Introduction
Cushing's disease (CD), which is characterized by
excessive secretion of the adrenocorticotrophic hormone (ACTH),
usually results from a pituitary adenoma or corticotroph
hyperplasia (1). High levels of
ACTH stimulate the adrenal glands to release excess
glucocorticoids, resulting in hypercortisolism, which can induce a
series of long-term complications, including cardiovascular
disease, metabolic syndrome, muscle atrophy, osteoporosis,
recurrent infections and psychiatric disorders that can cause
substantial morbidity and mortality if not appropriately treated
(2,3). To date, pituitary surgery remains the
first line therapy, while pituitary radiotherapy or bilateral
adrenalectomy have traditionally been used as adjuvant therapies
for persistent hypercortisolism (4). However, even with combined therapy,
the remission rate is low and the risk of recurrence is high
(5). Thus, the development of
novel therapeutic approaches for CD is required, which will depend
closely on an improved understanding of the genetic pathogenesis of
the disease.
To detect the intrinsic molecular defects of CD, RNA
sequencing (RNA-Seq) was used to determine the gene expression
profile of CD in the current study. Using RNA-Seq, the
transcriptome of 13 cases of CD and 5 normal human pituitaries
(NHPs) were analyzed. Marked downregulation of the gene secreted
frizzled-related protein 2 (SFRP2) was identified in CD. SFRP2
belongs to the SFRP family and acts as a negative regulator of the
canonical Wnt signaling pathway. It can competitively bind frizzled
receptors that accelerate β-catenin degradation and reduce the
nuclear translocation of β-catenin (6). Thus, decreased expression of SFRP2
results in upregulation of Wnt/β-catenin signaling, which regulates
diverse developmental processes, such as differentiation, cell
migration, cell adhesion and proliferation (7). In fact, it has been reported that
decreased expression of SFRP2 activates Wnt/β-catenin signaling,
which promotes the development of various tumors types, including
colorectal cancer (8), esophageal
carcinoma (9), medulloblastoma
(10), hepatocellular carcinoma
(11) and aggressive
nonfunctioning pituitary adenoma (12).
The findings of the current study confirmed that
SFRP2 expression is downregulated in human pituitary corticotroph
tumors compared with normal pituitary tissue. In clinical analysis,
a correlation between lower SFRP2 expression and larger tumor size
and invasiveness was identified. Furthermore, it was demonstrated
that SFRP2 is closely associated to corticotroph cell proliferation
and invasiveness through downregulation of Wnt signaling in
vitro. These results suggest that SFRP2 is a novel molecular
marker underlying tumorigenesis of pituitary corticotroph adenoma,
which may provide a rationale for SFRP2 as a potential therapeutic
target for CD.
Materials and methods
Patients and human tissue samples
NHP glands (n=8; 3 males, 5 females; 22–67 years
old) were obtained from fresh autopsy specimens between February,
2013 and October, 2015 at the Shanghai Jiaotong University School
of Medicine (Shanghai, China). The 8 samples were rinsed in sterile
saline and snap-frozen in liquid nitrogen. Suitable parts of each
normal pituitary tissue were embedded in paraffin. All 36 human
ACTH-secreting pituitary adenoma samples (30 females, 6 males;
24–59 years old) were obtained through endonasal transsphenoidal
surgery between January, 2013 and September, 2015 at the Department
of Neurosurgery of Rui-Jin Hospital, Shanghai Jiaotong University
(Shanghai, China). Tumors were immediately 'flash-frozen' in liquid
nitrogen until use. Suitable parts of each sample were embedded in
paraffin. Clinical diagnoses were confirmed via endocrine
assessment, clinical features, magnetic resonance imaging,
histology and pituitary hormone immunohistochemistry. Clinical data
were obtained from Rui-Jin Hospital (Shanghai, China), including
plasma ACTH, urinary free cortisol and midnight serum cortisol.
Informed consent was obtained from all participating patients,
which was approved by the Ruijin Hospital Ethics Committee
(Shanghai, China).
RNA extraction and purification
Total RNA was extracted from tissues and cells using
TRIzol reagent (Thermo Fisher Scientific, Inc., Waltham, MA, USA)
according to the manufacturer's instructions. The RNA integrity
number was verified to inspect RNA integrity with an Agilent
Bioanalyzer 2100 (Agilent Technologies, Inc., Santa Clara, CA,
USA). The RNeasy Micro kit (cat. no. 74004) and the RNase-Free
DNase Set (cat. no. 79254) (both from Qiagen GmbH, Hilden, Germany)
were used to further purify total RNA.
RNA-Seq expression profiling
Preparation of the RNA libraries was performed
according to the manufacturer's recommendations. All 18 libraries
(5 NHP, 13 CD) were constructed using the TruSeq RNA Sample Prep
Kit V2 (RS-122-2001; Illumina, Inc., San Diego, CA, USA). All
sequencing was performed with the Illumina HiSeq 2500. HiSeq
Control Software 2.0.12 (Illumina, Inc.) was used to analyze array
images and extract the raw data. FASTX-Toolkit (version 0.0.13;
hannonlab.cshl.edu/fastx_toolkit/) was used to filter
the raw reads to acquire clean reads. TopHat (version 2.0.9;
ccb.jhu.edu/software/tophat/downloads/) was employed
to map the reads. Subsequently, cufflinks (version 2.1.1;
ccb.jhu.edu/software/tophat/index.shtml) was used to
quantify the mapping results. Differentially expressed genes (DEGs)
were then identified through fold change and P-values calculated
with a t-test. The threshold set for up and downregulated genes was
fold change ≥2.0 and P≤0.05.
Functional enrichment analysis
Functional enrichment analysis was applied to
investigate the potential functions of the DEGs. Kyoto Encyclopedia
of Genes and Genomes (KEGG) pathway analysis (www.kegg.jp) was performed to reveal the physiological
function of the DEGs in CD, and Fisher's exact test was applied to
select significant pathways. The threshold of significance was
defined according to the P-value and false discovery rate.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Following extraction of total RNA, 1 µg total
RNA was converted into first-strand cDNA using an RT kit (Promega
Corporation, Madison, WI, USA), with incubation at 42°C for 15 min,
95°C for 5 min and then 4°C for 5 min. The cDNA was the used as a
template for qPCR using SYBR Green Supermix (Takara Bio, Inc.,
Otsu, Japan), and performed with the LightCycler 480 system (Roche
Diagnostics, Basel, Switzerland) subjected to 40 cycles of
amplification (95°C for 5 sec and 60°C for 1 min). The relative RNA
levels were determined with the 2−ΔΔCq method (13) and normalized to the mRNA level of
β-actin. Quantification was performed in quadruplicate, and the
experiments were repeated in triplicate. The sequences of the
primers are listed in Table I.
 | Table ISequences of the primers for reverse
transcription-quantitative polymerase chain reaction. |
Table I
Sequences of the primers for reverse
transcription-quantitative polymerase chain reaction.
| Species Gene
name | Primer sequences
(5′–3′) |
|---|
| Human SFRP2 | F:
ACGGCATCGAATACCAGAACA |
| R:
CTCGTCTAGGTCATCGAGGCA |
| Human GADD45B | F:
CAGATCCATTTTACGCTGATCCA |
| R:
TCCTCGCAAAACAGGCTGAG |
| Human JUNB | F:
ACAAACTCCTGAAACCGAGCC |
| R:
CGAGCCCTGACCAGAAAAGTA |
| Human FOSL2 | F:
CAGAAATTCCGGGTAGATATGCC |
| R:
GGTATGGGTTGGACATGGAGG |
| Human TFPI2 | F:
TCCTGCCCCTAGACTACGG |
| R:
CTCCCAGGTGTAGAAATTGTTGG |
| Human ACTB | F:
CATGTACGTTGCTATCCAGGC |
| R:
CTCCTTAATGTCACGCACGAT |
| Human TBX19 | F:
ACGGACCAATCAATCCAGGG |
| R:
TACACCCCATCCTGGTCTCA |
| Human CYR61 | F:
GCATTCCTCTGTGTCCCCAA |
| R:
TTAGTGTCCATCCGCACCAG |
| Mouse Sfrp2 | F:
CGTGGGCTCTTCCTCTTCG |
| R:
ATGTTCTGGTACTCGATGCCG |
| Mouse Pomc | F:
ATGCCGAGATTCTGCTACAGT |
| R:
TCCAGCGAGAGGTCGAGTTT |
| Mouse c-myc | F:
ATGCCCCTCAACGTGAACTTC |
| R:
GTCGCAGATGAAATAGGGCTG |
| Mouse Pttg1 | F:
TCTGATCCGCTGTACTCTCCT |
| R:
AGGCGGCAATTCAACATCCA |
| Mouse MMP2 | F:
CTGCCACTGTCCCAGGAAG |
| R:
CTCGCGGCAAGTCTTCAGAG |
| Mouse MMP7 | F:
CTTACCTCGGATCGTAGTGGA |
| R:
CCCCAACTAACCCTCTTGAAGT |
| Mouse MMP9 | F:
GCAGAGGCATACTTGTACCG |
| R:
TGATGTTATGATGGTCCCACTTG |
| Mouse Ctnnb1 | F:
ATGGAGCCGGACAGAAAAGC |
| R:
CTTGCCACTCAGGGAAGGA |
| Mouse Actb | F:
GGCTGTATTCCCCTCCATCG |
| R:
CCAGTTGGTAACAATGCCATGT |
Protein extraction and western blot
analysis
NHP (n=3) and CD (n=23) fresh-frozen tumor tissues
were used for western blotting. Total protein of each tissue was
extracted using in radioimmunoprecipitation assay (RIPA) buffer
containing Protease Inhibitor Cocktail (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) for 15 min at 0°C, and then supernatant was
collected. For cells, total cell lysate was also prepared by
incubating in RIPA buffer containing protease inhibitor cocktail on
ice. Protein concentrations were measured using a Bio-Rad protein
assay reagent (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Nuclear and cytoplasmic protein extracts were prepared using the
Nuclear Extract Kit (Active Motif, Carlsbad, CA, USA) according to
the manufacturer's instructions. Equal amounts of protein (20
µg per sample) were boiled for 5 min and resolved by
SDS-PAGE on 8 or 10% gels. Following transfer, membranes were
blocked for 1 h at room temperature in TBS-Tween-20 containing 5%
non-fat dried milk. The membranes were incubated with the following
primary antibodies: Anti-SFRP2 (dilution 1:1,000; cat. no.
ab86379), anti-β-catenin (dilution, 1:5,000; cat. no. ab32572; all
from Abcam, Cambridge, UK); anti-α-tubulin (dilution 1:1,000; cat.
no. T8328; Sigma-Aldrich; Merck KGaA); anti-cyclin E (dilution
1:1,000; cat. no. 4129), anti-proliferating cell nuclear antigen
(PCNA; dilution 1:2,000; cat. no. 2586), anti-phospho-extracellular
signal-regulated kinase (p-Erk; dilution 1:2,000; cat. no. 9101),
anti-Erk (dilution 1:1,000; cat. no. 9102), anti-c-myc (dilution
1:1,000; cat. no. 9402; all from Cell Signaling Technology, Inc.
Danvers, MA, USA); anti-β-actin (dilution 1:10,000; cat. no.
60008-1-Ig), anti-lamin (dilution 1:5,000; cat. no. 66095-1-Ig),
anti-GAPDH (dilution 1:10,000; cat. no. 10494-1-AP; all from
ProteinTech Group, Inc., Chicago, IL, USA). Following washing,
membranes were incubated with horseradish peroxidase-conjugated
secondary antibodies (dilution 1:4,000; cat. nos. 7076 and 7074;
Cell Signaling Technology, Inc.) for 1 h at room temperature. The
signal was detected using enhanced chemiluminescence (NEL104001EA;
PerkinElmer, Inc., Waltham, MA, USA) on the 5200 Chemiluminescence
Imager (Tanon Science and Technology Co., Ltd., Shanghai, China).
The densitometry analysis was performed using ImageJ software
(version 1.47v; National Institutes of Health, Bethesda, MD,
USA).
Immunohistochemistry staining (IHC)
The 5 µm thick sections were prepared from
the formalin-fixed and paraffin embedded tissues (3 NHP and 23 CD).
IHC staining was performed as previously reported (12). Specific primary antibody against
SFRP2 (dilution 1:200; cat. no. ab86379; Abcam) was used. All the
IHC slides were evaluated by two pathologists (Department of
Pathology, Rui-Jin Hospital, Shanghai, China). The staining
intensity was graded as follows: No staining, 0; weakly positive,
1; moderately positive, 2; and strongly positive, 3. The staining
percentage was graded as follows: 0–25% staining, 1; 26–50%
staining, 2; 51–75% staining, 3; and 76–100% staining, 4. The
immunoreactive score was calculated as intensity of the staining
multiplied by the percentage of positive cells, which was then
categorized as low (0–6) and high (7–12)
expression.
Cell culture and transfection
The AtT20 cell line and 293T cells were obtained
from the American Type Tissue Collection (Manassas, VA, USA). The
cells were cultured in Dulbecco's modified Eagle's medium
containing 10% fetal bovine serum (FBS) and 2 mM L-glutamine and
100 µg/ml penicillin/streptomycin (all from Gibco; Thermo
Fisher Scientific, Inc.) in a humidified incubator under 5%
CO2 conditions at 37°C. The murine SFRP2 coding sequence
was cloned into the retroviral pMSCVpuro vector (Clontech
Laboratories, Inc., Mountainview, CA, USA) to generate SFRP2
expression plasmids. The retroviral vector pMSCVpuro vector was
used as the negative control. 293T cells (2.5×105 in 2
ml media per well) were seeded in a 6-well plate. After 24 h, the
recombinant construct was co-transfected into 293T cells together
with two packaging vectors psPAX2 and pMD2.G (plasmid cat. nos.
12260 and 12259; obtained from Addgene, Inc., Cambridge, MA, USA)
in a ratio of 3:2.25:0.75 µg of pMSCVpuro, psPAX2 and
pMD2.G, respectively, per well. Lipofectamine 2000 transfection
reagent (Thermo Fisher Scientific, Inc.) was used according to the
manufacturer's instructions. The supernatant was collected at 48 h
post-transfection, and filtered through 0.45-µm pore size
filter. Viral titers were determined by QuickTiter Quantitation Kit
(cat. no. VPK-120; Cell Biolabs, Inc., San Diego, CA, USA)
following manufacturer's instructions. AtT20 cells were infected
with pMSCVpuro-SFRP2 vector and pMSCVpuro vector at the
multiplicity of infection of 10. Stably overexpressed SFRP2 AtT20
and control cells were established following puromycin selection
(100 ng/ml) for 14 days. To determine whether recombinant SFRP2
protein can effect ACTH secretion, proliferation and clonogenic
ability of AtT20 cells, the cells were treated with recombinant
mouse SFRP2 protein (cat. no. 1169-FR-025; R&D Systems, Inc.,
Minneapolis, MN, USA) at different doses (0–100 nM) and time points
(24–72 h).
Hormone assays
The culture media was collected from each well,
which was incubated for a specified time interval, and the level of
ACTH was measured using the ACTH ELISA kit (cat. no. EK-001-21;
Phoenix Pharmaceuticals, Inc., Burlingame, CA, USA). Each
experimental group was analyzed five times and the experiments were
repeated independently three times.
Cell proliferation and colony formation
assay
Cell viability was tested using the Cell Counting
Kit-8 assay (Dojindo Molecular Technologies, Inc., Kumamoto, Japan)
at the indicated time points. In the colony formation test, AtT20
cells were plated in a 6-well plate at a density of 103
cells/well. After 3 weeks, colonies were fixed, stained and counted
as previously described (14).
Flow cytometry
The cells were digested with 0.5% trypsin-EDTA,
transferred into flow tubes and centrifuged at 800 × g for 5 min.
Then cells were washed twice with cold PBS and resuspended cells in
binding buffer, and incubated in the dark at room temperature for 5
min with 5 µl Annexin V-fluorescein isothiocyanate (FITC)
and 10 µl propidium iodide prior to detection (FITC Annexin
V Apoptosis Detection kit I; BD Pharmingen, San Jose, CA, USA). The
apoptotic results were analyzed using FlowJo software (Tree Star,
Inc., Ashland, OR, USA) The results of the flow cytometry
experiments were displayed as four quadrants, in which the upper
right quadrant indicated advanced apoptosis rate, the lower right
quadrant represents the apoptosis rate during the early stages, the
upper left quadrant represents dead cells, and the lower left
quadrant represents living cells. This experiment was repeated six
times.
Scratch assay
Cells were cultured to 90% confluence in 6-well
plates. A small scratching area was then removed with a 100
µl plastic pipette tip, following which the cells were
cultured in low serum medium (containing 2% FBS). Cells were
inspected microscopically over 30 h and ImageJ software (version
1.47v; National Institutes of Health, Bethesda, MD, USA) was
applied to measure the relative scratch width.
Statistical analysis
Statistical analysis was performed using SPSS
version 19.0 software (IBM Corporation, Armonk, NY, USA). All
experimental data were presented as the mean ± standard deviation.
The correlation of expression of the DEGs between CD and NHP
samples, and the correlation between SFRP2 expression and hormone
parameters were analyzed using Spearman's correlation. Association
between clinicopathological parameters and SFRP2 expression were
evaluated by χ2 tests. When sample numbers in some
categorical cells were less than five, Fisher's exact test was
employed. Unpaired Student's t-test or the Mann-Whitney U test was
applied to analyze significant differences. One-way analysis of
variance was used to compare the differences among subgroups with
post hoc contrasts by Student-Newman-Keuls tests. P<0.05 was
considered to indicate a statistically significant difference.
Results
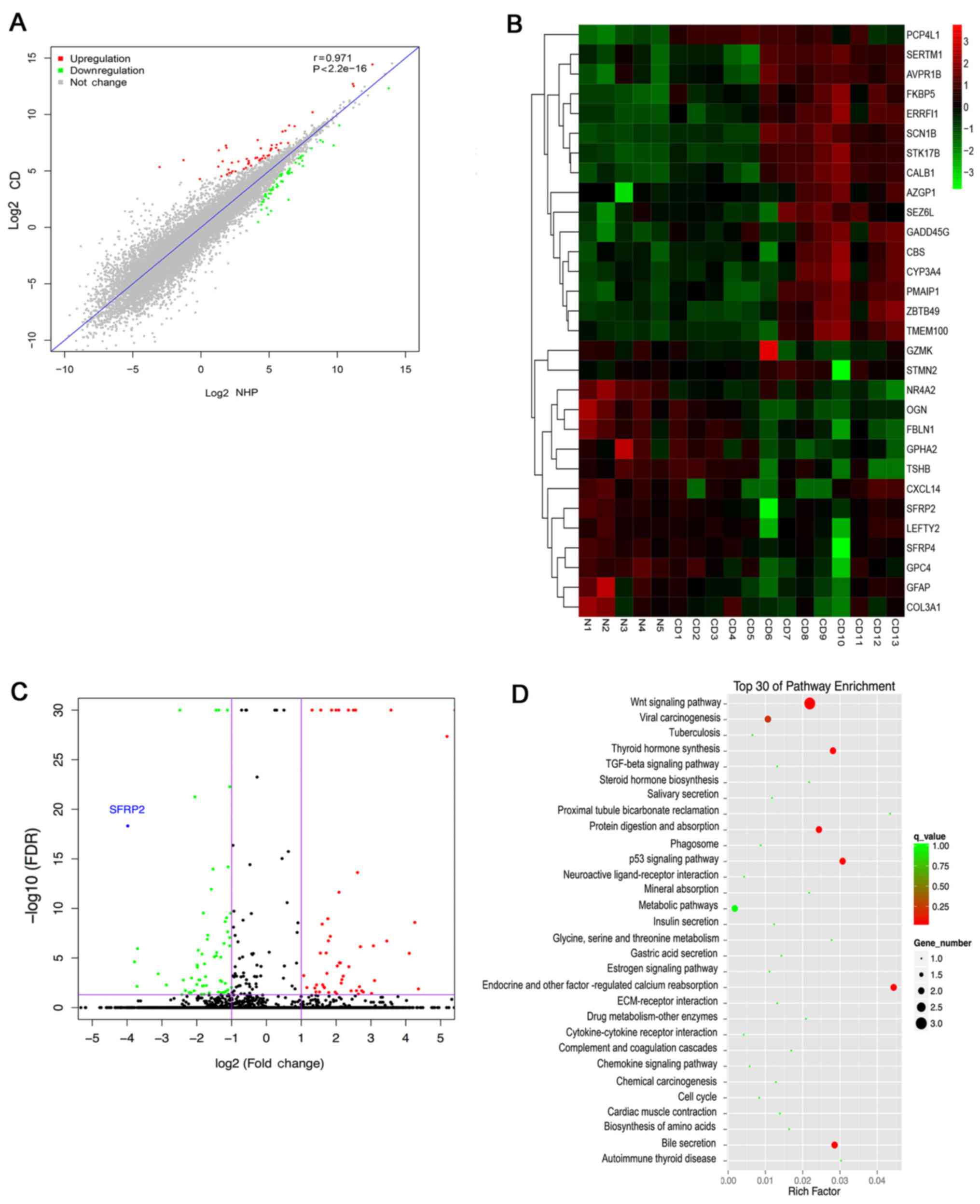
RNA-Seq reveals decreased expression of
SFRP2 in human pituitary corticotroph adenoma
To determine the differences in mRNA expression
levels between CD and NHP samples, the transcriptomes of 13 CD and
5 NHP samples were analyzed using RNA-Seq. Stringent statistical
filters with P<0.05 and fold change >2 were applied. The
correlation analysis revealed that the expression of genes between
CD and NHP samples were highly correlated, with a correlation
coefficient r of 0.97 (Fig. 1A).
Based on the RNA-Seq data, 122 differentially expressed genes were
identified in tumor tissues; 57 genes were upregulated and 65 genes
were downregulated in CD samples compared with the NHP samples. The
30 most significant DEGs are listed in Table II. α-2-glycoprotein 1 zinc-binding
(fold change 331.2874) exhibited the greatest upregulation, while
SFRP2 (fold change 15.9491) exhibited the greatest downregulation.
Hierarchical clustering analysis was also used to show the mRNA
expression levels of all specimens (Fig. 1B). As shown in the heat map and
volcano plot (Fig. 1C), SFRP2 was
the most downregulated gene of all the identified DEGs.
Furthermore, the top 30 pathways associated with these genes were
determined using KEGG pathway enrichment analysis. This analysis
indicted that the greatest number of genes were classified as
associated with the Wnt signaling pathway, which was significantly
enriched (Fig. 1D). These results
suggest that the decreased expression of SFRP2 may result in
aberrant Wnt signaling in ACTH-secreting pituitary adenomas.
| Table IIAberrantly expressed mRNAs and six
random verified genes in microarray for Cushing's disease
pituitaries and normal human pituitaries. |
Table II
Aberrantly expressed mRNAs and six
random verified genes in microarray for Cushing's disease
pituitaries and normal human pituitaries.
| Gene name | GenBank accession
no. | Fold change
(absorbance) | P-value | Regulation |
|---|
| AZGP1 | NM_001185.3 | 331.2874 | 1.38e-10 | Up |
| CYP3A4 | NM_017460.5 | 150.9702 | 2.24e-16 | Up |
| PMAIP1 | NM_021127.2 | 45.64176 | 2.08e-27 | Up |
| CALB1 | NM_004929.3 | 36.33607 | 3.56e-31 | Up |
| GZMK | NM_002104.2 | 20.56787 | 6.93e-05 | Up |
| PCP4L1 | NM_001102566.1 | 17.1511 | 8.66e-09 | Up |
| ERRFI1 | NM_018948.3 | 11.91691 | 3.42e-38 | Up |
| STK17B | XM_011512169.1 | 10.96635 | 4.21e-10 | Up |
| AVPR1B | NM_000707.3 | 8.601595 | 8.06e-06 | Up |
| SERTM1 | NM_203451.2 | 8.40439 | 1.37e-09 | Up |
| TMEM100 | NM_001099640.1 | 8.094779 | 0.0002328 | Up |
| SCN1B | NM_199037.4 | 6.870989 | 0.000160445 | Up |
| CBS | XM_017028490.1 | 6.652037 | 0.000109376 | Up |
| GADD45G | NM_006705.3 | 6.484037 | 1.70e-09 | Up |
| SEZ6L | NM_021115.4 | 6.369464 | 3.95e-05 | Up |
| ZBTB49 | NM_001330625.1 | 6.233656 | 0.00026919 | Up |
| FKBP5 | NM_001145775.2 | 6.123551 | 2.88e-17 | Up |
| STMN2 | NM_001199214.1 | 6.000352 | 0.000109376 | Up |
| SFRP2 | NM_003013.2 | 15.77754 | 4.56e-22 | Down |
| SFRP4 | NM_003014.3 | 13.79746 | 2.44e-05 | Down |
| GPHA2 | XM_011544776.2 | 13.12417 | 3.61e-05 | Down |
| GFAP | NM_002055.4 | 13.00376 | 2.65e-09 | Down |
| LEFTY2 | NM_001172425.2 | 8.61589 | 1.38e-06 | Down |
| OGN | NM_024416.4 | 7.314456 | 2.44e-05 | Down |
| TSHB | XM_011542065.2 | 5.587824 | 8.25e-140 | Down |
| NR4A2 | NM_006186.3 | 4.645393 | 0.000184 | Down |
| COL3A1 | NM_000090.3 | 4.219145 | 0.000112 | Down |
| CXCL14 | NM_004887.4 | 4.147009 | 5.25e-25 | Down |
| GPC4 | NM_001448.2 | 3.987747 | 2.14e-05 | Down |
| FBLN1 | NM_006487.2 | 3.951956 | 2.88e-07 | Down |
| FOSL2 | NM_005253.3 | 1.956347 | 3.48e-06 | Up |
| GADD45B | NM_015675.3 | 3.139243 | 4.41e-05 | Up |
| TBX19 | NM_005149.2 | 4.090872 | 2.02e-07 | Up |
| CYR61 | NM_001554.4 | 5.372227 | 0.00032136 | Down |
| JUNB | NM_002229.3 | 2.079143 | 1.33e-09 | Down |
| TFPI2 | NM_006528.3 | 1.893254 | 4.26e-07 | Down |
Validation of the expression of SFRP2 in
CD
To verify the RNA-Seq data, further analysis was
performed for six differentially expressed mRNAs, which were
included in the 122 DEGs, in another cohort of 23 CD patients and
three NHP samples by RT-qPCR. The DEGs, namely FOS like 2 AP-1
transcription factor subunit, growth arrest and DNA damage
inducible β, T-box 19, cysteine rich angiogenic inducer 61, JunB
proto-oncogene AP-1 transcription factor subunit and tissue factor
pathway inhibitor 2, were randomly selected (Table II). The qPCR results of the six
mRNAs were consistent with the RNA-Seq data, indicating the
validity of the RNA-Seq data (Fig.
2A). Furthermore, the RNA-Seq results were validated with
respect to SFRP2 by measuring its mRNA levels in samples procured
from 23 patients diagnosed with CD and three NHPs. There was a
significant decrease in SFRP2 mRNA expression in CD compared with
the NHP samples (Fig. 2B). IHC and
western blot analysis also revealed lower levels of SFRP2
expression in tumor tissues compared with normal controls (Fig. 2C and D). These results further
confirmed the decreased expression of SFRP2 in corticotroph
adenomas. The analysis of SFRP2 expression and association with
clinicopathological features of patients with CD is shown in
Table IV.
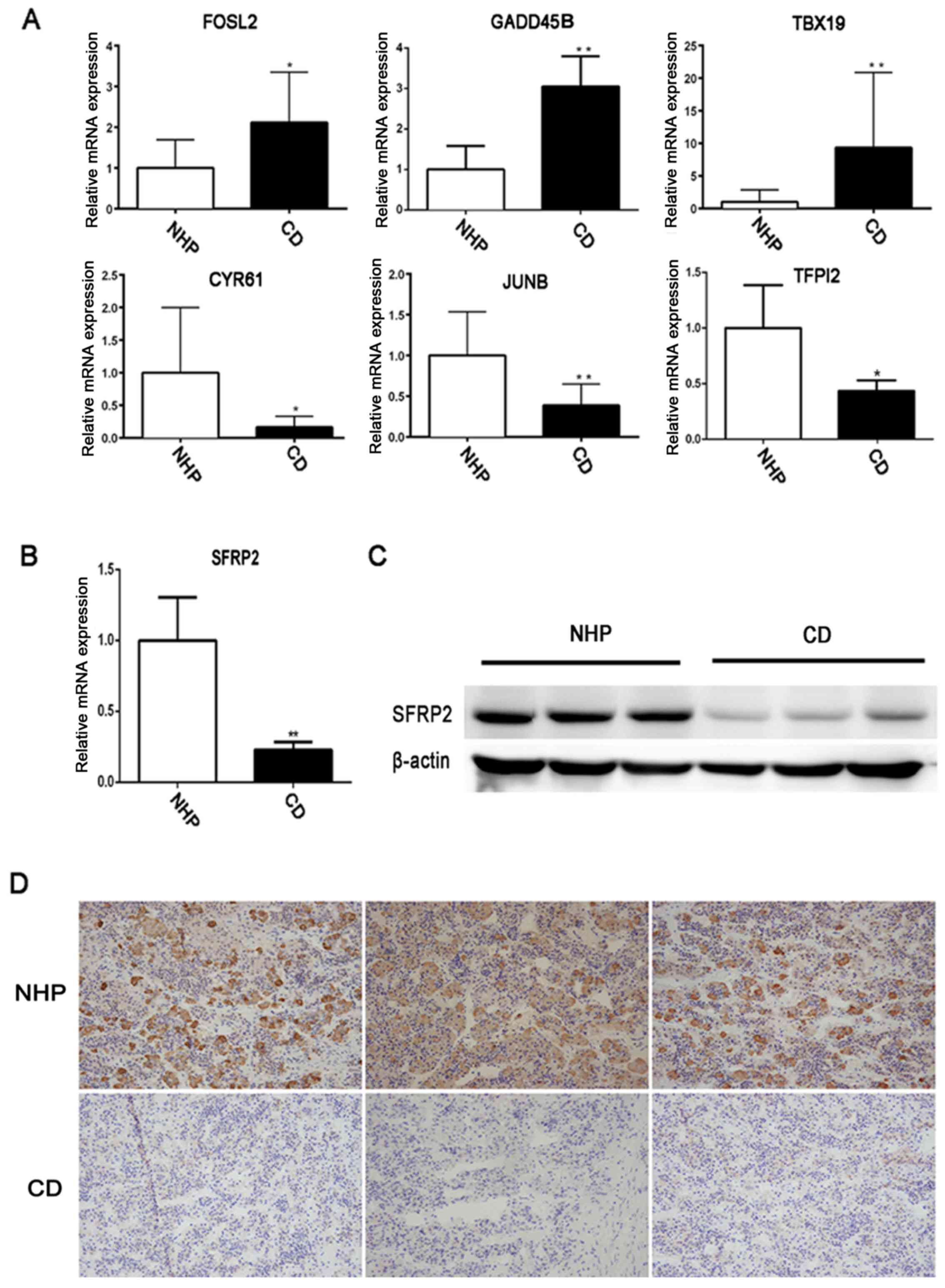 | Figure 2Validation of the expression of SFRP2
in human pituitary corticotroph adenoma. (A) Expression of six
randomly selected differentially expressed genes in CD tumor and
NHP samples were tested by RT-qPCR. (B) RT-qPCR and (C) western
blot analysis were used to detect SFRP2 expression in CD and NHP
samples. (D) Immunohistochemical staining of SFRP2 in a
representative NPH and CD corticotroph adenoma.
*P<0.05 and **P<0.01. RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; NHP, normal
human pituitary; CD, Cushing's disease; FOSL2, FOS like 2 AP-1
transcription factor subunit; GADD45B, growth arrest and DNA damage
inducible β; TBX19, T-box 19; CYR61, cysteine rich angiogenic
inducer 61; JUNB, JunB proto-oncogene AP-1 transcription factor
subunit; TFPI2, tissue factor pathway inhibitor 2; SFRP2, secreted
frizzled-related protein 2. |
| Table IVSFRP2 expression and
clinicopathological features in patients with Cushing's
disease. |
Table IV
SFRP2 expression and
clinicopathological features in patients with Cushing's
disease.
| Variable | SFRP2 expression [n
(%)]
| P-value |
|---|
| High
expression | Low expression |
|---|
| Sex | | | 1.000 |
| Male | 1 (4.35%) | 2 (8.70%) | |
| Female | 9 (39.13%) | 11 (47.82%) | |
| Age (years) | 45±10.88 | 44.69±11.58 | 0.949 |
| Tumor size | | | 0.104 |
| Microadenoma | 10 (43.48%) | 9 (39.13%) | |
| Macroadenoma | 0 (0%) | 4 (17.39%) | |
| Invasiveness | | | 0.339 |
| Yes | 1 (4.35%) | 4 (17.39%) | |
| No | 9 (39.13%) | 9 (39.13%) | |
| Midnight serum
cortisol (µg/24 h) | 21.08±5.81 | 20.74±8.08 | 0.912 |
| Urinary free
cortisol (µg/24 h) | 524.09±239.90 | 710.53±369.07 | 0.181 |
| Plasma ACTH
(pg/ml) | 148.01±173.53 | 309.79±508.49 | 0.348 |
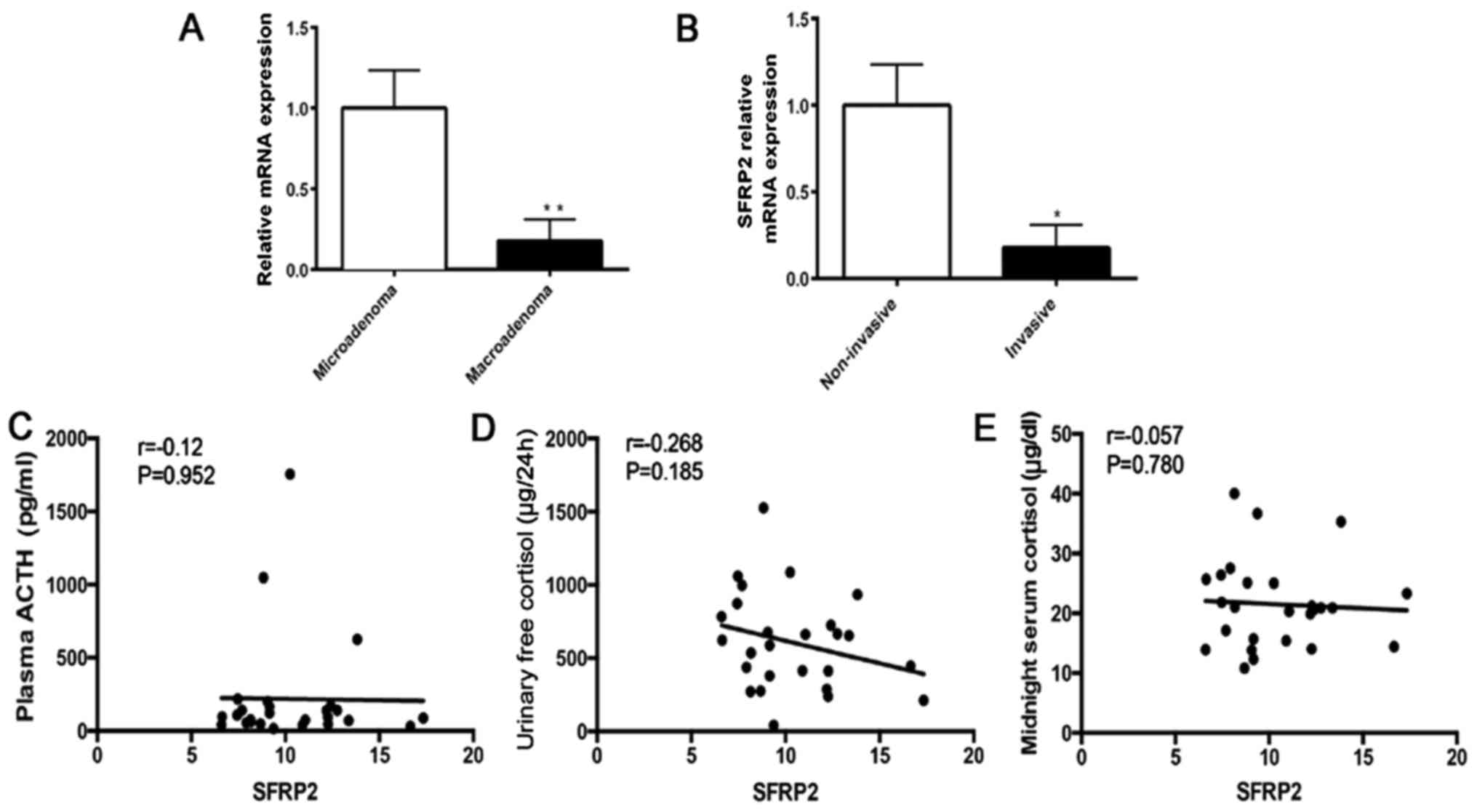
Lower expression of SFRP2 in corticotroph
adenoma is associated with larger size and increased
invasiveness
To further analyze the association between decreased
SFRP2 expression and clinical phenotype, the SFRP2 expression in 23
patients with CD and 3 NHP samples was evaluated (Table III). Analysis of relative RNA
expression indicated that the expression of SFRP2 was significantly
decreased in macroadenoma compared with mircroadenoma of CD
(Fig. 3A). Significant reduction
of SFRP2 was observed in invasive adenoma compared with
non-invasive adenoma of CD (Fig.
3B). These results indicated that lower expression of SFRP2 in
tumors is associated with a more aggressive character of pituitary
ACTH-secreting adenomas. In addition, the correlation between SFRP2
expression and hormone parameters in patients with CD was analyzed,
including plasma ACTH, urinary free cortisol and midnight serum
cortisol (data from the Department of Clinical Laboratory, Ruijin
Hospital, Shanghai, China). There was no significant correlation
between SFRP2 expression and these parameters (Fig. 3C–E). However, the results of the
correlation analysis may not be indicative of the true correlation,
because of the limited sample number.
| Table IIIClinical characteristics of the 26
patients with Cushing's disease. |
Table III
Clinical characteristics of the 26
patients with Cushing's disease.
| Case | Sex/age
(years) | Midnight serum
cortisol (µg/dl) | Urinary free
cortisol (µg/24 h) | Plasma ACTH
(pg/ml) | Tumor size | Invasiveness |
|---|
| 1 | F/51 | 13.95 | 239.02 | 94.29 | Microadenoma | No |
| 2 | F/52 | 15.39 | 414.46 | 37.575 | Negative | No |
| 3 | F/53 | 20.96 | 535.32 | 63.31 | Microadenoma | No |
| 4 | F/54 | 10.8 | 275.1 | 48.04 | Microadenoma | No |
| 5 | F/55 | 20.3 | 662.88 | 74.355 | Microadenoma | No |
| 6 | F/56 | 23.3 | 213.01 | 87.875 | Microadenoma | No |
| 7 | F/57 | 35.28 | 934.65 | 625.835 | Microadenoma | No |
| 8 | F/58 | 13.79 | 675.64 | 198.79 | Microadenoma | No |
| 9 | F/59 | 40.02 | 273.03 | 76.23 | Microadenoma | No |
| 10 | M/41 | 25.03 | 1085.83 | 1754 | Microadenoma | No |
| 11 | F/34 | 14.4 | 445.83 | 30.13 | Microadenoma | No |
| 12 | F/35 | 21.77 | 1059.03 | 217.56 | Huge adenoma | Invasive |
| 13 | F/36 | 19.85 | 287.36 | 139.055 | Microadenoma | No |
| 14 | F/37 | 12.34 | 379.49 | 122.12 | Macroadenoma | Invasive |
| 15 | F/38 | 25.7 | 623.1 | 95.09 | Microadenoma | No |
| 16 | F/39 | 15.69 | 587.5 | 166.37 | Microadenoma | No |
| 17 | F/40 | 20.91 | 654.26 | 71.19 | Microadenoma | No |
| 18 | M/24 | 25.08 | 1525.94 | 1048.19 | Macroadenoma | Invasive |
| 19 | F/31 | 21.2 | 412.62 | 46.29 | Microadenoma | No |
| 20 | F/32 | 17.09 | 996.96 | 140.595 | Huge adenoma | Invasive |
| 21 | F/33 | 26.39 | 872.34 | 107.58 | Negative | No |
| 22 | M/34 | 20.67 | 724.75 | 170.045 | Microadenoma | No |
| 23 | F/54 | 13.86 | 782.85 | 40.515 | Microadenoma | No |
| 24 | F/55 | 36.65 | 40.28 | 16.47 | Negative | No |
| 25 | F/56 | 20.93 | 666.54 | 141 | Microadenoma | Invasive |
| 26 | F/57 | 27.51 | 437.1 | 56.52 | Microadenoma | No |
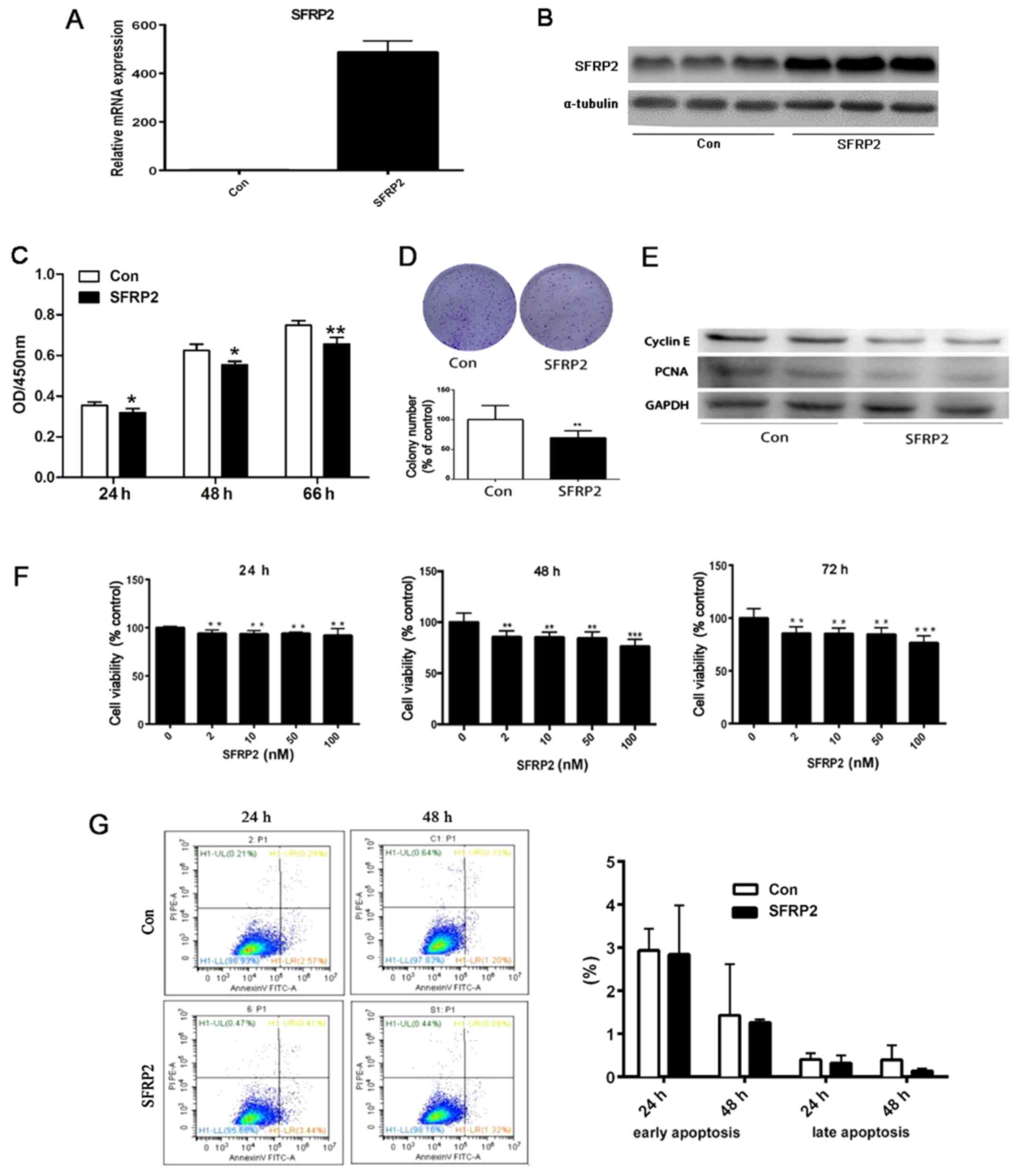
SFRP2 decreased the proliferation and
clonogenic ability of AtT20 cells in vitro
Since the expression of SFRP2 is downregulated in CD
and associated with larger tumor size, whether the overexpression
of SFRP2 reversed the biological characteristics of murine
pituitary corticotroph AtT20 cells was evaluated. A retroviral
vector containing wild type SFRP2 cDNA or empty vector (EV) to
generate stably transfected AtT20 cells. RT-qPCR and immunoblotting
were performed to evaluate the efficiency of overexpression. The
results demonstrated abundant SFRP2 mRNA and protein expression in
SFRP2-transfected AtT20 cells, but not in EV transfected cells
(Fig. 4A and B). Subsequently, it
was determined that cell proliferation was partly inhibited in a
time-dependent manner in AtT20 cells overexpressing SFRP2 with a
decrease in proliferation observed from 24 to 66 h (P<0.05)
(Fig. 4C). In addition, SFRP2
overexpression resulted in a reduction in the number AtT-20 cell
colonies compared with the vector control (P<0.01) (Fig. 4D). This decrease in cell growth in
AtT-20 cells overexpressing SFRP2 was confirmed by the reduction in
the levels of the proteins, PCNA and cyclin E, which are indicative
of proliferation, as determined by western blot analysis (Fig. 4E). In addition, AtT20 cells were
treated with recombinant SFRP2 protein and the absorbance was
detected at 450 nm. There was a decrease in cell proliferation in a
dose-dependent manner from 24–72 h (Fig. 4F). However, there was no marked
difference in the percentage of apoptotic cells between the
SFRP2-transfected or vector-transfected AtT20 cells when analyzed
by flow cytometry (Fig. 4G).
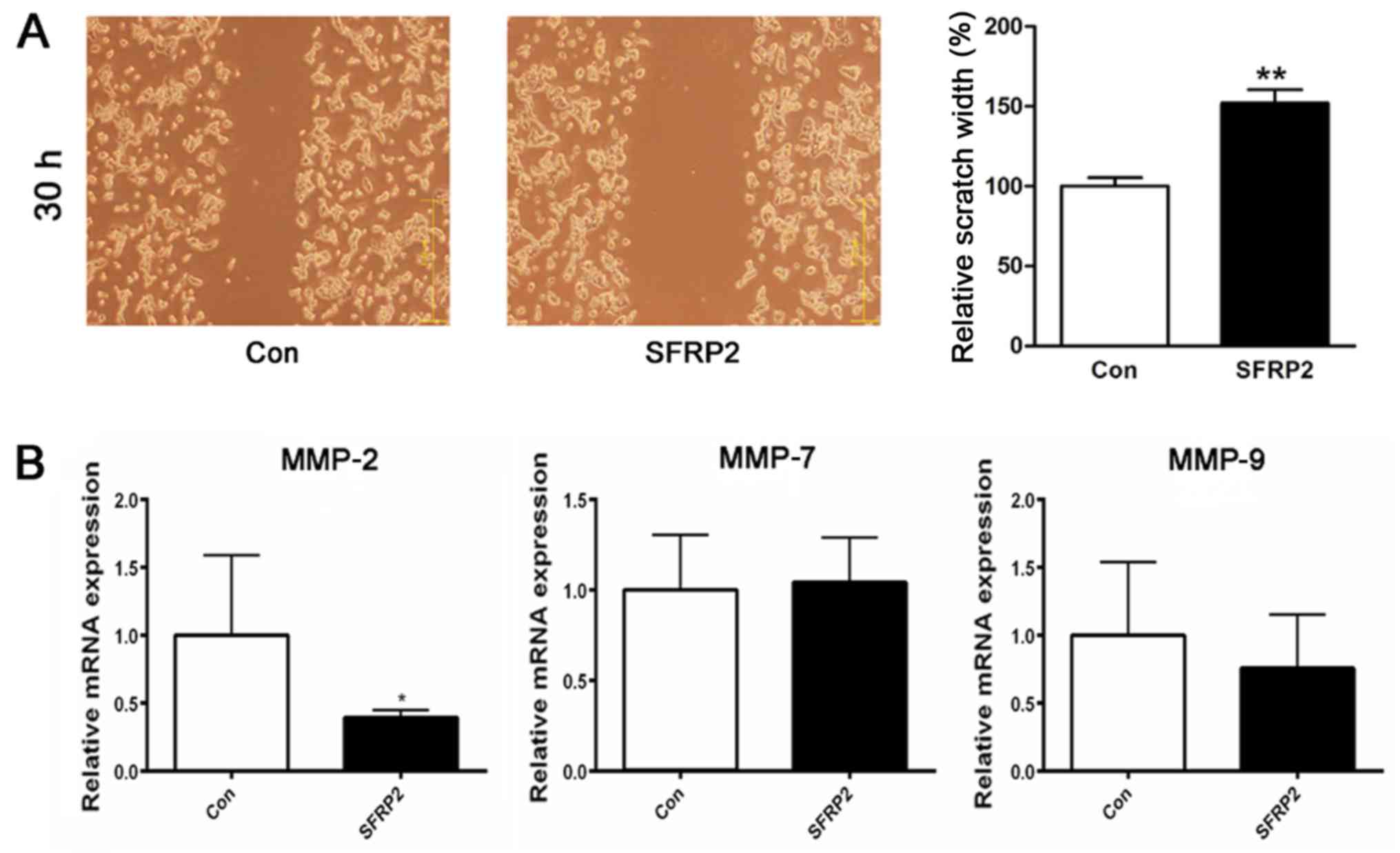
SFRP2 attenuates the migration of AtT20
cells in vitro
Previously, analysis of the clinical phenotype
indicated that the decreased expression of SFRP2 in corticotroph
adenoma was associated with higher invasiveness. Thus, it was
sought to determine whether SFRP2 expression also affected the
cellular migration of AtT20 cells. Migration was assayed in
SFRP2-transfected and vector-transfected AtT20 cells using a
wound-healing assay; the results revealed an evident delay in the
wound closure rate of the SFRP2-transfected cells at 30 h compared
with vector-transfected AtT20 cells (Fig. 5A). For further validation, the
expression of the matrix metallo-proteinases MMP-2, MMP-7 and
MMP-9, which are known to be associated with tumor invasion, were
analyzed RT-qPCR revealed that SFRP2 overexpression attenuated the
expression of MMP-2 in AtT20 cells (Fig. 5B).
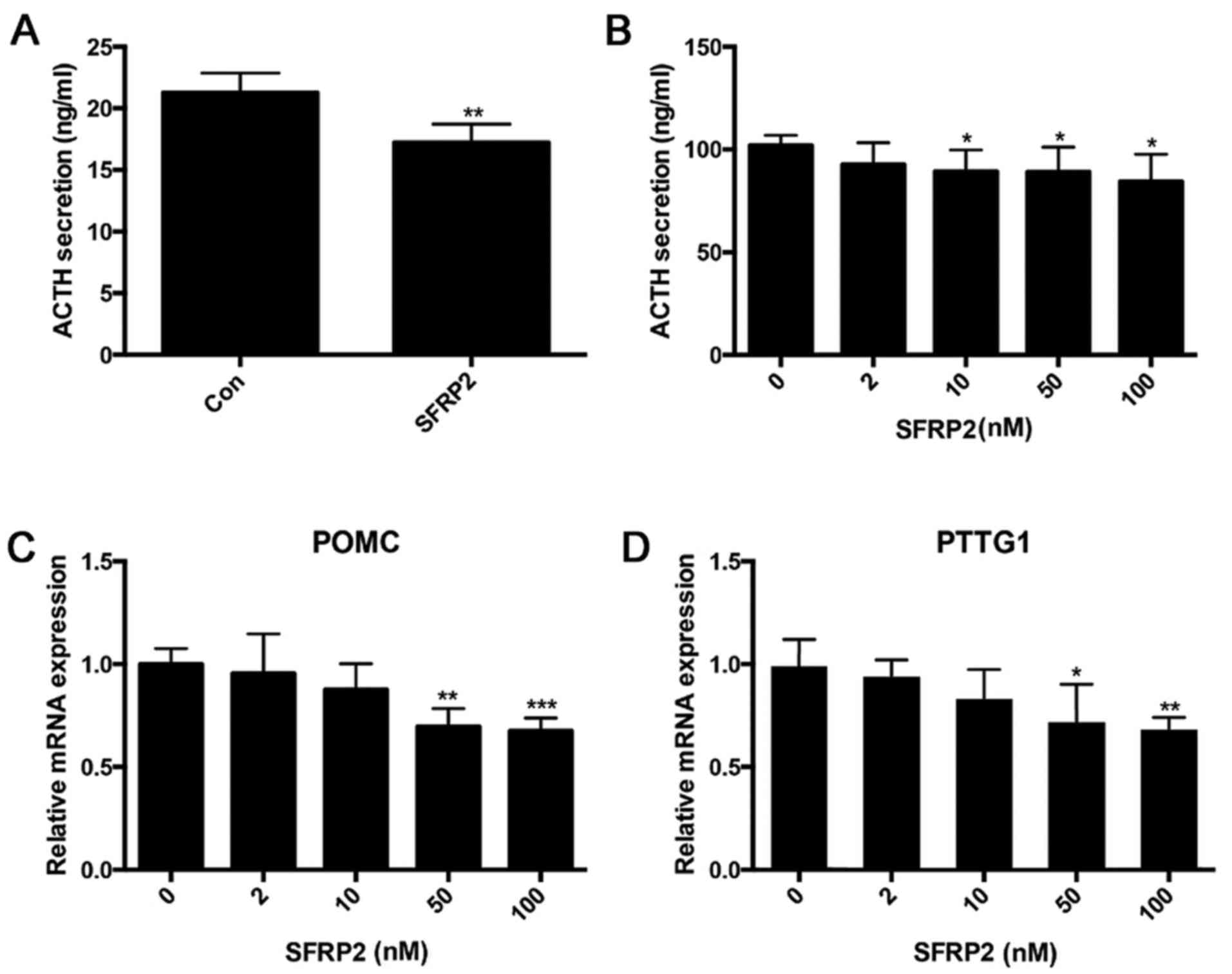
Overexpression of SFRP2 inhibits ACTH
secretion in AtT20 cells
Given the obvious decrease in SFRP2 expression in
pituitary corticotroph tumors, it was determined whether the level
of SFRP2 affected the secretion of ATCH. ACTH secretion was
measured in SFRP2-transfected and vector-transfected AtT20 cells,
the results demonstrated that overexpression of SFRP2 suppressed
the secretion of ACTH (Fig. 6A).
Treating AtT20 cells with a different dose of recombinant SFRP2
protein demonstrated that SFRP2 inhibited the secretion of ACTH in
a dose-dependent manner (Fig. 6B).
Additionally, SFRP2 suppressed the transcription of POMC and PTTG
(Fig. 6C and D).
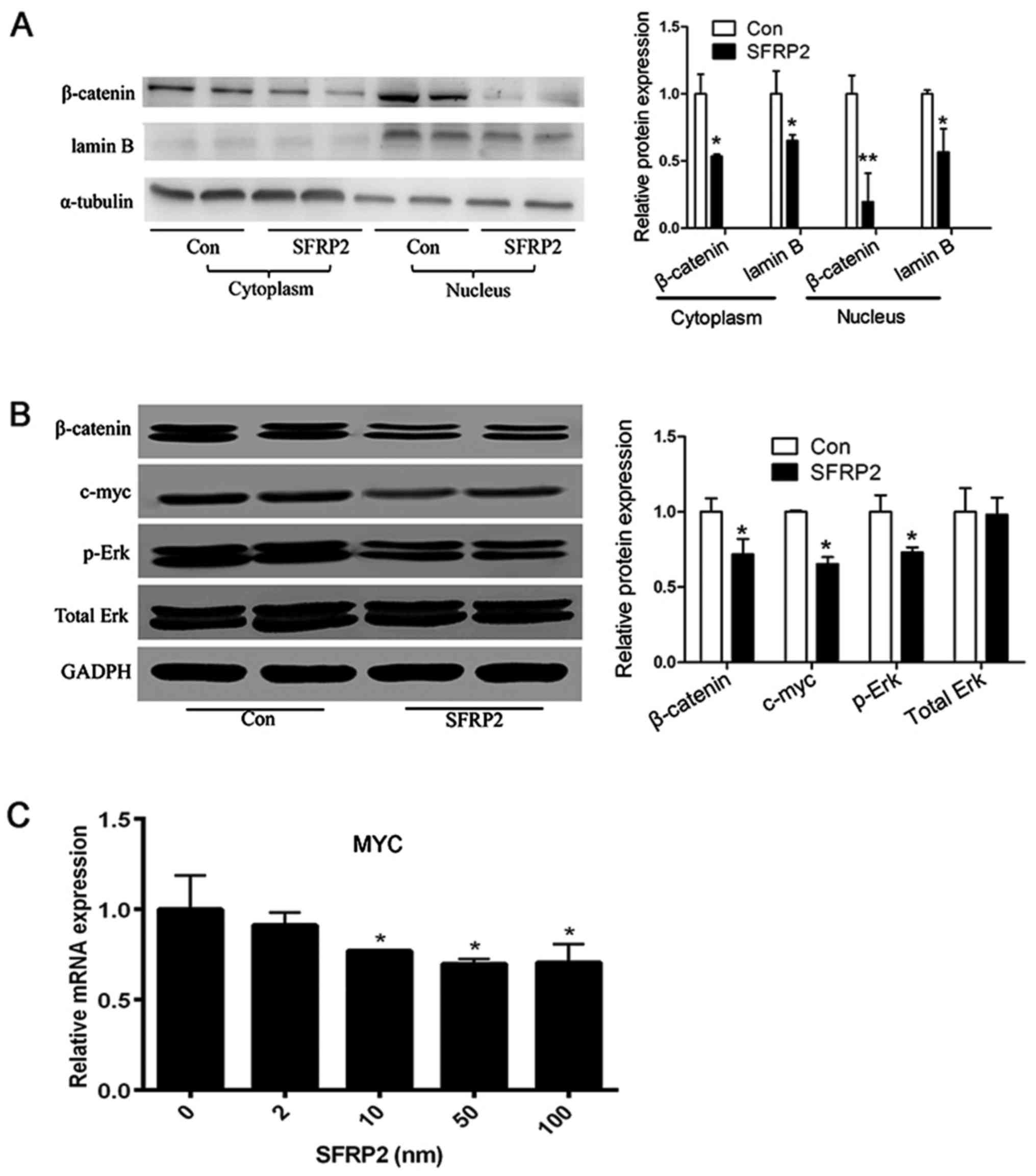
SFRP2 leads to decreased Wnt signaling
activity in AtT20 cells
Given that SFRP2 is an antagonist of the Wnt
signaling pathway, whether SFRP2 influences the activity of Wnt
signaling was investigated in vitro. In SFRP2-transfected
AtT20 cells, over-expression of SFRP2 reduced the amount of
β-catenin in the cytoplasm and nucleus (Fig. 7A). Furthermore, the expression
level of c-myc was detected, which is a downstream target of Wnt
signaling. The level of c-myc was markedly decreased in
SFRP2-transfected AtT20 cells, and the level of Erk phosphorylation
was also decreased compared with the vector-transfected AtT20 cells
(Fig. 7B). In addition, the
expression level of c-myc was decreased in AtT20 cells treated with
recombinant SFRP2 (Fig. 7C). These
data demonstrated that SFRP2 could attenuated the activity of Wnt
signaling in corticotroph adenoma AtT20 cells.
Discussion
As a typical type of next-generation sequencing
technology, high-throughput RNA-Seq has been widely used in the
recent studies of oncology due to its high sensitivity and
efficiency. Numerous valuable studies have reported on the
underlying molecular etiology or therapeutic targets in diverse
malignancies, including gallbladder carcinoma (15), prostate cancer (16) and liver cancer (17). This is the first report, however,
of the use of RNA-Seq in the study of CD.
In the present study, RNA-Seq was used to sequence
the transcriptome of human pituitary corticotroph adenoma, which
detected decreased expression of SFRP2. In addition, IHC and
western blot analysis of CD and NHP samples also confirmed the
decreased expression of SFRP2 in tumor tissues. Clinically, lower
expression of SFRP2 was associated with larger tumor sizes and
increased invasiveness. Thus, SFRP2 may be a useful marker for
predicting characteristics of pituitary ACTH adenoma. In fact,
researchers have already reported that SFRP2 expression is
associated with the degree of tumor malignancy and invasive ability
in various human cancers (12,18).
However, this is the first description of the aberrant expression
of SFRP2 in the tumorigenesis of CD.
The effects of SFRP2 overexpression on tumor cell
biology were analyzed in vitro. The data suggested that
overexpression of SFRP2 decreased the proliferation of AtT20 cells,
and reduce clonal ability and migration. In addition, increased
SFRP2 attenuated the secretion of ACTH in AtT20 cells. These data
are consistent with the clinical analysis of CD.
KEGG pathway enrichment analysis revealed that genes
classified in the Wnt signaling pathway were significantly enriched
in the CD samples. Indeed, several lines of evidence suggested that
SFRP2 has a pivotal role in the Wnt pathway, where it primarily
functions as an antagonist of Wnt signaling (19,20).
The Wnt signaling pathway has been reported to be involved in the
tumorigenesis of several cell types (21–25).
It has been reported that decreased expression of SFRP2 in
aggressive nonfunctioning pituitary adenoma results in nuclear
accumulation of β-catenin, and inhibitors of the Wnt pathway are
strongly downregulated in pituitary tumors (12,26).
To investigate whether SFRP2 influences Wnt signaling in CD, it was
confirmed that SFRP2 treatment reduced β-catenin and c-myc
expression. Similarly, AtT20 cells treated with SFRP2 exhibited
reduced expression of c-myc. These data demonstrate that SFRP2 can
antagonize Wnt signaling in AtT20 cells.
The major limitation of the current study is that
all the effects of SFRP2 were only verified in AtT20 cells. The
reason the AtT20 cell line was selected for the study is that it is
a very typical and representative cell line in CD research
(27). Currently, experiments are
being performed involving SFRP2 expression, its functional
verification and associated mechanism in other cell lines and mouse
models.
In conclusion, the findings of the present study
demonstrated that SFRP2 was downregulated in pituitary corticotroph
adenomas. In addition, lower expression of SFRP2 resulted in a
dysregulation of Wnt signaling. Thus, SFRP2 represents a promising
biomarker and therapeutic target for pituitary ACTH adenoma. A
pituitary tissue-specific SFRP2 knockdown mouse model will be
established in future research. Using transgenic mice and primary
cells, the phenotypes will be determined following SFRP2
overexpression and knockdown, and the underlying mechanism of SFRP2
in CD will be investigated.
Acknowledgments
Not applicable.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (nos. 81770779 and
81700692).
Availability of data and materials
All data generated or analyzed during this study are
available from the corresponding author on reasonable request.
Authors' contributions
JR, FJ, HJ collected the data and wrote the
manuscript. YS and SP analyzed the correlation between SFRP2
expression and clinical characteristics. CG and XC were involved in
data analysis and interpretation. As endocrinologists, WW and GN
made substantial contribution to the diagnosis of Cushing's
disease, provided technical assistance and revised the manuscript.
LB and QS were involved in the conception and design of the study,
and QS was the leader of this study. The final version of the
manuscript was read and approved by all authors.
Ethics approval and consent to
participate
Informed consent was obtained from all participating
patients. For the use of human samples, the protocol for this study
was approved by the Ruijin Hospital Ethics Committee (Shanghai,
China).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Nieman LK: Update in the medical therapy
of Cushing's disease. Curr Opin Endocrinol Diabetes Obes.
20:330–334. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Prague JK, May S and Whitelaw BC:
Cushing's syndrome. BMJ. 346:f9452013. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gadelha MR and Vieira Neto L: Efficacy of
medical treatment in Cushing's disease: A systematic review. Clin
Endocrinol (Oxf). 80:1–12. 2014. View Article : Google Scholar
|
|
4
|
Tritos NA, Biller BM and Swearingen B:
Management of Cushing disease. Nat Rev Endocrinol. 7:279–289. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wagenmakers MA, Netea-Maier RT, van
Lindert EJ, Timmers HJ, Grotenhuis JA and Hermus AR: Repeated
transsphenoidal pituitary surgery (TS) via the endoscopic
technique: A good therapeutic option for recurrent or persistent
Cushing's disease (CD). Clin Endocrinol (Oxf). 70:274–280. 2009.
View Article : Google Scholar
|
|
6
|
Saini S, Majid S and Dahiya R: The complex
roles of Wnt antagonists in RCC. Nat Rev Urol. 8:690–699. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Clevers H and Nusse R: Wnt/β-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Suzuki H, Watkins DN, Jair KW, Schuebel
KE, Markowitz SD, Chen WD, Pretlow TP, Yang B, Akiyama Y, Van
Engeland M, et al: Epigenetic inactivation of SFRP genes allows
constitutive WNT signaling in colorectal cancer. Nat Genet.
36:417–422. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Saito T, Mitomi H, Imamhasan A, Hayashi T,
Mitani K, Takahashi M, Kajiyama Y and Yao T: Downregulation of
sFRP-2 by epigenetic silencing activates the β-catenin/Wnt
signaling pathway in esophageal basaloid squamous cell carcinoma.
Virchows Arch. 464:135–143. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kongkham PN, Northcott PA, Croul SE, Smith
CA, Taylor MD and Rutka JT: The SFRP family of WNT inhibitors
function as novel tumor suppressor genes epigenetically silenced in
medulloblastoma. Oncogene. 29:3017–3024. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Takagi H, Sasaki S, Suzuki H, Toyota M,
Maruyama R, Nojima M, Yamamoto H, Omata M, Tokino T, Imai K, et al:
Frequent epigenetic inactivation of SFRP genes in hepatocellular
carcinoma. J Gastroenterol. 43:378–389. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu Y, Bai J, Hong L, Liu C, Yu S, Yu G and
Zhang Y: Low expression of secreted frizzled-related protein 2 and
nuclear accumulation of β-catenin in aggressive nonfunctioning
pituitary adenoma. Oncol Lett. 12:199–206. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
14
|
Jian FF, Li YF, Chen YF, Jiang H, Chen X,
Zheng LL, Zhao Y, Wang WQ, Ning G, Bian LG, et al: Inhibition of
ubiquitin-specific peptidase 8 suppresses adrenocorticotropic
hormone production and tumorous corticotroph cell growth in AtT20
cells. Chin Med J (Engl). 129:2102–2108. 2016. View Article : Google Scholar
|
|
15
|
Gu X, Li B, Jiang M, Fang M, Ji J, Wang A,
Wang M, Jiang X and Gao C: RNA sequencing reveals differentially
expressed genes as potential diagnostic and prognostic indicators
of gallbladder carcinoma. Oncotarget. 6:20661–20671. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pflueger D, Terry S, Sboner A, Habegger L,
Esgueva R, Lin PC, Svensson MA, Kitabayashi N, Moss BJ, MacDonald
TY, et al: Discovery of non-ETS gene fusions in human prostate
cancer using next-generation RNA sequencing. Genome Res. 21:56–67.
2011. View Article : Google Scholar :
|
|
17
|
Ho DW, Yang ZF, Yi K, Lam CT, Ng MN, Yu
WC, Lau J, Wan T, Wang X, Yan Z, et al: Gene expression profiling
of liver cancer stem cells by RNA-sequencing. PLoS One.
7:e371592012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Marsit CJ, Karagas MR, Andrew A, Liu M,
Danaee H, Schned AR, Nelson HH and Kelsey KT: Epigenetic
inactivation of SFRP genes and TP53 alteration act jointly as
markers of invasive bladder cancer. Cancer Res. 65:7081–7085. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chung MT, Lai HC, Sytwu HK, Yan MD, Shih
YL, Chang CC, Yu MH, Liu HS, Chu DW and Lin YW: SFRP1 and SFRP2
suppress the transformation and invasion abilities of cervical
cancer cells through Wnt signal pathway. Gynecol Oncol.
112:646–653. 2009. View Article : Google Scholar
|
|
20
|
Zhang Y and Chen H: Genistein attenuates
WNT signaling by up-regulating sFRP2 in a human colon cancer cell
line. Exp Biol Med (Maywood). 236:714–722. 2011. View Article : Google Scholar
|
|
21
|
Sano M, Driscoll DR, De Jesus-Monge WE,
Klimstra DS and Lewis BC: Activated wnt signaling in stroma
contributes to development of pancreatic mucinous cystic neoplasms.
Gastroenterology. 146:257–267. 2014. View Article : Google Scholar :
|
|
22
|
Zhang Y IV, Morris JP IV, Yan W, Schofield
HK, Gurney A, Simeone DM, Millar SE, Hoey T, Hebrok M and Pasca di
Magliano M: Canonical wnt signaling is required for pancreatic
carcinogenesis. Cancer Res. 73:4909–4922. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chambers TJ, Giles A, Brabant G and Davis
JR: Wnt signalling in pituitary development and tumorigenesis.
Endocr Relat Cancer. 20:R101–R111. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li Q, Shen K, Zhao Y, He X, Ma C, Wang L,
Wang B, Liu J and Ma J: MicroRNA-222 promotes tumorigenesis via
targeting DKK2 and activating the Wnt/β-catenin signaling pathway.
FEBS Lett. 587:1742–1748. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nojima M, Suzuki H, Toyota M, Watanabe Y,
Maruyama R, Sasaki S, Sasaki Y, Mita H, Nishikawa N, Yamaguchi K,
et al: Frequent epigenetic inactivation of SFRP genes and
constitutive activation of Wnt signaling in gastric cancer.
Oncogene. 26:4699–4713. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Elston MS, Gill AJ, Conaglen JV, Clarkson
A, Shaw JM, Law AJ, Cook RJ, Little NS, Clifton-Bligh RJ, Robinson
BG, et al: Wnt pathway inhibitors are strongly down-regulated in
pituitary tumors. Endocrinology. 149:1235–1242. 2008. View Article : Google Scholar
|
|
27
|
Liu N-A, Araki T, Cuevas-Ramos D, Hong J,
Ben-Shlomo A, Tone Y, Tone M and Melmed S: Cyclin E-mediated human
proopiomelanocortin regulation as a therapeutic target for cushing
disease. J Clin Endocrinol Metab. 100:2557–2564. 2015. View Article : Google Scholar : PubMed/NCBI
|