Introduction
Bladder cancer is a common malignancy of the urinary
system, and is clinically characterized by a high recurrence rate
and rapid progression (1). An
estimated 429,800 new bladder cancer cases and 165,100
cancer-associated mortalities occurred in 2012 worldwide (2). In United States, an estimated 79,030
new bladder cancer cases and 16,870 bladder cancer-associated
mortalities were reported in 2017 (3). Histologically, ~75% of cases with
non-muscle-invasive bladder cancer result in a favorable prognosis;
however, the remaining 25% of cases with muscle-invasive bladder
cancer exhibit a poor prognosis (4-6). To
date, the molecular mechanism underlying bladder cancer oncogenesis
remains not fully defined; therefore, the molecular mechanisms
underlying the carcinogenic function and progression of bladder
cancer should be urgently investigated. This research will help to
develop novel approaches for early detection, prediction of
prognosis and effective therapy for invasive bladder cancer, as
well as provide a novel method for bladder cancer prevention,
eventually improving the prognosis and reducing the incidence of
bladder cancer.
MicroRNAs (miRNAs or miRs) are a class of non-coding
small RNA molecules of approximately 22 nucleotides in length,
which possess the capacity of oncogenic or anti-oncogenic activity
by binding to the 3′-untranslated regions (UTRs) of their target
mRNA molecules. miRNAs negatively regulate the expression of their
target genes and serve a vital role in the manipulation of human
diseases and tumorigenesis (7).
Accumulating studies have indicated that miRNAs function to
regulate various physiological and pathological processes,
including cell growth, migration, invasion, cell cycle
distribution, cell differentiation, apoptosis, oncogenesis, lipid
metabolism and drug resistance (8,9).
Since alterations in miRNA expression are associated with the
development and progression of bladder cancer, certain miRNAs may
serve as potential diagnostic markers or therapeutic strategies for
bladder cancer (10-14). For instance, Zhang et al
(15) have observed that miR-497
expression is able to inhibit bladder cancer cell proliferation,
migration and invasion. In addition, Yu et al (16) have demonstrated that miR-613
significantly inhibits bladder cancer cell proliferation, migration
and invasion. Wu et al (17) have also reported that miR-429
overexpression can reverse epithelial-mesenchymal transition (EMT)
by restoring the expression of E-cadherin in bladder cancer. The
present study focuses on miR-612, which has recently been reported
to be downregulated in numerous types of human cancer, and
functions as a putative tumor suppressor in hepatocellular
carcinoma (18,19) and colorectal cancer (20).
In the present study, the level of miR-612
expression in bladder cancer tissues was investigated, and its
association with the clinicopathological data of patients was
determined. The effects of miR-612 expression in bladder cancer
cells in vitro and in nude mouse xenografts were then
assessed, while bioinformatics analysis was performed to identify
and confirm the miR-612 target genes. The present study is expected
to provide novel insightful information regarding miR-612 in
bladder cancer development and progression to assist in further
investigation of this miRNA as a novel tumor biomarker or
therapeutic strategy for bladder cancer in the future.
Materials and methods
Patients and tissue specimens
Bladder cancer and adjacent non-cancerous tissue
(ANT) specimens were collected from 46 patients who underwent
surgical resection of tumor lesions at the Shanghai Tenth People's
Hospital of Tongji University (Shanghai, China) between January
2012 and December 2015. The clinical characteristics of these
patients are shown in Table I.
None of the patients had received any chemotherapy or radiotherapy
prior to surgery. Following surgery, tissue specimens were
immediately placed into liquid nitrogen or were snap-frozen in
liquid nitrogen, and stored at -80°C until further use. All
patients were diagnosed with invasive bladder cancer according to
the 2002 version of the American Joint Committee on Cancer/Union
for International Cancer Control tumor, lymph node and metastasis
(TNM) staging system (21). This
study was approved by the Ethics Committee of Shanghai Tenth
People's Hospital of Tongji University, and informed consent was
obtained from all patients or their relatives.
 | Table IClinicopathological characteristics
of the patients with bladder cancer. |
Table I
Clinicopathological characteristics
of the patients with bladder cancer.
| Variables | No. of cases |
|---|
| Age (years) | |
| <60 | 8 |
| ≥60 | 38 |
| Sex | |
| Male | 41 |
| Female | 5 |
| TNM stage | |
| T1-T2 | 19 |
| T3-T4 | 27 |
| Distant
metastasis | |
| No | 32 |
| Yes | 14 |
Target gene prediction of miR-612
To predict the target genes of miR-612 we conducted
a bioinformatics analysis using a panel of different algorithms
including DIANAmT (http://www.microrna.gr/microT) miRanda (http://www.microrna.org/microrna/home.do) miRDB
(http://www.mirdb.org/) PICTAR5 (http://www.pictar.org/) RNA22 (http://cbcsrv.watson.ibm.com/rna22.html) TargetScan
(http://www.targetscan.org/) and miRWalk
(http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2/).
miRWalk2.0 is an updated version of the database
miRWalk that can used to validated miRNAs binding sites from the
human, mouse and rat on their target genes (http://zmf.umm.uniheidelberg.de/apps/zmf/mirwalk2/)
(22,23). MiRWalk2.0 not only recorded the
gene sequence of micrornas binding sites, but also combines this
information with a comparison of binding sites resulting from 12
existing miRNA-target prediction programs (DIANA-microTv4.0,
DIANA-microT-CDS, miRanda-rel2010, mirBridge, miRDB4.0, miRmap,
miRNAMap, doRiNA i.e., PicTar2, PITA, RNA22v2, RNAhybrid 2.1 and
Targetscan 6.2), we predicted 21 possible target genes in 7
databases (DIANAmT, miRanda, miRDB, miRWalk PICTAR5, RNA22 and
Targetscan). Subsequently, based on the combined score of mirna-612
on Targetscan, 9 possible target genes above 95 score were selected
(LRRC41, DUSP14, ZNF318, GABRA5, SMAD6, YTHDF1, HOXA13, TSG101 and
ME1).
To further narrow the screening range, we retrieved
the 9 genes on PubMed by keywords '(Tumor) AND LRRC41', '(Tumor)
AND DUSP14', '(Tumor) AND ZNF318', '(Tumor) AND GABRA5', '(Tumor)
AND SMAD6', '(Tumor) AND YTHDF1', '(Tumor) AND HOXA13', '(Tumor)
AND TSG101', '(Tumor) AND ME1', excluding the genes that have been
studied in bladder cancer (HOXA13{Hu, 2017 #120} (24) and no high expression in other
tumors [LRRC41 (25), DUSP14
(26), ZNF318 (27), GABRA5 (28)].
Cell lines and culture
Human bladder cancer T24, UMUC3, 5637 and J82 cell
lines, and the immortalized human normal bladder epithelial cell
line SV-HUC-1 were obtained from type culture collection of the
Chinese Academy of Sciences (Shanghai, China). SV-HUC-1 cells were
maintained in F12K medium (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany), while T24, UMUC3 and 5637 cells were cultured in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) and J82 cells were cultured in Dulbecco's modified Eagle's
medium (Gibco; Thermo Fisher Scientific, Inc.). Cells were cultured
at 37°C in a humidified incubator with 5% CO2. All cell
culture media were supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin (Hyclone; GE Healthcare Life Sciences,
Logan, UT, USA).
Transient gene transfection
Human miR-612 mimic (miR-612) and the corresponding
negative control mimic (miR-NC), as well as malic enzyme 1 siRNA
(si-ME1) and the corresponding negative control siRNA (si-NC), were
purchased from GenePharma Co., Ltd. (Shanghai, China). A plasmid
carrying malic enzyme 1 (ME1) cDNA was purchased from RiboBio Co.,
Ltd. (Guangzhou, China). For transient gene transfection, bladder
cancer T24 cells were grown and transiently transfected with these
reagents using Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol for 6 h
at 37°C. The cells were then subjected to different assays.
RNA isolation and reverse
transcriptase-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was isolated from frozen tissues or
cultured cells using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. The
concentration and purity of the RNA samples were measured by a
NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Inc.).
For detection of miR-612, 1 µg total RNA was reverse
transcribed into cDNA using a One Step Prime script miRNA cDNA
Synthesis kit (Qiagen, Inc., Valencia, CA, USA), and qPCR was
performed using the KAPA SYBR FAST qPCR kit (Kapa Biosystems, Inc.,
Boston, MA, USA), with U6 mRNA used to normalize the miR-612
expression level. The primers used were as follows: miR-612
forward, 5-GCAGGGCTTCTGAGCTCCTTAA-3, U6 forward,
5-CAAATTCGTGAAGCGTTCCATAT-3; the common reverse primer was
purchased from RiboBio Co., Ltd. The amplification procedure was as
follows: 5 min at 95°C, followed by 40 cycles at 95°C for 30 sec
and 65°C for 45 sec.
For detection of the ME1 mRNA level, cDNA was
synthesized using the PrimeScript RT Reagent kit (Takara Bio, Inc.,
Tokyo, Japan), according to the manufacturer's protocol. qPCR for
mRNA detection was also performed using the KAPA SYBR FAST qPCR kit
(Kapa Biosystems, Inc.), The primers used were as follows: ME1
forward, 5′-GCAGGTCTCCTTGCAGCTCT-3′ and reverse, 5′-TCCAAGGCCATCACA
ATCAG-3′; GAPDH forward, 5′-ATGTCGTGGAGTCTACTGGC-3′ and reverse,
5′-TGACCTTGCCCACAGCCTTG-3′. The PCR parameters for relative
quantification were as follows: 2 min at 95°C, followed by 40
cycles of 45 sec at 57°C and 45 sec at 72°C. The ME1 mRNA level was
normalized to that of GAPDH. The relative expression levels of
miR-612 and ME1 were analyzed by the 2−∆∆Cq method
(29).
Cell proliferation and colony formation
assays
The cell proliferation rate was measured using the
Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan). Briefly, the transfected cells were seeded into
96-well plates at a density of 1×103 cells/well and
grown for up to 5 days. Subsequently, 10 µl CCK-8 reagent
was added into each well, and the cells were further incubated at
37°C for 2 h. Next, the optical density value was measured at 450
nm using a microplate spectrophotometer obtained from BioTek
Instruments, Inc. (Winooski, VT, USA).
For the tumor cell colony formation assay, 1,000
transfected cells/well were seeded into 6-well plates and cultured
for 14 days. Subsequent to culturing, the cell colonies were washed
three times with phosphate-buffered saline (PBS), then fixed with
75% ethanol and stained with 0.1% crystal violet solution. Cell
colonies with ≥50 cells were counted under a light microscope
(Olympus Corporation, Tokyo, Japan). The experiments were performed
in triplicate and repeated at least once.
Cell migration and invasion assays
Transwell chambers (Corning, Inc., Lowell, MA, USA)
containing 6.5-mm-diameter polycarbonate filters with an
8-µm pore size were used to measure the cell migration
capacity. Briefly, 5×104 transfected cells in RPMI-1640
medium (200 µl) without FBS were seeded into each insert
(24-well plates), and 600 µl cell culture medium containing
10% FBS was added to the bottom chamber. The cells were allowed to
migrate for 16 h at 37°C and 5% CO2. For the tumor cell
invasion assay, the filter of the Transwell chamber was precoated
with Matrigel (BD Biosciences, Franklin Lakes, NJ, USA), and the
cells were allowed to invade for 24 h. The remaining experimental
procedures were the same as those of the tumor cell migration
assay. At the end of the experiments, the cells remaining in the
upper inserts were carefully removed using a cotton swab, whereas
cells that had migrated or invaded into the reverse side of the
filters were fixed with 70% ethanol for 30 min and stained with
0.1% crystal violet for 10 min. Images of five randomly selected
microscopic fields were captured under a microscope (Olympus
Corporation), and then the number of cells was counted. The
experiments were performed in triplicate and repeated at least
once.
Luciferase reporter assay
The wild-type 3′-UTR segment and a mutant 3′-UTR
segment of ME1 cDNA were amplified using PCR, and then inserted
into the luciferase gene using the psiCHECK-2 vector (Promega
Corporation, Madison, WI, USA), according to the manufacturer's
instructions. Luciferase assay was then performed in T24 cells,
during which tumor cells (1×105) were seeded into
24-well plates and grown overnight. On the following day, 100 ng
luciferase reporter vectors and 100 nM miR-612 or miR-NC were
transfected into the T24 cells using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) for 48 h. Thereafter,
the cell protein was extracted and the luciferase activity was
measured using a luciferase reporter assay system (Promega
Corporation), according to the manufacturer's protocol.
Western blot analysis
Total cellular protein was extracted from the
transfected cells using a radioimmunoprecipitation assay buffer
(Sigma-Aldrich) containing protease inhibitor, and the protein
concentration of these samples was measured using a bicinchoninic
acid assay kit (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Protein samples of 20 µg each were
separated with 10% sodium dodecyl sulfate-polyacrylamide gels by
electrophoresis and then blotted onto nitrocellulose membranes
(Sigma-Aldrich; Merck KGaA). For western blotting, the membranes
were blocked with 5% non-fat milk for 1 h at 37°C and immunoblotted
at 4°C overnight with primary antibodies against: ME1 (sc-365891,
1:1000, secondary antibodies: Mouse), E-cadherin (sc-52327, 1:1000
secondary antibodies: Mouse), N-cadherin (sc-53488, 1:1000,
secondary antibodies: Mouse), vimentin (sc-373717, 1:1000,
secondary antibodies: Mouse), matrix metalloproteinase-9 (MMP-9)
(sc-12759, 1:1,000, secondary antibodies: Mouse), and anti-β-actin
(sc-47778, 1:1000, secondary antibodies: Mouse) (all purchased from
Santa Cruz Biotechnology, Dallas, TX, USA). The membranes were
subsequently incubated with the fluoresence-conjugated secondary
antibodies (926-68072 or 926-32210, LI-COR Biosciences, Shanghai,
China) for 1 h at room temperature, and the protein bands were
visualized and quantified by using the Odyssey two-color infrared
laser imaging system (LI-COR Biosciences, Lincoln, NE, USA).
Animal experiments
The animal experiments conducted in the present
study were approved by the Animal Care and Use Committee of Tongji
University. Briefly, 6 male nude BALB/c mice (age, 6 weeks old;
weight, 18-22 g) were purchased from Shanghai SIPPR-Bk Lab Animal
Co., Ltd. (Shanghai, China) and housed in specific-pathogen-free
conditions with a 12-h light/dark cycle (temperature, 18–22°C;
humidity, 50–60; irradiated feed and double-distilled water).
Bladder cancer T24 cells stably expressing miR-612 or miR-NC were
harvested, washed twice with PBS and resuspended in PBS. A total of
5×106 cells were then subcutaneously injected into the
left flank regions of each nude mouse. Tumor cell xenograft
formation was measured every 5 days using a Vernier caliper, and
the tumor volume was calculated using the following formula: Volume
(mm3) = 0.5 × width2 × length. At 5 weeks
after tumor cell inoculation, the mice were sacrificed by cervical
dislocation. Tumor xenografts were resected, weighed and then
stored at −80°C until further use.
Statistical analysis
SPSS software (version 16.0; SPSS, Inc., Chicago,
IL, USA) was used for all statistical analyses. The experimental
data were statistically analyzed using the Student's t-test or
one-way analysis of variance, and are expressed as the mean ±
standard deviation from three independent experiments. The
χ2 test or Fisher's exact test was performed to examine
the correlation of miR-612 expression with the clinicopathological
data of bladder cancer patients, whereas the association of miR-612
with ME1 expression was statistically analyzed using Spearman's
correlation test. P<0.05 was regarded to indicate a
statistically significant difference.
Results
Downregulation of miR-612 expression in
bladder cancer tissues and cell lines
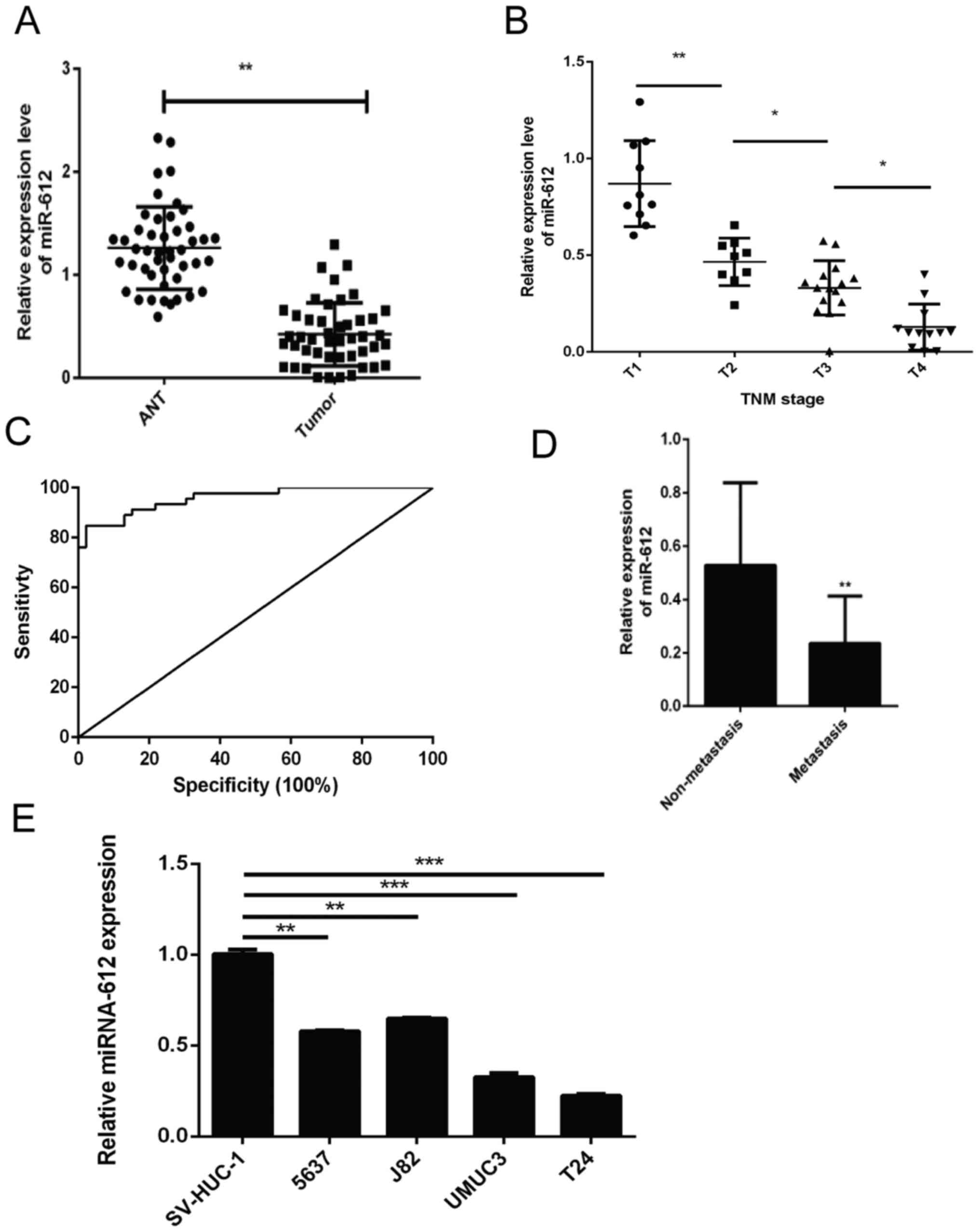
In the present study, the levels of miR-612
expression in 46 pairs of bladder cancer and ANT specimens were
initially assessed using RT-qPCR. It was observed that the miR-612
expression level was significantly lower in bladder cancer tissues
compared with that in ANTs (P<0.01; Fig. 1A). Next, the association of miR-612
expression with the clinicopathological features of the patients
was examined, and loss of miR-612 expression was found to be
significantly associated with advanced TNM stages (Fig. 1B) and tumor distant metastasis
(Fig. 1D). Furthermore, the
receiver operating characteristic curve analysis demonstrated that
tissue miR-612 levels may be a potential biomarker for
distinguishing bladder cancer patients from the controls (area
under the curve, 0.958; Fig. 1C).
Consistently, miR-612 expression was also significantly lower in
T24, UMUC3, 5637 and J82 bladder cancer cells as compared with that
in the normal bladder epithelial cell line SV-HUC-1. T24 cells
exhibited the lowest miR-612 levels among these four bladder cancer
cell lines (Fig. 1E).
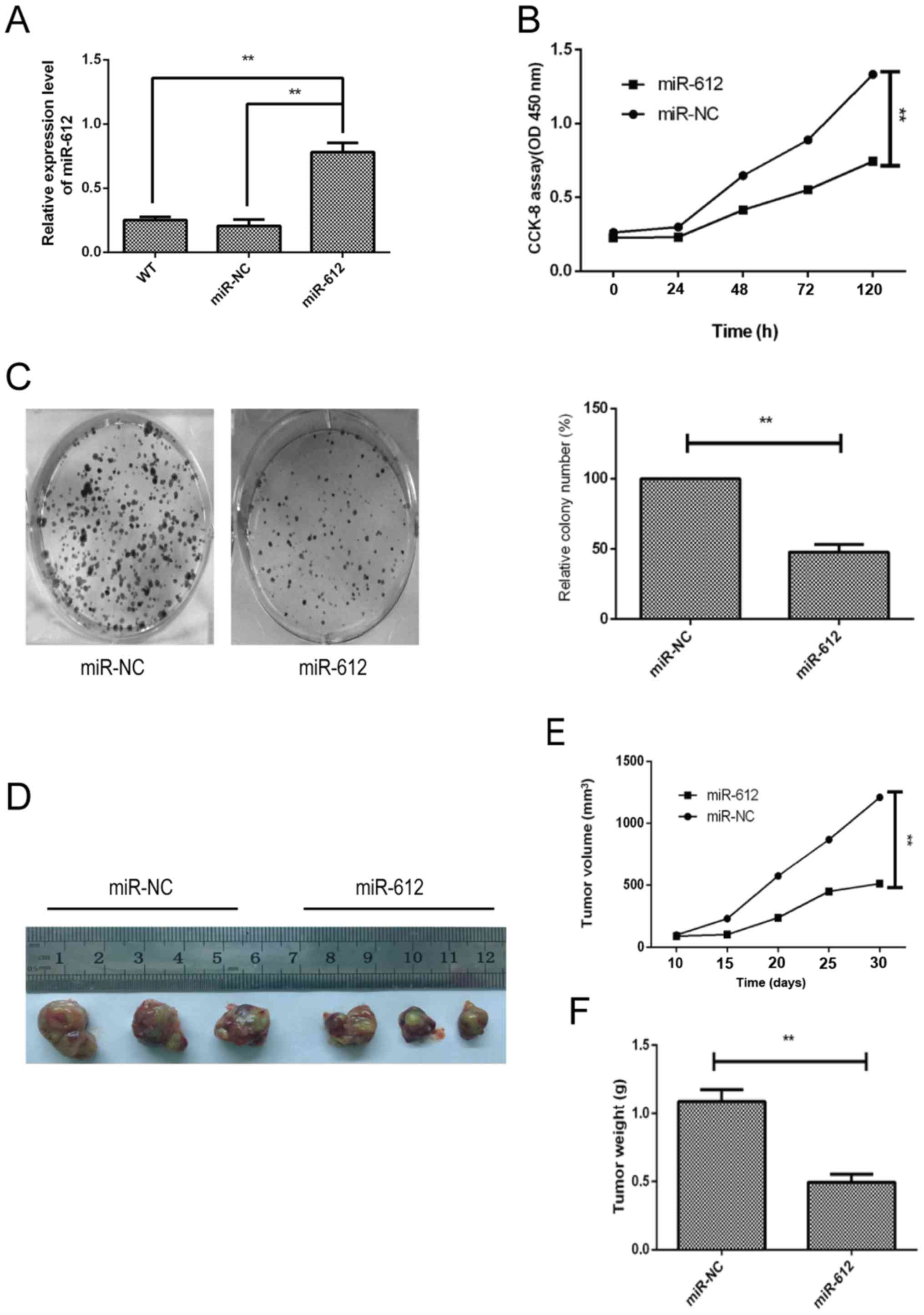
Inhibition of bladder cancer growth
following miR-612 overexpression in vitro and in vivo
To determine the biological effects of miR-612 on
bladder cancer cell growth, miR-612 mimic or negative control was
transfected into T24 cells, since this cell line had the lowest
level of miR-612 expression (Fig.
1E). The data revealed that the miR-612 mimic was successfully
transfected into the cells and increased miR-612 expression; miR-NC
transfection had no significant effect on the T24 cells (Fig. 2A). The CCK-8 and colony formation
assays demonstrated that miR-612 overexpression in T24 cells
significantly reduced tumor cell growth (Fig. 2B) and colony formation (Fig. 2C). Furthermore, the nude mouse
experiments indicated that T24 cells with miR-612 overexpression
resulted in slower growth of the tumor xenografts as compared with
that for miR-NC-transfected T24 cells (Fig. 2E). Consistently, the weight
(Fig. 2F) and tumor xenograft size
(Fig. 2D) of the
miR-612-transfected group were significantly reduced in comparison
with those of the miR-NC-transfected group.
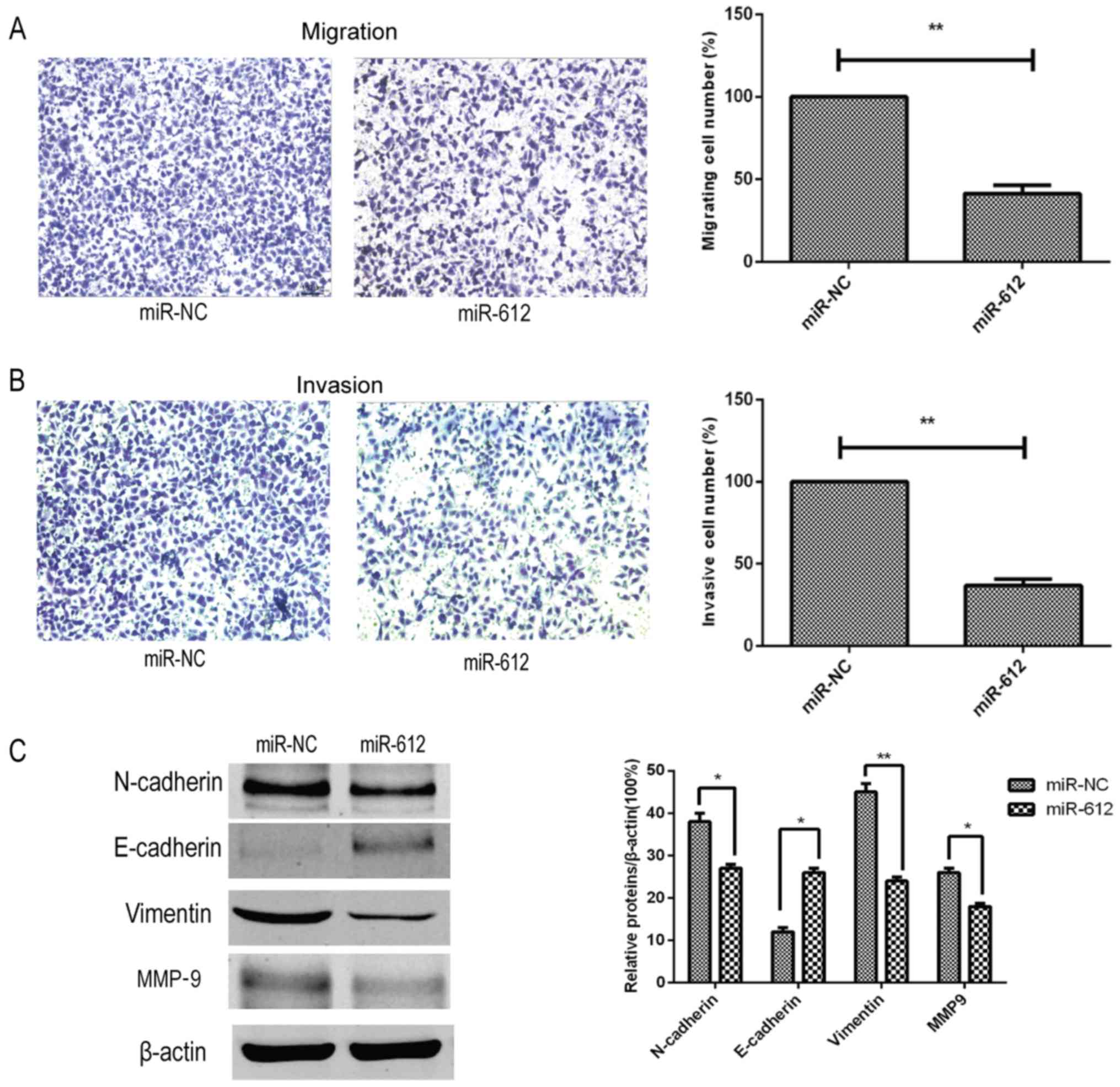
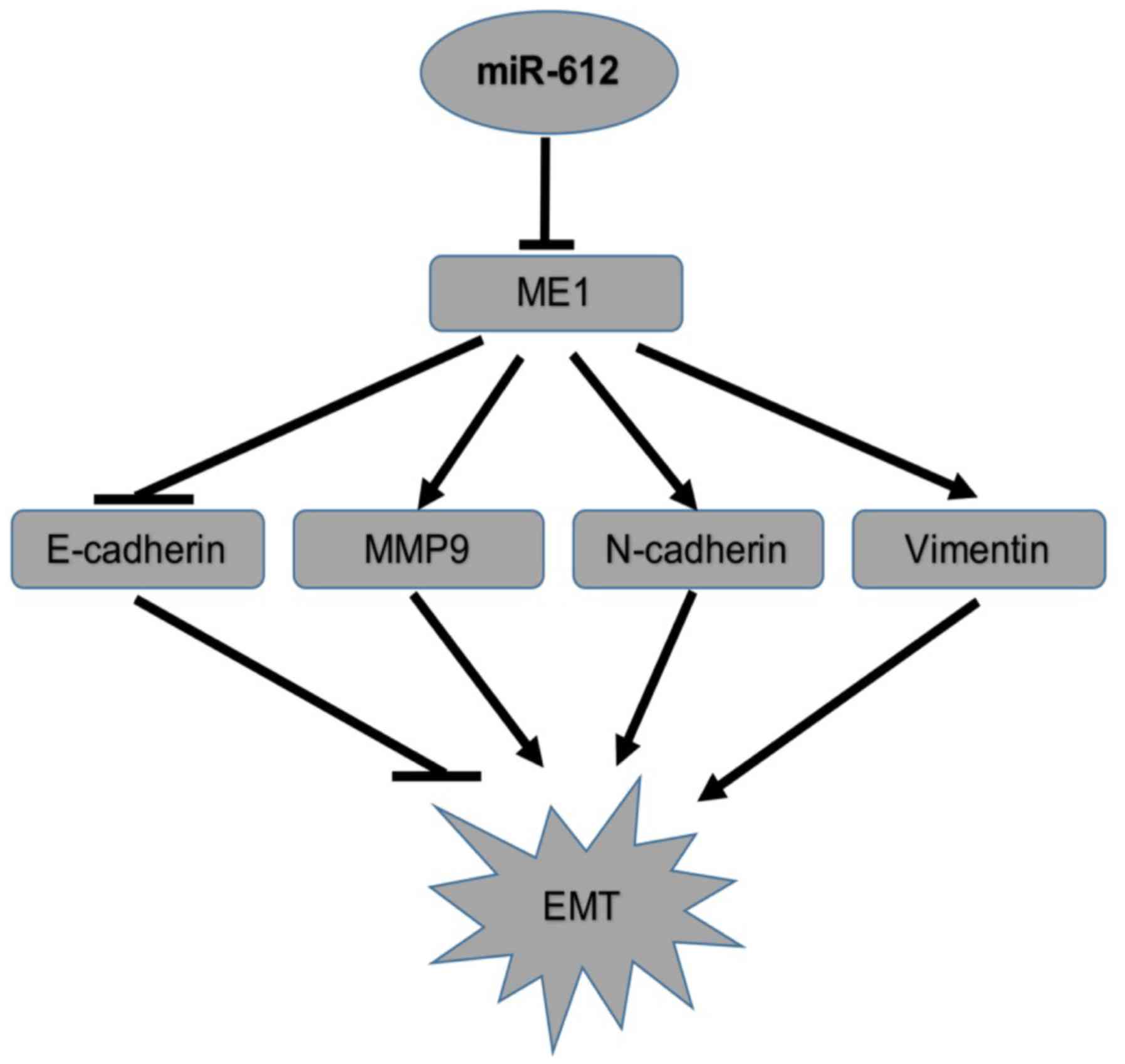
miR-612 inhibition of tumor cell
migration, invasion and EMT in vitro
Next, the effects of miR-612 on the metastatic
capacity of bladder cancer cells were assessed in vitro, and
it was observed that restoration of miR-612 expression in T24 cells
using the miR-612 mimic significantly reduced the tumor cell
migration and invasion abilities in vitro (Fig. 3A and B). Furthermore, the
expression of several accepted EMT-associated markers was also
affected by the miR-612 mimic. Compared with the miR-NC group, the
expression levels of N-cadherin, vimentin and MMP-9 were
significantly decreased by miR-612 mimic transfection, whereas the
expression level of E-cadherin was significantly upregulated in the
miR-612 group (Fig. 3C). During
EMT, N-cadherin, vimentin and MMP-9 levels are upregulated, and
E-cadherin levels are downregulated. Thus, our findings indicated
that miR-612 overexpression inhibited EMT.
ME1 is a the direct target of
miR-612
Since miRNAs alter the expression of their target
genes to exert their biological functions in cells, bioinformatic
analysis was performed using several software (DIANAmT, miRanda,
miRDB, miRWalk, PICTAR5, RNA22 and Targetscan) to predict the
possible targets of miR-612. Subsequently, relevant publications on
these genes were retrieved from Pubmed, and genes that have been
widely studied in bladder cancer and were not highly expressed in
other tumors were excluded. Finally, it was detected that miR-612
was able to bind to the 3′-UTR of ME1 cDNA (Fig. 4A). This binding ability of miR-612
to the ME1 3′-UTR was then confirmed using a luciferase reporter
assay. The results revealed that miR-612 was able to precisely bind
to the wild-type 3′-UTR of ME1, which resulted in significantly
inhibited luciferase activity (Fig.
4B). Furthermore, it was demonstrated that miR-612
overexpression in T24 cells significantly downregulated the mRNA
(Fig. 4C) and protein (Fig. 4D) levels of ME1. The ex vivo
data further supported this observation, since the level of ME1
mRNA was markedly higher in bladder cancer tissues as compared with
that in ANTs (Fig. 4E). In
addition, the level of ME1 mRNA was inversely associated with the
level of miR-612 in bladder cancer tissues (Fig. 4F; r=−0.403; P<0.0001). These
results indicated that ME1 is indeed the direct target of miR-612
in bladder cancer.
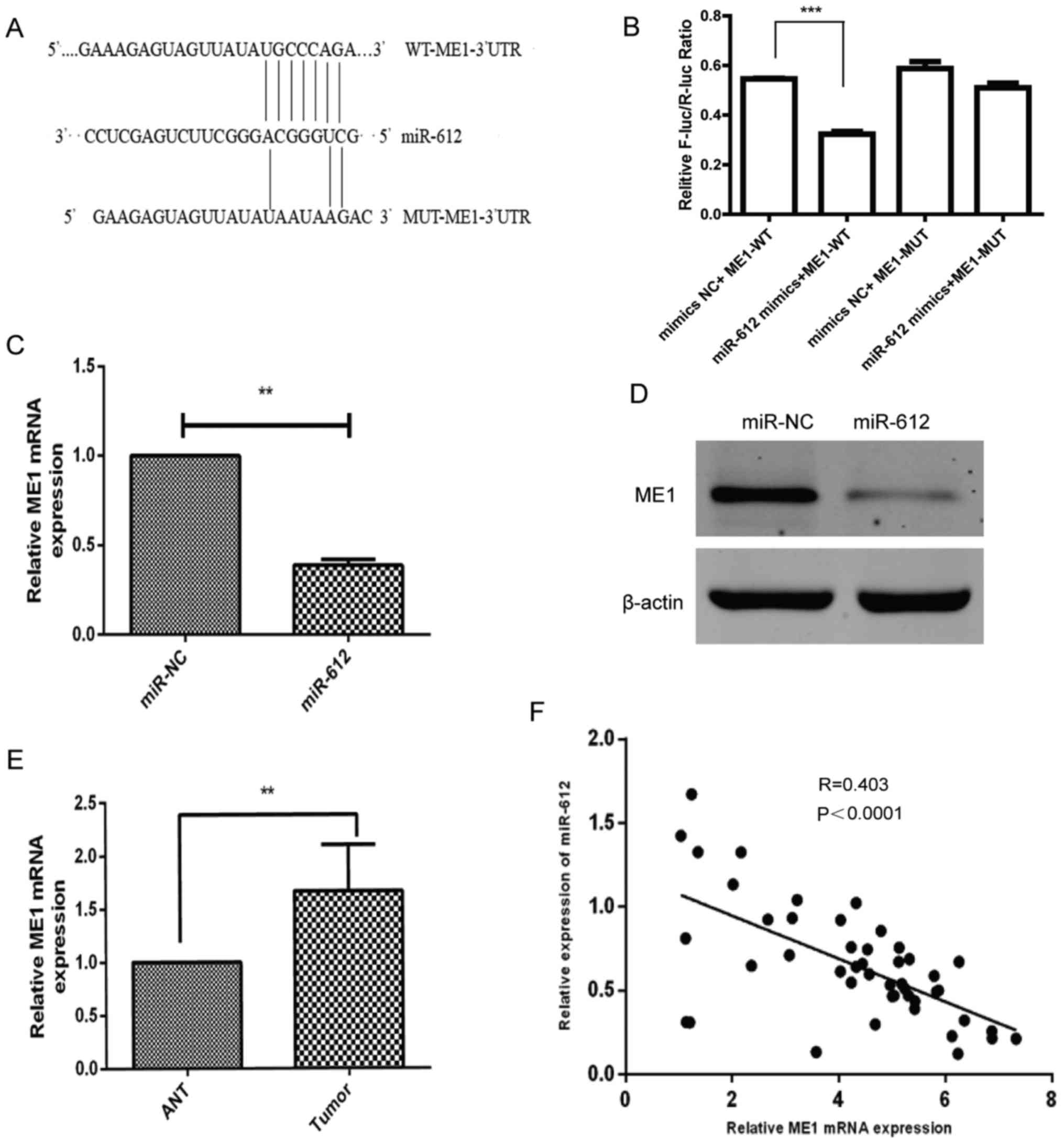 | Figure 4ME1 is a direct target of miR-612.
(A) Bioinformatic analysis indicated that ME1 is the target of
miR-612. The presumable miR-612-binding sites in the 3′-UTR of ME1
cDNA and the mutant binding sites are shown. (B) The relative
luciferase activity was determined in T24 cells following
co-transfection with a luciferase reporter plasmid (WT or MUT
3′-UTR ME1 cDNA), and miR-612 mimic or miR-NC. (C) mRNA and (D)
protein levels of ME1 in T24 cells following transfection with
miR-612 mimic or miR-NC were analyzed using RT-qPCR and western
blot analysis, respectively. GAPDH and β-actin served as the
internal controls, respectively. (E) The levels of ME1 mRNA were
measured using RT-qPCR in 46 paired bladder cancer and ANT
specimens. GAPDH was used as an internal control. (F) The
correlation between ME1 and miR-612 expression in bladder cancer
tissue samples (n=46) was analyzed using Spearman's correlation
test. **P<0.01 and ***P<0.001. miR,
microRNA; NC, negative control; ME1, malic enzyme 1; UTR,
untranslated region; WT, wild-type; MUT, mutated; ANT, adjacent
normal tissue; RT-qPCR, reverse transcription-quantitative
polymerase chain reaction. |
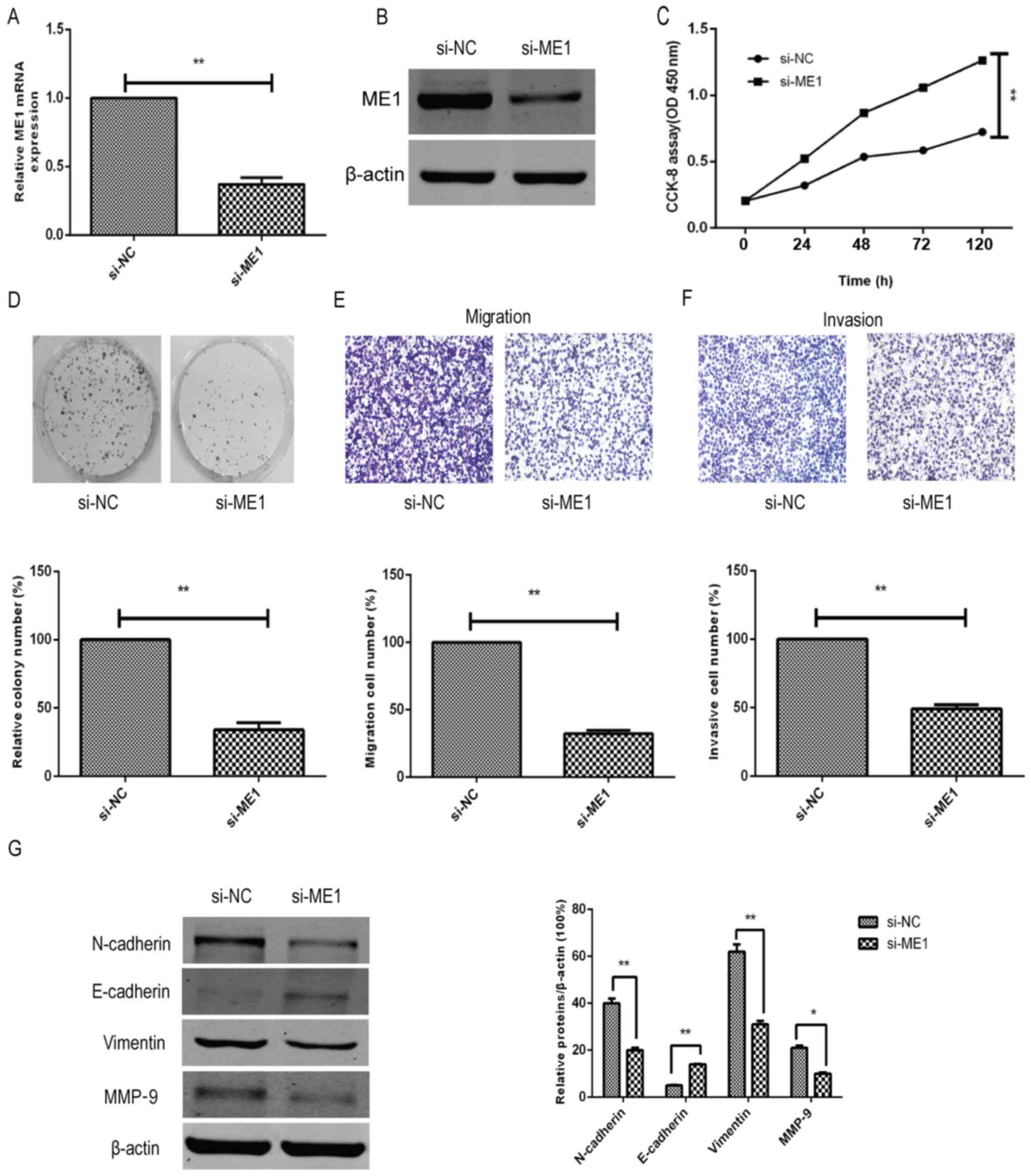
Parallel effects of ME1 knockdown and
overexpression with miR-612 manipulation on bladder cancer cells in
vitro
To further confirm the effect and biological
functions of ME1 in bladder cancer cells, ME1 expression was
knocked down in T24 cells using si-ME1 and compared against the
si-NC group. The data demonstrated that si-ME1 significantly
reduced the ME1 mRNA and protein expression levels in T24 cells, as
compared with the miR-NC-transfected T24 cells (Fig. 5A and B). In addition, functional
assays revealed that ME1 knockdown significantly inhibited the
tumor cell growth (Fig. 5C),
colony formation (Fig. 5D),
migration (Fig. 5E) and invasion
capacity (Fig. 5F), and affected
the tumor cell EMT marker levels (Fig.
5G) in T24 cells, mimicking the effects of miR-612
overexpression on T24 cells (Figs.
2 and 3).
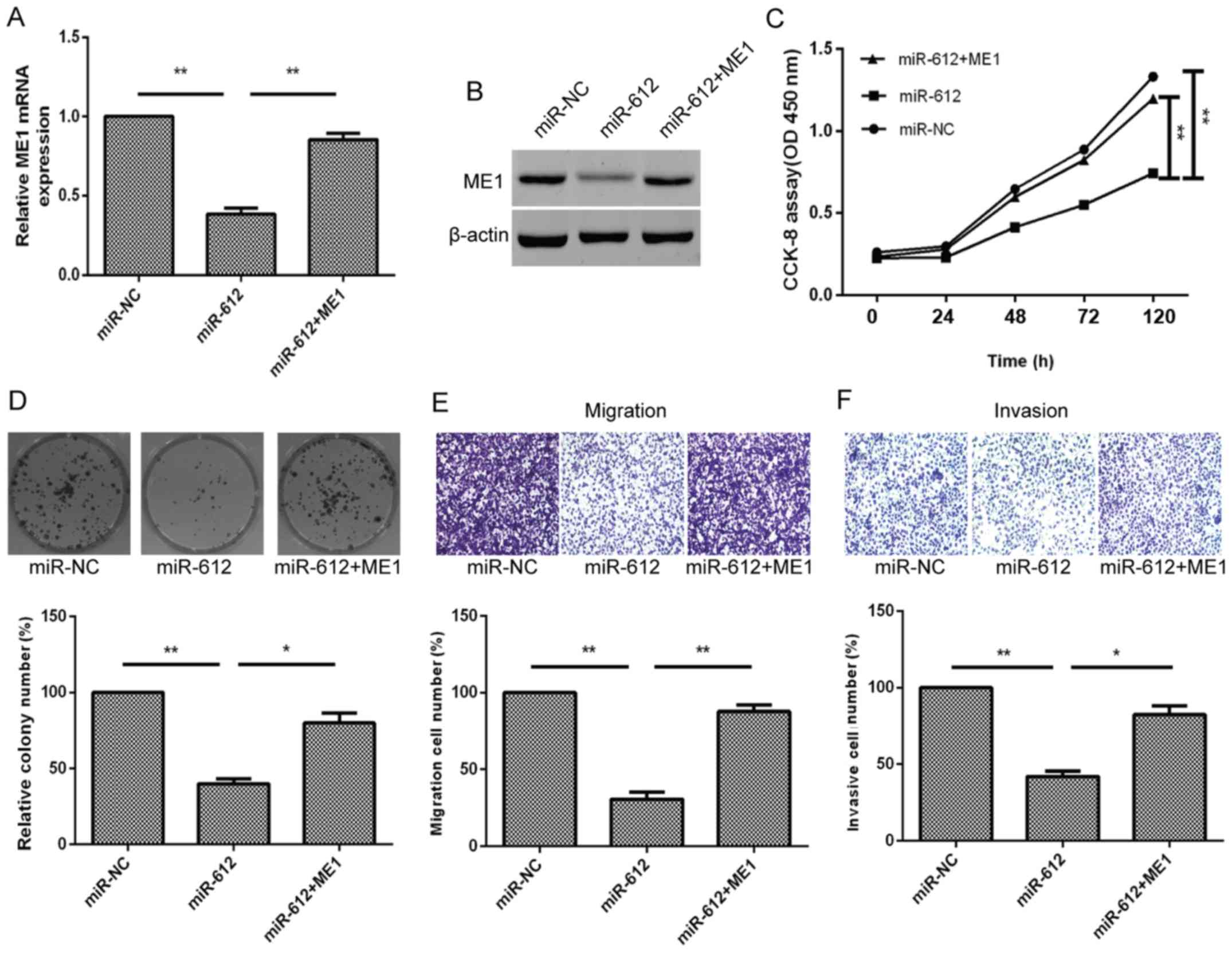
Furthermore, to better verify that ME1 was a direct
target of miR-612, the T24 cells were simultaneously co-transfected
with the miR-612 mimic or miR-NC and ME1 overexpression plasmid. We
found that ME1 overexpression using ME1 cDNA transfection in
miR-612-expressing T24 cells was able to partially reverse miR-612
overexpression-induced reduction of ME1 expression in T24 cells
(Fig. 6A and B). The ME1
overexpression also functionally rescued the inhibitory effect of
miR-612 overexpression on tumor cell growth, colony formation,
migration and invasion (Fig.
6C–F). Taken together, the results indicated that miR-612
inhibited tumor cell growth, colony formation, migration, invasion
and EMT by targeting ME1 in vitro (Fig. 7).
Discussion
Accumulating evidence has demonstrated that aberrant
miRNA expression contributes to the development and progression of
human cancer, including bladder cancer (30-34).
The present study further confirmed this notion for miR-612 in
bladder cancer tissues and cell lines. It was observed that miR-612
expression was downregulated in bladder cancer tissues and cell
lines as compared with that in normal tissue and cells. The data
also revealed that loss of miR-162 expression was associated with
advanced bladder cancer TNM stages and metastasis. Additionally, it
was demonstrated that miR-612 suppressed bladder cancer cell
proliferation in vitro and in vivo by targeting ME1
expression. These results suggest that miR-612 may be a tumor
suppressor gene or possess tumor-suppressing functions in bladder
cancer. Further studies will investigate whether miR-612 serves as
a potential tumor biomarker or therapeutic target for bladder
cancer.
Previous studies have reported that altered miR-612
expression is involved in the development and progression of many
types of human cancer, including hepatocellular carcinoma and
colorectal cancer. For instance, miR-612 expression was able to
inhibit liver cancer stemness and target AKT2 expression via the
Wnt/β-catenin signaling pathway (19). In addition, miR-612 expression
suppressed colorectal cancer growth and metastasis by targeting
AKT2 expression (20), while
miR-612 was shown to serve an inhibitory effect on hepatocellular
carcinoma cell stemness through modulation of the Sp1/Nanog
signaling pathway (18). These
previous findings suggest that miR-612 functions as a putative
tumor suppressor in the aforementioned types of cancer.
Furthermore, the present study demonstrated low expression of
miR-612 in bladder cancer tissues and cell lines; however, the
target gene ME1 was highly expressed in bladder cancer tissues.
This indicated that, while miR-612 serves as a tumor suppressor in
bladder cancer, the target gene ME1 was cancer-dependent. Further
studies should investigate the association between miR-612 and the
target genes possessing miR-612 tumor-suppressing functions in
human cancer.
In the present study, the effects of miR-612 in
bladder cancer were found to occur via the targeting of ME1
expression, according to the following observations: i) High
expression of ME1 mRNA was inversely associated with miR-612
expression in bladder cancer tissues; ii) miR-612 was able to bind
to the 3′-UTR of ME1 cDNA, but not to the mutated ME1 molecule;
iii) miR-612 mimic transfection was able to downregulate ME1
expression; iv) ME1 knockdown significantly inhibited tumor cell
growth, colony formation, and migration and invasion capacities,
which mimicked the effects of miR-612 overexpression on T24 cells;
and v) ME1 overexpression in miR-612-expressing T24 cells partially
reversed miR-612 overexpression-reduced ME1 expression and
functionally rescued the inhibitory effect of miR-612
overexpression on tumor cell growth, colony formation, migration
and invasion.
ME1 is a cytosolic NADP+-dependent enzyme
this is critically responsible for NADPH production, fatty acid
biosynthesis, and lipogenesis in cells and tissues (35). ME1 expression is upregulated in
various types of human cancer, such as in colorectal cancer, and is
associated with a poor overall survival of patients (36,37).
It was also reported that ME1 expression induces EMT of
hepatocellular carcinoma cells and is associated with a poor
prognosis of these patients (25).
In addition, knockdown of ME1 expression has been demonstrated to
inhibit the nasopharyngeal carcinoma cell migration and invasion
capacity. Another study reported that mutant KRAS was associated
with ME1 overexpression and predicted the response of non-small
cell lung cancer cells to radiation therapy (38). Furthermore, the p53 protein
reciprocally regulated ME1 expression to modulate cell metabolism
and senescence (35). To the best
of our knowledge, the present study demonstrated for the first time
that the levels of ME1 mRNA were upregulated in bladder cancer
tissues, but inversely associated with miR-612 expression.
Meanwhile, the in vivo data revealed that miR-612 suppressed
the growth of bladder cancer cell-induced xenografts in mice by
repression of ME1 expression.
In conclusion, the current results provided evidence
indicating that miR-612 expression is downregulated in bladder
cancer tissues and cell lines, and that decreased miR-612
expression is associated with advanced bladder cancer TNM stages
and distant metastasis. The in vitro and in vivo data
further suggested that miR-612 expression was able to inhibit
bladder cancer cell growth, colony formation, migration, invasion
and EMT through directly targeting the expression of ME1. These
findings indicate that miR-612 may serve as a novel therapeutic
target for bladder cancer.
Acknowledgments
The authors would like to thank the personnel of the
Central Laboratory of the Shanghai Tenth People's Hospital for
their assistance and support.
Funding
This study was supported in part by a grant from the
National Natural Science Foundation of China (no. 81272836).
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
ML and XY made substantial contributions to the
conception and design of the study; ML and YC drafted the
manuscirpt; ML made substantial contributions to the acquisition,
analysis, and interpretation of the data for the study; YC made
substantial contributions to the interpretation of the data for the
study; BH contributed to the design of the study; SM, KC, LW and XY
contributed to the acquisition and analysis of the data for the
study; BH, SM, KC, LW and XY revised the manuscript critically for
important intellectual content. All authors gave the final approval
of the manuscript to be published and agree to be accountable for
all aspects of the study in ensuring that questions related to the
accuracy or integrity of any part of the study are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The patient study was approved by the Ethics
Committee of Shanghai Tenth People's Hospital of Tongji University.
Informed consent was obtained from all patients or their relatives.
The animal experiments were approved by the Animal Care and Use
Committee of Tongji University.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests
References
|
1
|
McGuire S: World Cancer Report 2014.
Geneva, Switzerland: World Health Organization, International
Agency for Research on Cancer, WHO Press; 2015, Adv Nutr. 7. pp.
418–419. 2016, View Article : Google Scholar
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zeng LP, Hu ZM, Li K and Xia K: miR-222
attenuates cisplatin-induced cell death by targeting the
PPP2R2A/Akt/mTOR Axis in bladder cancer cells. J Cell Mol Med.
20:559–567. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang X, Cheng Y, Li P, Tao J, Deng X,
Zhang X, Gu M, Lu Q and Yin C: A lentiviral sponge for miRNA-21
diminishes aerobic glycolysis in bladder cancer T24 cells via the
PTEN/PI3K/AKT/mTOR axis. Tumour Biol. 36:383–391. 2015. View Article : Google Scholar
|
|
6
|
Hanke M, Hoefig K, Merz H, Feller AC,
Kausch I, Jocham D, Warnecke JM and Sczakiel G: A robust
methodology to study urine microRNA as tumor marker: microRNA-126
and microRNA-182 are related to urinary bladder cancer. Urol Oncol.
28:655–661. 2010. View Article : Google Scholar
|
|
7
|
Bushati N and Cohen SM: microRNA
functions. Annu Rev Cell Dev Biol. 23:175–205. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer.
96(Suppl): R40–R44. 2007.PubMed/NCBI
|
|
9
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vinall RL, Ripoll AZ, Wang S, Pan CX and
deVere White RW: miR-34a chemosensitizes bladder cancer cells to
cisplatin treatment regardless of p53-Rb pathway status. Int J
Cancer. 130:2526–2538. 2012. View Article : Google Scholar
|
|
11
|
Shen J, Zhang J, Xiao M, Yang J and Zhang
N: miR-203 suppresses bladder cancer cell growth and targets the
Twist1. Oncol Res. Sep 6–2017.(Epub ahead of print). doi:
10.3727/096504017X15041934685237.print) 2017. View Article : Google Scholar
|
|
12
|
Pop-Bica C, Gulei D, Cojocneanu-Petric R,
Braicu C, Petrut B and Berindan-Neagoe I: Understanding the role of
non-coding RNAs in bladder cancer: From dark matter to valuable
therapeutic targets. Int J Mol Sci. 18:182017. View Article : Google Scholar
|
|
13
|
Wei Z, Hu X, Liu J, Zhu W, Zhan X and Sun
S: MicroRNA-497 upregulation inhibits cell invasion and metastasis
in T24 and BIU-87 bladder cancer cells. Mol Med Rep. 16:2055–2060.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu D, Niu X, Tao J, Li P, Lu Q, Xu A, Chen
W and Wang Z: MicroRNA-379-5p plays a tumor-suppressive role in
human bladder cancer growth and metastasis by directly targeting
MDM2. Oncol Rep. 37:3502–3508. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Y, Zhang Z, Li Z, Gong D, Zhan B,
Man X and Kong C: MicroRNA-497 inhibits the proliferation,
migration and invasion of human bladder transitional cell carcinoma
cells by targeting E2F3. Oncol Rep. 36:1293–1300. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu H, Duan P, Zhu H and Rao D: miR-613
inhibits bladder cancer proliferation and migration through
targeting SphK1. Am J Transl Res. 9:1213–1221. 2017.PubMed/NCBI
|
|
17
|
Wu CL, Ho JY, Chou SC and Yu DS: miR-429
reverses epithelial-mesenchymal transition by restoring E-cadherin
expression in bladder cancer. Oncotarget. 7:26593–26603.
2016.PubMed/NCBI
|
|
18
|
Liu Y, Liu DL, Dong LL, Wen D, Shi DM,
Zhou J, Fan J and Wu WZ: miR-612 suppresses stem cell-like property
of hepatocellular carcinoma cells by modulating Sp1/Nanog
signaling. Cell Death Dis. 7:e23772016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tang J, Tao ZH, Wen D, Wan JL, Liu DL,
Zhang S, Cui JF, Sun HC, Wang L, Zhou J, et al: miR-612 suppresses
the stemness of liver cancer via Wnt/β-catenin signaling. Biochem
Biophys Res Commun. 447:210–215. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sheng L, He P, Yang X, Zhou M and Feng Q:
miR-612 negatively regulates colorectal cancer growth and
metastasis by targeting AKT2. Cell Death Dis. 6:e18082015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Umetsu SE, Shafizadeh N and Kakar S:
Grading and staging mucinous neoplasms of the appendix: A case
series and review of the literature. Hum Pathol. 69:81–89. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dweep H, Sticht C, Pandey P and Gretz N:
miRWalk–database: Prediction of possible miRNA binding sites by
'walking' the genes of three genomes. J Biomed Inform. 44:839–847.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dweep H and Gretz N: miRWalk2.0: A
comprehensive atlas of microRNA-target interactions. Nat Methods.
12:6972015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hu H, Chen Y, Cheng S, Li G and Zhang Z:
Dysregulated expression of homebox gene HOXA13 is correlated with
the poor prognosis in bladder cancer. Wien Klin Wochenschr.
129:391–397. 2017. View Article : Google Scholar
|
|
25
|
Schenková K, Lutz J, Kopp M, Ramos S and
Rivero F: MUF1/leucine-rich repeat containing 41 (LRRC41), a
substrate of RhoBTB-dependent cullin 3 ubiquitin ligase complexes,
is a predominantly nuclear dimeric protein. J Mol Biol.
422:659–673. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zheng H, Li Q, Chen R, Zhang J, Ran Y, He
X, Li S and Shu HB: The dual-specificity phosphatase DUSP14
negatively regulates tumor necrosis factor- and
interleukin-1-induced nuclear factor-κB activation by
dephosphorylating the protein kinase TAK1. J Biol Chem.
288:819–825. 2013. View Article : Google Scholar
|
|
27
|
Mukhtar YM, Huang Y, Liu J, Chen D and
Zheng W: Acetanilide and bromoacetyl-lysine derivatives as
activators for human histone deacetylase 8. Bioorg Med Chem Lett.
27:2319–2323. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sengupta S, Weeraratne SD, Sun H, Phallen
J, Rallapalli SK, Teider N, Kosaras B, Amani V, Pierre-Francois J,
Tang Y, et al: α5-GABAA receptors negatively regulate MYC-amplified
medulloblastoma growth. Acta Neuropathol. 127:593–603. 2014.
View Article : Google Scholar
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
30
|
Braicu C, Cojocneanu-Petric R, Chira S,
Truta A, Floares A, Petrut B, Achimas-Cadariu P and Berindan-Neagoe
I: Clinical and pathological implications of miRNA in bladder
cancer. Int J Nanomedicine. 10:791–800. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yoshino H, Seki N, Itesako T, Chiyomaru T,
Nakagawa M and Enokida H: Aberrant expression of microRNAs in
bladder cancer. Nat Rev Urol. 10:396–404. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hu Y, Cheng C, Hong Z and Shi Z:
Independent prognostic miRNAs for bladder urothelial carcinoma.
Oncol Lett. 14:3001–3005. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu X, Liu X, Wu Y, Wu Q, Wang Q, Yang Z
and Li L: MicroRNAs in biofluids are novel tools for bladder cancer
screening. Oncotarget. 8:32370–32379. 2017.PubMed/NCBI
|
|
34
|
Wang X, Liang Z, Xu X, Li J, Zhu Y, Meng
S, Li S, Wang S, Xie B, Ji A, et al: miR-148a-3p represses
proliferation and EMT by establishing regulatory circuits between
ERBB3/AKT2/c-myc and DNMT1 in bladder cancer. Cell Death Dis.
7:e25032016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jiang P, Du W, Mancuso A, Wellen KE and
Yang X: Reciprocal regulation of p53 and malic enzymes modulates
metabolism and senescence. Nature. 493:689–693. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wen D, Liu D, Tang J, Dong L, Liu Y, Tao
Z, Wan J, Gao D, Wang L, Sun H, et al: Malic enzyme 1 induces
epithelial-mesenchymal transition and indicates poor prognosis in
hepatocellular carcinoma. Tumour Biol. 36:6211–6221. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shen H, Xing C, Cui K and Li Y, Zhang J,
Du R, Zhang X and Li Y: MicroRNA-30a attenuates mutant KRAS-driven
colorectal tumorigenesis via direct suppression of ME1. Cell Death
Differ. 24:1253–1262. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chakrabarti G: Mutant KRAS associated
malic enzyme 1 expression is a predictive marker for radiation
therapy response in non-small cell lung cancer. Radiat Oncol.
10:1452015. View Article : Google Scholar : PubMed/NCBI
|