Introduction
Worldwide, the number of diagnosed lung cancer cases
amounts to 1.8 million every year. In fact, globally, this is the
leading cause of cancer-related deaths (1). Small cell lung cancer (SCLC) accounts
for ~15–17% of such lung cancers and is clearly related to
cigarette smoking since nearly all SCLC patients were or are active
smokers (2). This lung cancer
subtype is an aggressive malignancy associated with a poor patient
prognosis, with a 5-year survival rate at diagnosis rarely
exceeding 15% (3,4). Between 75 and 80% of patients suffer
from metastatic disease at the time of diagnosis. For this reason,
few patients may benefit from curative surgery (4). On the other hand, in the advanced
setting, conventional treatment includes combinations of different
chemotherapeutic agents such as cisplatin, carboplatin, etoposide,
irinotecan, topotecan, doxorubicin, adriamycin or cyclophosphamide
(4). Nonetheless, a significant
clinical challenge is the extremely frequent development of
multi-drug resistance, which makes the chemotherapy ineffective
after some time.
During the last few years, evidence has accumulated
in regards to the increased antitumoral effect of administering
conventional chemotherapy in combination with several antioxidants
(5,6). Moreover, natural products may be
useful anticancer agents. For example, the green tea polyphenol
epigallocathechin-3-gallate (EGCG) has been shown in different
studies to prevent cancer (7),
including lung cancer (8–11). Specifically in regards to SCLC,
EGCG has been demonstrated to induce cytotoxicity by the reduction
of telomerase activity (12). In
addition, DNA fragmentation and apoptosis as well as cell cycle
arrest in the S phase were observed. These preclinical in
vitro data point to the potential use of EGCG in the treatment
of SCLC patients.
Ocoxin® oral solution (OOS) is a
nutritional supplement whose formulation includes several compounds
with anticancer activity, including EGCG (13), vitamin B6, vitamin C, or cinnamic
acid (6,14,15).
In addition, OOS contains glycyrrhizinic acid, which exhibits
anti-inflammatory and immunomodulatory effects (16). Given such composition, OOS is
currently being investigated in clinical trials as part of the
treatment of several types of cancer, demonstrating, to date, an
improvement in the quality of life of such patients (17,18).
Moreover, several recent studies have investigated the potential
antitumor effect of OOS on different tumor models, including
HER2-positive breast cancer (13),
acute myeloid leukemia (19) and
hepatocellular carcinoma (20). In
all these models, OOS exhibited clear antitumor properties both
in vitro and in vivo in xenograft mouse models. At
the mechanistic level, OOS seemed to induce a general delay of cell
cycle progression. In fact, in breast cancer as well as in AML
models, cell cycle blockage seems to be mediated by the increase in
the cell cycle inhibitor p27 (13,19).
Based on these precedents, the effects of OOS on
preclinical models of SCLC were explored. The potential
antiproliferative action of this formulation was initially assessed
in vitro using two different SCLC cell lines alone and in
combination with other conventional antitumoral drugs. Its in
vivo action was analyzed using a xenograft model that showed a
reduction in tumor growth in animals that received OOS. Finally,
the mechanisms responsible for such decrease were examined both
in vitro and in vivo.
Materials and methods
Reagents and antibodies
Cell culture media,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
hematoxylin and eosin (H&E) were purchased from Sigma-Aldrich
(St. Louis, MO, USA). Fetal bovine serum (FBS) and antibiotics were
from Life Technologies (Carlsbad, CA, USA) and Immobilon P (PVDF)
membranes from Millipore (Billerica, MA, USA). OOS was provided by
Catalysis, S.L. (Madrid, Spain). Its formulation included per 50 ml
of solution: L-glycine 1,000 mg, glucosamine sulfate 1,000 mg,
L-arginine 320 mg, L-cysteine 102 mg, licorice (Glycyrrhiza
glabra L.) 100 mg, vitamin C (L-ascorbic acid) 60 mg, water,
zinc sulfate 40 mg, green tea extract (Camellia sinensis L.
Kuntze) 12.5 mg, vitamin B5 (D-calcium pantothenate) 6 mg, vitamin
B6 (pyridoxine hydrochloride) 2 mg, manganese sulfate 2 mg,
cinnamon extract (Cinnamomum zeylanicum Blume) 1.5 mg, folic
acid (pteroylmonoglutamic acid) 200 μg, vitamin B12
(cyanocobalamin) 1 μg, acidulant (malic acid) and
preservative (sodium methylparaben). Other generic chemicals were
purchased from Sigma-Aldrich, Roche Biochemicals (Basel,
Switzerland) or Merck (Darmstadt, Germany).
The origins of the different antibodies used in the
western blot analyses were as follows: the anti-glyceraldehyde
3-phosphate dehydrogenase (GAPDH) (sc-166574, mouse monoclonal,
used at 1:10,000) and the anti-PARP (sc-8007, mouse monoclonal,
1:5,000) antibodies were purchased from Santa Cruz Biotechnology
(Santa Cruz, CA, USA); the anti-Rb (#554136, mouse monoclonal,
1:1,000) and anti-caspase-3 (#610323, rabbit polyclonal, 1:5,000)
from BD Biosciences (Franklin Lakes, NJ, USA) and the anti-p27
(#3686, rabbit polyclonal, 1:5,000), anti-caspase-8 (#9746, mouse
monoclonal, 1:1,000), anti-caspase-9 (#9502, rabbit polyclonal,
1:1,000), anti-caspase-7 (#9492, rabbit polyclonal, 1:1,000),
anti-cleaved caspase-3 (#9664, rabbit polyclonal, 1:1,000) and
anti-bid (#2002, rabbit polyclonal, 1:1,000) from Cell Signaling
Technology, Inc. (Danvers, MA, USA). The horseradish
peroxidase-conjugated secondary antibodies (goat anti-rabbit,
170-6515, 1:20,000 and goat anti-mouse 170-6516, 1:10,000) were
from Bio-Rad Laboratories Inc. (Hercules, CA, USA).
Cell culture, protein extraction and
western blotting (WB)
SCLC cell lines were grown in RPMI-1640 medium
supplemented with 10% FBS and antibiotics. Cells were cultured at
37°C in a humidified atmosphere in the presence of 5%
CO2-95% air. The two SCLC cell lines used in this study
have already been described (21–23)
and were generously provided by Dr Clementi (CNR Center of
Cytopharmacology, Milan, Italy). Furthermore, GLC-8 cells were
initially isolated from a tumor biopsy of an SCLC (22), while DMS 92 cells came from a bone
marrow metastasis of an SCLC (21).
To prepare for protein purification, cells were
harvested by centrifugation, washed in phosphate-buffered saline
(PBS) and lysed in ice-cold lysis buffer (24). Protein quantification, sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
WB analysis were performed as previously described (24).
Cell cycle and BrDU incorporation
assay
For cell cycle analysis by flow cytometry,
ethanol-fixed cells were stained with 5 μg/ml propidium
iodide (PI) and 250 μg DNase-free RNAse. A total of 50,000
cells were acquired in the PI gate by using a BD Accuri™ C6 flow
cytometer and the C6 (version 1.0.264.21) software (BD
Biosciences).
To measure bromodeoxyuridine (BrDU) incorporation,
the Cell Proliferation ELISA, BrdU (colorimetric) kit (Roche
Diagnostics GmbH, Mannheim, Germany) was used following the
manufacturer's instructions. Data were acquired using a BioTek
Synergy 4 reader with Gen5 1.05 software (both from BioTek
Instruments, Inc., Winooski, VT, USA).
Assessment of cell death and cell
proliferation
For analysis of apoptotic cell death, GLC-8 cells
were treated for 48 h with OOS diluted 1:25 in culture media or
left untreated (control), resuspended in binding buffer (10 mM
HEPES, 140 mM NaCl, 2.5 nM CaCl2, pH 7.4) containing 5
μl Annexin V-FITC (BD Biosciences) and 5 μl 50
μg/ml PI, and stained at room temperature for 15 min. A
total of 50,000 cells were acquired using the BD Accuri™ C6 flow
cytometer (BD Biosciences) as previously described.
Cell proliferation of SCLC cells was examined using
a modified MTT metabolization assay (19). At least three wells were analyzed
for each condition, and the results are presented as the mean ± SD
of a representative experiment repeated at least twice.
In vivo experiments
For the animal studies, 12 7-week-old female athymic
mice (CB17-SCID) were purchased from Charles River Laboratories
(Wilmington, MA, USA) and maintained in pathogen-free housing at
our Institutional Animal Care Facility. Animal experiments were
performed according to the institutional guidelines and protocol
approved by the Ethics Committee of CSIC-Universidad de Salamanca
(Salamanca, Spain). One week later, 6×106 GLC-8 cells
resuspended in 50 μl of RPMI-1640 and 50 μl of
Matrigel were subcutaneously injected into the right caudal flank
of each animal. When tumors became palpable, mice were randomized
into two groups (n=6 per group), that were administered vehicle
alone (control group) or 100 μl OOS per mouse (20 g).
Treatments were administered for 31 days with a daily schedule
(Monday to Friday) by oral gavage. Mice were weighed and tumors
measured twice a week with a digital caliper (Proinsa, Vitoria,
Spain). Tumor volumes were calculated using the following formula:
V = (L/2) × (W/2)2 × 4/3 × π, where V, volume (cubic
mm); L, length (mm); and W, width (mm). At the time of sacrifice,
tumor tissue was resected and part of the tumor was immediately
frozen at −80°C. Another part was fixed in formalin for further
analyses.
Histological and immunohistochemical
(IHC) analyses
Representative tumor areas were fixed in formalin,
paraffin-embedded, cut in 2- to 3-μm sections, and either
stained with H&E or prepared for IHC, that was performed as
previously described (19). Thus,
two cell conditioning periods of 8 min at 95°C and 4 min at 100°C
on a hot plate using Tris-EDTA (pH 8.0) buffer were performed on
previously dewaxed formalin-fixed paraffin-embedded sections.
Sections were then incubated with the a 1:50 dilution of an
anti-Ki-67 antibody (clon SP6; Master Diagnóstica, Granada, Spain)
and the staining was performed with the IHC 3,3′-diaminobenzidine
(DAB) system (Ventana Medical Systems, Tucson, AZ, USA). For TUNEL
staining, the In Situ Cell Death Detection kit (Roche
Diagnostics GmbH, Mannheim, Germany) was used, and counterstained
with 4′,6-diamino-2-phenylindole. Results were evaluated in a
manner blinded to the clinicopathological and molecular data. The
number and intensity of immunoreactive cells were evaluated in at
least 10 randomly selected fields. These procedures were conducted
by independent personnel of the pathology unit of our center.
Conflict measurements were solved by consensus.
RNA isolation, cDNA synthesis and
microarray hybridization and analysis
After thawing, tumors were excised and lysed in
TRIzol reagent (Life Technologies), according to the manufacturer's
instructions. Briefly, tumors were homogenized (Dispomix; L&M
Biotech, Holly Springs, NC, USA) and incubated in TRIzol solution
for 2 min at room temperature, before the addition of chloroform.
Tubes were vigorously shaken and the different phases were
separated by centrifugation at 18,000 × g and 4°C for 15 min. The
upper, aqueous phase was recovered and the RNA present was
precipitated with isopropyl alcohol. Once washed in 70% ethanol,
the resultant RNA was column-purified (RNeasy Mini kit; Qiagen,
Inc., Valencia, CA, USA) and its integrity was assessed (Agilent
2100 Bioanalyzer; Agilent Technologies, Inc., Santa Clara, CA,
USA). Biotinylated complementary RNA was then synthesized (Enzo
Life Sciences, Inc., Farmingdale, NY, USA) and hybridized to human
Clarion™ S GeneChip oligonucleotide arrays (Affymetrix; Thermo
Fisher Scientific, Inc.). Quantitation of fluorescence intensities
of probesets was conducted using the GenArray Scanner (Hewlett
Packard). Unprocessed files were normalized using the RMA algorithm
implemented in the Affymetrix Expression Console (Thermo Fisher
Scientific, Inc.). Differentially expressed genes were identified
using significant analysis of microarrays, selecting all genes with
a value of Q≤0.05.
Statistical analysis
Each condition was analyzed in triplicate or
quadruplicate and data are presented as the mean ± SD of an
experiment that was repeated at least three times. Comparisons of
continuous variables between two groups were performed using
two-sided Student's t-test. Differences were considered to be
statistically significant when P-values were <0.05.
Results
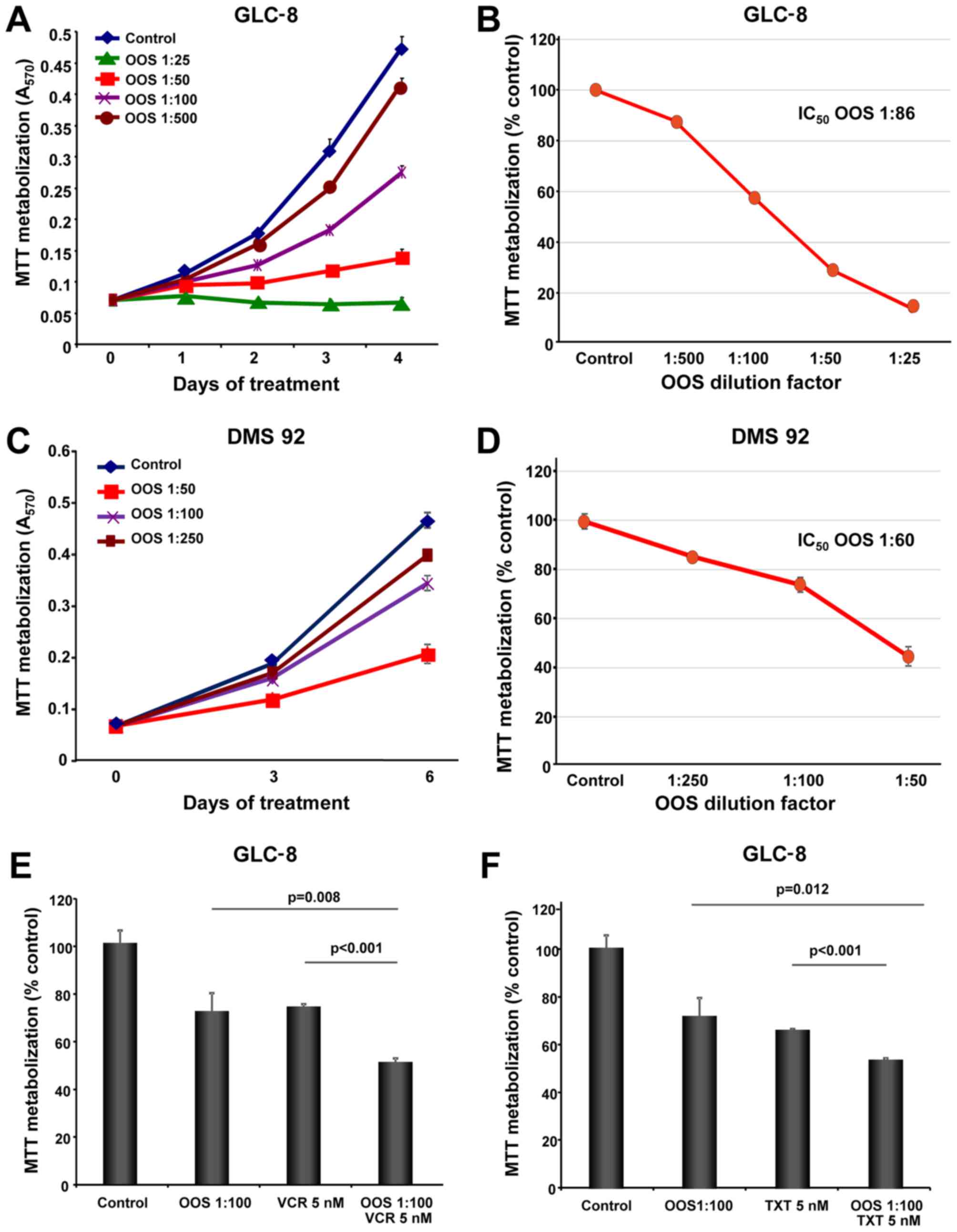
Effect of OOS on the proliferation of
SCLC cell lines
The in vitro action of OOS was evaluated in
the SCLC cell lines GLC-8 and DMS 92. With this purpose, cells were
grown in the presence of increasing doses of OOS diluted in the
culture medium and their proliferation was evaluated using MTT
metabolization assays performed at different days of treatment. OOS
decreased MTT metabolization in these cells in a time- and
dose-dependent manner (Fig. 1A and
C). The IC50 values determined for GLC-8 cells after
4 days and DMS 92 cells after 6 days of treatment were a 1:86
dilution and a 1:60 dilution, respectively (Fig. 1B and D).
Effect of OOS in combination with
standard of care SCLC treatments
In most of the cases, the success of antitumor
therapies is based on the combination of different agents. For this
reason, we aimed to ascertain whether the combination of OOS with
drugs commonly used in the SCLC clinic could improve their
antitumor effect. Thus, GLC-8 cells were treated for 2 days with
OOS alone (diluted 1:100 in regular media), or in combination with
vincristine (VCR, 5 nM), docetaxel (TXT, 5 nM) or cisplatin (2 or
20 μM). The combination of OOS with this latter compound did
not exhibit a superior antiproliferative effect to the one observed
for each of the individual treatments (data not shown). In the case
of VCR or TXT, the combinations were more effective for inhibiting
MTT metabolization than the independent treatment with any of the
agents (Fig. 1E and F). In fact,
when the combination index (CI) of OOS and VCR was calculated using
Calcusyn software (Biosoft, Cambridge, UK) an additive effect of
both drugs was demonstrated (CI between 0.9 and 0.96, data not
shown).
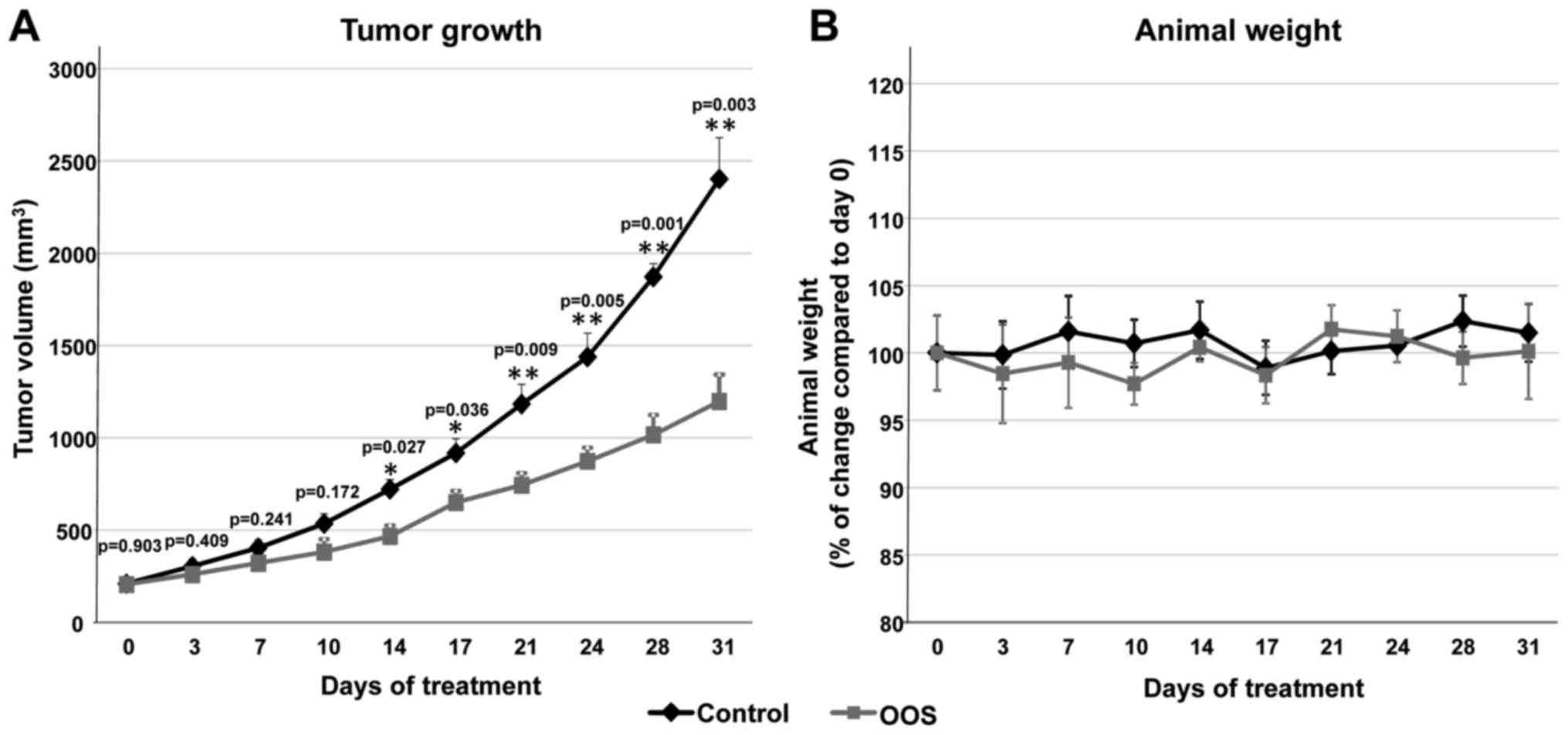
In vivo efficacy of OOS in SCLC murine
models
Given the antitumoral action of OOS in vitro,
we explored whether this effect could also be observed in
vivo. With this aim, we used a xenograft murine model to
determine the effect of OOS treatment once the tumors were already
established. CB17-SCID athymic mice were injected with GLC-8 cells
and when tumors were palpable and had correctly been engrafted and
started to grow, the animals were randomized into two different
groups that were orally treated with water (control) or 100
μl OOS per 20 g of animal weight (OOS). Initially, tumors
included in the two groups had a similar volume (209.3±17.2 vs.
206.1±16 mm3, mean ± SEM, respectively). Tumoral sizes
were measured twice a week for a total time period of 31 days.
After 14 days of oral treatment with OOS, a significant decrease in
the growth of the tumors was observed (p=0.027) (Fig. 2A). Nonetheless, such differences
increased along the time of treatment. Thus, at the end of the
experiment, the mean tumor volume of the control mice was
2,402.6±223.6 mm3, as compared with 1,197±144.4
mm3 for the group treated with OOS (p=0.003) (Fig. 2A). The mean body weight of the
control or treated mice did not substantially change throughout the
duration of the experiment (Fig.
2B), indicative of good tolerability to OOS in our experimental
conditions.
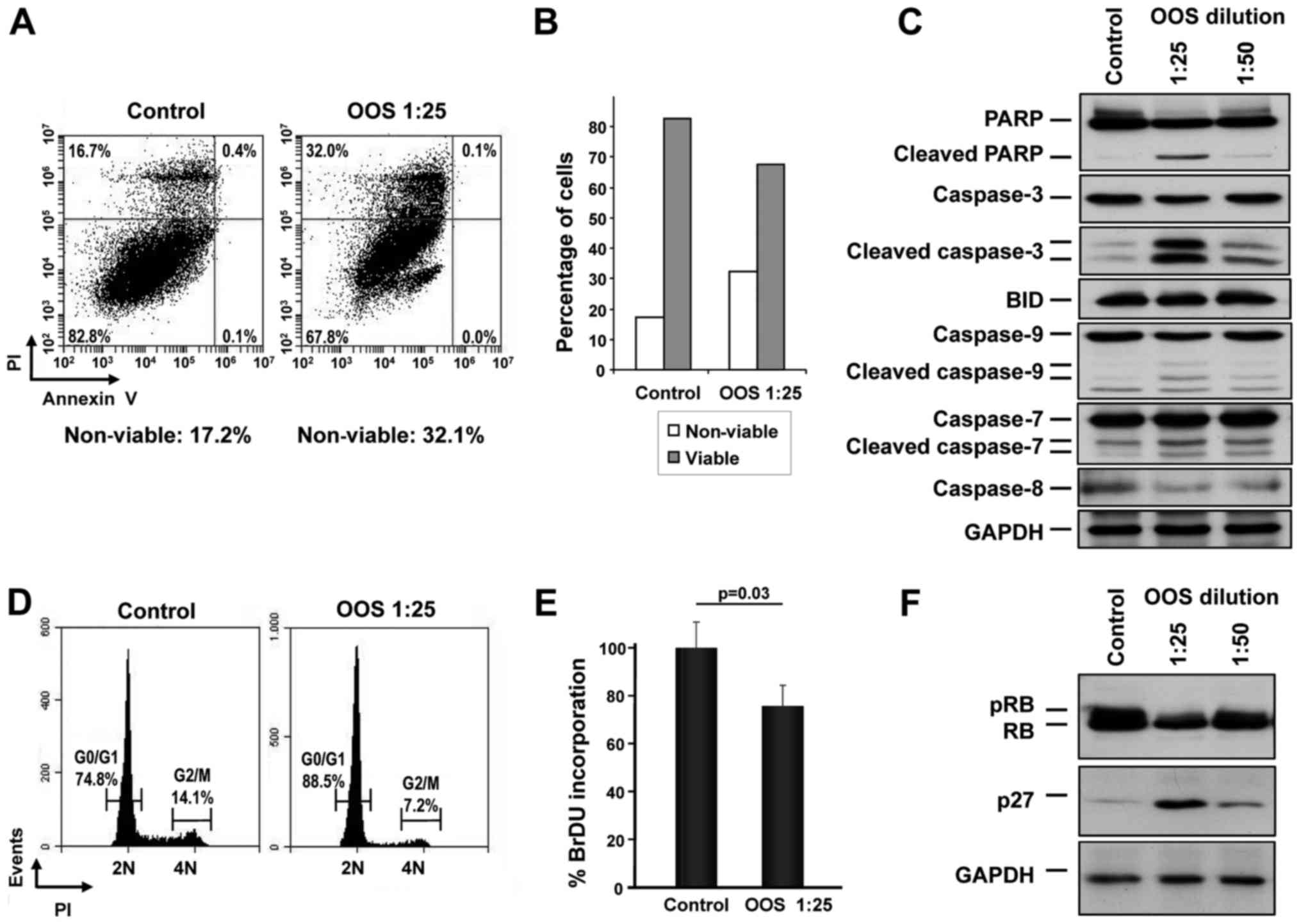
Mechanism of action of OOS on SCLC
cells
The mechanisms leading to the in vitro
reduction of MTT metabolization were next analyzed. We proposed
that such a reduction could be due to an increase in cell death,
reduced cell cycle progression or a combination of both. Thus, the
capability of OOS to induce apoptotic cell death was assessed.
GLC-8 cells were treated for 24 h with OOS diluted at 1:25 in
culture medium and apoptotic cell death was determined by double
staining with Annexin V and PI. A slight increase in the percentage
of cells stained by both Annexin V and PI was observed after OOS
treatment (control, 16.7% vs. OOS-treated, 32.0%) (Fig. 3A). Analysis of the total viable vs.
non-viable population confirmed the induction of cell death by OOS
(Fig. 3B). Moreover, when the
levels of the cleavage of several proteins involved in apoptosis
such as PARP or different caspases were determined by western
blotting, a clear induction of their proteolytic processing
indicative of activation was detected, especially at dilutions of
OOS of 1:25 (Fig. 3C).
We next investigated whether OOS could induce
changes in cell cycle progression. Cells were treated with OOS and
cytometrically analyzed after PI staining of the DNA. In those
conditions, treatment with OOS resulted in an increase in the G0/G1
population, increasing from 74.8 to 88.5%. Consequently, there was
a concomitant decrease in the percentage of cells in the G2/M (from
14.1 to 7.2%) or S phases (from 11.1 to 4.3%) following treatment
with OOS (Fig. 3D). The capability
of OOS to interfere with DNA synthesis was evaluated using an
alternative technique that allows the quantification of cells
progressing through S phase by the measure of its BrDU
incorporation. With this assay, a reduction in the S phase
population was demonstrated (Fig.
3E), pointing to a slowdown in cell cycle progression as part
of the mechanism of action of OOS.
The cell cycle effects of OOS in other cellular
models seems to be mediated by an increase in the cell cycle
inhibitory protein p27 (13,19).
To explore whether such a mechanism of action could as well
participate in the effect of OOS on SCLC cells, p27 protein levels
were determined by western blotting both in control untreated cells
or after 24 h of treatment with 1:25 or 1:50 dilutions of OOS. When
a dilution factor of 1:25 was used, a clear increase in the p27
level was observed (Fig. 3F).
Moreover, this increase was correlated with a decrease in the
phosphorylation and total levels of the retinoblastoma protein (pRB
and RB, respectively) (Fig.
3F).
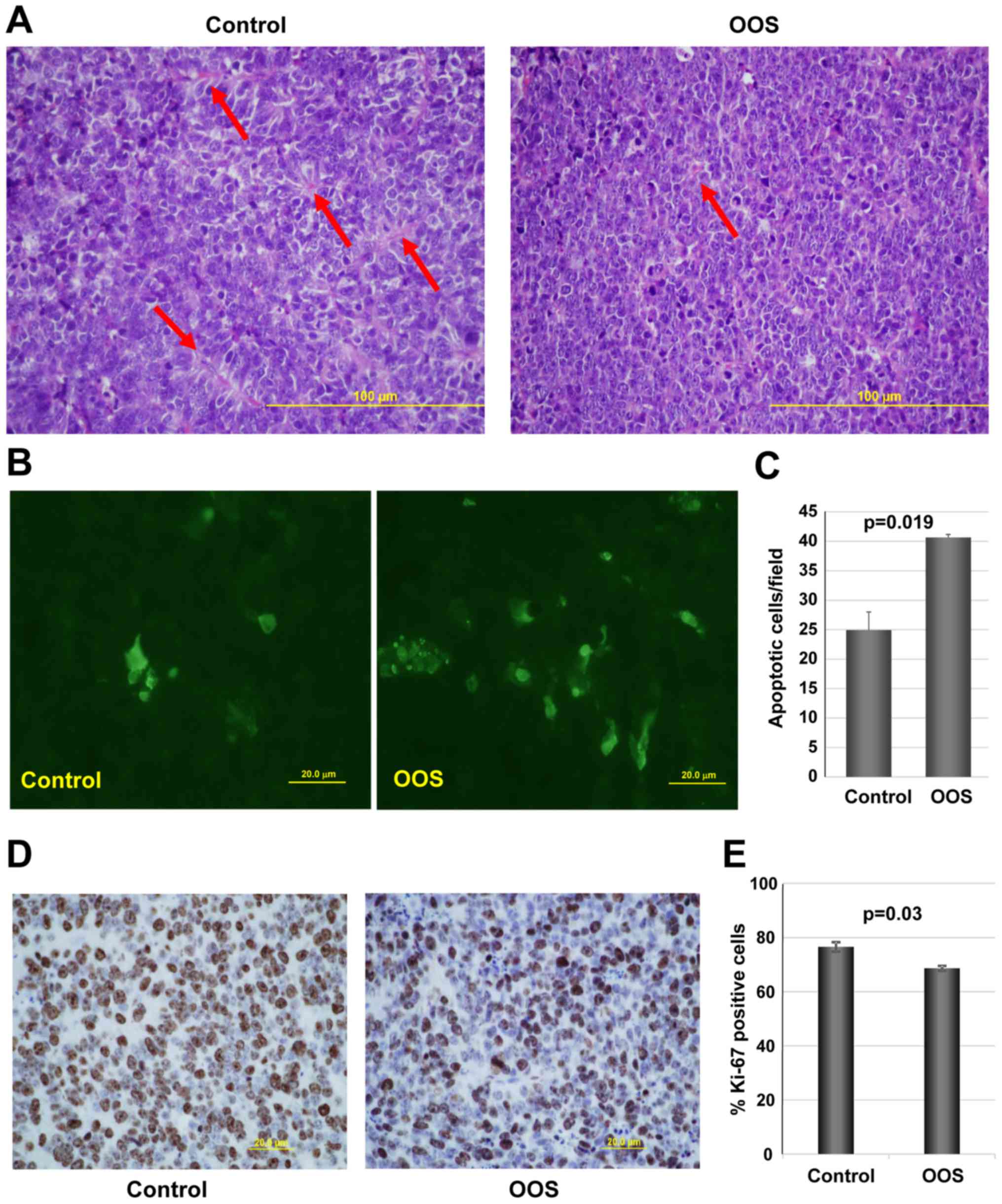
Effect of OOS on cell proliferation and
apoptosis in vivo
We next investigated the potential mechanisms
leading to the decrease in tumor growth observed in OOS-treated
animals, when compared to the untreated controls. We first
evaluated the general tumor aspects after H&E staining.
Independently of whether the animal received treatment of not, the
tumoral masses dissected from the animals presented a large central
apoptotic/necrotic area that extended to up to 60% of the tumor
(data not shown). Tumor cells were in general poorly
differentiated, with a high mitotic index. Additionally, control
tumors seemed to be better vascularized than OOS-treated ones. In
fact, control tumors seemed to be more pseudo-glandular than the
treated ones (arrows in Fig. 4A).
In line with this, tumors from animals which received OOS exhibited
a more compact aspect (Fig.
4A).
We next evaluated the level of apoptotic cells in
the control and OOS-treated animals. While, as mentioned above,
when observed under a microscope the central apoptotic/necrotic
area observed was similar in both experimental conditions, in
OOS-treated tumors more apoptotic regions within non-necrotic
tissue could be observed. Analysis of apoptotic cell death after
TUNEL staining technique showed that tumors from the OOS-treated
animals contained a larger number of apoptotic cells than tumors
from untreated animals (Fig. 4B).
Moreover, quantitative analyses of the TUNEL-stained tumors
corroborated that the number of apoptotic cells detected per field
was significantly lower in the control condition than that in the
treated tumors (p=0.019) (Fig.
4C). Together, these data demonstrate that OOS triggers the
apoptotic cell death of SCLC cells in the in vivo
setting.
The expression of human Ki-67, a protein associated
with cell proliferation, was also evaluated. Treatment with OOS
induced a statistically significant decrease in the percentage of
Ki-67-positive cells (Fig. 4D and
E). The decrease in Ki-67 levels was indicative of a reduced
proliferation of OOS-treated tumors.
Transcriptional effect of OOS in
vivo
To gain further insight into the molecular mechanism
leading to the in vivo reduction in tumor growth, we
investigated the changes in gene expression profiles induced by OOS
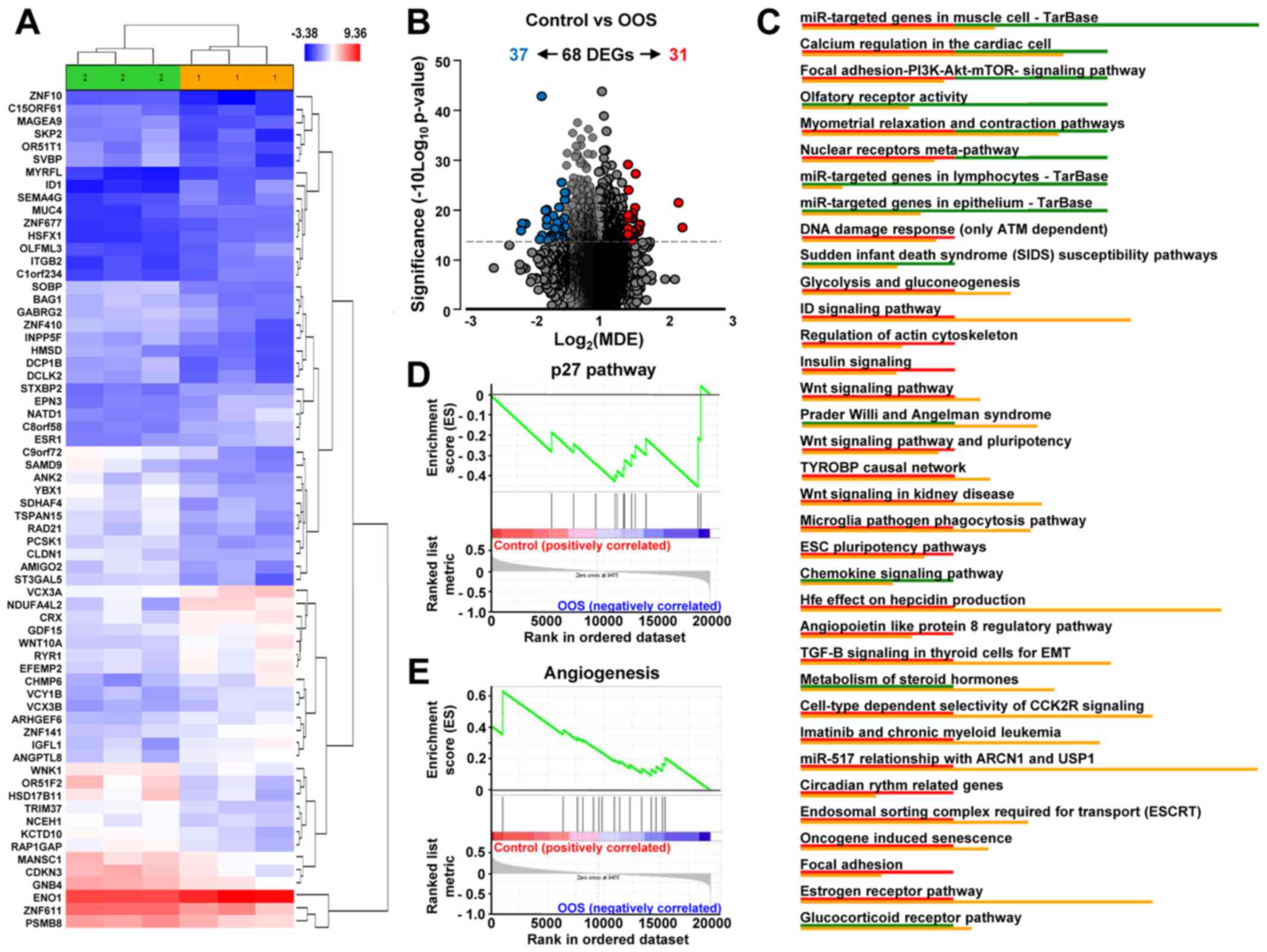
treatment. Unsupervised clustering of the gene expression data
clearly associated samples into two groups, control and
OOS-treated, indicating that treatment with OOS was able to change
the transcriptome to make treated samples to diverge from the
untreated control tumors (Fig.
5A). Quantitative comparison of both conditions defined 68
deregulated genes (fold change of at least ±1.5) (Table I). Volcano plots indicated that
among them, 31 genes were upregulated in the control condition
whereas 37 were increased after OOS treatment (Fig. 5B). Among the deregulated genes,
NADH dehydrogenase NDUFA4L2 (2.26-fold higher expression), the ID1
inhibitor of DNA binding (2.2-fold) or ITGB2 (1.7-fold) had a
higher expression in the control samples than in those from animals
treated with OOS. In contrast, the opposite situation was the
olfactory receptor OR51F2 (2.17-fold higher expression in
OOS-treated samples) or YBX1 acceptor (2.12-fold), among others
(Table I). Additionally, several
intracellular pathways were deregulated, including the DNA damage
response pathways, cell cycle regulation, or the PI3K/AKT/mTOR, Wnt
or insulin signaling, all of them involved in cell
proliferation/apoptosis (Fig. 5C).
Moreover, when the GSEA software was used to analyze these
gene-expression data, the p27 pathway (Fig. 5D) and certain pro-apoptotic
pathways (data not shown) were found to be upregulated in the
OOS-treated tumors. In addition, several angiogenesis-related
datasets were downregulated in tumors derived from OOS-treated
animals, as shown in Fig. 5E.
 | Table IIn vivo effect of OOS on gene
expression profiles. |
Table I
In vivo effect of OOS on gene
expression profiles.
| Transcript cluster
ID | Control Avg
signal | OOS Avg signal | Fold change (C vs.
OOS) | ANOVA p-value (C
vs. OOS) | FDR p-value (C vs.
OOS) | Gene symbol | Description |
|---|
|
TC1200010961.hg.1 | 6.89 | 5.71 | 2.26 | 0.026035 | 0.698253 | NDUFA4L2 | NADH dehydrogenase
(ubiquinone) 1 α subcomplex, 4-like 2 |
|
TC2000007083.hg.1 | 4.89 | 3.75 | 2.2 | 0.00829 | 0.698253 | ID1 | Inhibitor of DNA
binding 1, dominant negative helix-loop-helix protein |
|
TC2100007394.hg.1 | 4.84 | 4.07 | 1.7 | 0.022428 | 0.698253 | ITGB2 | Memczak2013
ANTISENSE, CDS, coding, INTERNAL best transcript NM_001127491 |
|
TC1000008663.hg.1 | 4.99 | 4.24 | 1.69 | 0.023254 | 0.698253 | SEMA4G | Sema domain (Dom),
Ig Dom, transmembrane Dom and short cytoplasmic Dom; semaphorin
4G |
|
TC1700012281.hg.1 | 5.62 | 4.88 | 1.68 | 0.028637 | 0.698253 | EPN3 | Epsin 3 |
|
TC0200011898.hg.1 | 7.56 | 6.82 | 1.67 | 0.032135 | 0.698253 | SDC1 | Syndecan 1 |
|
TC0100013266.hg.1 | 4.94 | 4.22 | 1.64 | 0.02815 | 0.698253 | C1orf234 | Chromosome 1 open
reading frame 234 |
|
TC0X00008694.hg.1 | 4.67 | 3.97 | 1.63 | 0.002167 | 0.698253 | HSFX1 | Heat shock
transcription factor family, X-linked 1 |
|
TC0300013665.hg.1 | 4.72 | 4.03 | 1.62 | 0.010431 | 0.698253 | MUC4 | Mucin 4, cell
surface associated |
|
TC0X00006567.hg.1 | 6 | 5.3 | 1.62 | 0.03384 | 0.698253 | VCX3B; VCX | Variable charge,
X-linked 3B; variable charge, X-linked |
|
TC0400006449.hg.1 | 6.39 | 5.7 | 1.61 | 0.033002 | 0.698253 | ZNF141 | Transcript
Identified by AceView, Entrez Gene ID(s) 100288237; 7700 |
|
TC0Y00007078.hg.1 | 5.99 | 5.31 | 1.6 | 0.038427 | 0.698253 | VCY; VCY1B | Variable charge,
Y-linked; variable charge, Y-linked 1B |
|
TC1700010088.hg.1 | 5.62 | 4.94 | 1.6 | 0.0348 | 0.698253 | NATD1 | N-acetyltransferase
domain containing 1 |
|
TC0100006755.hg.1 | 9.2 | 8.54 | 1.59 | 0.013559 | 0.698253 | ENO1 | Memczak2013
ANTISENSE, coding, INTERNAL, UTR3 best transcript NM_001428 |
|
TC0100018271.hg.1 | 4.91 | 4.24 | 1.59 | 0.045745 | 0.698253 | OLFML3 | Olfactomedin like
3 |
|
TC0X00006558.hg.1 | 6.93 | 6.26 | 1.59 | 0.017512 | 0.698253 | VCX; VCX3A | Variable charge,
X-linked; variable charge, X-linked 3A |
|
TC1100011257.hg.1 | 6.5 | 5.83 | 1.59 | 0.030276 | 0.698253 | EFEMP2 | EGF containing
fibulin-like extracellular matrix protein 2 |
|
TC1700009144.hg.1 | 6.03 | 5.37 | 1.58 | 0.043374 | 0.698253 | CHMP6 | Charged
multivesicular body protein 6 |
|
TC1200012664.hg.1 | 4.37 | 3.72 | 1.57 | 0.047618 | 0.698253 | MYRFL | Myelin regulatory
factor-like |
|
TC1900011334.hg.1 | 4.7 | 4.04 | 1.57 | 0.016798 | 0.698253 | ZNF677 | Zinc finger protein
677 |
|
TC0600009871.hg.1 | 5.52 | 4.88 | 1.56 | 0.029687 | 0.698253 | ESR1 | Estrogen receptor
1 |
|
TC1900008366.hg.1 | 6.17 | 5.55 | 1.54 | 0.038319 | 0.698253 | IGFL1 | IGF like family
member 1 |
|
TC1900011772.hg.1 | 6.53 | 5.91 | 1.54 | 0.015269 | 0.698253 | CRX | Cone-rod
homeobox |
|
TC0200010818.hg.1 | 6.34 | 5.73 | 1.53 | 0.016051 | 0.698253 | WNT10A | Wingless-type MMTV
integration site family, member 10A |
|
TC0800012281.hg.1 | 5.44 | 4.84 | 1.52 | 0.004602 | 0.698253 | C8orf58 | Chromosome 8 open
reading frame 58 |
|
TC1900008017.hg.1 | 6.5 | 5.89 | 1.52 | 0.03701 | 0.698253 | RYR1 | Ryanodine receptor
1 (skeletal) |
|
TC0100012921.hg.1 | 7.49 | 6.9 | 1.51 | 0.027124 | 0.698253 | DHRS3; MIR6730 |
Dehydrogenase/reductase (SDR family)
member 3; microRNA 6730 |
|
TC0X00010933.hg.1 | 6.1 | 5.51 | 1.51 | 0.017036 | 0.698253 | ARHGEF6 | Rac/Cdc42 guanine
nucleotide exchange factor 6 |
|
TC1900007020.hg.1 | 6.28 | 5.68 | 1.51 | 0.03582 | 0.698253 | ANGPTL8 | Angiopoietin like
8 |
|
TC1900007384.hg.1 | 6.52 | 5.93 | 1.51 | 0.014667 | 0.698253 | GDF15 | Growth
differentiation factor 15 |
|
TC1900011656.hg.1 | 5.27 | 4.67 | 1.51 | 0.001409 | 0.698253 | STXBP2 | Syntaxin binding
protein 2 |
|
TC0300013151.hg.1 | 5.75 | 6.34 | −1.51 | 0.024943 | 0.698253 | NCEH1 | Neutral cholesterol
ester hydrolase 1 |
|
TC0300013264.hg.1 | 6.75 | 7.34 | −1.51 | 0.007192 | 0.698253 | GNB4 | Guanine nucleotide
binding protein (G protein), β polypeptide 4 |
|
TC1000007895.hg.1 | 5.36 | 5.96 | −1.51 | 0.047298 | 0.698253 | TSPAN15 | Tetraspanin 15 |
|
TC1700011277.hg.1 | 5.68 | 6.27 | −1.51 | 0.005171 | 0.698253 | TRIM37 | Transcript
Identified by AceView, Entrez Gene ID(s) 4591 |
|
TC1200006454.hg.1 | 6.08 | 6.69 | −1.52 | 0.017021 | 0.698253 | WNK1 | Transcript
Identified by AceView, Entrez Gene ID(s) 378465; 65125 |
|
TC0600011507.hg.1 | 6.91 | 7.54 | −1.55 | 0.034697 | 0.698253 | PSMB8 | Proteasome subunit
β8 |
|
TSUnmapped00000243.hg.1 | 6.34 | 6.97 | −1.55 | 0.041704 | 0.698253 | MANSC1 | MANSC domain
containing 1 |
|
TC0100013902.hg.1 | 4.58 | 5.22 | −1.56 | 0.039592 | 0.698253 | SVBP | Small vasohibin
binding protein |
|
TC0500011513.hg.1 | 5.24 | 5.88 | −1.56 | 0.003247 | 0.698253 | PCSK1 | Proprotein
convertase subtilisin/kexin type 1 |
|
TC1900011312.hg.1 | 7.48 | 8.12 | −1.56 | 0.044255 | 0.698253 | ZNF611 | Zinc finger protein
611 |
|
TC0300013520.hg.1 | 5.41 | 6.08 | −1.59 | 0.019529 | 0.698253 | CLDN1 | Claudin 1 |
|
TC0100013223.hg.1 | 5.83 | 6.51 | −1.6 | 0.027675 | 0.698253 | RAP1GAP | RAP1 GTPase
activating protein |
|
TC0X00011080.hg.1 | 4.29 | 4.97 | −1.6 | 0.010211 | 0.698253 | MAGEA9;
MAGEA9B | MAGE family member
A9; MAGE family member A9B |
|
TC0800011566.hg.1 | 5.2 | 5.89 | −1.61 | 0.041099 | 0.698253 | RAD21 | Transcript
Identified by AceView, Entrez Gene ID(s) 5885 |
|
TC1200011870.hg.1 | 5.78 | 6.47 | −1.61 | 0.030776 | 0.698253 | KCTD10 | Potassium channel
tetramerization domain containing 10 |
|
TC0500007145.hg.1 | 4.22 | 4.92 | −1.62 | 0.049614 | 0.698253 | SKP2 | Transcript
Identified by AceView, Entrez Gene ID(s) 6502 |
|
TC1200012736.hg.1 | 3.76 | 4.46 | −1.63 | 0.020998 | 0.698253 | ZNF10 | Zinc finger protein
10 |
|
TC0400008450.hg.1 | 5.23 | 5.94 | −1.64 | 0.035161 | 0.698253 | ANK2 | Ankyrin 2,
neuronal |
|
TC1100006674.hg.1 | 4.4 | 5.13 | −1.65 | 0.037934 | 0.698253 | OR51T1 | Olfactory receptor,
family 51, subfamily T, member 1 |
|
TC1000009092.hg.1 | 4.64 | 5.38 | −1.66 | 0.028207 | 0.698253 | INPP5F | Inositol
polyphosphate-5-phosphatase F |
|
TC1800009244.hg.1 | 4.68 | 5.41 | −1.66 | 0.027752 | 0.698253 | HMSD | Histocompatibility
(minor) serpin domain containing |
|
TC0900009855.hg.1 | 4.81 | 5.56 | −1.69 | 0.015155 | 0.698253 | BAG1 | Bcl-2-associated
athanogene |
|
TC0500009304.hg.1 | 4.84 | 5.62 | −1.71 | 0.04978 | 0.698253 | GABRG2 | γ-aminobutyric acid
(GABA) A receptor, γ 2 |
|
TC1200010538.hg.1 | 5.22 | 6 | −1.72 | 0.037044 | 0.698253 | AMIGO2 | Adhesion molecule
with Ig-like domain 2 |
|
TC1400010625.hg.1 | 4.83 | 5.61 | −1.72 | 0.040343 | 0.698253 | ZNF410 | Zinc finger protein
410 |
|
TC0400008972.hg.1 | 4.53 | 5.33 | −1.74 | 0.04379 | 0.698253 | DCLK2 | Doublecortin-like
kinase 2 |
|
TC0200013298.hg.1 | 5.03 | 5.86 | −1.78 | 0.045777 | 0.698253 | ST3GAL5 | ST3 β-galactoside
α-2.3-sialyltransferase 5 |
|
TC1200009590.hg.1 | 4.5 | 5.34 | −1.79 | 0.011405 | 0.698253 | DCP1B | Decapping mRNA
1B |
|
TC1400007201.hg.1 | 6.39 | 7.23 | −1.79 | 0.02332 | 0.698253 | CDKN3 | Cyclin-dependent
kinase inhibitor 3 |
|
TC1500007644.hg.1 | 4.13 | 4.98 | −1.79 | 0.018087 | 0.698253 | C15orf61 | Chromosome 15 open
reading frame 61 |
|
TC0600008471.hg.1 | 5.3 | 6.16 | −1.81 | 0.011906 | 0.698253 | SDHAF4 | Succinate
dehydrogenase complex assembly factor 4 |
|
TC0700011796.hg.1 | 5.17 | 6.06 | −1.86 | 0.037538 | 0.698253 | SAMD9 | Sterile α motif
domain containing 9 |
|
TC0600009019.hg.1 | 4.77 | 5.68 | −1.87 | 0.00006 | 0.429919 | SOBP | Sine oculis binding
protein homolog |
|
TC0400012945.hg.1 | 5.78 | 6.7 | −1.9 | 0.045226 | 0.698253 | HSD17B11 | Hydroxysteroid
(17-β) dehydrogenase 11 |
|
TC0100008026.hg.1 | 5.32 | 6.4 | −2.12 | 0.021804 | 0.698253 | YBX1 | Zhang2013
ALT_ACCEPTOR, ALT_DONOR, coding, INTERNAL, intronic best
transcript |
|
TC1100006671.hg.1 | 5.76 | 6.88 | −2.17 | 0.021524 | 0.698253 | OR51F2 | Olfactory receptor,
family 51, subfamily F, member 2 |
|
TC0900009779.hg.1 | 5.16 | 6.3 | −2.19 | 0.030103 | 0.698253 | C9orf72 | Chromosome 9 open
reading frame 72 |
Discussion
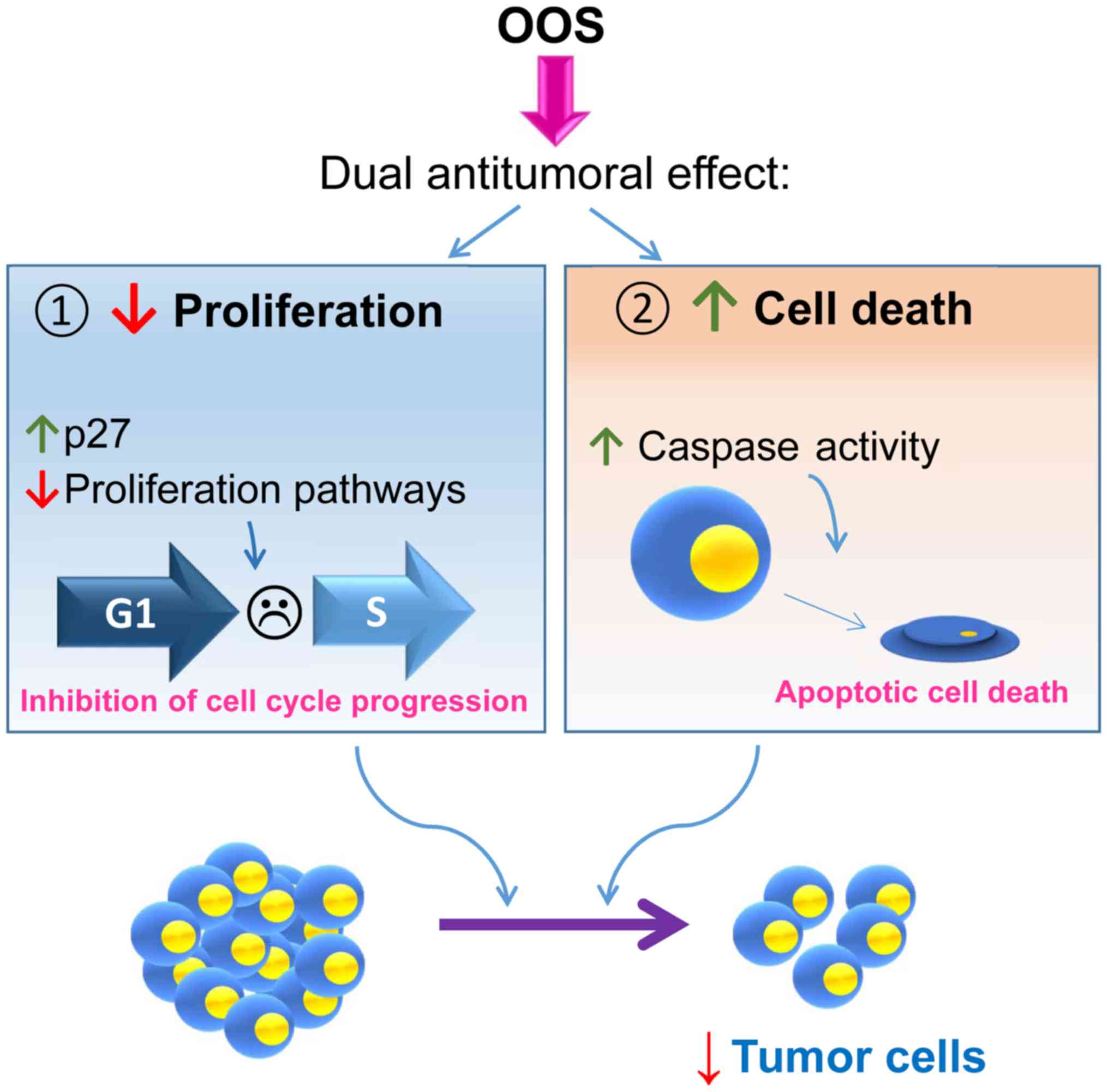
In the present study, we evaluated the antitumoral
effect of OOS in SCLC using both in vitro and in vivo
models. OOS induced a clear decrease in MTT metabolization,
indicative of an antiproliferative/pro-apoptotic effect in the
different SCLC-derived cell lines tested. Such a reduction was both
time and dose dependent. Moreover, OOS reduced in vivo
progression of tumors generated by injection of GLC-8 cells in
mice. OOS treatment slowed down tumor growth without signs of major
toxicity, since the weights of the control and treated mice were
analogous, and no detectable changes in the behavior of the mice
were observed.
To explore the mechanisms leading to such tumor
growth reduction in vivo, several approaches were carried
out. First, a general evaluation of tumor characteristics indicated
that tumors from control or treated mice exhibited large
necrotic/apoptotic areas in the central region of the tumor that is
clearly related to the tumor nature, independently of the
treatment. Moreover, microscopic inspection of the tumors showed
that control tumors seemed to be more vascularized. In fact,
treated tumors appeared to be more compact. The presence of blood
vessels is extremely important for tumor growth, since those tumor
masses larger than a few millimeters in size need these structures
for correct nutrient/oxygen supply (25). To evaluate whether such a decrease
in blood vessels affected critical oncogenic properties such as
survival or proliferation, sections of the tumors were stained with
markers of these biological processes. When tumors were stained
with the proliferation marker Ki-67, a reduction in proliferating
cells after OOS treatment was observed. In addition, a clear
increase in cell death was detected under those same conditions
after staining for apoptotic cell death by TUNEL. These results
point to a combined pro-apoptotic as well as anti-proliferative
action of OOS in the SCLC in vivo models.
These in vivo mechanisms of action were
confirmed in vitro, where OOS induced both cell death and
cell cycle delay. This last action may result from a complex
network that regulates cell proliferation, as suggested by the
deregulation of several pathways that have been linked to the
control of cell proliferation identified in the transcriptomic
analyses (Fig. 6). Within such a
network, one of the actual major players responsible for such
anti-proliferative action appears to be the cell cycle inhibitory
protein p27. These data corroborate previous studies in which p27
seems to be a key intermediate in OOS anti-proliferative action in
other cellular systems such as breast cancer (13), hepatocarcinoma or acute myeloid
leukemia (19,20).
The success of most antitumor therapies is based on
the combination of different agents to increase the antitumor
action of the individual agents. With this aim, OOS was used in
combination with treatments commonly used in the clinic for SCLC,
such as cisplatin, docetaxel or vincristine. When OOS was used
together with these last two compounds, a clear potentiation of the
antitumor properties was observed, opening the possibility of
exploring such effect in patients. This is especially important
since most of the patients suffer from metastatic disease at the
time of diagnosis and, in those cases, only palliative chemotherapy
is offered. For these patients, improvements in the standard of
care treatment by the addition of other agents such as OOS could be
beneficial as well as a very interesting therapeutic option.
Lung cancer represents the most lethal of all
cancers worldwide. The lack of effective treatments, despite recent
advances, calls for efforts to improve its management. OOS, which
includes several vitamins and antioxidants, could have beneficial
effects, always in adequate combination with standard of care
therapies. To date, in this study a clear antitumoral effect of OOS
on SCLC tumorigenesis has been demonstrated. Moreover, the dual
action of the drug that inhibited cell cycle progression and also
stimulated cell death may offer therapeutic benefit to the
classical drugs used in lung cancer therapy. It would also be
valuable to determine whether OOS exerts preventive action on SCLC
using, if possible, genetic models for this disease.
Acknowledgments
This study was partially supported by the Ministry
of Economy and Competitiveness of Spain (BFU2015-71371-R) and
Catalysis S.L. (Madrid, Spain). Our Cancer Research Institute and
the work carried out at our laboratory receive support from the
European Community through the Regional Development Funding Program
(FEDER) and from the CRIS Cancer Foundation.
Competing interests
E.S. is an employee of Catalysis S.L. The research
expenses for this study were partially supported by Catalysis S.L.
(Madrid, Spain).
References
|
1
|
Riaz SP, Lüchtenborg M, Coupland VH,
Spicer J, Peake MD and Møller H: Trends in incidence of small cell
lung cancer and all lung cancer. Lung Cancer. 75:280–284. 2012.
View Article : Google Scholar
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Govindan R, Page N, Morgensztern D, Read
W, Tierney R, Vlahiotis A, Spitznagel EL and Piccirillo J: Changing
epidemiology of small-cell lung cancer in the United States over
the last 30 years: Analysis of the surveillance, epidemiologic, and
end results database. J Clin Oncol. 24:4539–4544. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kahnert K, Kauffmann-Guerrero D and Huber
RM: SCLC-state of the art and what does the future have in store?
Clin Lung Cancer. 17:325–333. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lamson DW and Brignall MS: Antioxidants in
cancer therapy; their actions and interactions with oncologic
therapies. Altern Med Rev. 4:304–329. 1999.PubMed/NCBI
|
|
6
|
Yang CS, Chung JY, Yang G, Chhabra SK and
Lee MJ: Tea and tea polyphenols in cancer prevention. J Nutr.
130(Suppl 2): 472S–478S. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fujiki H: Green tea: Health benefits as
cancer preventive for humans. Chem Rec. 5:119–132. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Clark J and You M: Chemoprevention of lung
cancer by tea. Mol Nutr Food Res. 50:144–151. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Khan N and Mukhtar H: Dietary agents for
prevention and treatment of lung cancer. Cancer Lett. 359:155–164.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liang W, Binns CW, Jian L and Lee AH: Does
the consumption of green tea reduce the risk of lung cancer among
smokers? Evid Based Complement Alternat Med. 4:17–22. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lu Y, Yao R, Yan Y, Wang Y, Hara Y, Lubet
RA and You M: A gene expression signature that can predict green
tea exposure and chemopreventive efficacy of lung cancer in mice.
Cancer Res. 66:1956–1963. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sadava D, Whitlock E and Kane SE: The
green tea polyphenol, epigallocatechin-3-gallate inhibits
telomerase and induces apoptosis in drug-resistant lung cancer
cells. Biochem Biophys Res Commun. 360:233–237. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hernández-García S, González V, Sanz E and
Pandiella A: Effect of oncoxin oral solution in HER2-overexpressing
breast cancer. Nutr Cancer. 67:1159–1169. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hollman PC, Feskens EJ and Katan MB: Tea
flavonols in cardiovascular disease and cancer epidemiology. Proc
Soc Exp Biol Med. 220:198–202. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu L, Hudgins WR, Shack S, Yin MQ and
Samid D: Cinnamic acid: A natural product with potential use in
cancer intervention. Int J Cancer. 62:345–350. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gomez EV, Perez YM, Sanchez HV, Forment
GR, Soler EA, Bertot LC, García AY, del Rosario Abreu Vazquez M and
Fabian LG: Antioxidant and immunomodulatory effects of Viusid in
patients with chronic hepatitis C. World J Gastroenterol.
16:2638–2647. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Al-Mahtab M, Akbar SM, Khan MS and Rahman
S: Increased survival of patients with end-stage hepatocellular
carcinoma due to intake of ONCOXIN®, a dietary
supplement. Indian J Cancer. 52:443–446. 2015. View Article : Google Scholar
|
|
18
|
Dayem Uddin MAI, Mahmood I, Ghosha AK and
Khatuns RA: Findings of the 3-month supportive treatment with
ocoxin solution beside the standard modalities of patients with
different neoplastic diseases. TAJ. 22:172–175. 2009.
|
|
19
|
Díaz-Rodríguez E, Hernández-García S, Sanz
E and Pandiella A: Antitumoral effect of ocoxin on acute myeloid
leukemia. Oncotarget. 7:6231–6242. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Díaz-Rodríguez E, El-Mallah AM, Sanz E and
Pandiella A: Antitumoral effect of Ocoxin in hepatocellular
carcinoma. Oncol Lett. 14:1950–1958. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huqun EY, Endo Y, Xin H, Takahashi M,
Nukiwa T and Hagiwara K: A naturally occurring p73 mutation in a
p73-p53 double-mutant lung cancer cell line encodes p73 alpha
protein with a dominant-negative function. Cancer Sci. 94:718–724.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Postmus PE, de Ley L, van der Veen AY,
Mesander G, Buys CH and Elema JD: Two small cell lung cancer cell
lines established from rigid bronchoscope biopsies. Eur J Cancer
Clin Oncol. 24:753–763. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sher E, Pandiella A and Clementi F:
Voltage-operated calcium channels in small cell lung carcinoma cell
lines: Pharmacological, functional, and immunological properties.
Cancer Res. 50:3892–3896. 1990.PubMed/NCBI
|
|
24
|
Cabrera N, Díaz-Rodríguez E, Becker E,
Martín-Zanca D and Pandiella A: TrkA receptor ectodomain cleavage
generates a tyrosine-phosphorylated cell-associated fragment. J
Cell Biol. 132:427–436. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Folkman J: Angiogenesis in cancer,
vascular, rheumatoid and other disease. Nat Med. 1:27–31. 1995.
View Article : Google Scholar : PubMed/NCBI
|