi-square test was used for categorical data comparisons between 2
groups. Survival was estimated using the Kaplan-Meier method and
the log-rank test. A P-value of 0.05 was considered to indicate a
statistically significant difference.
Results
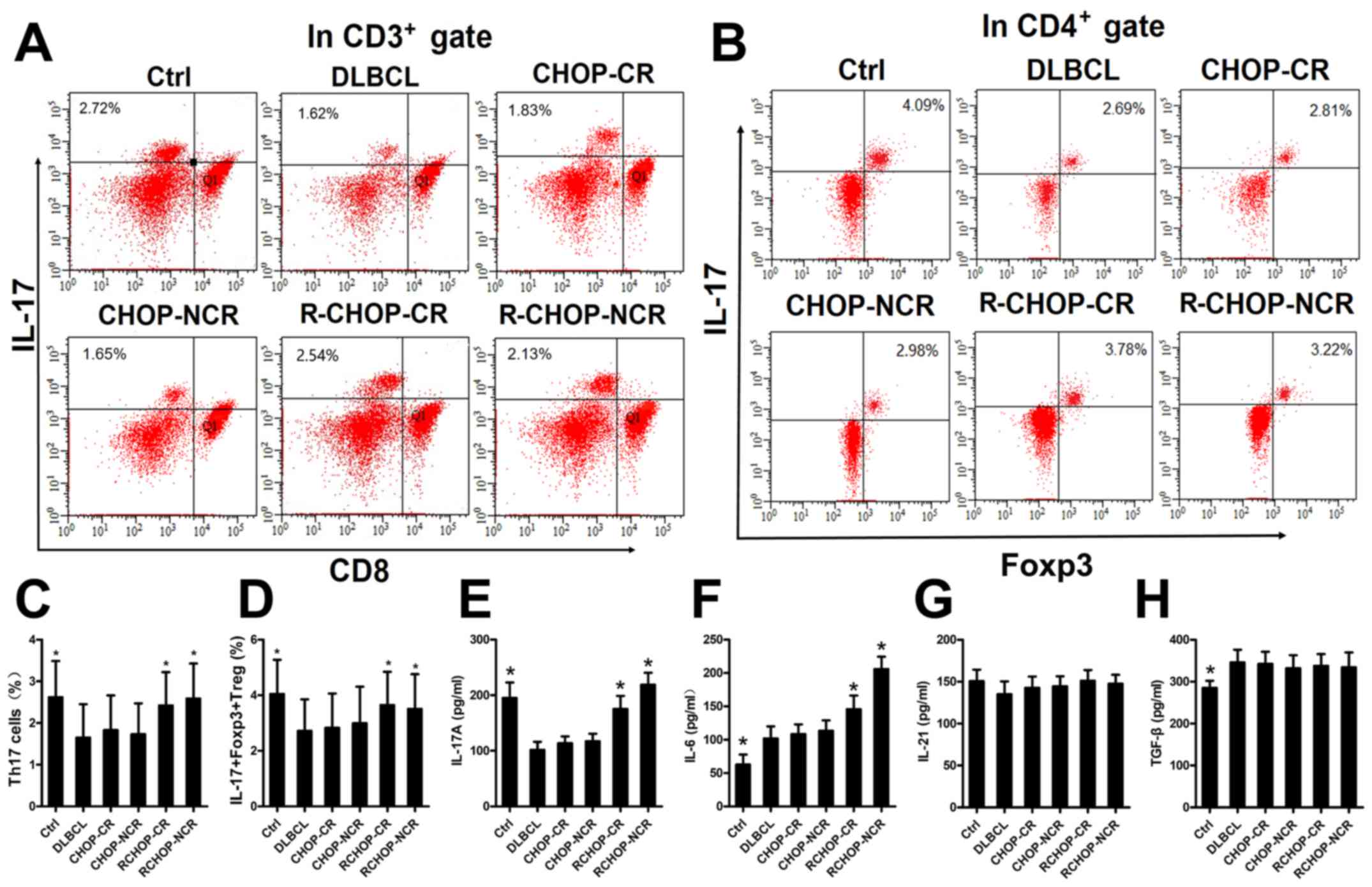
Rituximab increases IL-17A levels and the
proportions of Th17 and IL-17+Foxp3+ Treg
cells in the PBMCs from patients with DLBCL
We examined the mechanisms through which rituximab
affects the expression of IL-17A and its two associated secretory
cells in patients with DLBCL by detecting the IL-17A levels and the
proportions of Th17 and IL-17+Foxp3+ Treg
cells in the PBMCs by ELISA and FACS analysis, respectively. A
total of 113 patients newly diagnosed with DLBCL and 20 healthy
donors were divided into 6 groups, as described in the Materials
and methods. Representative FACS plots of the Th17 and
IL-17+Foxp3+ Treg cells from each group are
shown in Fig. 1A and B. The IL-17A
levels and the proportions of Th17 and
IL-17+Foxp3+ Treg cells were significantly
higher in the Ctrl, R-CHOP-CR and R-CHOP-NCR groups, compared with
the DLBCL, CHOP-CR and CHOP-NCR groups (Fig. 1C-E), implying that rituximab
elevated the IL-17A levels and the differentiation of its two
associated secretory cells. The IL-6 levels in the R-CHOP-CR and
R-CHOP-NCR groups were also significantly increased compared with
those in the DLBCL, CHOP-CR and CHOP-NCR groups (Fig. 1F). The TGF-β levels were higher in
the DLBCL group than in the Ctrl group (Fig. 1H). Rituximab had no significant
effect on the IL-21 and TGF-β levels in any group (Fig. 1G and H).
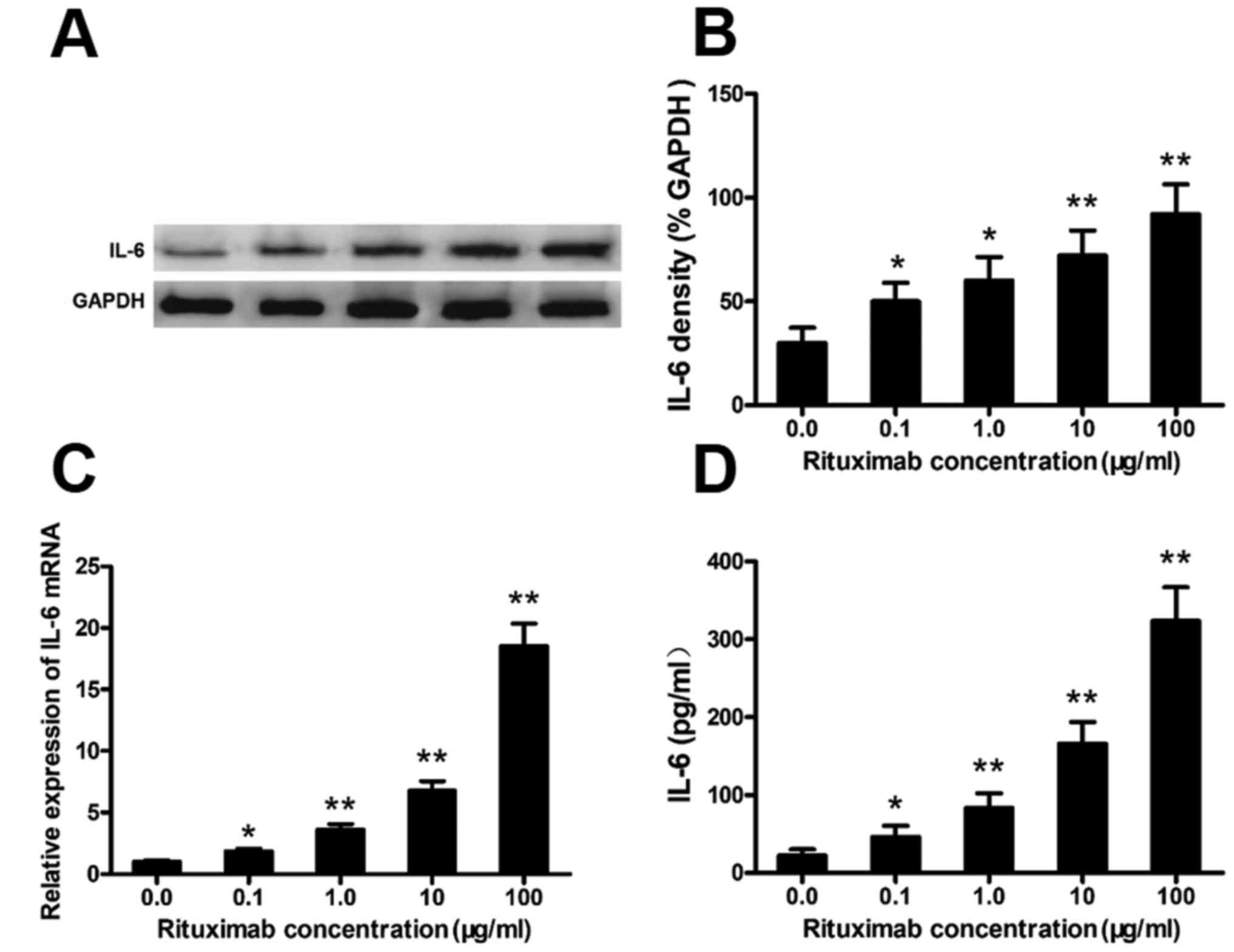
Rituximab increases IL-6 expression in
DLBCL cell lines
Rituximab increased IL-6 expression in the PBMCs
from patients with DLBCL (Fig. 1).
To confirm this phenomenon and the cell source, we cultured the
SU-DHL-4 cells with various concentrations of rituximab and
analyzed the expression of IL-6 in the SU-DHL-4 cells. Rituximab
markedly increased the protein and mRNA levels of IL-6 in the
SU-DHL-4 cells in a concentration-dependent manner (Fig. 2A–C). Typical western blots of IL-6
protein are shown in Fig. 2A. IL-6
in the culture supernatants was also significantly elevated in a
rituximab concentration-dependent manner, as detected by ELISA
(Fig. 2D).
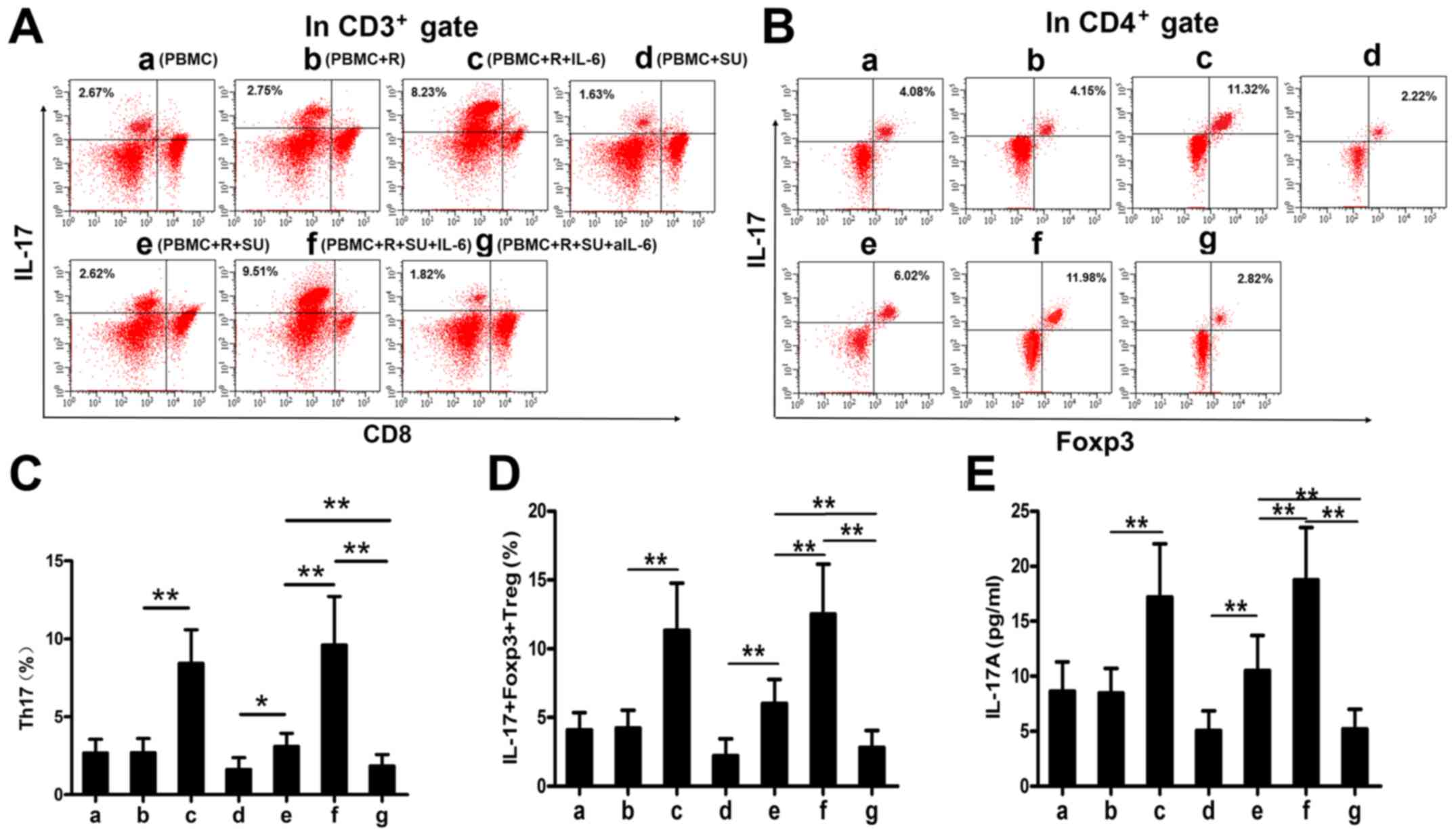
Rituximab upregulates IL-17A levels and
the proportions of Th17 and IL-17+Foxp3+ Treg
cells in vitro
IL-6 enhances the differentiation of Th17 cells and
IL-17+Foxp3+ Treg cells together with TGF-β
(12). As demonstrated above,
rituximab elevated IL-6 expression in DLBCL cells and in PBMCs from
patients with DLBCL, and that TGF-β levels were increased in
patients with DLBCL (Fig. 1H). We
therefore hypothesized that rituximab would increase the
proportions of Th17 and IL-17+Foxp3+ Treg
cells and upregulate the IL-17A levels in the SU-DHL-4 cells and
PBMCs in vitro. To examine this hypothesis, we cultured the
cells as described in the Materials and methods and analyzed the
cells and supernatants by FACS analysis and ELISA. Representative
FACS plots of Th17 and IL-17+Foxp3+ Treg
cells from each group are shown in Fig. 3A and B. Rituximab had no effect on
the proportions of Th17 and IL-17+Foxp3+ Treg
cells or on the IL-17A levels in the PBMCs cultured without the
SU-DHL-4 cells (Fig. 3C-E, bars a
and b); however, both secretory cell populations and IL-17A
expression levels were increased in the presence of the SU-DHL-4
cells (Fig. 3C-E, bars d and e).
This upregulation was significantly enhanced by exogenous IL-6
(Fig. 3C, bars c and e; D, bars c
and f; and E, bars c and f) and abolished by aIL-6 (Fig. 3C-E, bar g).
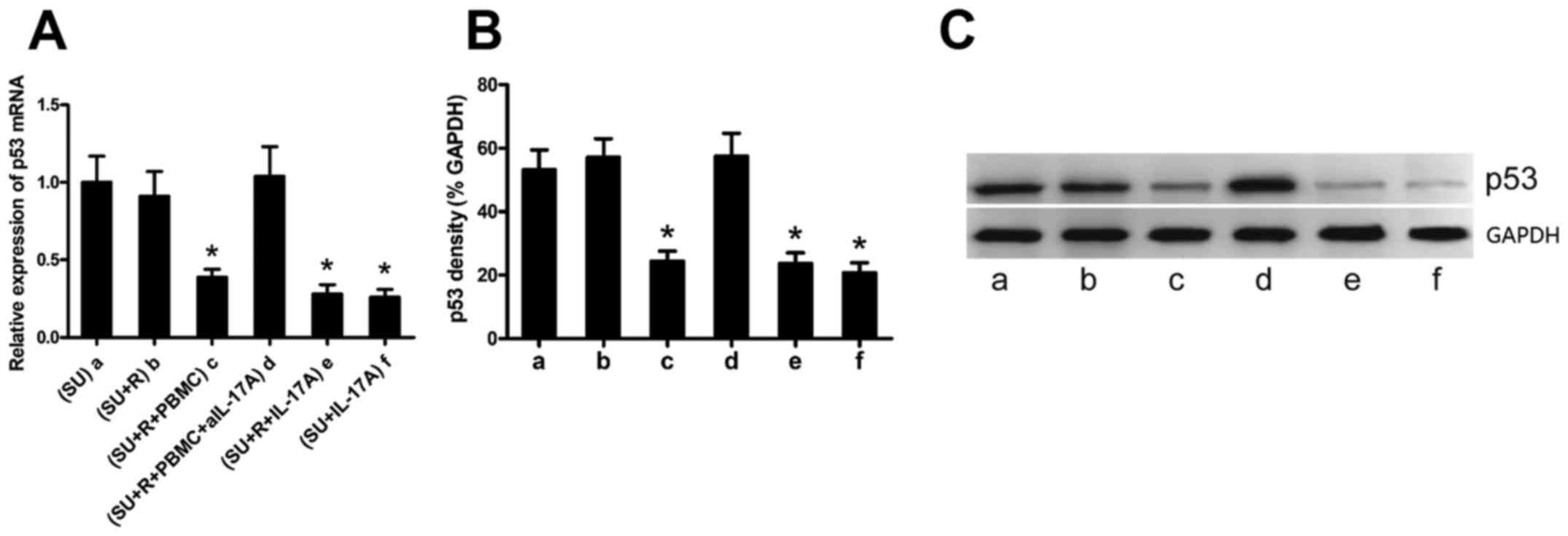
IL-17A suppresses p53 expression in the
DLBCL cell lines
IL-17A has been reported to promote the growth of
DLBCL (20), although the
mechanism involved remain elusive. p53 is a tumor suppressor
protein, and we have previously demonstrated that IL-17
suppressesp53 expression and induces the radiation resistance of
k1106 B cell lymphoma cells (21).
We thus hypothesized that rituximab-induced IL-17A expression may
suppress p53 expression in the SU-DHL-4 cells. Co-culture
experiments demonstrated that rituximab alone did not affect p53
expression in the SU-DHL-4 cells (Fig.
4A and B, bars a and b); however, p53 expression was
significantly decreased in the SU-DHL-4 cells co-cultured with
PBMCs and rituximab (Fig. 4A and
B, bar c). To determine whether IL-17A is required for this
p53-suppressing process, we added a neutralizing anti-IL-17A
antibody to the co-cultures, which resulted in a significantly
increased p53 expression in the SU-DHL-4 cells (Fig. 4 and B, bar d). By contrast,
exogenous IL-17A significantly decreased the expression of p53 in
the SU-DHL-4 cells (Fig. 4A and B,
bars e and f). Representative western blots of p53 protein are
shown in Fig. 4C.
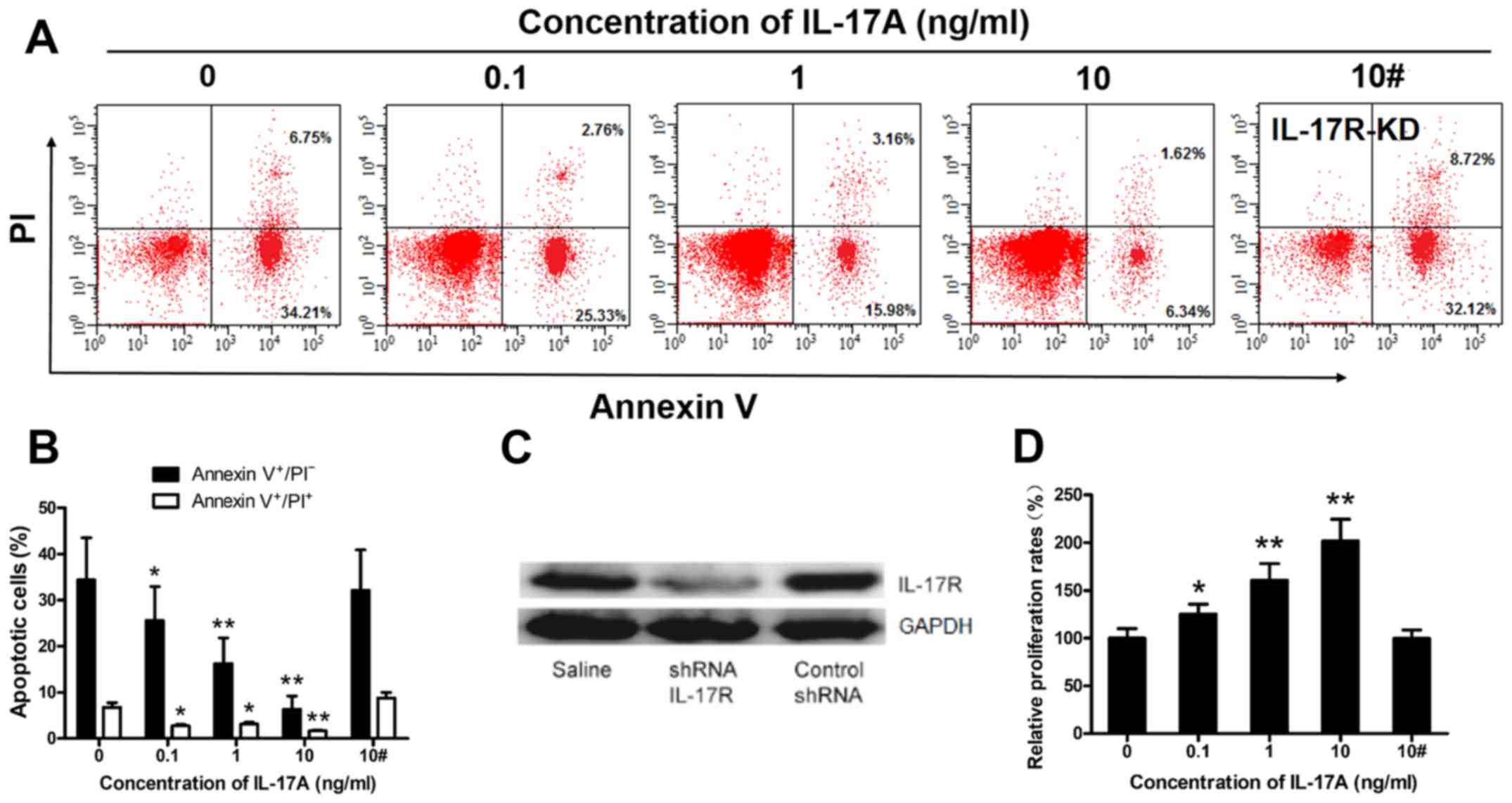
IL-17A prevents rituximab-induced
SU-DHL-4 cell apoptosis and promotes SU-DHL-4 cell
proliferation
The above-mentioned results suggest that IL-17A may
promote SU-DHL-4 cell growth (Fig.
4), in accordance with previous findings (20). To examine this hypothesis, we
cultured IL-17R-KD and wild-type SU-DHL4 cells with rituximab in
the presence of various concentrations of IL-17A (0-10 ng/ml), and
then analyzed the cells by FACS analysis and CCK-8 assay. The
frequency of apoptotic wild-type SU-DHL-4 cells was markedly
suppressed by IL-17A in a concentration-dependent manner (Fig. 5A and B). We successfully knocked
down IL-17R expression in the SU-DHL-4 cells using
lentiviral-mediated shRNA, resulting in the significant
down-regulation of IL-17R protein levels in the SU-DHL-4 cells
(Fig. 5C). IL-17A (10 ng/ml) had
no effect on the frequency of apoptotic IL-17R-KD SU-DHL4 cells
(Fig. 5A and B) compared to the
control (10 ng/ml IL-17A). Furthermore, the proliferation rate of
wild-type SU-DHL-4 cells, but not that of IL-17R-KD SU-DHL4 cells,
was increased in an concentration-dependent manner by IL-17A
(Fig. 5D).
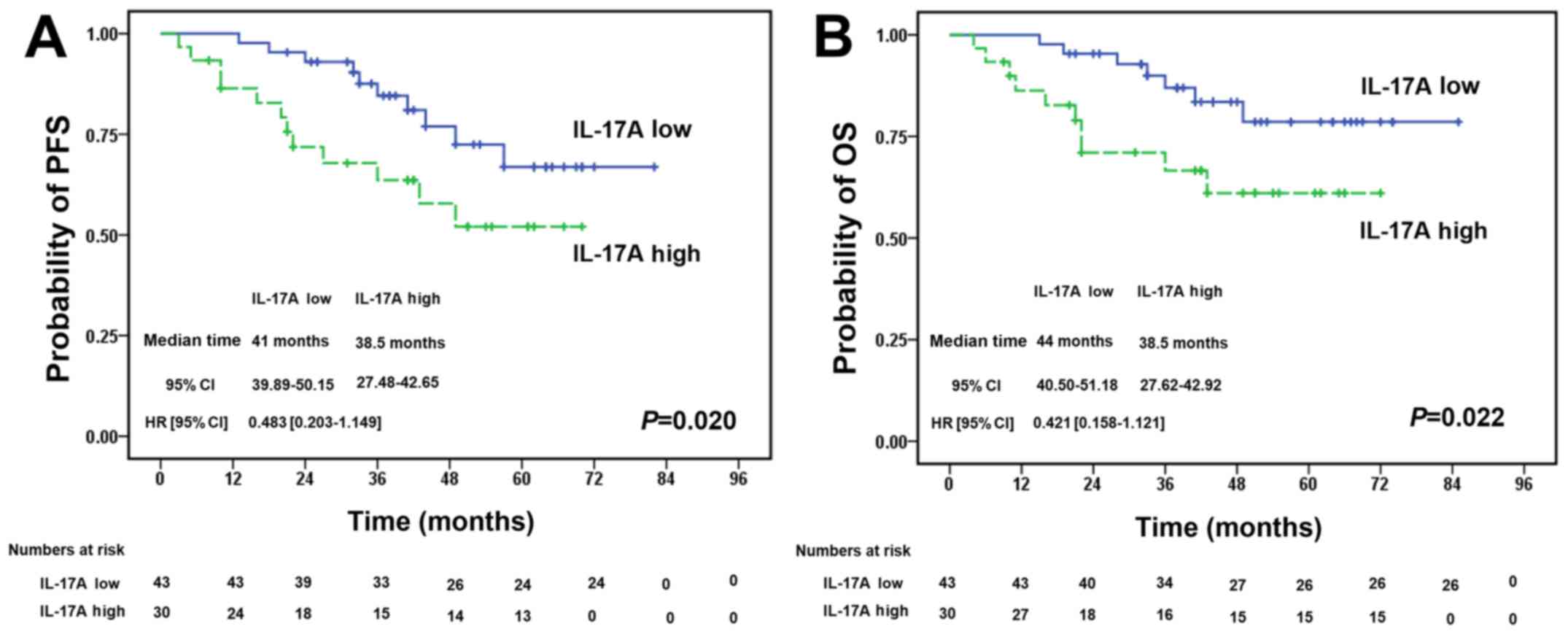
Increased expression of IL-17A in PB
predicts a poorer survival of patients with DLBCL
We investigated the association betweenIL-17A
expression in PB from patients with DLBCL and their prognosis. We
detected the IL-17A levels in PB from 73 patients with DLBCL prior
to treatment with R-CHOP regimens. The patients were separated into
2 groups according to the median PB level of IL-17A (54.18 pg/ml).
There was no significant difference in the baseline clinical
characteristics between the 2 groups (Table II). Survival analysis revealed
that patients with high IL-17A levels had a significantly poorer
progression-free survival (Fig.
6A) and overall survival (Fig.
6B) compared with the low IL-17A expression group (P=0.020 and
P=0.022, respectively).
| Table IIBaseline clinical characteristics of
patients with DLBCL with varying IL-17A expression levels. |
Table II
Baseline clinical characteristics of
patients with DLBCL with varying IL-17A expression levels.
| Variable | IL-17A in
peripheral blood
|
|---|
| Low (n=42) | High (n=31) |
|---|
| Age (years) | | |
| <50 | 15 | 10 |
| ≥50 | 27 | 21 |
| P-valuea | 0.807 | |
| Sex | | |
| Male | 26 | 17 |
| Female | 16 | 14 |
| P-valuea | 0.633 | |
| Ann Arbor
stageb | | |
| I–II | 17 | 13 |
| III–IV | 25 | 18 |
| P-valuea | 0.231 | |
| IPI scorec | | |
| 1–3 | 24 | 19 |
| 4–5 | 18 | 12 |
| P-valuea | 0.812 | |
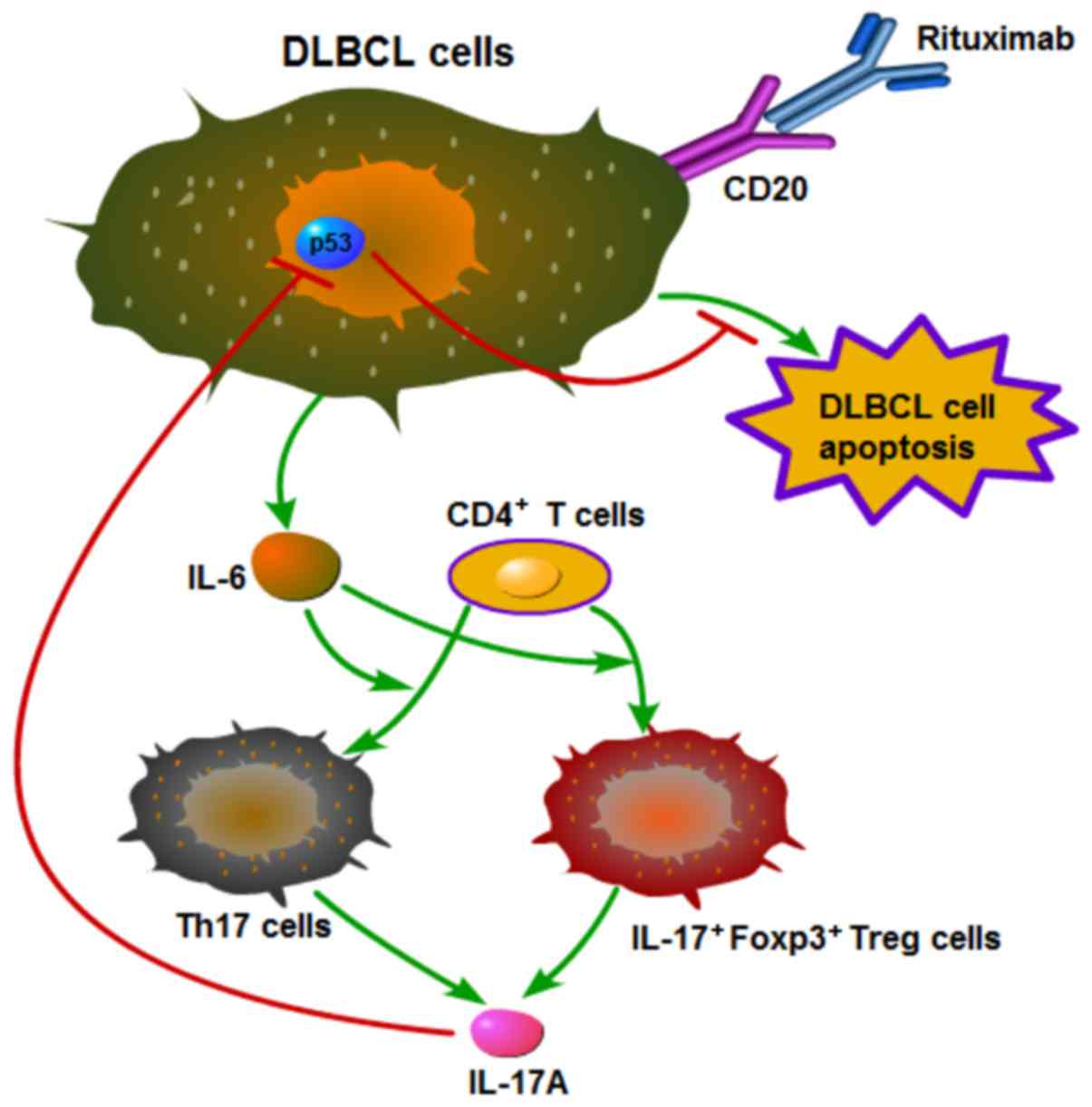
Mechanistic outline of RR in DLBCL
Based on the above-mentioned results, we summarized
the mechanistic outline of RR in DLBCL in Fig. 7. Rituximab induces DLBCL cells to
secrete IL-6, which promotes the differentiation of Th17 and
IL-17+Foxp3+ Treg cells from CD4+
T cells, and these two types of cells then secrete IL-17A. IL-17A
prevents rituximab-induced apoptosis and promotes the proliferation
of DLBCL cells by suppressing p53 expression.
Discussion
RR presents a challenge to clinical specialists by
limiting and weakening the therapeutic efficacy of rituximab
inpatients with DLBCL. However, the mechanisms responsible for of
RR remains elusive (5). In the
current study, we demonstrated that rituximab promoted the
differentiation of Th17 and IL-17+Foxp3+ Treg
cells, and increased the secretion of IL-17A by elevating IL-6
levels, both in patients with DLBCL and in vitro. IL-17A
suppressed p53 expression, leading to the inhibition of
rituximab-induced apoptosis and enhanced the proliferation of DLBCL
cells. Furthermore, high IL-17A levels in the PB of patients with
DLBCL predicted a relatively poor prognosis.
We previously demonstrated that B-NHL cells secreted
IL-6, which was upregulated by irradiation (21). Other studies have confirmed that
rituximab induced IL-6 production in human B cells in vitro
(26) and elevated serum IL-6
levels in patients with DLBCL following chemotherapy (23,24).
By comparing patients treated with and without rituximab (R-CHOP
and CHOP regimens), the current results indicated that rituximab
significantly promoted IL-6 secretion in patients with DLBCL, and
this finding was supported by in vitro experiments. Our
results revealed that IL-6 levels were also elevated in patients in
the R-CHOP-CR group. In addition, a previous study found that
rituximab induced IL-6 production in human B cells (26); thus, we considered that human B
cells and other cells apart from DLBCL cells secreted IL-6
following rituximab administration. Our results were thus in
accordance with the above conclusions. High plasma IL-6 levels have
been shown to be associated with poorer clinical outcomes following
rituximab-combined therapy in patients with DLBCL (27), suggesting that IL-6 may be a
potential therapeutic target in DLBCL. However, previous
experimental data demonstrated that anti-IL-6 therapy was
ineffective in irradiation-resistant lymphoma and myeloma cells
(28). We thus speculated that
increased IL-6 levels may play a role by influencing other cytokine
networks or signaling pathways. Further studies are warranted in
order to elucidate the mechanisms whereby increased IL-6 influences
the therapeutic effects of rituximab.
Previous studies have demonstrated that IL-17A plays
a critical role in promoting the growth of germinal cell BDLBCL
cells in vitro and in a mouse model (20), and in inhibiting
irradiation-induced apoptosis of NHL cells (21), which are mainly derived from Th17
and IL-17+Foxp3+ Treg cells. However, to the
best of our knowledge, no previous studies have focused on the
effects of rituximab on these two cell types and their associated
cytokines in DLBCL patients or in vitro. Yin et al
found that PBMC Th1 and Th2 cells from patients with DLBCL were
influenced by the presence or absence of rituximab in the
chemotherapy regimen, which was also related to the patients'
response to treatment (29). Two
previous studies have both shown that a low level of circulating
Treg cells predicts a poor prognosis in patients with DLBCL
(30,31); however, to the best of our
knowledge, no studies to date have determined whether rituximab
affects Th17 cell differentiation and IL-17 levels in patients with
DLBCL.T cells present developmental plasticity in the tumor
microenvironment (13). As we
speculated, our results provide the first evidence to indicate that
rituximab increases the proportions of Th17 and
IL-17+Foxp3+ Treg cells, and increases IL-17A
levels in vivo and in vitro, probably as a result of
increased IL-6 and TGF-β levels. These effects may be ascribed to
the plasticity of CD4+ T cell differentiation.
In this study, increased IL-6 levels induced the
differentiation of Th17 cells, which secrete IL-17A. Conversely,
other studies have shown that rituximab inhibits Th17 cells and
IL-17A expression in anti-neutrophil cytoplasmic
antibody-associated vasculitis and primary Sjogren's syndrome
(32,33). We therefore consider that rituximab
may affect Th17 and IL-17+Foxp3+ Treg cells
and IL-17A levels differently in NHL and in non-malignant B
cell-associated immunological diseases. Lymphoma B cells exhibit an
aberrant expression of CD70, CD80 or CD86 compared with normal B
cells, and these costimulatory molecules interact with their
ligands (CD27 or CD28) to play an important role in T cell
differentiation and development (34). Lymphoma B cells have also been
shown to upregulate Treg cells and inhibit Th17 cells by the
above-mentioned CD27-CD70 or CD28-CD80/86 interactions between
lymphoma cells and CD4+ T cells (18). We hypothesized that rituximab
deleted lymphoma B cells and abolished these costimulatory molecule
interactions, and increased the expression of Th17 cells. Moreover,
we demonstrated that patients with DLBCL had higher TGF-β levels
and that rituximab increased IL-6 levels in DLBCL cells, suggesting
that IL-6 and TGF-β may plastically promote Foxp3+ Treg
cells to secrete IL-17A and thereby upregulate
IL-17+Foxp3+ Treg cell development.
The anticancer or cancer-promoting effects of IL-17A
remain controversial, and may depend on the type of cancer or the
functions of immune cells and related cytokines in the tumor
microenvironment (13). A number
of studies have suggested that IL-17A can promote the growth of
many types of tumor (35-37). Ferretti et al demonstrated
that IL-17A boosted the growth of human germinal center-derived B
cell NHL in vitro and in a mouse model by promoting tumor
cell proliferation and neo-angiogenesis (20). Having demonstrated that rituximab
promoted IL-17A levels by inducing IL-6 expression both in
vivo and in vitro, we also examined the effects of
IL-17A on the development of DLBCL cells and showed that IL-17A
boosted their proliferation. Rituximab is known to induce DLBCL
cell apoptosis, and we therefore determined whether IL-17A affects
rituximab-induced DLBCL cell apoptosis and the expression of the
classical apoptosis regulator, p53. We found that rituximab-induced
IL-17A expression prevented rituximab-induced DLBCL cell apoptosis,
indicating that increased IL-17A thus promoted RR in patients with
DLBCL. These conclusions are in agreement with the findings of our
with our previous study (21) and
with other published research (20).
p53 protein is an important tumor suppressor
protein, and p53 dysfunction is involved in the pathogenesis of
B-cell malignancies, including DLBCL (38,39).
The dysfunction of p53 inhibits DLBCL apoptosis and promotes DLBCL
generation, progression and invasion (38). IL-17 has been reported to regulate
matrix metalloproteinase-9 mRNA levels in a p53-dependent manner
and to promote lung tumor growth (35), and the data from a previous study
indicated that high levels of IL-17 were associated with low levels
of p53 in colorectal cancer (40).
In accordance with the findings of previous studies, the results of
the present study demonstrated that IL-17A suppressed the
expression of p53 in DLBCL cells, which may partially account for
the mechanism where by IL-17A inhibits the apoptosis of DLBCL
cells.
We confirmed the association between IL-17A and the
prognosis of patients with DLBCL treated with rituximab by
analyzing the survival of 73 patients with DLBCL treated with the
R-CHOP regimen, and demonstrated that the increased expression of
IL-17A in the PB of the patients predicted a poorer survival.
IL-17A is therefore a prognostic factor in DLBCL. We previously
identified interferon regulatory factor 8 expression in the tumor
microenvironment as a prognostic factor for DLBCL (25), and other investigators have also
found that non-tumor immune cell components in DLBCL lesions
affected the efficacy of the R-CHOP regimen (17). However, serum IL-17A levels are
more easily measured and are thus more useful in a clinical
setting. In addition, this clinical result supports our hypothesis
that increased IL-17A levels promote RR in patients with DLBCL.
Patients with DLBCL usually receive rituximab combined chemotherapy
for at least 4-6 months. Furthermore, the half-life of rituximab is
long and it can persist for 3-6 months in vivo, suggesting
that IL-17A may promote RR continuously for a prolonged period.
Further clarification of the role of IL-17A in the pathogenesis of
DLBCL may allow for the development of anti-IL-17A therapies to
overcome RR.
Although the clinical part of this study was
conducted retrospectively, we carried out in vitro
experiments to confirm our conclusions. In the future, we aim to
conduct a randomized controlled trial with a greater number of
patients with DLBCL. We also aim to investigate other mechanisms
whereby IL-17A may promote RR, in addition to suppressing p53
expression in DLBCL cells. In addition, we aim to examine more
DLBCL cell lines, such as ABC-subtype cell lines, in our future
research.
In conclusion, in this study, we demonstrate that
rituximab elevates IL-6 levels, which promote Foxp3+
Treg cells to secrete IL-17A, consequently promoting the
differentiation of Th17 and IL-17+Foxp3+ Treg
cells, and increasing IL-17A expression in patients with DLBCL, as
well as in vitro. IL-17A prevents rituximab-induced
apoptosis and promotes the proliferation of DLBCL cells by
suppressing p53 expression (Fig.
7). Furthermore, increased PB levels of IL-17A predict an
unfavorable prognosis inpatients with DLBCL. These results suggest
that IL-17A promotes RR by suppressing p53 expression, and may thus
be a useful prognostic factor for DLBCL. IL-17A may also be a
potential future therapeutic target for RR.
Acknowledgments
Not applicable.
Notes
[1]
Funding
The present study was supported by the Guangzhou
Planned Project of Science and Technology, China (grant nos.
201707010279, 201300000100 and 201704020105).
[2] Availability
of data and materials
The analyzed datasets generated during the study
are available from the corresponding author on reasonable
request.
[3] Authors'
contributions
WZ contributed to the experimental design and
implementation, performed the experiments, drafted the manuscript,
and performed data analysis. QL and YW conceived or designed the
experiments, modified the manuscript. XX, ZZ, LY and ZY contributed
to experiment implementation and data analysis. QD, HD, HX and ZX
performed the experiments and data analysis. All authors have read
and approved the final manuscript.
[4] Ethics
approval and consent to participate
This study was approved by the Ethics Committee of
Guangzhou First People's Hospital (2014-SYL-034). Written informed
consent was obtained from all participants or their families prior
to obtaining the samples.
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Sehn LH and Gascoyne RD: Diffuse large
B-cell lymphoma: Optimizing outcome in the context of clinical and
biologic heterogeneity. Blood. 125:22–32. 2015. View Article : Google Scholar
|
|
2
|
Lenz G, Wright G, Dave SS, Xiao W, Powell
J, Zhao H, Xu W, Tan B, Goldschmidt N, Iqbal J, et al
Lymphoma/Leukemia Molecular Profiling Project: Stromal gene
signatures in large-B-cell lymphomas. N Engl J Med. 359:2313–2323.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nakajima Y, Tomita N, Itabashi M,
Miyashita K, Watanabe R, Miyazaki T, Tachibana T, Takasaki H,
Kawasaki R, Tanaka M, et al: Analysis of outcomes in patients with
supra-diaphragmatic vs infra-diaphragmatic diffuse large B cell
lymphoma treated with R-CHOP therapy. Leuk Res. 39:198–203. 2015.
View Article : Google Scholar
|
|
4
|
Fowler NH: R2-CHOP vs R-CHOP for diffuse
large B-cell lymphoma. Clin Adv Hematol Oncol. 12:608–610.
2014.
|
|
5
|
Rezvani AR and Maloney DG: Rituximab
resistance. Best Pract Res Clin Haematol. 24:203–216. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stolz C and Schuler M: Molecular
mechanisms of resistance to Rituximab and pharmacologic strategies
for its circumvention. Leuk Lymphoma. 50:873–885. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bello C and Sotomayor EM: Monoclonal
antibodies for B-cell lymphomas: Rituximab and beyond. Hematology
(Am Soc Hematol Educ Program). 2007.233–242. 2007.
|
|
8
|
Seyfizadeh N, Seyfizadeh N, Hasenkamp J
and Huerta-Yepez S: A molecular perspective on rituximab: A
monoclonal antibody for B cell non Hodgkin lymphoma and other
affections. Crit Rev Oncol Hematol. 97:275–290. 2016. View Article : Google Scholar
|
|
9
|
Pavanello F, Zucca E and Ghielmini M:
Rituximab: 13 open questions after 20 years of clinical use. Cancer
Treat Rev. 53:38–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Iwakura Y, Ishigame H, Saijo S and Nakae
S: Functional specialization of interleukin-17 family members.
Immunity. 34:149–162. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gaffen SL: Structure and signalling in the
IL-17 receptor family. Nat Rev Immunol. 9:556–567. 2009. View Article : Google Scholar
|
|
12
|
Bettelli E, Carrier Y, Gao W, Korn T,
Strom TB, Oukka M, Weiner HL and Kuchroo VK: Reciprocal
developmental pathways for the generation of pathogenic effector
TH17 and regulatory T cells. Nature. 441:235–238. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hemdan NY: Anti-cancer versus
cancer-promoting effects of the interleukin-17-producing T helper
cells. Immunol Lett. 149:123–133. 2013. View Article : Google Scholar
|
|
14
|
Du R, Zhao H, Yan F and Li H:
IL-17+ Foxp3+ T cells: An intermediate
differentiation stage between Th17 cells and regulatory T cells. J
Leukoc Biol. 96:39–48. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang C and Fu ZX: Localization of
IL-17+Foxp3+ T cells in esophageal cancer.
Immunol Invest. 40:400–412. 2011. View Article : Google Scholar
|
|
16
|
Li L and Boussiotis VA: The role of
IL-17-producing Foxp3+ CD4+ T cells in
inflammatory bowel disease and colon cancer. Clin Immunol.
148:246–253. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gomez-Gelvez JC, Salama ME, Perkins SL,
Leavitt M and Inamdar KV: Prognostic impact of tumor
microenvironment in diffuse large B-cell lymphoma uniformly treated
with R-CHOP chemotherapy. Am J Clin Pathol. 145:514–523. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang ZZ, Novak AJ, Ziesmer SC, Witzig TE
and Ansell SM: Malignant B cells skew the balance of regulatory T
cells and TH17 cells in B-cell non-Hodgkin's lymphoma. Cancer Res.
69:5522–5530. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lu T, Yu S, Liu Y, Yin C, Ye J, Liu Z, Ma
D and Ji C: Aberrant circulating Th17 cells in patients with B-cell
non-Hodgkin's lymphoma. PLoS One. 11:e01480442016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ferretti E, Di Carlo E, Ognio E, Guarnotta
C, Bertoni F, Corcione A, Prigione I, Fraternali-Orcioni G, Ribatti
D, Ravetti JL, et al: Interleukin-17A promotes the growth of human
germinal center derived non-Hodgkin B cell lymphoma.
OncoImmunology. 4:e10305602015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Q, Xu X, Zhong W, Du Q, Yu B and Xiong
H: IL-17 induces radiation resistance of B lymphoma cells by
suppressing p53 expression and thereby inhibiting
irradiation-triggered apoptosis. Cell Mol Immunol. 12:366–372.
2015. View Article : Google Scholar :
|
|
22
|
Obenauf AC, Zou Y, Ji AL, Vanharanta S,
Shu W, Shi H, Kong X, Bosenberg MC, Wiesner T, Rosen N, et al:
Therapy-induced tumour secretomes promote resistance and tumour
progression. Nature. 520:368–372. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Khan MA, Garg K, Bhurani D and Agarwal NB:
Early manifestation of mild cognitive impairment in B-cell
non-Hodgkin's lymphoma patients receiving CHOP and rituximab-CHOP
chemotherapy. Naunyn Schmiedebergs Arch Pharmacol. 389:1253–1265.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zimmer P, Mierau A, Bloch W, Strüder HK,
Hülsdünker T, Schenk A, Fiebig L, Baumann FT, Hahn M, Reinart N, et
al: Post-chemotherapy cognitive impairment in patients with B-cell
non-Hodgkin lymphoma: A first comprehensive approach to determine
cognitive impairments after treatment with rituximab,
cyclophosphamide, doxorubicin, vincristine and prednisone or
rituximab and bendamustine. Leuk Lymphoma. 56:347–352. 2015.
View Article : Google Scholar
|
|
25
|
Zhong W, Xu X, Zhu Z, Du Q, Du H, Yang L,
Ling Y, Xiong H and Li Q: Increased expression of IRF8 in tumor
cells inhibits the generation of Th17 cells and predicts
unfavorable survival of diffuse large B cell lymphoma patients.
Oncotarget. 8:49757–49772. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jones JD, Hamilton BJ, Skopelja S and
Rigby WF: Induction of interleukin-6 production by rituximab in
human B cells. Arthritis Rheumatol. 66:2938–2946. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Giachelia M, Voso MT, Tisi MC, Martini M,
Bozzoli V, Massini G, D'Aló F, Larocca LM, Leone G and Hohaus S:
Interleukin-6 plasma levels are modulated by a polymorphism in the
NF-κB1 gene and are associated with outcome following
rituximab-combined chemotherapy in diffuse large B-cell non-Hodgkin
lymphoma. Leuk Lymphoma. 53:411–416. 2012. View Article : Google Scholar
|
|
28
|
Gougelet A, Mansuy A, Blay JY, Alberti L
and Vermot-Desroches C: Lymphoma and myeloma cell resistance to
cytotoxic agents and ionizing radiations is not affected by
exposure to anti-IL-6 antibody. PLoS One. 4:e80262009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yin Q, Chen L, Li Q, Mi R, Li Y, Wei X and
Song Y: Changes of T-lymphocyte subpopulation and differential
expression pattern of the T-bet and GATA-3 genes in diffuse large
B-cell lymphoma patients after chemotherapy. Cancer Cell Int.
14:852014. View Article : Google Scholar
|
|
30
|
Rusak M, Bołkun Ł, Chociej-Stypułkowska J,
Pawlus J, Kłoczko J and Dąbrowska M: Flow-cytometry-based
evaluation of peripheral blood lymphocytes in prognostication of
newly diagnosed DLBCL patients. Blood Cells Mol Dis. 59:92–96.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
G łowala-Kosińska M, Chwieduk A, Nieckula
J, Saduś-Wojciechowska M, Grosicki S, Rusin A, Nowara E and Giebel
S: Association of circulating regulatory T cell number with the
incidence and prognosis of diffuse large B-cell lymphoma. Eur J
Haematol. 91:122–128. 2013. View Article : Google Scholar
|
|
32
|
Pullerits R, Ljevak M, Vikgren J and
Bokarewa M: Off-trial evaluation of the B cell-targeting treatment
in the refractory cases of antineutrophil cytoplasmic antibodies
(ANCA)-associated vasculitis: Long-term follow-up from a single
centre. Scand J Immunol. 76:411–420. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ciccia F, Guggino G, Rizzo A, Alessandro
R, Carubbi F, Giardina A, Cipriani P, Ferrante A, Cannizzaro A,
Giacomelli R, et al: Rituximab modulates IL-17 expression in the
salivary glands of patients with primary Sjögren's syndrome.
Rheumatology (Oxford). 53:1313–1320. 2014. View Article : Google Scholar
|
|
34
|
Yang ZZ, Novak AJ, Ziesmer SC, Witzig TE
and Ansell SM: CD70+ non-Hodgkin lymphoma B cells induce
Foxp3 expression and regulatory function in intratumoral
CD4+CD25 T cells. Blood. 110:2537–2544. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu B, Guenther JF, Pociask DA, Wang Y,
Kolls JK, You Z, Chandrasekar B, Shan B, Sullivan DE and Morris GF:
Promotion of lung tumor growth by interleukin-17. Am J Physiol Lung
Cell Mol Physiol. 307:L497–L508. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang Q, Liu S, Zhang Q, Xiong Z, Wang AR,
Myers L, Melamed J, Tang WW and You Z: Interleukin-17 promotes
development of castration-resistant prostate cancer potentially
through creating an immunotolerant and pro-angiogenic tumor
microenvironment. Prostate. 74:869–879. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu X, Zeng Z, Xu L, Yu J, Cao Q, Chen M,
Sung JJ and Hu P: Increased expression of IL17A in human gastric
cancer and its potential roles in gastric carcinogenesis. Tumour
Biol. 35:5347–5356. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tessoulin B, Eveillard M, Lok A, Chiron D,
Moreau P, Amiot M, Moreau-Aubry A, Le Gouill S and
Pellat-Deceunynck C: p53 dysregulation in B-cell malignancies: More
than a single gene in the pathway to hell. Blood Rev. 31:251–259.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lu TX, Young KH, Xu W and Li JY: TP53
dysfunction in diffuse large B-cell lymphoma. Crit Rev Oncol
Hematol. 97:47–55. 2016. View Article : Google Scholar
|
|
40
|
Radosavljevic G, Ljujic B, Jovanovic I,
Srzentic Z, Pavlovic S, Zdravkovic N, Milovanovic M, Bankovic D,
Knezevic M, Acimovic LJ, et al: Interleukin-17 may be a valuable
serum tumor marker in patients with colorectal carcinoma.
Neoplasma. 57:135–144. 2010. View Article : Google Scholar : PubMed/NCBI
|