Introduction
MicroRNAs (miRNAs or miRs) are a major type of small
noncoding RNA molecules involved in gene regulation via binding to
the 3′ untranslated region of their target mRNAs, and subsequently
causing mRNA degradation or translation inhibition (1–4). An
increasing volume of research has provided evidence that miRNAs act
as oncogenes or tumor suppressors in various malignant tumors by
influencing tumor growth, proliferation, apoptosis, invasion and
metastasis (5–8). The close association between miRNAs
and breast cancer (BC) has also been investigated. For instance,
Ding et al (9) revealed
that miR-145 is a tumor suppressor in BC and performs its
anti-oncogenic role by inhibiting a cancer-associated gene,
transforming growth factor-β1. Xiao et al (10) observed that miR-129 blocks BC
regeneration by degrading estrogen receptor 1 mRNA at the
posttranscriptional level, inhibiting the subsequent
estrogen-induced NOTCH cascade and ultimately reducing the number
of stem-like cells. A recent study conducted by Lu et al
(11) demonstrated that miR-129-5p
increases the sensitivity of human epidermal growth factor receptor
2 (Her2)-positive BC to trastuzumab by downregulating ribosomal
protein S6. In addition, due to the ease of detecting miRNAs in
tumor biopsies and body fluids, numerous researchers have
highlighted the diagnostic and prognostic roles of miRNAs in
cancer, including in BC. Jang et al (12) examined miR-9 and miR-155 expression
levels in triple-negative BC and found that they functioned as
biomarkers for prognosis prediction. Zhang et al (13) detected circulating miRNAs
expression using a serum-direct multiplex detection assay based on
reverse transcription-polymerase chain reaction (RT-PCR). The
authors determined a 3-miRNA signature (miR-424, miR-199a and
miR-29c) that exhibited the highest diagnostic accuracy for BC
diagnosis (13). Our research
group has also identified a 9-miRNA signature for BC diagnosis
based on data from The Cancer Genome Atlas (TCGA) (14). Therefore, the investigation of
miRNAs in cancer has provided promising results, and exploring
miRNAs involved in the occurrence and development of BC is
conducive to the diagnosis and treatment of patients with BC.
miR-671 precursors form two mature miRNAs, namely
miR-671-5p and miR-671-3p. A recent study conducted by Tan et
al (15) has demonstrated that
miR-671-5p is an antioncogene in BC. The authors also observed that
miR-671-5p was downregulated in BC and that forced miR-671-5p
expression in the BC cell line MDA-MB-231 was able to inhibit cell
proliferation and invasion, and sensitize cells to chemotherapy
(15). Furthermore, Godfrey et
al (16) compared global miRNA
expression in 205 BC patients and 205 healthy volunteers using a
microarray method, and observed a high expression of miR-671-3p in
the serum samples of BC patients. However, a study concentrating on
the biological function and molecular mechanism of miR-671-3p in BC
has yet to be reported.
The present study focused on the expression level,
biological function and potential molecular mechanism of miR-671-3p
in BC. In vitro experiments were conducted to determine the
biological effects of miR-671-3p on BC cells, and miR-671-3p
expression was detected using RT-quantitative PCR (RT-qPCR).
Simultaneously, all available microarray datasets were combined
from the Gene Expression Omnibus (GEO) and ArrayExpress databases
to verify the expression of miR-671-3p in BC. Furthermore, the
predicted targets of miR-671-3p were examined, Gene Ontology (GO)
annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG)
pathway analyses were conducted, and a protein-protein interaction
(PPI) network was constructed to gain insight into the underlying
molecular mechanism of miR-671-3p in BC.
Materials and methods
Patients and samples
A total of 38 pairs of BC and adjacent non-tumorous
tissues were obtained from patients who were pathologically
diagnosed with BC (17). To ensure
that the RT-qPCR results were not affected by the size of
paraffin-embedded tissues, samples with a surface area that was too
small or large were excluded. Finally, 38 BC tissues and 11
adjacent non-tumorous tissues with a surface area of ~1.5
cm2 were selected. All patients underwent initial
surgery at the First Affiliated Hospital of Guangxi Medical
University (Nanning, China) between January 2012 and December 2013,
and had not received any preoperative radiotherapy or chemotherapy.
Adjacent non-tumorous tissues were collected at >5 cm away from
the tumorous node. Following excision, tissues were cryopreserved
in liquid nitrogen and stored at −80°C prior to RNA extraction. All
participants provided informed consent prior to sample collection.
The Ethics Committee of the First Affiliated Hospital of Guangxi
Medical University approved this investigation. The workflow of the
present study is exhibited in Fig.
1.
Cell culture and transfection with
miR-671-3p mimic
The human BC-derived cell line MDA-MB-231 was
provided by the American Type Culture Collection (Manassas, VA,
USA) and cultured as described in previous studies (18,19).
For transfection, MDA-MB-231 cells were seeded in a 96-well plate
(2.5×103 cells per well), and subsequently incubated at
37°C for 24 h. Next, miR-671-3p mimic and negative mimic control
(Ambion; Thermo Fisher Scientific, Inc., Waltham, MA, USA) were
transfected into the cells at a concentration of 200 nmol/l with a
CombiMag Magnetofection transfection kit (OZ Biosciences,
Marseille, France) according to the manufacturer's protocol. A
blank control was set with nothing transfected into cells.
Following transfection, cell samples were collected at 0, 24, 48,
72 and 96 h for further analyses. All in vitro experiments
were conducted in triplicate.
RT-qPCR
Total RNA was isolated using the miRNeasy FFPE kit
(Qiagen, Duesseldorf, German) following the manufacturer's
protocol. The measurement of total RNA quality was conducted on
NanoDrop-2000 Ultra Micro Spectrophotometer (Thermo Fisher
Scientific, Inc.), and an RNA concentration >45 ng/µl was
required based on the calculation by the instructions of miScript
II RT kit and miScript SYBR-Green PCR kit. Then, total RNA was
reversed into cDNA with miScript II RT kit (Qiagen). Subsequently,
RT-qPCR was conducted with 7900HT Fast Real-Time PCR System
(Applied Biosystems; Thermo Fisher Scientific, Inc.) in accordance
with the manufacturer's protocol. The thermal cycling conditions
are shown in Table I. RNU44 was
used as a reference gene for the normalization of miR-671-3p
abundance. The primers for miR-671-3p (cat. no. 1000066;
CCGGUUCUCAGGGCUCCACC) and RNU44 (cat. no. MS00033855) were
synthesized by Qiagen. All the samples were examined in triplicate,
and the mean of the three well values were used as the Cq value.
The relative expression level of miR-671-3p was determined
according to the 2−ΔCq method (ΔCq =
CqmiR-671-3p-CqRNU44), as previously
described (20,21).
 | Table IThermal cycling conditions of
RT-qPCR. |
Table I
Thermal cycling conditions of
RT-qPCR.
| Step | Time | Temperature
(°C) | Cycle |
|---|
|
Pre-denaturation | 15 min | 95 | 1 |
| Three steps of one
cycle | | | 45 |
| Denaturation | 15 sec | 94 | |
| Annealing | 30 sec | 55 | |
| Elongation | 30 sec | 70 | |
Cell function detection
Cell viability and proliferation were examined by
fluorometric detection with resorufin (CellTiter-Blue Cell
Viability Assay; cat. no. G8080; Promega Corportation, Madison, WI,
USA) and a colorimetric tetrazolium (MTS) assay (CellTiter96
AQUEOUS One Solution Cell Proliferation Assay; cat. no. G3580;
Promega Corporation), respectively. Furthermore, cell apoptosis was
assessed by Hoechst 33342 and propidium iodide (PI; both purchased
from Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
double-fluorescent chromatin staining. Caspase-3/7 activity was
also examined with a synthetic rhodamine-labeled caspase-3/7
substrate (Apo-ONE Homogeneous Caspase-3/7 Assay; cat. no. G7790;
Promega Corporation). The protocols of these assays were performed
as reported in previous studies (18,19,21–23).
Studies on GEO and ArrayExpress
databases
To verify the expression level of miR-671-3p in BC,
a meta-analysis was conducted based on data obtained from the GEO
(www.ncbi.nlm.nih.gov/geo) and
ArrayExpress (www.ebi.ac.uk/array-express) databases. Relevant
records were retrieved with the following keywords: (cancer OR
carcinoma OR tumor OR neoplas* OR malignan*) AND (breast OR mammary
OR mastocarcinoma) AND (miRNA OR microRNA OR miR OR 'non-coding
RNA' OR ncRNA). The last update time was October 25th, 2017.
Datasets were included once they met the following inclusion
criteria: i) The records provided expression data of miR-671-3p in
BC; ii) the number of samples in each dataset in the cancerous and
non-tumorous groups was >3; and iii) the subjects involved in
the study were humans. The following datasets were excluded: i)
Studies not associated with BC; ii) records with insufficient data
to calculate the expression level of miR-671-3p; and iii) animal
studies.
Two trained investigators independently screened all
available datasets and extracted the following characteristics from
all the enrolled studies: First author, publication year, country,
data source, platform, sample size, and expression values of
miR-671-3p in cancer and normal groups. Any disagreement was
resolved through discussion with a third researcher.
The standard mean difference (SMD) with the 95%
confi-dence interval (CI) was calculated to appraise miR-671-3p
expression in BC. SMD <0 indicated that miR-671-3p was
downregulated, and the result was consider as statistically
significant if the corresponding 95% CI of SMD did not overlap
zero. Heterogeneity among studies was evaluated by the
χ2-based Q test and I2 statistic. If a
statistically significant heterogeneity existed (I2
>50% or P<0.05), a random-effects model was applied.
Otherwise, a fixed-effects model was selected (24). Furthermore, sensitivity analysis
was conducted to evaluate whether the pooled result was stable.
Begg's funnel plot and Egger's test were also used to determine the
possible publication bias, with P>0.05 suggesting no publication
bias. All the aforementioned calculations were performed with Stata
software, version 12.0 (Stata Corporation, College Station, TX,
USA).
Target prediction and bioinformatics
analyses
The predicted target genes of miR-671-3p were
determined using the miRWalk 2.0 database (zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2), which
contains 12 online tools, namely Targetscan, RNAhybrid, RNA22,
PITA, Pictar2, miRWalk, Microt4, miRNAMap, miRDB, mirbridge,
miRanda and miRMap. Targets predicted by >3 algorithms were
selected for GO functional annotation and KEGG pathway analyses
using the DAVID online tool (david.ncifcrf.gov). Subsequently, the enriched results
were visualized with the R package 'ggplot2' (cran.r-project.org/web/packages/ggplot2/index.html).
To better understand the molecular mechanism of miR-671-3p in BC,
target genes enriched in cancer-associated pathways were uploaded
to the STRING database (string-db.org)
for PPI network construction. Meanwhile, the regulatory network of
the miRNA-gene was constructed and visualized with Cytoscape
version 3.4.0 software (25).
Furthermore, the expression of these target mRNAs was validated
using data from TCGA (cancergenome.nih.gov). A heatmap was drawn with the R
package 'pheatmap' (cran.r-project.org/web/packages/pheatmap/index.html),
and a box-scatter plot was generated with GraphPad Prism 5 software
(GraphPad Software, Inc., La Jolla, CA, USA).
Statistical analysis
Values are represented as the mean ± standard
deviation. Mann-Whitney U test and Student's t-test were applied
for comparison between continuous variables. Fisher's exact test
was also conducted to evaluate the correlation between miR-671-3p
expression and clinicopathological parameters of age, histological
grade (26), T stage, N stage, M
stage, TNM stage (27), molecular
subtype, ER/PR status and Her2 status. A receiver operating
characteristic (ROC) curve was generated to determine the ability
of miR-671-3p to distinguish BC from non-tumorous breast tissues. A
Kaplan-Meier survival curve with log-rank test was utilized to
estimate the prognostic power of miR-671-3p in BC. All these
analyses were conducted using SPSS version 22.0 software (IBM
Corp., Armonk, NY, USA), and P<0.05 was considered to indicate a
difference that was statistical significance.
Results
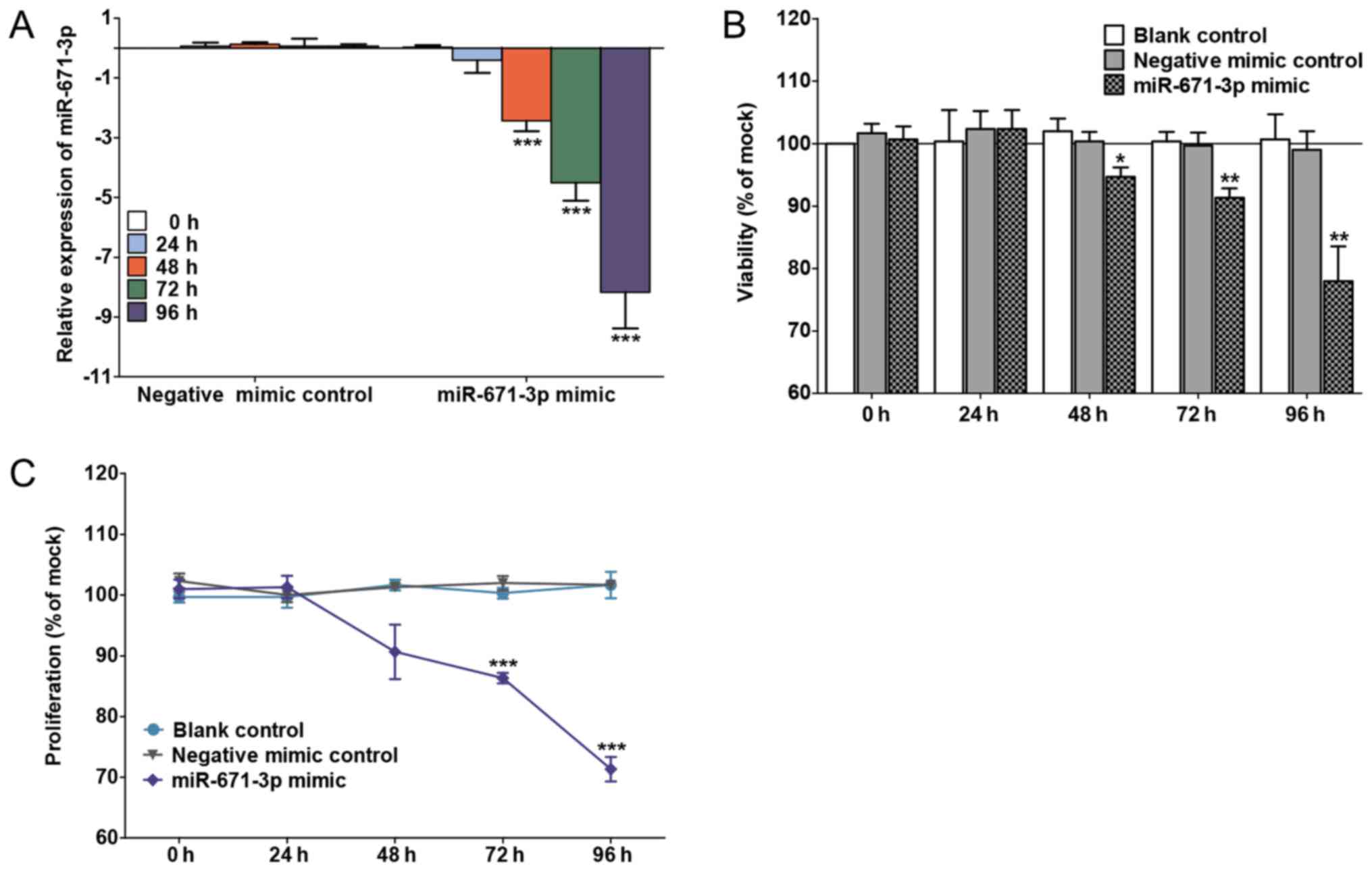
miR-671-3p mimic arrests the
proliferation and induces the apoptosis of BC cells in vitro
The transfection efficiency is shown in Fig. 2A. miR-671-3p was significantly
upregulated in the miR-671-3p mimic group at 48 h (P<0.0001), 72
h (P<0.0001) and 96 h (P<0.0001) compared with the negative
mimic control group. An unregulated miR-671-3p decreased cell
viability and proliferation, as determined by the fluoro-metric
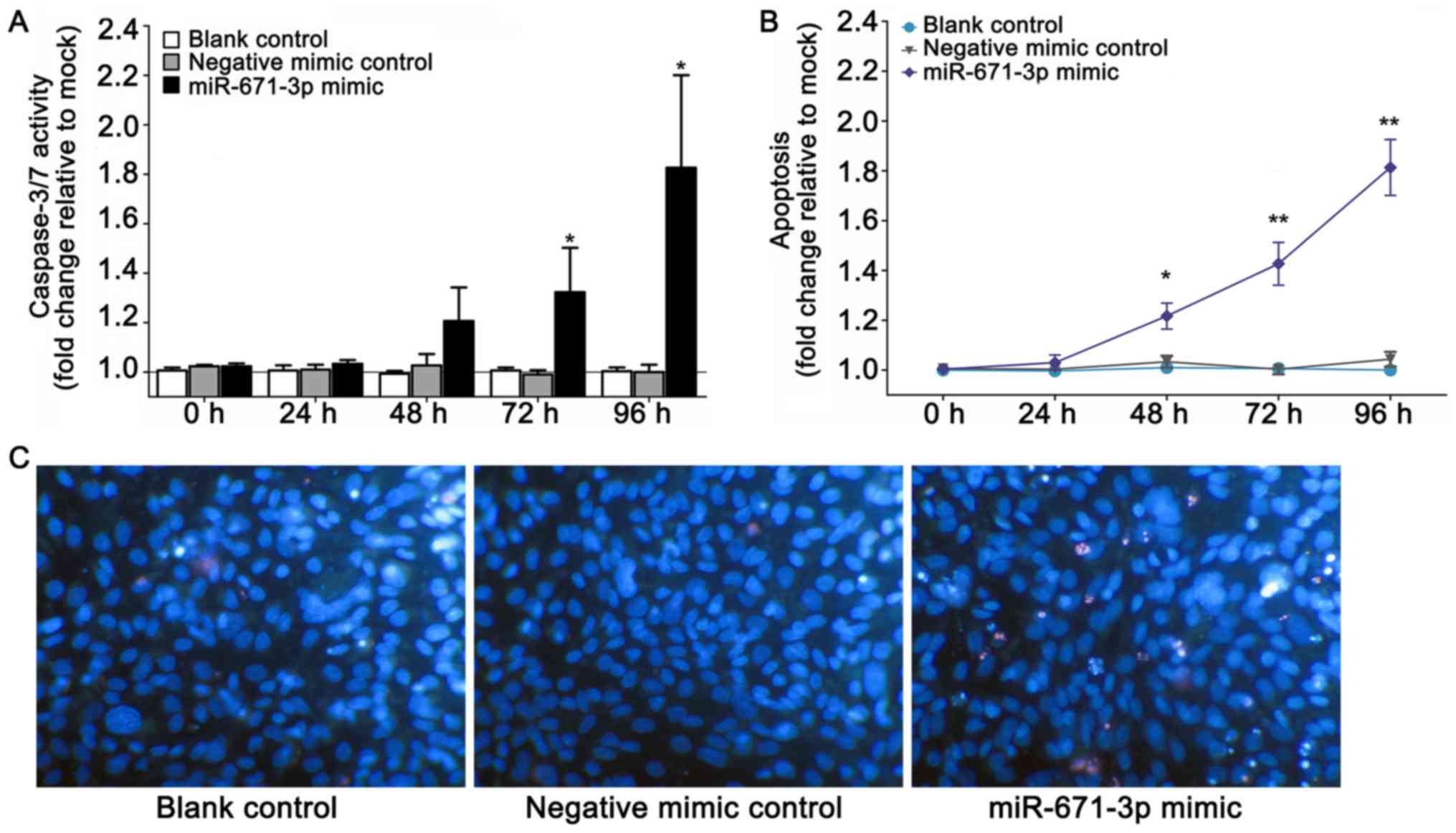
resorufin and MTS assays, respectively (Fig. 2B and C). Furthermore, to explore
the effects of the miR-671-3p mimic on cell apoptosis,
CellTiter-Blue and fluorescent caspase-3/7 assays were conducted.
The results indicated that the caspase-3/7 activity and apoptosis
of cells transfected with miR-671-3p mimic increased compared the
caspase-3/7 activity and apoptosis of cells in the blank and
negative mimic control groups (Fig. 3A
and B). In addition, viable and apoptotic cells were observed
under a microscope subsequent to Hoechst 33342/PI
double-fluorescent chromatin staining at 96 h. The results revealed
that miR-671-3p overexpression evidently inhibited the viability
and boosted the apoptosis of MDA-MB-231 cells in vitro
(Fig. 3C).
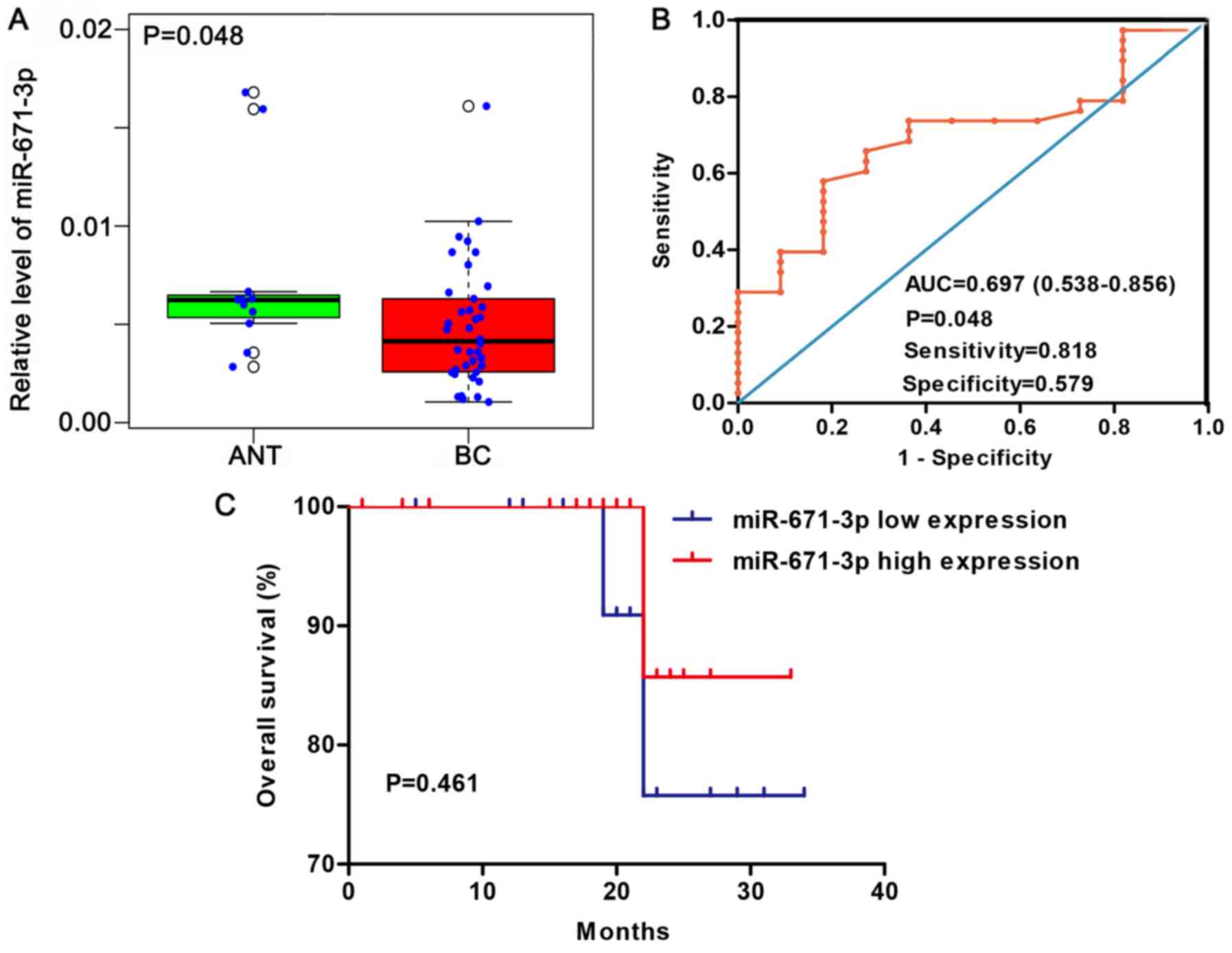
Expression level and clinical value of
miR-671-3p in BC via in-house RT-qPCR
According to the median expression of miR-671-3p in
38 BC tissues and 11 adjacent non-tumorous tissues, samples were
divided into high- and low-miR-671-3p group. A total of 17 tumor
samples (44.7%) exhibited high miR-671-3p expression, while 9
non-tumorous samples (81.8%) presented high miR-671-3p expression
(81.8%). The expression pattern of miR-671-3p was visualized as a
box-scatter plot and ROC curve. According to the results, the
miR-671-3p expression was significantly lower in cancer tissues
when compared with that in adjacent non-tumorous tissues (P=0.048;
Fig. 4A). The area under the ROC
curve was 0.697 (95% CI=0.538-0.856; P=0.048; Fig. 4B), with a sensitivity and
specificity of 0.818 and 0.579, respectively.
The association of miR-671-3p expression with the
clinicopathological characteristics of patients, including the age,
histological grade, T stage, N stage, M stage, TNM stage, molecular
subtype, ER/PR status and Her2 status, was investigated. As shown
in Table II, no statistically
significant differences were detected. Furthermore, the association
between miR-671-3p expression and prognosis was examined. The
patients were divided into high and low miR-671-3p expression
groups according to the median value of miR-671-3p expression.
Patients with highly expressed miR-671-3p had a mean survival time
of 31.429 months, while patients with low miR-671-3p expression had
a mean survival time of 30.818 months. However, no statistically
significant difference was observed between the two groups
(P=0.461; Fig. 4C).
| Table IIAssociation between miR-671-3p
expression and clinicopathological parameters of patients
(n=38). |
Table II
Association between miR-671-3p
expression and clinicopathological parameters of patients
(n=38).
| Characteristic | Group | Count | miR-671-3p
expression
| P-value |
|---|
| Low (%) | High (%) |
|---|
| Age (years) | ≤50 | 18 | 11 (61.1) | 7 (38.9) | 0.532 |
| >50 | 20 | 10 (50) | 10 (50) | |
| Tumor size | ≤2.5 cm | 11 | 7 (63.6) | 4 (36.4) | 0.721 |
| >2.5 cm | 27 | 14 (51.9) | 13 (48.1) | |
| Pathological
grade | IDC I | 5 | 3 (60) | 2 (40) | 0.396 |
| IDC II | 24 | 15 (62.5) | 9 (37.5) | |
| IDC III | 9 | 3 (33.3) | 6 (66.7) | |
| T stage | T1-T2 | 30 | 17 (56.7) | 13 (43.3) | 1 |
| T3-T4 | 8 | 4 (50) | 4 (50) | |
| N stage | N0 | 18 | 10 (55.6) | 8 (44.4) | 1 |
| N1-N3 | 20 | 11 (55) | 9 (45) | |
| M stage | M0 | 38 | 21 (55.3) | 17 (44.7) | – |
| M1 | 0 | – | – | |
| TNM stage | I | 6 | 4 (66.7) | 2 (33.3) | 0.91 |
| II | 19 | 10 (52.6) | 9 (47.4) | |
| III | 13 | 7 (46.2) | 6 (53.8) | |
| Molecular
subtype | Luminal | 10 | 6 (60) | 4 (40) | 0.315 |
| Her2-positive | 18 | 7 (38.9) | 11 (61.1) | |
| Triple
negative | 10 | 7 (70) | 3 (30) | |
| ER/PR | Negative | 28 | 15 (53.6) | 13 (46.4) | 1 |
| Positive | 10 | 6 (60) | 4 (40) | |
| Her2 | Negative | 20 | 14 (70) | 6 (30) | 0.101 |
| Positive | 18 | 7 (38.9) | 11 (61.1) | |
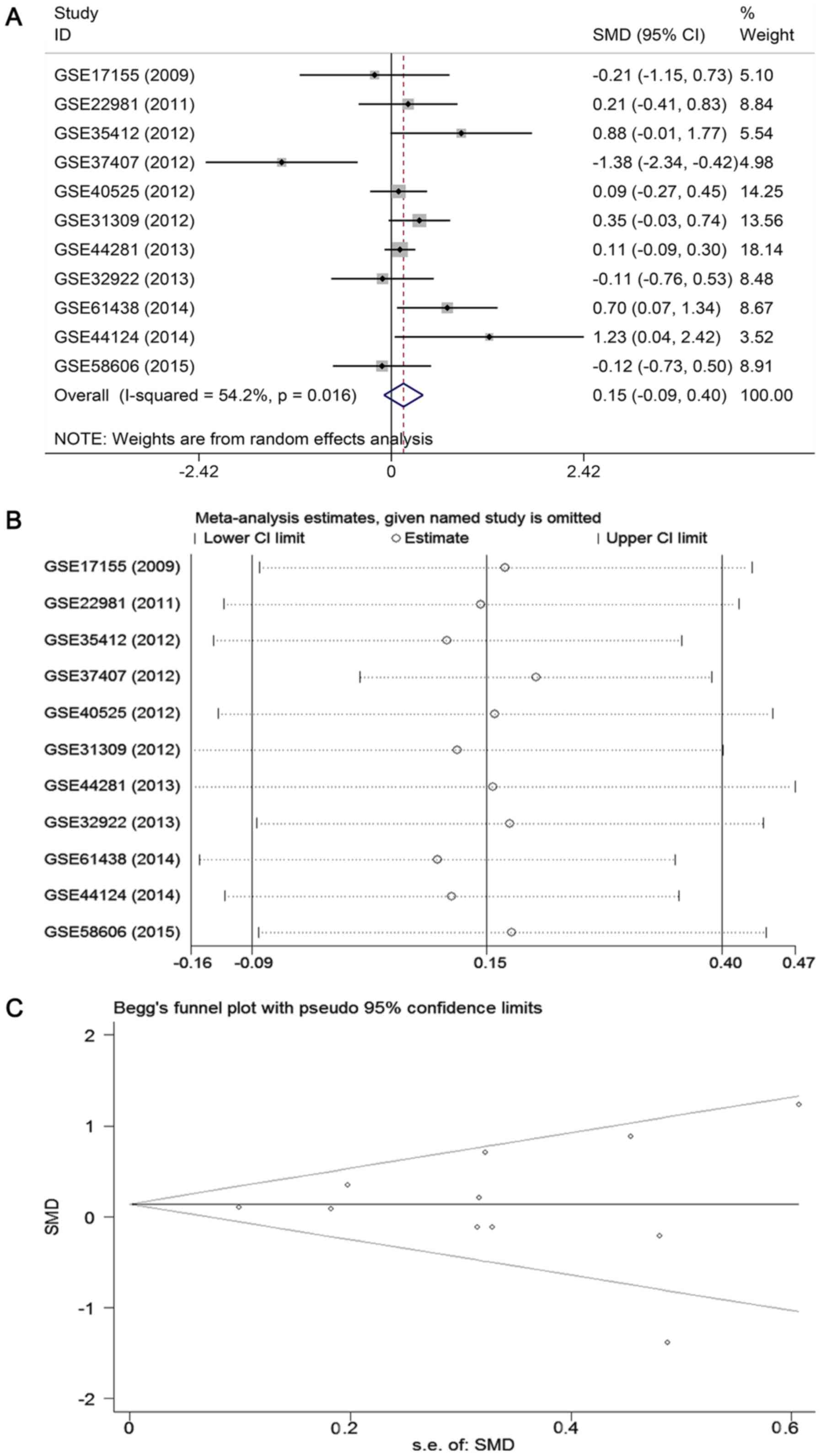
Results of meta-analyses based on GEO and
ArrayExpress databases
To further verify the expression level of miR-671-3p
in BC, a meta-analysis based on microarray datasets was conducted.
A total of 11 studies with 688 BC patients and 400 healthy subjects
were included (16,28-36).
The basic characteristics of the 11 available datasets are
displayed in Table III. The
combined SMD from a random-effects model demonstrated that the
expression of miR-671-3p in BC samples was similar to that in
normal controls (SMD: 0.15, 95% CI: -0.09 to 0.40, P=0.222;
heterogeneity: I2=54.2%, P=0.016; Fig. 5A). Sensitivity analysis revealed
that the pooled result was stable (Fig. 5B). Begg's and Egger's tests
indicated that no publication bias was observed among the 11
records (Begg's test, P=0.755; Egger's test, P=0.564; Fig. 5C).
| Table IIIMain characteristics of the 11
included microarray datasets obtained from the GEO and ArrayExpress
databases. |
Table III
Main characteristics of the 11
included microarray datasets obtained from the GEO and ArrayExpress
databases.
| First author
(year) | Country | Data source | Platform | Sample size
(T/N) | miR-671-3p
expressiona
| Ref. |
|---|
| T | N |
|---|
| Fassan et al
(2009) | Italy | GEO: GSE17155 | GPL8871 | 33/5 | 7.98±0.95 | 8.17±0.32 | (28) |
| Zhao et al
(2010) | USA | GEO: GSE22981 | GPL8179 | 20/20 | 9.39±1.95 | 9.00±1.75 | (29) |
| Romero-Cordoba
et al (2012) | Mexico | GEO: GSE35412 | GPL9731 | 34/6 | 4.41±0.30 | 4.13±0.42 | (30) |
| Gravgaard et
al (2012) | Sweden | GEO: GSE37407 | GPL13703 | 50/5 | 6.23±0.03 | 6.27±0.01 | (31) |
| Biagioni et
al (2012) | Israel | GEO: GSE40525 | GPL8227 | 61/59 | 2.45±0.33 | 2.42±0.33 | (32) |
| Schrauder et
al (2012) | Germany | GEO: GSE31309 | GPL14132 | 48/57 | 5.81±0.79 | 5.55±0.69 | (33) |
| Godfrey (2013) | USA | GEO: GSE44281 | GPL14613 | 205/205 | 2.24±0.70 | 2.17±0.59 | (16) |
| Tanic (2013) | Spain | GEO: GSE32922 | GPL7723 | 22/16 | 6.08±0.09 | 6.09±0.09 | No ref.
available |
| Yan (2015) | Australia | GEO: GSE61438 | GPL8179 | 44/13 | 7.96±0.68 | 7.43±0.97 | (34) |
| Feliciano
(2013) | Spain | GEO: GSE44124 | GPL14767 | 50/3 | 5.42±0.09 | 5.31±0.07 | (35) |
| Matamala
(2015) | Spain | GEO: GSE58606 | GPL18838 | 121/11 | 6.43±0.71 | 6.51±0.04 | (36) |
Prospective molecular mechanism of
miR-671-3p in BC, as determined by enrichment analyses
The putative targets of miR-671-3p were predicted in
12 online prediction databases (Targetscan, RNAhybrid, RNA22, PITA,
Pictar2, miRWalk, Microt4, miRNAMap, miRDB, mirbridge, miRanda and
miRMap), and targets confirmed by >3 algorithms were selected
for further analyses. A total of 1,470 targets combined were
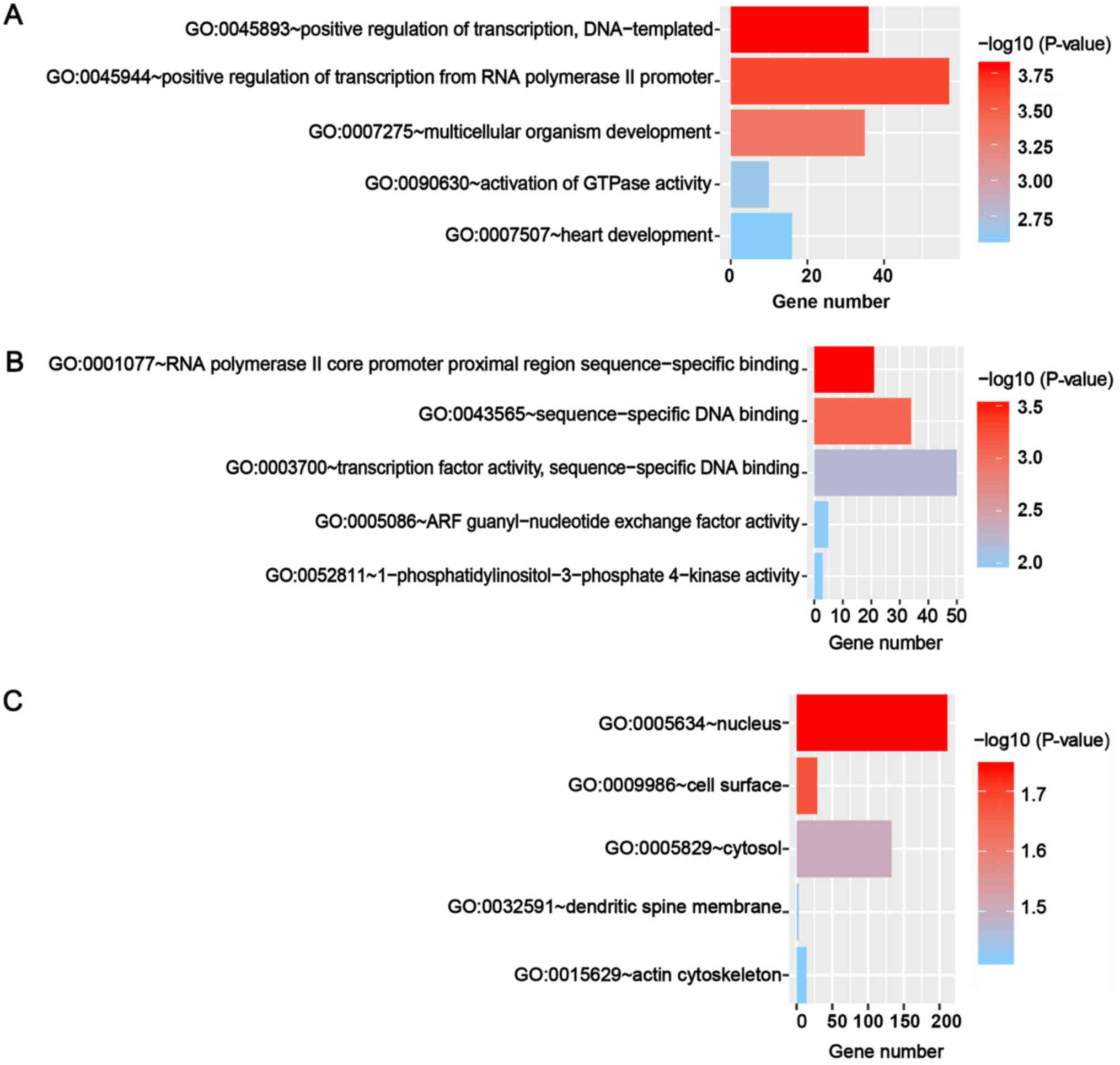
obtained by GO and KEGG analyses. The GO analyses included three
categories, namely biological process, molecular function and
cellular component. The top five GO annotations are shown in
Fig. 6, and the results suggested
that the targets of miR-671-3p were markedly enriched in the cell
nucleus and participated in transcription regulation. The top 10
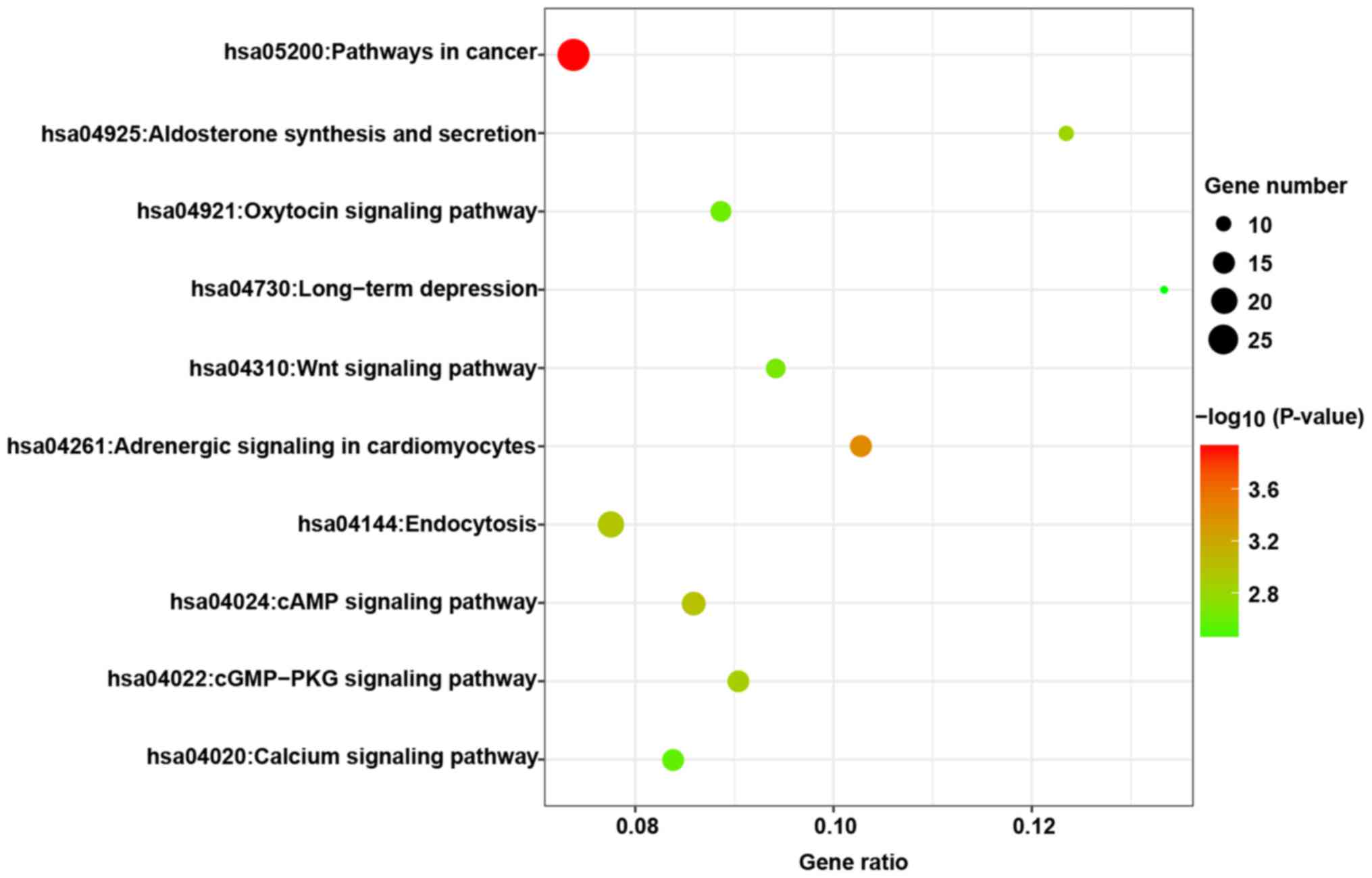
KEGG pathways are presented in Fig.
7, and it was observed that these predicted targets may be
involved in several tumor-associated pathways, including pathways
in cancer and the Wnt signaling pathway.
Construction of PPI network and
miRNA-gene regulatory network
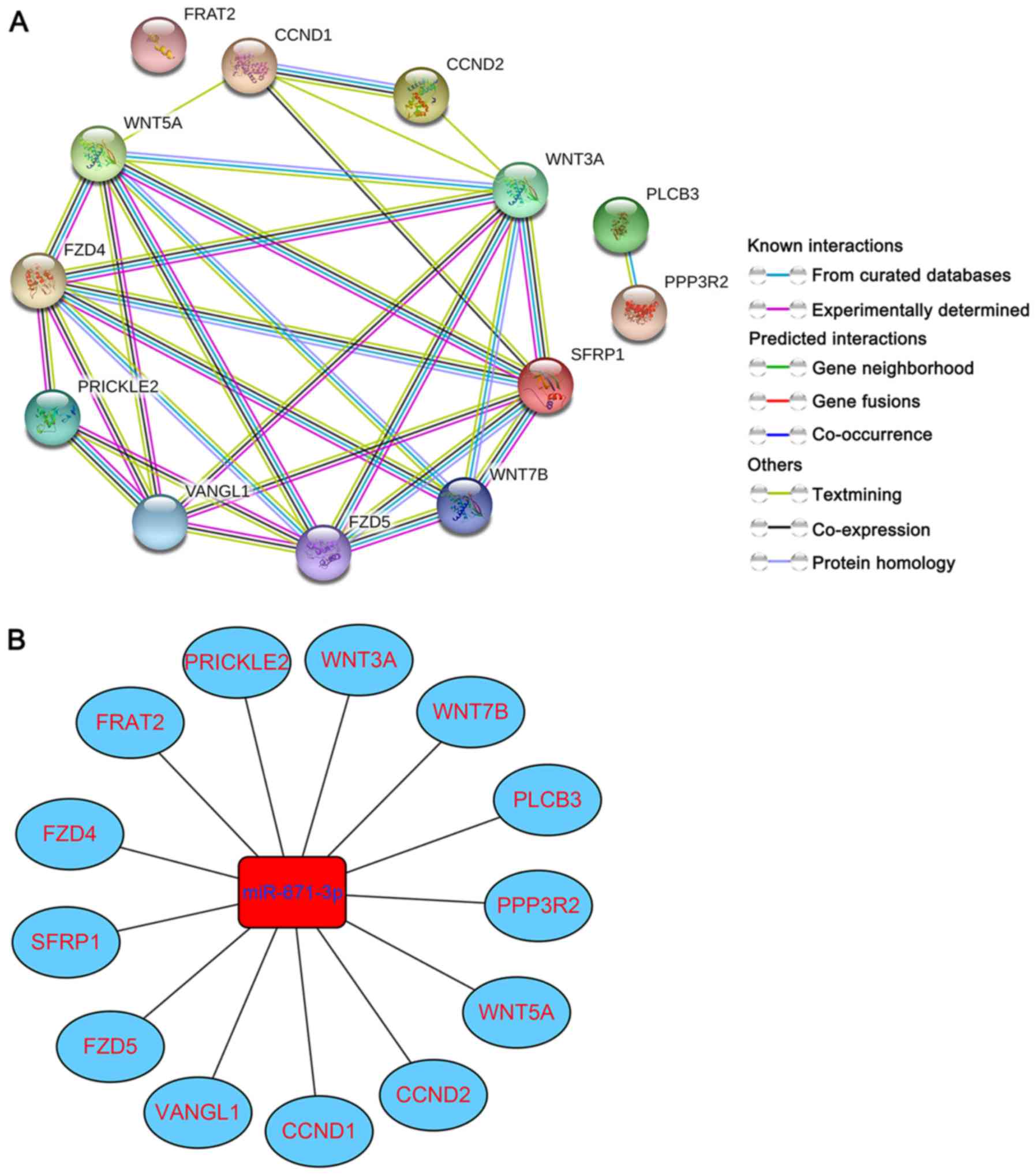
To further delve into the potential molecular
mechanism of miR-671-3p in BC, targets enriched in the Wnt
signaling pathway were selected for PPI network construction. The
following 13 genes were selected: WNT5A, WNT7B, WNT3A, PLCB3,
CCND1, CCND2, VANGL1, SFRP1, PRICKLE2, PPP3R2, FRAT2, FZD5 and
FZD4. As shown in Fig. 8A, a total
of 13 nodes and 29 edges were involved in the PPI network.
Simultaneously, the miRNA-gene regulatory network was formed based
on the 13 miRNA-gene pairs (Fig.
8B).
Subsequently, the expression levels of the 13 genes
were verified using data from TCGA. Due to the expression of genes
WNT3A and PPP3R2 being zero in >10% of samples, those genes were
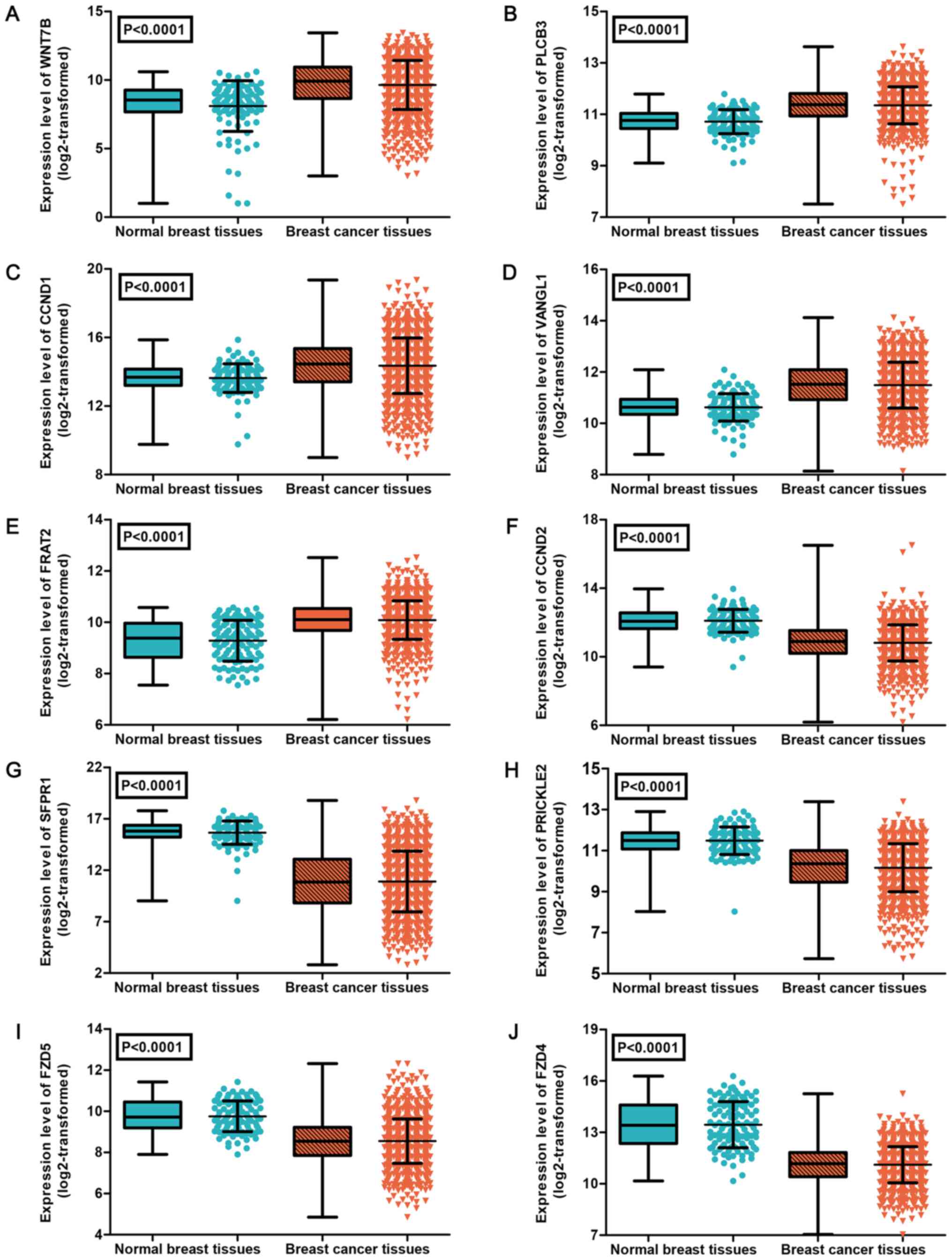
removed from the expression analysis (37). As shown in Table IV, the results identified 10
differently expressed target genes in BC, including 5 upregulated
genes (WNT7B, PLCB3, CCND1, VANGL1 and FRAT2) and 5 downregulated
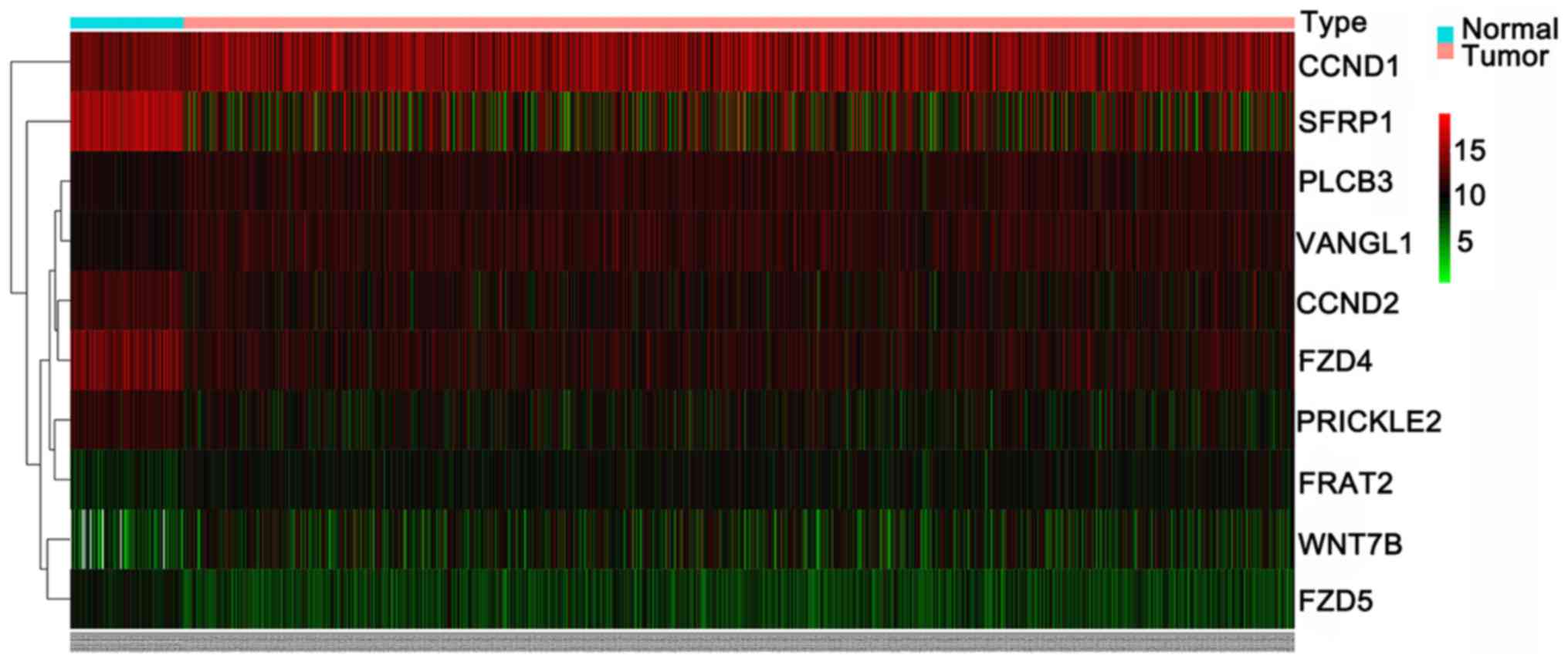
genes (CCND2, SFRP1, PRICKLE2, FZD5 and FZD4). The expression
patterns of the 10 differently expressed genes were visualized as a
heatmap (Fig. 9) and box-scatter
plots (Fig. 10).
| Table IVExpression levels of the 11 target
genes of miR-671-3p based on data obtained from TCGA. |
Table IV
Expression levels of the 11 target
genes of miR-671-3p based on data obtained from TCGA.
| Gene | Sample size
(T/N) | miR-671-3p
expressiona
| t-test
|
|---|
| Tumor | Normal | t-value | P-value |
|---|
| WNT5A | 1109/113 | 10.03±1.62 | 10.16±1.03 | 1.234 | 0.219 |
| WNT7B | 1109/108 | 9.64±1.79 | 7.61±2.66 | 7.975 | <0.0001 |
| PLCB3 | 1109/113 | 11.35±0.72 | 10.71±0.47 | 12.995 | <0.0001 |
| CCND1 | 1109/113 | 14.36±1.62 | 13.63±0.84 | 7.832 | <0.0001 |
| VANGL1 | 1109/113 | 11.49±0.89 | 10.62±0.54 | 15.203 | <0.0001 |
| FRAT2 | 1109/113 | 10.09±0.75 | 9.29±0.79 | 10.238 | <0.0001 |
| CCND2 | 1109/113 | 10.81±1.06 | 12.10±0.67 | 18.331 | <0.0001 |
| SFRP1 | 1109/113 | 10.89±2.96 | 15.66±1.13 | 34.327 | <0.0001 |
| PRICKLE2 | 1109/113 | 10.16±1.17 | 11.48±0.67 | 18.326 | <0.0001 |
| FZD5 | 1109/113 | 8.55±1.08 | 9.76±0.74 | 15.64 | <0.0001 |
| FZD4 | 1109/113 | 11.11±1.06 | 13.45±1.34 | 17.961 | <0.0001 |
Discussion
BC is an aggressive cancer that threatens the health
of women. It is the most prevalent tumor and the main cause of
oncogenic mortality in women worldwide, with an estimated 1,676,600
newly diagnosed cases and 521,900 associated mortalities annually
(38). Although specific molecular
biomarkers for optimizing clinical management have been explored
(39–41), their clinical application remains
limited due to the heterogeneity and complexity of BC. Therefore,
probing effective biomarkers to better understand the molecular
mechanism of BC and to provide appropriate target treatments for BC
patients is essential. The aim of the present study was to address
the expression level, clinical value, biological function and
molecular mechanism of miR-671-3p in BC and, further, to explore
the potential clinical usability of miR-671-3p for therapeutic
decisions in BC.
In the present study, in vitro experiments
were conducted to examine the biological effects of miR-671-3p on
the proliferation and apoptosis of BC cells by transfecting a
miR-671-3p mimic into MDA-MB-231 cells. The results revealed that
the miR-671-3p mimic evidently attenuated cell proliferation and
induced cell apoptosis, indicating that miR-671-3p was a tumor
suppressor in BC.
Only one previous study conducted by Godfrey et
al (16) has reported the
expression pattern of miR-671-3p in BC. The authors detected miRNA
expression in serum of BC patients and healthy participants using
array Affymetrix arrays and demonstrated that 16 miRNAs, including
miR-671-3p, were overexpressed in the serum of BC patients. The
authors validated the expression levels of the three miRNAs with
the highest expression using RT-qPCR; however, no statistically
significant differences were detected. Given the low accuracy of
gene chip technology, large-scale studies based on RT-qPCR are
imperative for verifying the serum level of miR-671-3p in BC
patients. In the present study, miR-671-3p expression was examined
in BC and non-tumorous tissues using RT-qPCR, and the results
indicated that miR-671-3p was downregulated in cancer tissues. All
available relevant microarray datasets were also collected in order
to further confirm the expression level of miR-671-3p in BC.
However, the result revealed that the expression of miR-671-3p in
BC tissues was similar to that in the normal controls, which was
not consistent with the results of RT-qPCR. Differences in sample
sources and detection methods may explain this inconsistency in the
findings. For instance, the expression of miR-671-3p may vary
across individuals. In addition, the accuracy of RT-qPCR is higher
compared with that of microarray analysis. Although a total of 11
microarray datasets were included to assess the expression level of
miR-671-3p in BC and the pooled result was stable, the results
should be interpreted with caution, since these datasets were
obtained from different platforms, resulting in a significant
heterogeneity in the current study. Additionally, the confounders
induced by different RNA extraction methods and diverse RNA
detection platforms may also limit the validity of the present
meta-analysis results. Thus, further and larger-scale RT-qPCR-based
investigations will be indispensable in deciphering the exact
expression of miR-671-3p in BC.
The association of miR-671-3p expression with the
clinicopathological parameters and overall survival of patients was
also analyzed to determine whether miR-671-3p can function as a
predictive indicator in BC. However, no association between
miR-671-3p expression and the clinicopathological factors was
detected. Patients with high miR-671-3p expression survived longer
in comparison with patients with low miR-671-3p expression,
however, no statistically significant difference was observed.
Considering the relatively small sample size of 38 cases in the
present study, further investigation including more samples is
required to determine the prognostic value of miR-671-3p in
patients with BC.
Finally, the in silico GO and KEGG analyses
in the present study revealed the potential molecular mechanism of
miR-6713p in the tumorigenesis of BC. It was observed that the
predicted targets of miR-671-3p were involved in several
tumor-associated pathways, including the Wnt signaling pathway. The
Wnt signaling pathway is known to be closely linked with the
initiation and progression of various malignant tumors, including
BC, by driving of the epithelial-mesenchymal transition and
metastasis of tumor cells (42-47).
Accumulating evidence has suggested that the Wnt signaling cascade
is regulated by miRNAs (46,48-51).
A study performed by Cai et al (46) indicated that miR-374a activates the
Wnt signaling pathway in BC by inhibiting negative modulators of
the Wnt signaling cascade, including WNT5A, WIF1 and PTEN. A study
conducted by Yi et al (51)
demonstrated that miR-214 negatively regulates the Wnt signaling
cascade and, consequently, inhibits cell proliferation in BC.
However, the regulatory effects of miR-671-3p on the Wnt signaling
pathway have yet to be determined. In the current study, it was
observed that 13 target genes (WNT5A, WNT7B, WNT3A, PLCB3, CCND1,
CCND2, VANGL1, SFRP1, PRICKLE2, PPP3R2, FRAT2, FZD5 and FZD4) of
miR-671-3p were significantly enriched in the Wnt signaling
pathway. To further determine the roles of the 13 genes in BC,
their expression patterns were investigated using data from TCGA.
The results revealed that the genes WNT7B, PLCB3, CCND1, VANGL1 and
FRAT2 were evidently upregulated in BC, indicating that they may
act as oncogenes, while WNT3A, CCND2, SFRP1, PRICKLE2, FZD5 and
FZD4 were markedly downregulated in BC, suggesting that they may
function as tumor suppressor genes in this tumor. Furthermore, it
is inferred that miR-671-3p may suppress the tumorigenesis and
development of BC by interacting with these genes, and further
inhibiting the Wnt signaling pathway. However, further rigorous
investigation is requited to confirm the targeting effects of
miR-671-3p on these genes.
In conclusion, the present study provided evidence
that miR-671-3p functions as a tumor suppressor to inhibit the
initiation and development of BC by influencing the Wnt signaling
pathway, as observed by in vitro experiments, in-house
RT-qPCR, microarray datasets and bioinformatics analyses. The
findings also demonstrated a possible molecular mechanism of
miR-671-3p in BC, indicating that miR-671-3p may be a prospective
molecular target for the therapy of BC. Further investigation is
required to verify these conclusions.
Acknowledgments
Not applicable.
Funding
This study was supported by grants from the Natural
Science Foundation of Guangxi, China (grant nos. 2017GXNSFAA198067
and 2015GXNSFAA139187).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DDX collected and analyzed the data from the GEO and
Oncomine databases, participated in the statistical analyses and
was a major contributor in the writing of the manuscript. HC
performed in-house RT-qPCR and in vitro experiments. RQH,
AHL and JCZ conducted the bioinformatics analyses. GC examined all
of the data presented in the manuscript. ZBF and KLW guided the
design of all of the experiments and the writing of the manuscript.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
All participants provided informed consent prior to
sample collection. The Ethical Committee of the First Affiliated
Hospital of Guangxi Medical University approved this
investigation.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sayed D and Abdellatif M: MicroRNAs in
development and disease. Physiol Rev. 91:827–887. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fan FY, Deng R, Yi H, Sun HP, Zeng Y, He
GC and Su Y: The inhibitory effect of MEG3/miR-214/AIFM2 axis on
the growth of T-cell lymphoblastic lymphoma. Int J Oncol.
51:316–326. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yu M, Xue H, Wang Y, Shen Q, Jiang Q,
Zhang X, Li K, Jia M, Jia J, Xu J, et al: miR-345 inhibits tumor
metastasis and EMT by targeting IRF1-mediated mTOR/STAT3/AKT
pathway in hepatocellular carcinoma. Int J Oncol. 50:975–983. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
He R, Yang L, Lin X, Chen X, Lin X, Wei F,
Liang X, Luo Y, Wu Y, Gan T, et al: MiR-30a-5p suppresses cell
growth and enhances apoptosis of hepatocellular carcinoma cells via
targeting AEG-1. Int J Clin Exp Pathol. 8:15632–15641. 2015.
|
|
6
|
Xie Z, Chen W, Chen Y, Wang X, Gao W and
Liu Y: miR-768-3p is involved in the proliferation, invasion and
migration of non-small cell lung carcinomas. Int J Oncol.
51:1574–1582. 2017. View Article : Google Scholar
|
|
7
|
Shang A, Yang M, Shen F, Wang J, Wei J,
Wang W, Lu W and Wang C and Wang C: MiR-1-3p suppresses the
proliferation, invasion and migration of bladder cancer cells by
up-regulating SFRP1 expression. Cell Physiol Biochem. 41:1179–1188.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen G, He M, Yin Y, Yan T, Cheng W, Huang
Z, Zhang L, Zhang H, Liu P, Zhu W, et al: miR-1296-5p decreases
ERBB2 expression to inhibit the cell proliferation in
ERBB2-positive breast cancer. Cancer Cell Int. 17:952017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ding Y, Zhang C, Zhang J, Zhang N, Li T,
Fang J, Zhang Y, Zuo F, Tao Z, Tang S, et al: miR-145 inhibits
proliferation and migration of breast cancer cells by directly or
indirectly regulating TGF-β1 expression. Int J Oncol. 50:1701–1710.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xiao G, Li X, Li G, Zhang B, Xu C, Qin S,
Du N, Wang J, Tang SC, Zhang J, et al: MiR-129 blocks estrogen
induction of NOTCH signaling activity in breast cancer stem-like
cells. Oncotarget. 8:103261–103273. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lu X, Ma J, Chu J, Shao Q, Zhang Y, Lu G,
Li J, Huang X, Li W, Li Y, et al: MiR-129-5p sensitizes the
response of Her-2 positive breast cancer to trastuzumab by reducing
Rps6. Cell Physiol Biochem. 44:2346–2356. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jang MH, Kim HJ, Gwak JM, Chung YR and
Park SY: Prognostic value of microRNA-9 and microRNA-155 expression
in triple-negative breast cancer. Hum Pathol. 68:69–78. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang L, Xu Y, Jin X, Wang Z, Wu Y, Zhao
D, Chen G, Li D, Wang X, Cao H, et al: A circulating miRNA
signature as a diagnostic biomarker for non-invasive early
detection of breast cancer. Breast Cancer Res Treat. 154:423–434.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiong DD, Lv J, Wei KL, Feng ZB, Chen JT,
Liu KC, Chen G and Luo DZ: A nine-miRNA signature as a potential
diagnostic marker for breast carcinoma: An integrated study of
1,110 cases. Oncol Rep. 37:3297–3304. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tan X, Fu Y, Chen L, Lee W, Lai Y, Rezaei
K, Tabbara S, Latham P, Teal CB, Man YG, et al: miR-671-5p inhibits
epithelial-to-mesenchymal transition by downregulating FOXM1
expression in breast cancer. Oncotarget. 7:293–307. 2016.
|
|
16
|
Godfrey AC, Xu Z, Weinberg CR, Getts RC,
Wade PA, DeRoo LA, Sandler DP and Taylor JA: Serum microRNA
expression as an early marker for breast cancer risk in
prospectively collected samples from the Sister Study cohort.
Breast Cancer Res. 15:R422013. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gradishar WJ, Anderson BO, Balassanian R,
Blair SL, Burstein HJ, Cyr A, Elias AD, Farrar WB, Forero A,
Giordano SH, et al: Invasive Breast Cancer Version 1.2016, NCCN
Clinical Practice Guidelines in Oncology. J Natl Compr Canc Netw.
14:324–354. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen G, Rong M and Luo D: TNFRSF6B
neutralization antibody inhibits proliferation and induces
apoptosis in hepatocellular carcinoma cell. Pathol Res Pract.
206:631–641. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rong M, Chen G and Dang Y: Increased
miR-221 expression in hepatocellular carcinoma tissues and its role
in enhancing cell growth and inhibiting apoptosis in vitro. BMC
Cancer. 13:212013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rong M, He R, Dang Y and Chen G:
Expression and clinicopathological significance of miR-146a in
hepatocellular carcinoma tissues. Ups J Med Sci. 119:19–24. 2014.
View Article : Google Scholar :
|
|
21
|
Chen G, Kronenberger P, Teugels E and De
Grève J: Influence of RT-qPCR primer position on EGFR interference
efficacy in lung cancer cells. Biol Proced Online. 13:12010.
View Article : Google Scholar
|
|
22
|
Chen G, Umelo IA, Lv S, Teugels E, Fostier
K, Kronenberger P, Dewaele A, Sadones J, Geers C and De Grève J:
miR-146a inhibits cell growth, cell migration and induces apoptosis
in non-small cell lung cancer cells. PLoS One. 8:e603172013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen G, Kronenberger P, Teugels E, Umelo
IA and De Grève J: Effect of siRNAs targeting the EGFR T790M
mutation in a non-small cell lung cancer cell line resistant to
EGFR tyrosine kinase inhibitors and combination with various
agents. Biochem Biophys Res Commun. 431:623–629. 2013. View Article : Google Scholar
|
|
24
|
Xiao F, Lan A, Lin Z, Song J, Zhang Y, Li
J, Gu K, Lv B, Zhao D, Zeng S, et al: Impact of CAG repeat length
in the androgen receptor gene on male infertility - a
meta-analysis. Reprod Biomed Online. 33:39–49. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kohl M, Wiese S and Warscheid B:
Cytoscape: Software for visualization and analysis of biological
networks. Methods Mol Biol. 696:291–303. 2011. View Article : Google Scholar
|
|
26
|
Phukan JP, Sinha A and Deka JP:
Cytological grading of breast carcinoma on fine needle aspirates
and its relation with histological grading. South Asian J Cancer.
4:32–34. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chavez-MacGregor M, Mittendorf EA, Clarke
CA, Lichtensztajn DY, Hunt KK and Giordano SH: Incorporating tumor
characteristics to the American Joint Committee on Cancer Breast
Cancer Staging System. Oncologist. 22:1292–1300. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fassan M, Baffa R, Palazzo JP, Lloyd J,
Crosariol M, Liu CG, Volinia S, Alder H, Rugge M, Croce CM, et al:
MicroRNA expression profiling of male breast cancer. Breast Cancer
Res. 11:R582009. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhao H, Shen J, Medico L, Wang D,
Ambrosone CB and Liu S: A pilot study of circulating miRNAs as
potential biomarkers of early stage breast cancer. PLoS One.
5:e137352010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Romero-Cordoba S, Rodriguez-Cuevas S,
Rebollar-Vega R, Quintanar-Jurado V, Maffuz-Aziz A, Jimenez-Sanchez
G, Bautista-Piña V, Arellano-Llamas R and Hidalgo-Miranda A:
Identification and pathway analysis of microRNAs with no previous
involvement in breast cancer. PLoS One. 7:e319042012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gravgaard KH, Lyng MB, Laenkholm AV,
Søkilde R, Nielsen BS, Litman T and Ditzel HJ: The miRNA-200 family
and miRNA-9 exhibit differential expression in primary versus
corresponding metastatic tissue in breast cancer. Breast Cancer Res
Treat. 134:207–217. 2012. View Article : Google Scholar
|
|
32
|
Biagioni F, Bossel Ben-Moshe N, Fontemaggi
G, Canu V, Mori F, Antoniani B, Di Benedetto A, Santoro R, Germoni
S, De Angelis F, et al: miR-10b*, a master inhibitor of the cell
cycle, is down-regulated in human breast tumours. EMBO Mol Med.
4:1214–1229. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schrauder MG, Strick R, Schulz-Wendtland
R, Strissel PL, Kahmann L, Loehberg CR, Lux MP, Jud SM, Hartmann A,
Hein A, et al: Circulating micro-RNAs as potential blood-based
markers for early stage breast cancer detection. PLoS One.
7:e297702012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yan M, Shield-Artin K, Byrne D, Deb S,
Waddell N, Haviv I and Fox SB; kConFab Investigators, kConFab:
Comparative microRNA profiling of sporadic and BRCA1 associated
basal- like breast cancers. BMC Cancer. 15:5062015. View Article : Google Scholar
|
|
35
|
Feliciano A, Castellvi J, Artero-Castro A,
Leal JA, Romagosa C, Hernández-Losa J, Peg V, Fabra A, Vidal F,
Kondoh H, et al: miR-125b acts as a tumor suppressor in breast
tumorigenesis via its novel direct targets ENPEP, CK2-α, CCNJ, and
MEGF9. PLoS One. 8:e762472013. View Article : Google Scholar
|
|
36
|
Matamala N, Vargas MT, González-Cámpora R,
Miñambres R, Arias JI, Menéndez P, Andrés-León E, Gómez-López G,
Yanowsky K, Calvete-Candenas J, et al: Tumor microRNA expression
profiling identifies circulating microRNAs for early breast cancer
detection. Clin Chem. 61:1098–1106. 2015. View Article : Google Scholar
|
|
37
|
Zeng JH, Liang L, He RQ, Tang RX, Cai XY,
Chen JQ, Luo DZ and Chen G: Comprehensive investigation of a novel
differentially expressed lncRNA expression profile signature to
assess the survival of patients with colorectal adenocarcinoma.
Oncotarget. 8:16811–16828. 2017.
|
|
38
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schnitt SJ: Classification and prognosis
of invasive breast cancer: From morphology to molecular taxonomy.
Mod Pathol. 23(Suppl 2): S60–S64. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Song C, Zhang L, Wang J, Huang Z, Li X, Wu
M, Li S, Tang H and Xie X: High expression of microRNA-183/182/96
cluster as a prognostic biomarker for breast cancer. Sci Rep.
6:245022016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Marino AL, Evangelista AF, Vieira RA,
Macedo T, Kerr LM, Abrahão-Machado LF, Longatto-Filho A, Silveira
HC and Marques MM: MicroRNA expression as risk biomarker of breast
cancer metastasis: A pilot retrospective case-cohort study. BMC
Cancer. 14:7392014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sun L, Liu T, Zhang S, Guo K and Liu Y:
Oct4 induces EMT through LEF1/β-catenin dependent WNT signaling
pathway in hepatocellular carcinoma. Oncol Lett. 13:2599–2606.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yanaka Y, Muramatsu T, Uetake H, Kozaki K
and Inazawa J: miR-544a induces epithelial-mesenchymal transition
through the activation of WNT signaling pathway in gastric cancer.
Carcinogenesis. 36:1363–1371. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jin B, Wang W, Meng XX, Du G, Li J, Zhang
SZ, Zhou BH and Fu ZH: Let-7 inhibits self-renewal of
hepatocellular cancer stem-like cells through regulating the
epithelial-mesenchymal transition and the Wnt signaling pathway.
BMC Cancer. 16:8632016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu ZJ, Liu HL, Zhou HC and Wang GC: TIPE2
Inhibits Hypoxia-induced Wnt/β-catenin pathway activation and EMT
in glioma cells. Oncol Res. 24:255–261. 2016. View Article : Google Scholar
|
|
46
|
Cai J, Guan H, Fang L, Yang Y, Zhu X, Yuan
J, Wu J and Li M: MicroRNA-374a activates Wnt/β-catenin signaling
to promote breast cancer metastasis. J Clin Invest. 123:566–579.
2013.PubMed/NCBI
|
|
47
|
Xiao C, Wu CH and Hu HZ: LncRNA UCA1
promotes epithelial-mesenchymal transition (EMT) of breast cancer
cells via enhancing Wnt/beta-catenin signaling pathway. Eur Rev Med
Pharmacol Sci. 20:2819–2824. 2016.PubMed/NCBI
|
|
48
|
Xiao G, Zhang B, Meng J, Wang J, Xu C,
Tang SC, Li X, Zhang J, Liang R, Ren H, et al: miR-367 stimulates
Wnt cascade activation through degrading FBXW7 in NSCLC stem cells.
Cell Cycle. 16:2374–2385. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Fasihi A, M Soltani B, Atashi A and Nasiri
S: Introduction of hsa-miR-103a and hsa-miR-1827 and hsa-miR-137 as
new regulators of Wnt signaling pathway and their relation to
colorectal carcinoma. J Cell Biochem. Aug 17–2017.Epub ahead of
print. PubMed/NCBI
|
|
50
|
Zhang W, Shen C, Li C, Yang G, Liu H, Chen
X, Zhu D, Zou H, Zhen Y, Zhang D, et al: miR-577 inhibits
glioblastoma tumor growth via the Wnt signaling pathway. Mol
Carcinog. 55:575–585. 2016. View Article : Google Scholar
|
|
51
|
Yi SJ, Li LL and Tu WB: MiR-214 negatively
regulates proliferation and WNT/β-catenin signaling in breast
cancer. Eur Rev Med Pharmacol Sci. 20:5148–5154. 2016.
|