Introduction
Liver cancer is a prevalent malignancy worldwide and
ranks as one of the leading causes of cancer-associated mortality
(1). The mechanism of liver cancer
is complicated and heterogeneous, and is accompanied by various
molecular abnormalities. It is necessary to identify novel
oncogenes and tumor suppressors for further investigation and
potential clinical application.
The microrchidia (MORC) family proteins are
conserved proteins with important roles in multiple biological
processes. MORC2, also known as ZCWCC1, ZCW3, KIAA0852 and
AC004542.C22.1, is a member of this family (2,3). It
contains an ATPase domain, a zinc finger type CW domain, and
nuclear localization signal and coiled-coil domains (2–4). It
has been reported that MORC2 can bind with histone deacetylase
(HDAC)4 and functions as a transcriptional repressor by mediating
the deacetylation of histone H3 (5). A limited number of studies have shown
that MORC2 functions in chromatin remodeling, facilitating DNA
damage repair and promoting lipogenesis (2,6,7),
however, its function in cancer remains to be fully elucidated. As
abnormal chromatin dynamics, enhancing DNA damage repair ability
and de novo lipogenesis are crucial events in cancer cells,
MORC2 may function as an oncogene by promoting the malignant
phenotype of cancer cells. MORC2 can promote the migration and
invasion of breast cancer cells, and is involved in a prognostic
prediction model for breast cancer containing six genes (8,9). Its
oncogenic role in gastric cancer has also been demonstrated
(10–12). For example, it has been reported
that MORC2 downregulates p21 by recruiting HDAC1 to the p21
promoter, in a p53-independent manner in gastric cancer; the
phosphorylation of MORC2 increases the expression of cyclin
D1-cyclin-dependent kinase (CDK)4 and cyclin D3-CDK6 complexes,
promotes gastric cell cycle transition from the G1 to S stage, and
indicates a poorer prognosis in patients with gastric cancer
(11,12).
However, to date, no studies have reported on the
clinicopathologic significance and functions of MORC2 in liver
cancer. The present study presented the first evidence, to the best
of our knowledge, of the expression pattern of MORC2 in human liver
cancer and its clinical significance. The roles of MORC2 in the
progression of liver cancer and its underlying mechanisms were
investigated. The data demonstrated that MORC2 was upregulated in
liver cancer, and contributed to the proliferation, metastasis and
chemoresistance of liver cancer cells via the p53 and Hippo
pathways.
Materials and methods
Cell culture, culture conditions and
antibodies
The HepG2, Bel-7402, Huh7, PLC/PRF-5, SMMC7721 and
LM3 liver cancer cell lines were obtained from the Cell Bank of the
Chinese Academy of Sciences Committee Type Culture Collection
(Shanghai, China), and the normal L02 liver cell line was conserved
at the Central Laboratory of Renmin Hospital of Wuhan University
(Wuhan, China). The cells were cultured in DMEM supplemented with
10% fetal bovine serum (FBS) (Zhejiang Tianhang Biotechnology Co.,
Ltd., Hangzhou, China) and 100 units penicillin/streptomycin. The
cells were cultured at 37°C and 5% CO2 in a humidified
chamber. Rabbit polyclonal anti-MORC2 antibody was purchased from
Abcam (Cambridge, UK). Mouse monoclonal anti-β-actin antibody was
purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
Anti-rabbit and anti-mouse horseradish peroxidase (HRP)-conjugated
secondary antibodies were purchased from Promega Corporation
(Madison, WI, USA).
Patients and histological and
immunohistochemical (IHC) staining
The GSE14520 and GSE22058 mRNA expression profile
were downloaded from the Gene Expression Omnibus (GEO) database
(13–15). The Cancer Genome Atlas (TCGA) copy
number-altered genome data for each patient was directly downloaded
from cBioPortal for Cancer Genomics (16,17).
All liver cancer samples and paired adjacent tissues were retrieved
from patients receiving surgery between December 1 and December 31,
2014, from the Department of Pathology, Zhongnan Hospital of Wuhan
University (Wuhan, China). All patients provided informed written
consent prior to the investigation. The inclusion of human samples
was approved by the Ethics Review Board of the Second People's
Hospital of Guangdong Province (Guangdong, China; approval no.
2015-KYLL-023). The tissues were first stained with hematoxylin and
eosin for histological examination. The deparaffinized sections
were treated with 3% H2O2 and subjected to
antigen retrieval by citric acid (pH 6.0). Following overnight
incubation with primary antibody (anti-MORC2 antibody; 1:200) at
4°C, the sections were incubated for 30 min at room temperature
with HRP-labeled polymer conjugated with secondary antibody
(MaxVision™ kits) and incubated for 1 min with diaminobenzidine.
The sections were then lightly counterstained with hematoxylin.
Sections without primary antibody served as negative controls. The
expression level of MORC2 was ascertained according to the average
score of two pathologists' evaluations using a CKX41 microscope
(Olympus Corporation, Tokyo, Japan). As MORC2 is mainly expressed
in the nucleus, the positive nuclear staining of MORC2 was used to
elucidate its expression level according to the following formula:
Immunostaining score = percentage score × intensity score, where
the percentage score represented the percentage of immunopositive
cells, and was graded as 0 (<6%), 1 (6–33%), 2 (34–66%) and 3
(>66%). The intensity score represented the intensity of
immunostaining, and was determined as 0 (absent), 1 (weak
staining), 2 (moderate staining) and 3 (strong staining). All cases
were diagnosed by two certificated pathologists without
discrepancy.
Small interfering RNA (siRNA)
transfection and establishment of stable expressing cells
The siRNAs were designed and purchased commercially
(Genepharma, Shanghai, China) as shown in Table I. The cells were transfected with
50 ng/µl targeting siRNA using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) for
48 h, following which the depletion efficiency was analyzed by
western blot analysis. siRNA-homo-1760 was selected for the
following experiments, as it exhibited the optimal efficiency. For
the establishment of stable MORC2-overexpressing cells, the
overexpression plasmid was constructed using the pLV-EGFP (2A) puro
plasmid; the plasmid was packed using a lentivirus system (both
from Inovogen Biotechnology Co., Ltd., Beijing, China). Following
infection, the cells were selected for 4 weeks using DMEM with 10
µg/ml puromycin (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany). The expression of MORC2 was lowest in the PLC cell line,
however, the overexpression efficiency was not satisfactory in this
cell line, thus the SMMC7721 cell line was used to construct the
stable MORC2-overexpressing cells in the present study.
 | Table IsiRNA sequences used for the
overexpression of MORC2. |
Table I
siRNA sequences used for the
overexpression of MORC2.
| siRNA | Sense (5′-3′) | Antisense
(5′-3′) |
|---|
|
MORC2-homo-1760 |
GCGGAACAUUGGUGAUCAUTT |
AUGAUCACCAAUGUUCCGCTT |
|
MORC2-homo-2439 |
GGAGCCUACACACAACAAATT |
UUUGUUGUGUGUAGGCUCCTT |
|
MORC2-homo-3972 |
GCAGCUGAGUGCUAUGAAUTT |
AUUCAUAGCACUCAGCUGCTT |
| Control siRNA |
UUCUCCGAACGUGUCACGUTT |
ACGUGACACGUUCGGAGAATT |
Western blot analysis
In brief, cells were lysed in lysis buffer
containing 50 mmol/l Tris (pH 8.0), 150 mmol/l NaCl, 1% NP-40, 0.5%
sodium deoxycholate, 0.1% SDS, 50 mmol/l NaF, 1 mmol/l
Na3VO4. Protein concentration was analyzed
and adjusted using a bicinchoninic acid protein assay.
Subsequently, equal quantities (50 µg) of samples were
separated by 10% SDS-PAGE (Bio-Rad Laboratories, Inc., Hercules,
CA, USA) and transferred onto nitrocellulose membranes (EMD
Millipore, Billerica, MA, USA). After blocking with 5% non-fat milk
in Tris-buffered saline containing 0.05% Tween-20, the membranes
were incubated with primary antibodies against MORC2 (1:400,
bs-0354R; BIOSS, Beijing, China), cytochrome c (1:200,
ab53056), caspase-3 (1:500, ab47131) (both from Abcam), caspase-9
(1:150, sc-56076; Santa Cruz Biotechnology, Inc.), poly
(ADP-ribose) polymerase (PARP) (1:150, ab4830; Abcam), p53 (1:500,
sc-6243), B-cell lymphoma-2 (Bcl-2) (1:500, sc-783),
Bcl-2-associated X protein (Bax) (1:300, sc-493) (all from Santa
Cruz Biotechnology, Inc.), p53 upregulated modulator of apoptosis
(PUMA)α (1:5000, ab33906; Abcam), Yes-associated protein 1 (YAP1)
(1:500, orb89757; Biorbyt Ltd., Cambridge, UK), phosphorylated
(p-)S127-YAP1 (1:10,000, ab76252; Abcam), Transcriptional
co-activator with PDZ-binding motif (TAZ) (1:500, #23306-1-AP;
ProteinTech Group, Inc., Chicago, IL, USA), p-S89-TAZ (1:500,
sc-17610), and β-actin (1:1,000, sc-47778) (both from Santa Cruz
Biotechnology, Inc.) at 4°C overnight. The primary antibodies
coupled to sample proteins were then visualized by incubation for 1
h at 37°C with HRP-conjugated secondary antibodies (1:1,000,
074-1506 and 074-1806; KPL, Inc., Gaithersburg, MD, USA) using a
chemiluminescence detection system (EMD Millipore) according to the
manufacturer's protocol.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA from the tissues and cells was isolated
using TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.).
Subsequently, 2 µg of total RNA from each sample was used
for cDNA synthesis. RT-qPCR analysis was performed in triplicate
with the SYBR® Green PCR Master mix (Takara Bio, Inc.,
Otsu, Japan) and β-actin was used as an internal control according
to the manufacturer's protocol. Each sample included: 4 µl
cDNA, 2 µl primers, 12.5 µl 2X SYBR Green master mix
and 6.5 µl ddH2O. The PCR thermo-cycling
conditions were as follows: 5 min at 94°C, followed by 40 cycles at
94°C for 20 sec, 60°C for 20 sec and at 72°C for 20 sec, and 5 min
at 72°C. Relative expression levels of target genes were determined
according to the 2−ΔΔCq method (18), where Cq represents the
quantification cycle for each transcript. The primer pairs for
human MORC2 and β-actin were designed as shown in Table II.
| Table IIPrimers used for human MORC2 and
β-actin. |
Table II
Primers used for human MORC2 and
β-actin.
| Primer | Sequence
(5′-3′) |
|---|
| Homo-MORC2-F |
GAAAGCCTGCCAACACTCTC |
| Homo-MORC2-R |
CTCATCAGAAACTGCGACA |
| Homo-β-actin-F |
CATTAAGGAGAAGCTGTGCT |
| Homo-β-actin-R |
GTTGAAGGTAGTTTCGTGGA |
Cell counting kit-8 (CCK-8) assay
In the cell proliferation assay, the cells
(1×105/well) were plated onto 96-well plates on the
first day and allowed to attach overnight. The following day, 10
µl of CCK-8 (Dojindo Molecular Technologies, Inc., Kumamoto,
Japan) was added to each well and incubated at 37°C and 5%
CO2 for 1 h between days 1 and 5. The absorbance was
then detected with the multifunctional microplate reader at 490 nm.
For the chemoresistance assay, following seeding of the cells
(1×105/well) in the plates, the cells were treated with
the chemotherapeutics, doxorubicin, 5-fluorouracil and cisplatin,
at different concentrations (0, 1, 5, 10, 20, 40, 80 and 100
µM) for 72 h at 37°C, following which CCK-8 was added.
Following incubation at 37°C and 5% CO2 for 1 h, the
absorbance was detected and the half maximal inhibitory
concentration (IC50) parameter was calculated.
Plate colony formation assay
The cells were digested with trypsin, centrifuged at
110 × g for 5 min at 37°C, resuspended in DMEM supplemented with
10% FBS and seeded in 6-well plates (1,000 cells/well), following
which the cells in each group were cultured for 2 weeks. The cells
were then washed twice with PBS, fixed with 4% paraformaldehyde for
20 min, and then stained with 0.1% crystal violet for 30 min. The
dishes were then carefully washed with PBS until the background was
clear. Finally, the number of colonies was counted under a
microscope (CKX41; Olympus Corporation).
Cell-cycle analysis
In brief, the cells were digested with trypsin,
centrifuged at 110 × g for 5 min at 37°C, resuspended in PBS,
washed twice with PBS and then fixed in 100% ice-cold methanol
overnight at -20°C. The cells were then incubated with 50 mg/ml
propidium iodide (PI) and 1 mg/ml RNAase in PBS for 20 min,
following which the samples were analyzed with BD FACSAria (BD
Biosciences, Franklin Lakes, NJ, USA).
In vitro migration and invasion
assays
In the scratch wound healing assay, cells were
cultured in serum-free medium for 24 h and wounded with pipette
tips. Subsequently, the medium was replaced with fresh medium. The
wound healing procedure was observed after 48 h, and images of the
cells were captured under a microscope (CKX41; Olympus
Corporation). Cell migration and invasion assays were performed
using Transwell chambers, as previously described (19). The cells were harvested and
resuspended in serum-free medium, and then added to the upper
chamber. Following incubation for 48 h, cells remaining on the
upper side of membrane were removed with a cotton swab. The cells
migrated to the lower membrane surface were fixed and stained with
0.1% crystal violet for 30 min, and the number of cells was counted
under a microscope. To assess invasion ability, the membranes were
pre-coated with diluted Matrigel, whereas the membranes in
migration experiments were not pre-coated with Matrigel.
Nude mice experiments
All animal experiments were approved by the Ethics
Review Board of the Second People's Hospital of Guangdong Province
(approval no. 2015-KYLL-063). For the experiments, 4-week-old male
BALB/C nude mice (Hangzhou Hibio Technology Co., Ltd., Hangzhou,
China) were used. The mice were maintained under the following
pathogen-free conditions: 60% humidity; room temperature; 12-h
light/dark cycle; ad libitum access to food and water. In a
subcutaneous xenograft procedure, 1×106 cells
(SMMC7721MORC2 or SMMC7721Vector) were
resuspended in PBS solution and then injected subcutaneously into
the left and right side of each of the mice (n=6). The tumor size
was measured every 3 days, and tumor volume was calculated using
the following formula: 1/2 length × width2. For the
assessment of pulmonary metastasis, 10 mice were included in each
group, and 1×106 cells (SMMC7721MORC2 or
SMMC7721Vector) were injected into the caudal vena.
After 2 weeks, the mice were sacrificed and lung colonization was
quantified by pathological examination.
Analysis of apoptosis
Annexin V-FITC/PI staining was used to investigate
whether MORC2 regulates the apoptosis of liver cancer cells. The
cells were seeded into 6-well plates. When the cells in each group
were at a log phase of growth, the cells were digested with
trypsin, centrifuged at 110 × g for 5 min at 37°C and resuspended
in PBS. The cells were then incubated with an ApoScreen Annexin V
Apoptosis kit and PI. Every sample containing 10,000 cells was
analyzed using BD FACSAria (BD Biosciences). The experiments were
performed in triplicate.
Statistical analysis
Statistical significances between values of
different experimental groups were analyzed using Student's t-test
or one-way analysis of variance. A χ2 test and Fisher's
test were used in analyzing enumeration data and pathway analysis.
P<0.05 was considered to indicate a statistically significant
difference. All statistical analyses were conducted using GraphPad
Prism 6.0 (GraphPad Software, Inc., La Jolla, CA, USA) or SPSS for
Windows 17.0.1 software (SPSS, Inc., Chicago, IL, USA). Expression
profile analysis was conducted on L02MORC2 and
L02Vector data (Chen et al, unpublished data)
using GCBI 1.0 software (Gminix Technology Co., Ltd., Shanghai,
China) with the Kyoto Encyclopedia of Genes and Genomes database
(http://www.genome.jp/kegg/) as
reference.
Results
MORC2 is overexpressed in liver cancer
samples at the mRNA and protein levels
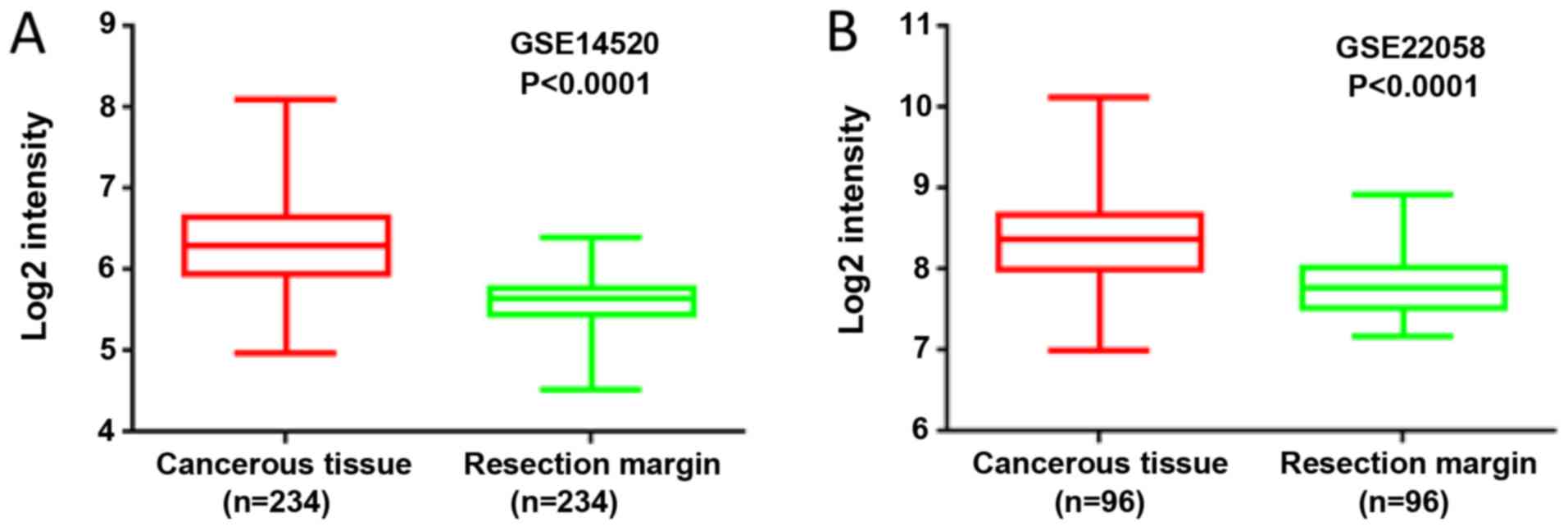
By analyzing the expression profile of liver cancer
samples from GEO datasets GSE14520 and GSE22058, it was found that
the mRNA levels of MORC2 were significantly upregulated in the
liver cancer tissues, compared with the matched non-tumorous
tissues (Fig. 1). The expression
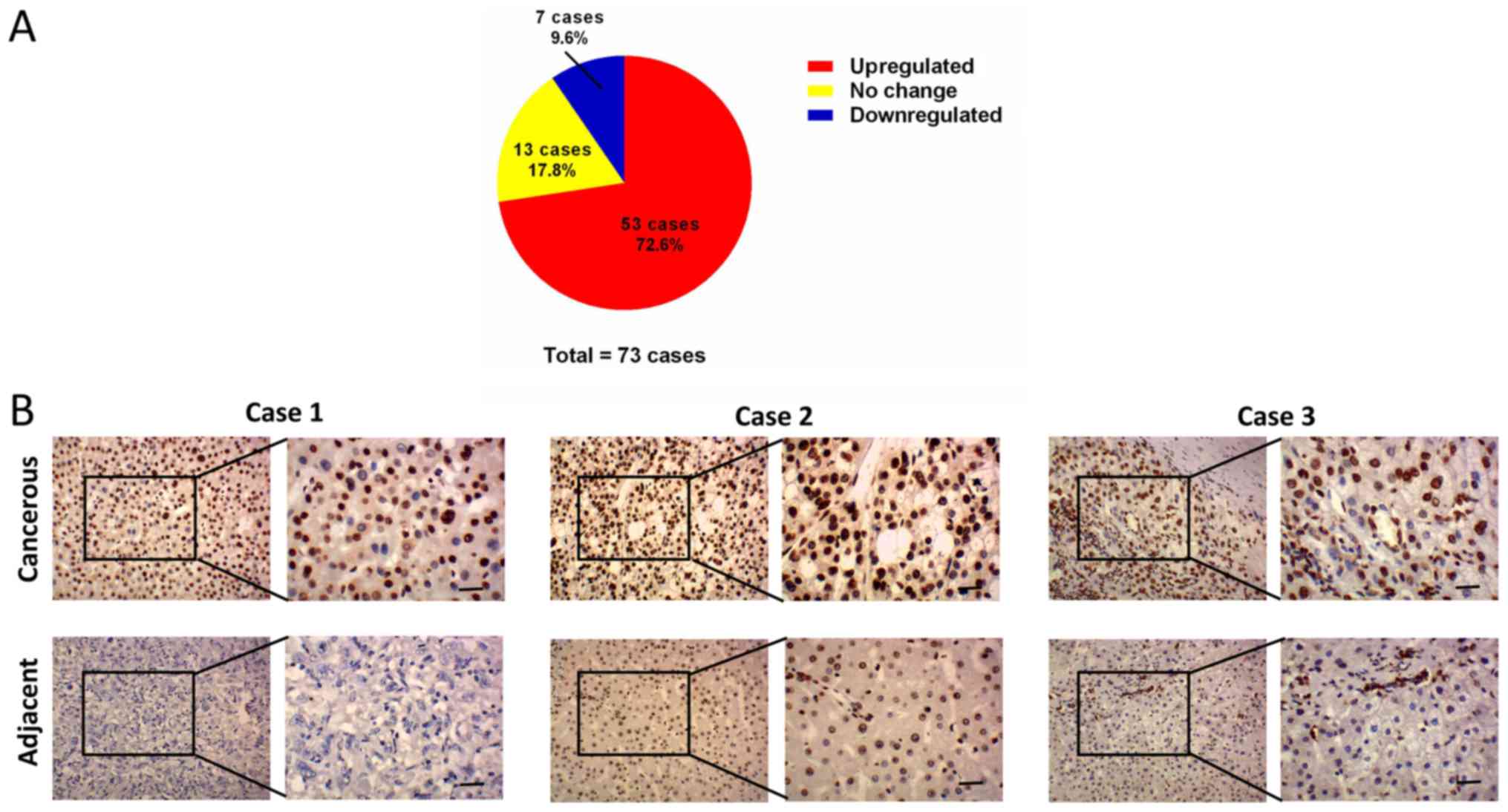
levels of MORC2 in 73 liver cancer and corresponding adjacent liver
tissues specimens were then examined using IHC staining. Consistent
with previous reports, MORC2 was expressed mainly in the nucleus
and was relatively weak in the cytoplasm (Fig. 2) (3,10).
The results showed that MORC2 was overexpressed in the majority
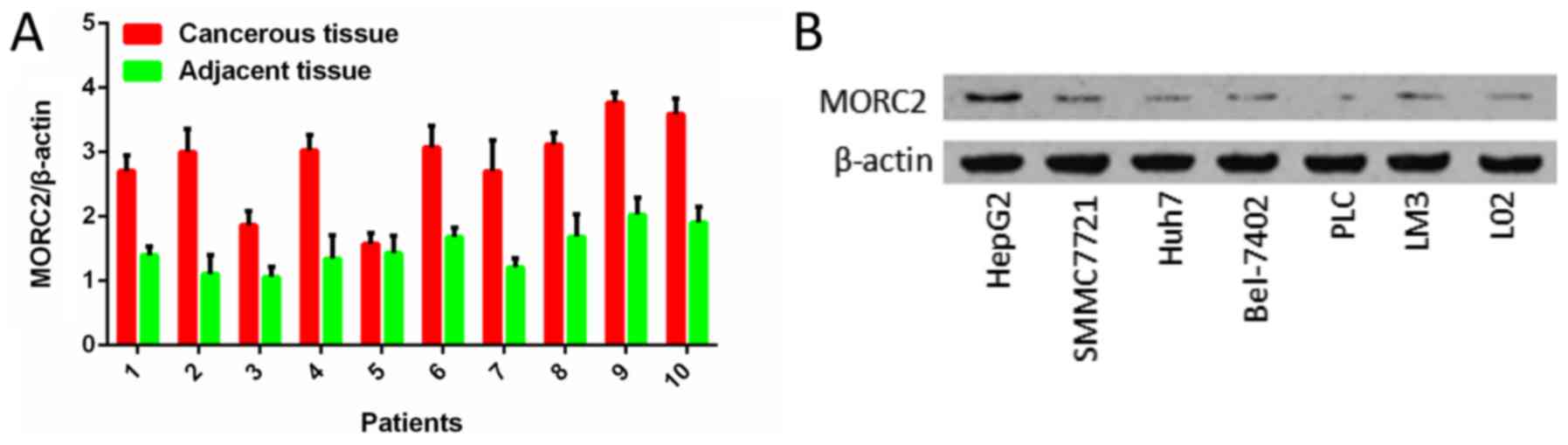
(72.6%, 53/73) of the liver cancer samples (Fig. 2). RT-qPCR analysis was also used to
examine the levels of MORC2 in 10 pairs of liver cancer and
pair-adjacent liver tissue specimens; accordingly, the expression
of MORC2 was upregulated in almost all the cancerous tissues
(Fig. 3A). The expression level of
MORC2 was then examined in HepG2, Bel-7402, Huh7, PLC/PRF-5,
SMMC7721, LM3 and L02 cells. Higher expression of MORC2 was
observed in the majority of the liver cancer cells, compared with
the L02 cells (Fig. 3B). From
these results, it was concluded that MORC2 was overexpressed in
liver cancer at the mRNA and protein level, demonstrating the
important role of MORC2 in the pathogenesis of liver cancer.
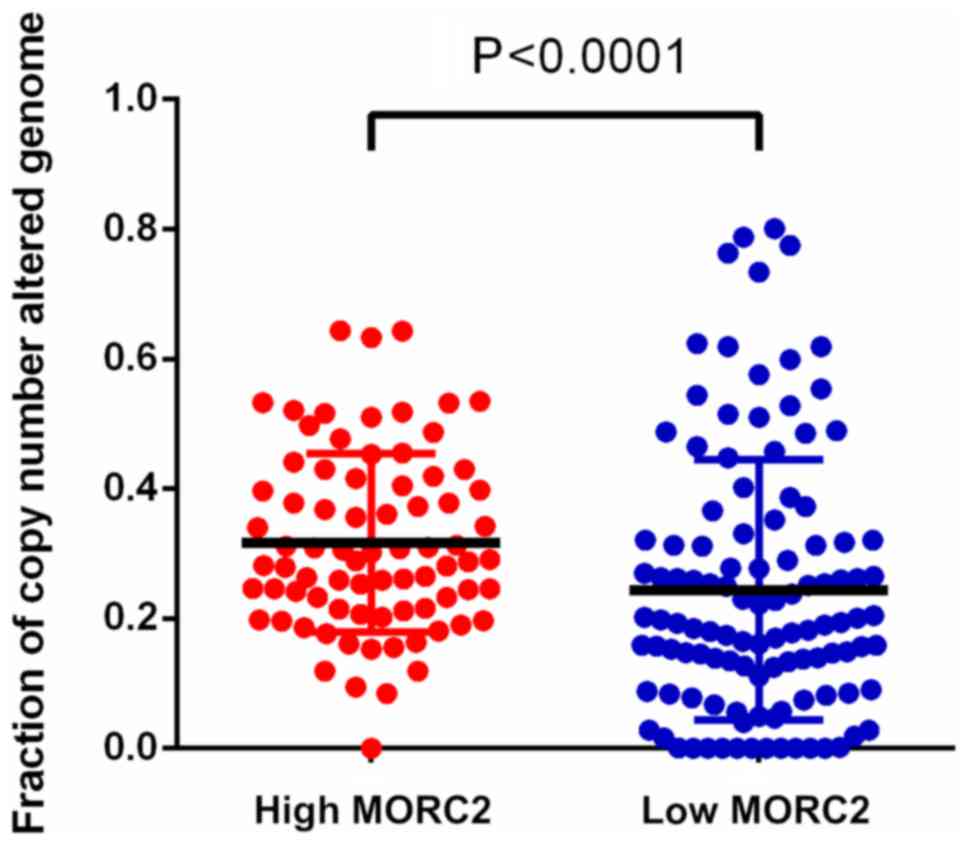
High expression of MORC2 leads to
accumulation of copy number variations and unfavorable pathological
characteristics
Dysfunctional DNA damage repair results in genetic
alterations, including somatic copy number alteration, and this
characteristic is increasingly recognized as common feature of
human liver cancer (20). It has
been reported that MORC2 is associated with chromatin remodeling,
promotes the induction of γ-H2AX and modulates DNA damage repair
(6). Utilizing the data of TCGA
database (16,17), it was detected that the fraction of
copy number alterations was higher in the genome of patients with
liver cancer and higher MORC2 (Fig.
4). This result indicated that the upregulation of MORC2 may
result in liver cancer progression via accumulating extra copies of
DNA.
Using the clinical information of GSE14520, the
correlation between the expression of MORC2 and clinicopathological
features was assessed to determine its clinical significance. The
samples were divided into two groups according to the expression
level and were analyzed with χ2 test. As shown in
Table III, a higher expression
of MORC2 was associated with larger tumor volume (P=0.009) and
higher American Joint Committee on Cancer T stage (P=0.007). These
results indicated that MORC2 may be a potential prognostic
biomarker in liver cancer.
| Table IIIAssociations between the expression
of MORC2 and the clinicopathologic features of liver cancer
(GSE14520). |
Table III
Associations between the expression
of MORC2 and the clinicopathologic features of liver cancer
(GSE14520).
| Characteristic | Patients (n) | Expression
ofmicrorchidia 2
| P-value |
|---|
| High | Low |
|---|
| Age (years) | | | | |
| ≤55 | 166 | 88 | 77 | 0.166 |
| >55 | 76 | 33 | 43 | |
| Sex | | | | |
| Male | 211 | 108 | 103 | 0.336 |
| Female | 31 | 13 | 18 | |
| AFP (ng/ml) | | | | |
| ≤200 | 128 | 58 | 70 | 0.119 |
| >200 | 110 | 61 | 49 | |
| ALT (U/l) | | | | |
| ≤50 | 142 | 64 | 78 | 0.068 |
| >50 | 100 | 57 | 43 | |
| Cirrhosis | | | | |
| Yes | 223 | 112 | 111 | 0.811 |
| No | 19 | 9 | 10 | |
| Tumor size
(d/cm) | | | | |
| <5 | 153 | 67 | 86 | 0.009 |
| ≥5 | 88 | 54 | 34 | |
| Tumor number | | | | |
| Solitary | 190 | 91 | 99 | 0.211 |
| Multiple | 52 | 30 | 22 | |
| AJCC T stage | | | | |
| T1 | 96 | 34 | 62 | 0.007 |
| T2 | 78 | 43 | 35 | |
| T3 | 51 | 30 | 21 | |
| BCLC stage | | | | |
| 0 | 20 | 8 | 12 | 0.283 |
| A | 152 | 68 | 84 | |
| B | 24 | 13 | 11 | |
| C | 29 | 18 | 11 | |
| CLIP stage | | | | |
| 0 | 98 | 41 | 57 | 0.297 |
| 1 | 79 | 40 | 39 | |
| 2,3,4,5 | 48 | 26 | 22 | |
| PRMS
classification | | | | |
| High | 121 | 74 | 47 | <0.001 |
| Low | 121 | 47 | 74 | |
In liver cancer cells, knockdown of MORC2
inhibits proliferation in vitro
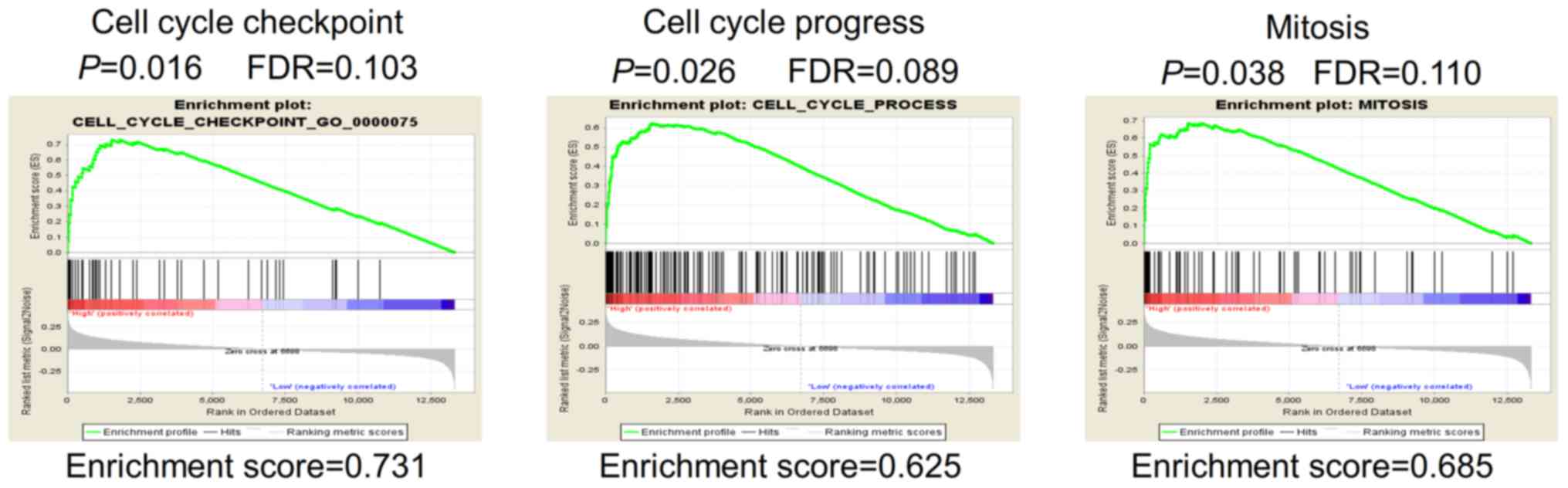
Gene set enrichment analysis (GSEA) was performed in
order to identify potential genes modulated by MORC2. The mRNA
expression profiling data in GSE14520 were used to outline
MORC2-correlated genes. In the GSEA analysis of Gene Ontology
terms, it showed that gene sets involved in 'cell cycle process'
and 'mitosis' were enriched in the MORC2-high expression samples
(Fig. 5). Enhanced mitogenic
signaling and aberrant process of cell cycle are essential for cell
proliferation and cancer progression (20). The bioinformatics data provided a
possible explanation of why a high expression of MORC2 was
associated with unfavorable clinicopathological features, including
tumor size and T stage (Table
III). The results suggested that MORC2 has an important
function in the proliferation of liver cancer cells.
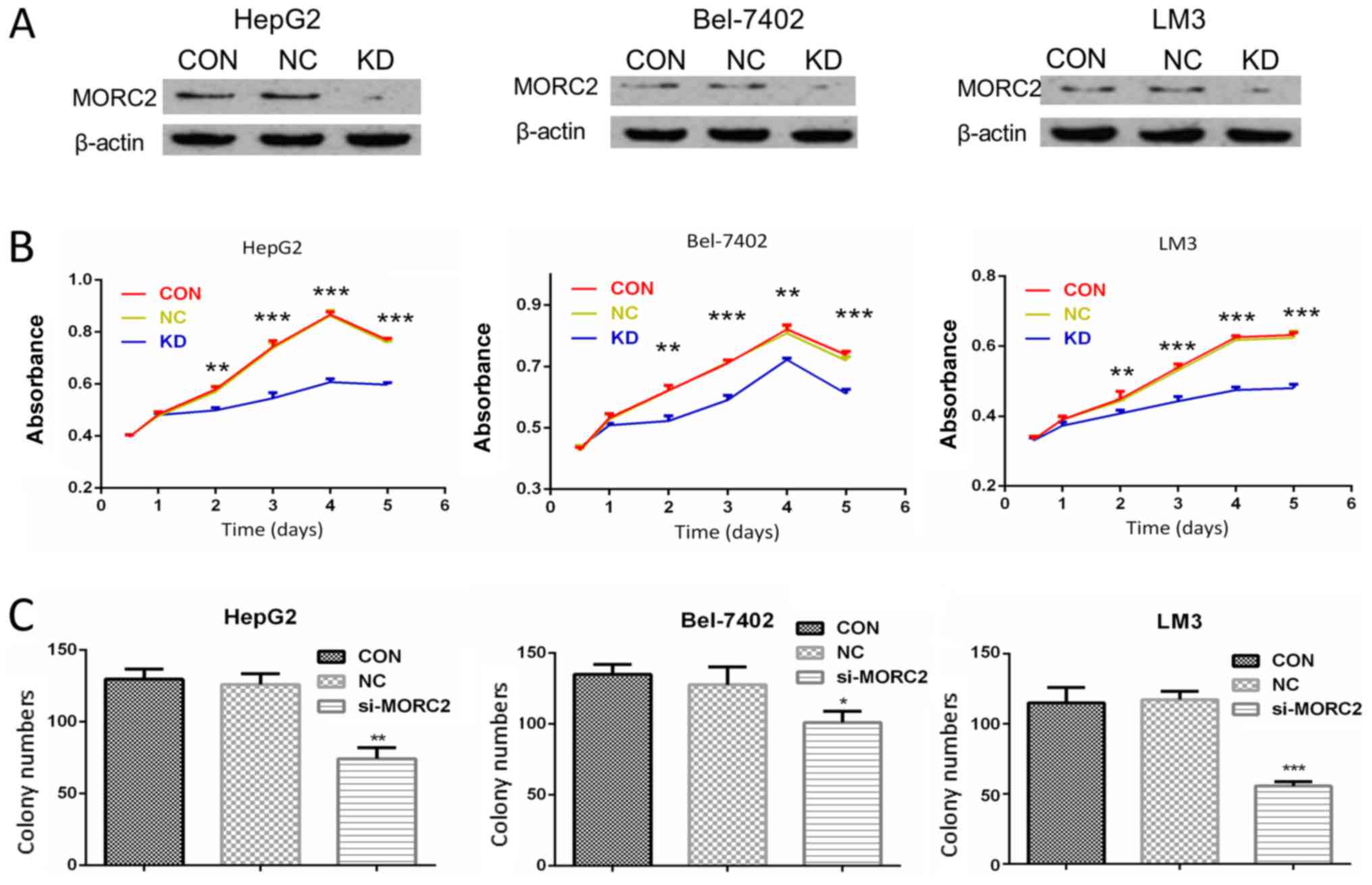
To verify the biological role of MORC2 in the
proliferation of liver cancer, MORC2 was knocked down using siRNAs
in HepG2, Bel-7402 and LM3 cells, which had a higher expression of
MORC2. Western blot analysis was used to determine the knockdown
efficiency (Fig. 6A).
The effect of MORC2 on cell proliferation was then
examined. As shown in Fig. 6B,
compared with the control groups, following MORC2 knockdown, the
HepG2, Bel-7402 and LM3 cells exhibited a significantly lower cell
proliferation rate (Fig. 6B).
Furthermore, cell proliferation was measured using a plate colony
formation assay. Compared with the control cells, MORC2 knockdown
in the HepG2, Bel-7402 and LM3 cells resulted in markedly decreased
colony formation abilities (Fig.
6C).
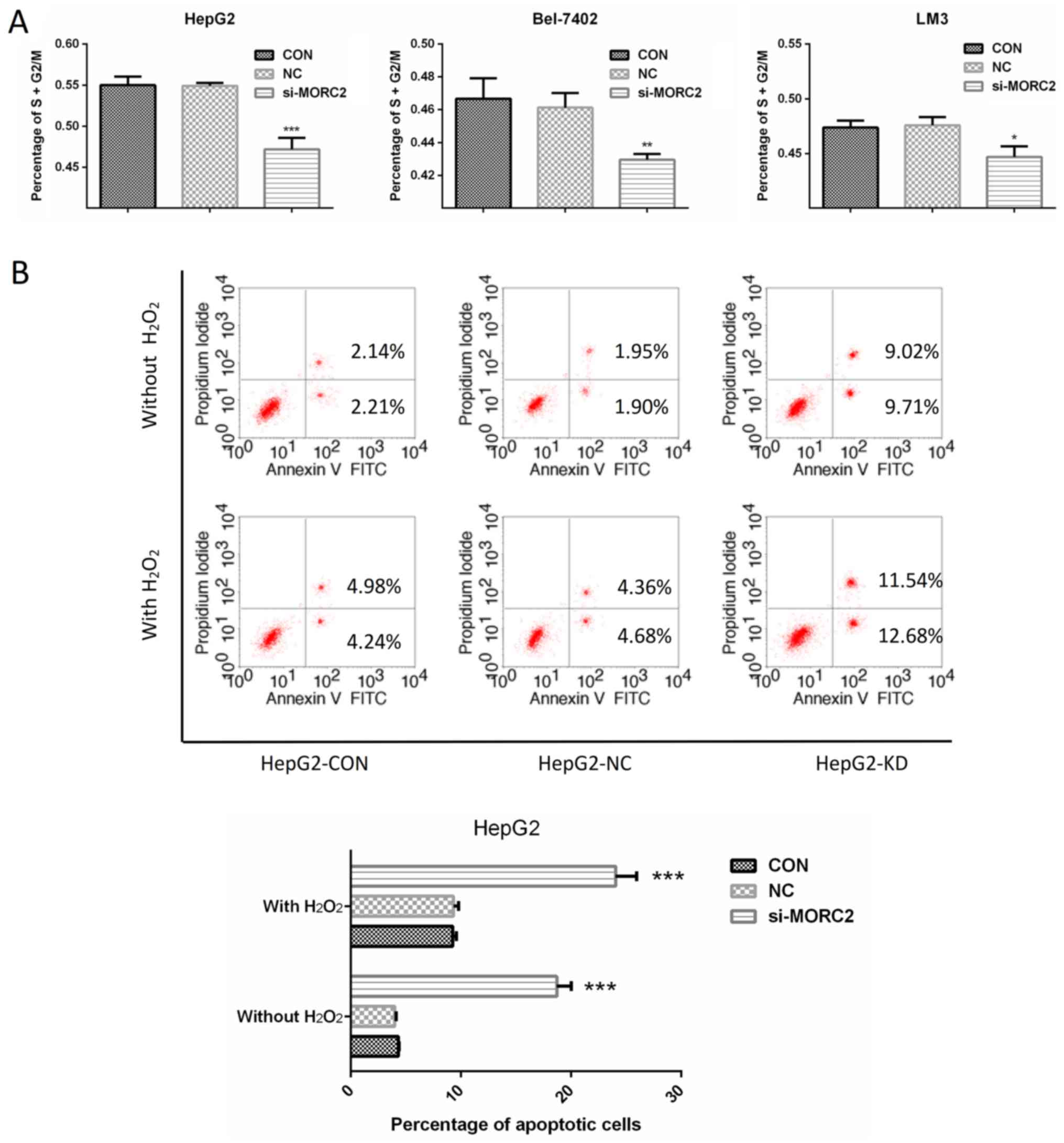
In liver cancer cells, inhibition of
MORC2 promotes cell cycle arrest and induces apoptosis
Enhanced cell cycle progression and reduced
apoptotic signaling are two important factors causing sustained
proliferation. To examine the factors involved in MORC2 regulating
the proliferation of liver cancer cells, the cell cycle was
detected in liver cancer cells through flow cytometry. Following
transfection with si-MORC2 or si-NC for 48 h, the inhibition of
MORC2 led to a significant accumulation of cells at the G0/G1-phase
and a marked decrease in cells at the S/G2/M-phase in HepG2,
Bel-7402 and LM3 cells (Fig. 7A).
Subsequently, the effects of MORC2 on apoptosis under normal
condition or oxidative stress (treated with
H2O2) were investigated. The knockdown of
MORC2 markedly increased apoptosis of the HepG2 cells (Fig. 7B). These results indicated that the
downregulation of MORC2 promoted cell cycle arrest and induced the
apoptosis of liver cancer cells.
Knockdown of MORC2 inhibits cell
migration and invasion of liver cancer cells in vitro
The clinicopathological analysis also revealed that
a higher expression level of MORC2 was significantly associated
with a gene expression signature of higher metastatic potential
(P<0.001, Table III),
however, the role of MORC2 in cancer cell metastasis has not been
investigated previously. Cell migration and invasion are critical
during the multistep process of cancer cell metastasis. The present
study assessed whether MORC2 was a crucial molecule involved in
cell migration and invasion using Transwell assays. MORC2-loss of
function inhibited the migration and invasion rates of the HepG2,
Bel-7402 and LM3 cells (Fig. 8A and
B). To confirm this result, a scratch wound healing assay was
also used to evaluate the effect of MORC2 on cell movement.
Consistent with the previous observations, the inhibition of MORC2
attenuated the mobility of the liver cancer cells (Fig. 8C).
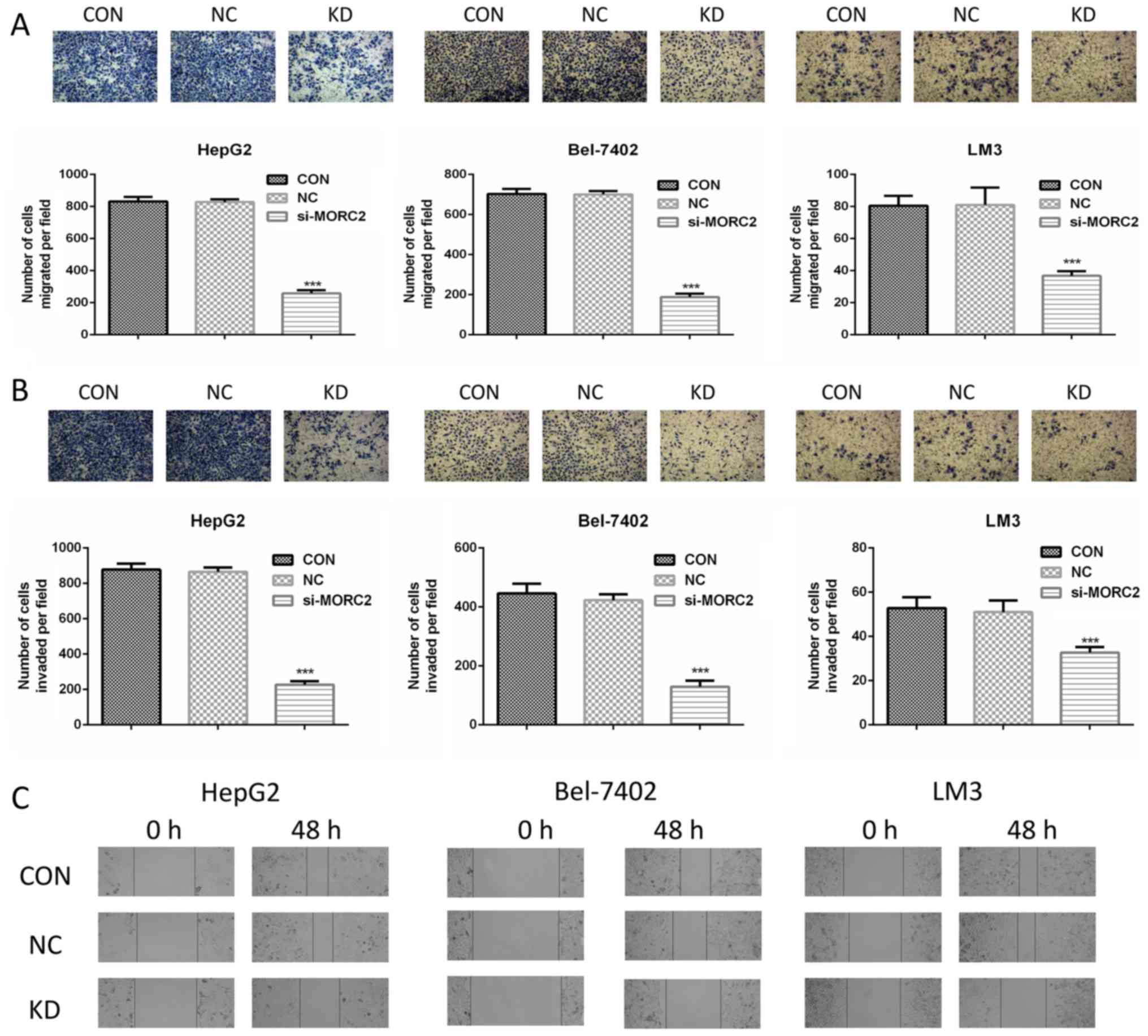 | Figure 8MORC2 modulates migration, invasion
and metastasis of liver cancer cells in vitro. (A) Cell
migration was assessed using a Transwell assay in HepG2 (left),
Bel-7402 (middle) and LM3 (right) cells following transfection of
the cells with MORC2 siRNA for 48 h. The cells that migrated into
the bottom surface of the filters were stained. Magnification, ×40.
(B) Cell invasion was assessed using a Transwell assay with
Matrigel in HepG2 (left), Bel-7402 (middle) and LM3 (right) cells
following transfection of the cells with MORC2 siRNA for 48 h. The
cells that invaded into the lower surface of the filters were
stained. Magnification, ×40. (C) Movement ability was detected by
scratch wound healing assays in HepG2 (left), Bel-7402 (middle) and
LM3 (right) cells following transfection of the cells with MORC2
siRNA for 48 h. Magnification, ×40. Data are presented as the mean
± standard deviation of six independent experiments.
***P<0.001. MORC2, microrchidia 2; siRNA, small
interfering RNA; CON, control; KD, knockdown; NC, negative
control. |
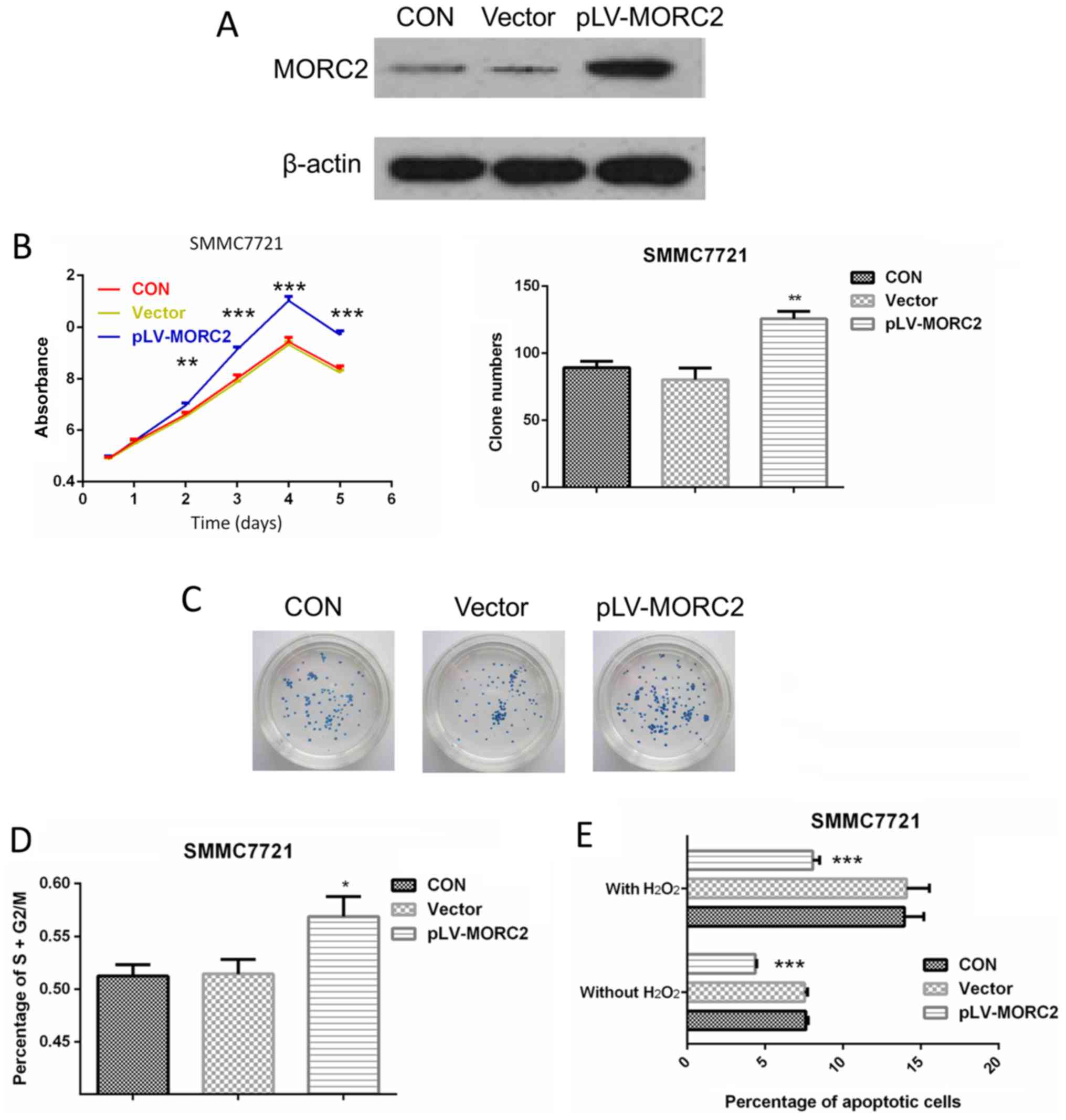
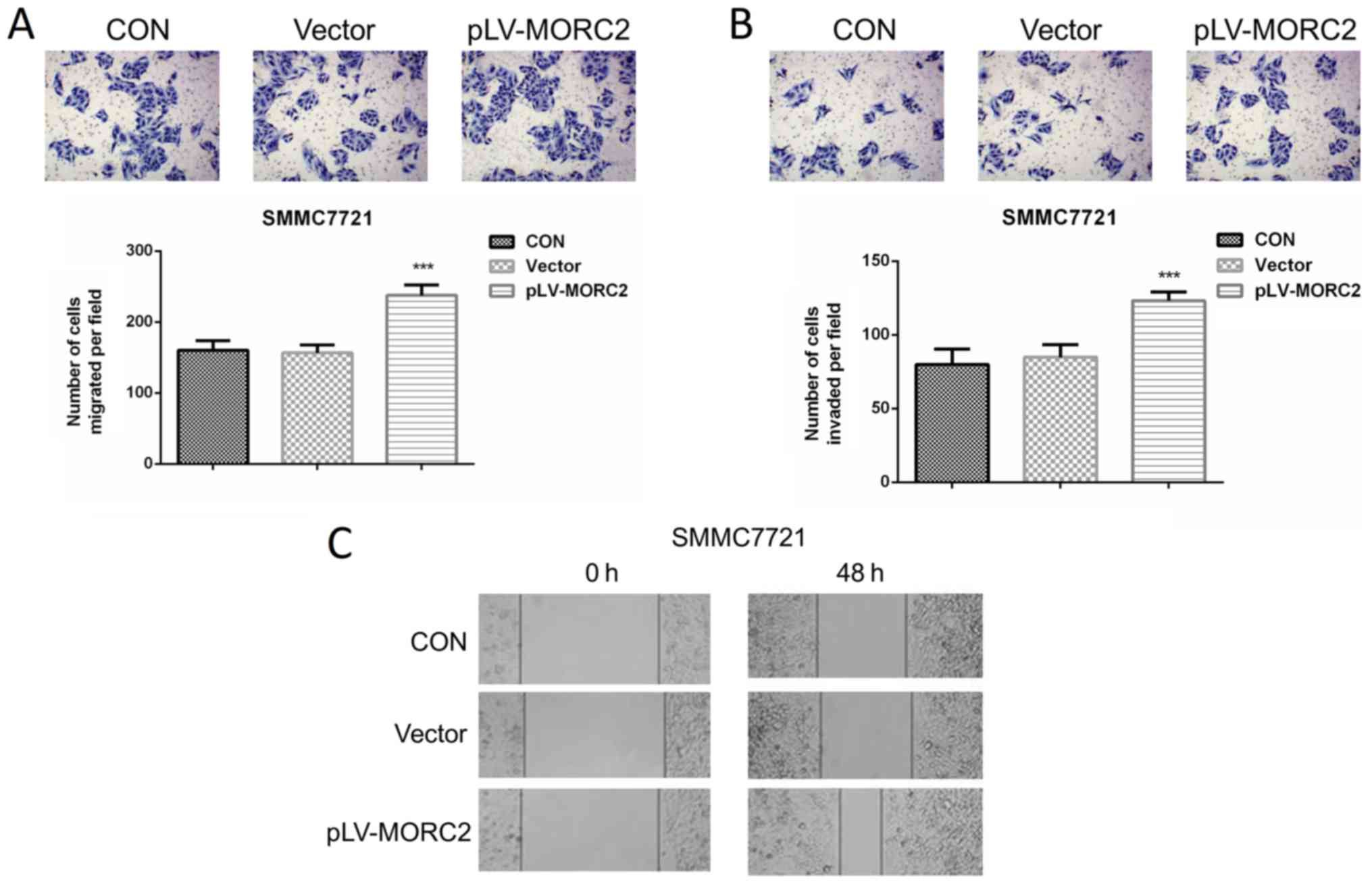
Overexpression of MORC2 promotes the
malignant phenotypes of the SMMC7721 liver cancer cell line in
vitro and in vivo
The SMMC7721 cell line, which had a relatively lower
expression of MORC2, was used to construct cells overexpressing
MORC2 using a lentivirus-mediated packed pLV-MORC2 vector. Western
blot analysis was used to determine the knockdown and ectopic
expression (Fig. 9A). Compared
with the control group, SMMC7721 cells exhibited a significantly
higher cell viability rate when MORC2 was overexpressed (Fig. 9B). Similarly, the
MORC2-overexpressing SMMC7721 cells exhibited significantly
increased colony formation (Fig.
9C). The overexpression of MORC2 increased the cell cycle
progression and decreased the number of apoptotic cells (Fig. 9D and E). Furthermore, SMMC-7721
cells stably expressing MORC2 exhibited enhanced migration,
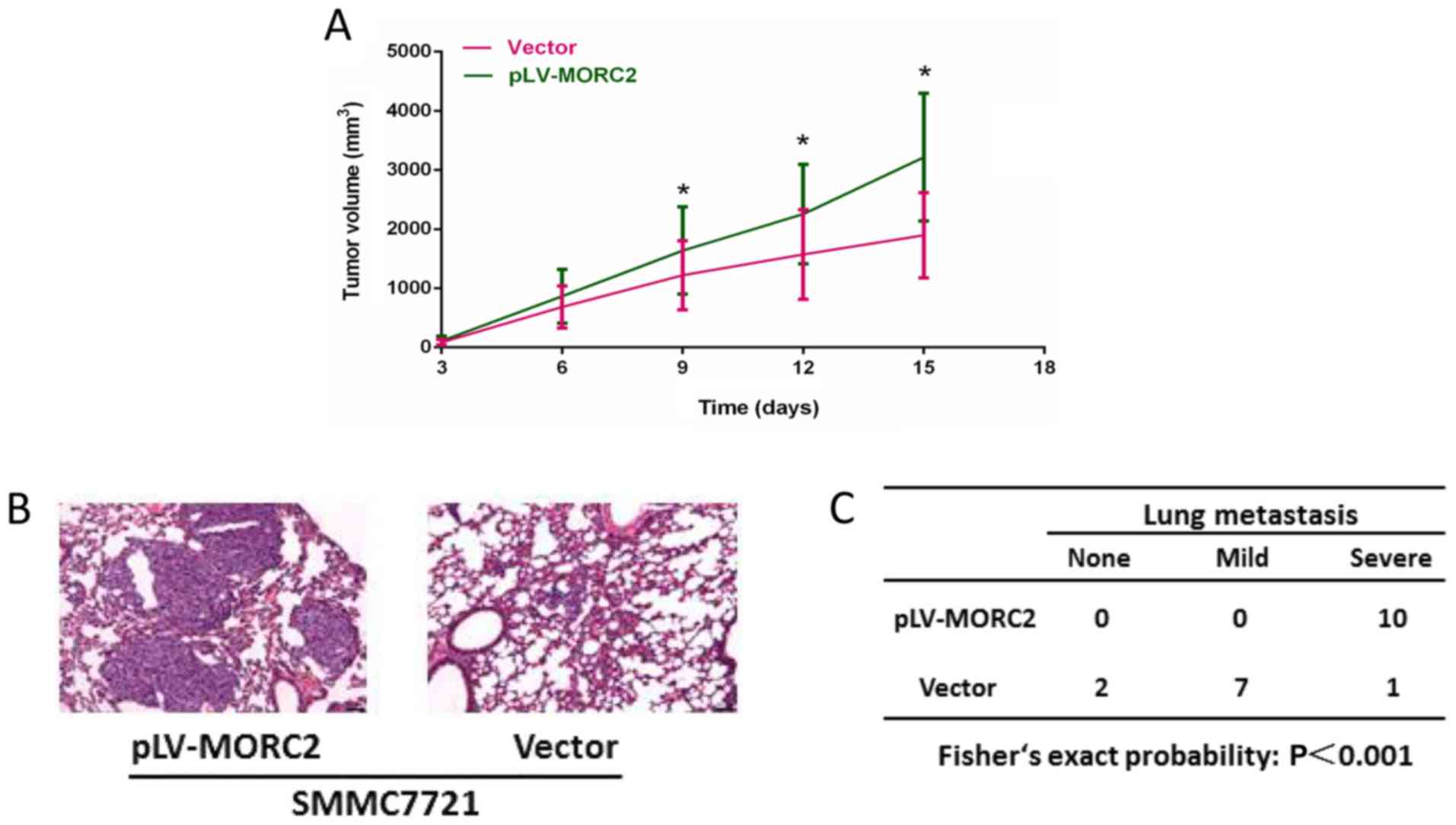
invasion and movement, compared with the control cells (Fig. 10). The in vivo experiments
showed that the overexpression of MORC2 significantly increased
tumor volume in a subcutaneous xenograft model (Fig. 11A). Colonization at a distant site
is the last key step in the metastatic cascade (21). The present study also used a tail
vein injection model to imitate the pathophysiological process to
determine whether MORC2 was involved in the distant colonization of
liver cancer cells. At 2 weeks post-injection, the mice were
sacrificed by cervical dislocation, and lung colonization was
quantified by pathological examination. In line with the
observation in vitro, all 10 mice exhibited severe lung
metastasis in the MORC2-overexpression group, the incidence of
which was significantly higher, compared that in the empty vector
group (1/10; P<0.001) (Fig. 11B
and C). These results revealed the promoting role of MORC2 in
liver cancer metastasis.
Inhibition of MORC2 improves the
sensitivity of liver cancer cells to chemotherapeutic drugs
The present study also examined whether MORC2 has
the potential to be applied in clinical liver cancer treatment.
Chemotherapy provides an optional strategy in the treatment of
liver cancer, particularly for patients with advanced tumors.
However, chemotherapy is unsatisfactory due to chemoresistance
(22). The present study
hypothesized that the negative effect of MORC2 on the apoptosis of
liver cancer cells may also contribute to the drug resistance of
liver cancer cells. To test this hypothesis, chemosensitivity to
the three most common chemotherapeutic drugs, doxorubicin,
cisplatin and 5-fluorouracil, was assayed in MORC2-knockdown groups
and control groups. The results showed that the IC50
values of all the chemotherapeutic drugs were significantly
decreased by the knockdown of MORC2 (Table IV). Therefore, MORC2 contributed
to enhancing the chemotherapeutic sensitivity of liver cancer
cells.
| Table IVsiRNA-mediated knockdown of MORC2
increases the sensitivity of liver cancer cells to
chemotherapeutics. |
Table IV
siRNA-mediated knockdown of MORC2
increases the sensitivity of liver cancer cells to
chemotherapeutics.
| Cell group | IC50
values
|
|---|
| Doxorubicin
(µM) | Cisplatin
(µM) | 5-fluorouracil
(µM) |
|---|
| HepG2 | | | |
| si-NC | 33.53 | 25.88 | 42.82 |
| si-MORC2 | 8.86 | 7.89 | 14.04 |
| Bel-7402 | | | |
| si-NC | 34.24 | 39.78 | 50.54 |
| si-MORC2 | 15.27 | 27.84 | 31.12 |
| LM3 | | | |
| si-NC | 29.72 | 43.23 | 23.68 |
| si-MORC2 | 19.89 | 35.77 | 12.06 |
To determine the mechanisms by which MORC2 affects
apoptosis and chemoresistance, the expression levels of
apoptosis-related proteins were examined following the knockdown of
MORC2 in the cells. As shown in Fig.
12A, in the HepG2, Bel-7402 and LM3 cells, si-MORC2
transfection induced the cleavage of caspase-9, caspase-3 and PARP,
and the release of mitochondrial cytochrome c into the
cytosol. These results demonstrated that MORC2 affected apoptosis
and chemoresistance by modulating the mitochondrial apoptotic
pathway.
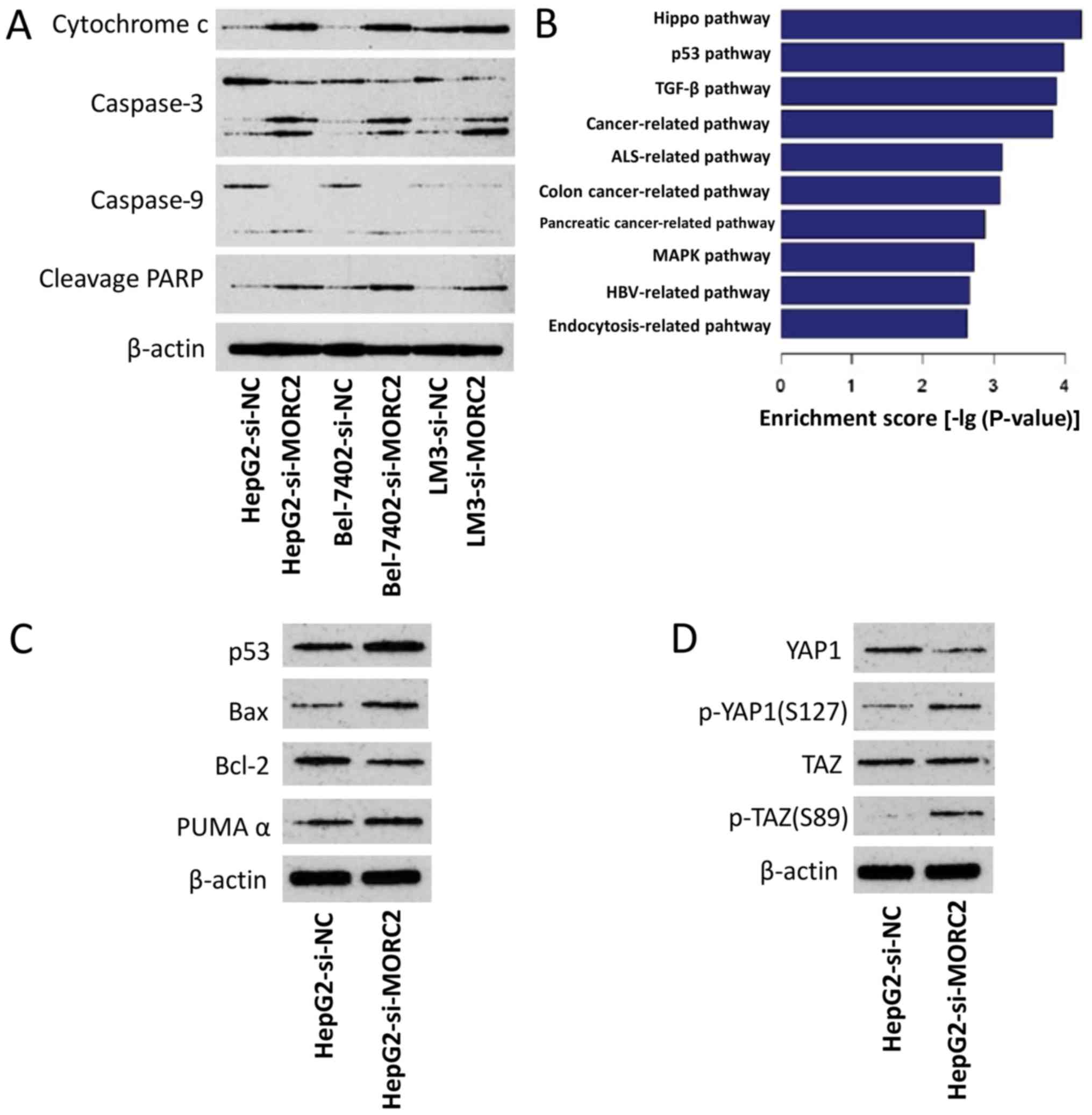 | Figure 12Dyregulation of MORC2 disrupts
several crucial cancer-related pathways. (A) Knockdown of MORC2
activates apoptotic pathways. (B) Based on the Kyoto Encyclopedia
of Genes and Genomes database, gene expression profile analysis
identified significant different pathways modulated by MORC2. (C)
In HepG2 cells, knockdown of MORC2 activated the p53 pathway. (D)
In HepG2 cells, knockdown of MORC2 inhibited the Hippo pathway.
MORC2, microrchidia; PARP, poly (ADP-ribose) polymerase; Bcl-2,
B-cell lymphoma-2; Bax, Bcl-2-associated X protein; PUMAα, p53
upregulated modulator of apoptosis α; p-, phosphorylated; si, small
interfering RNA; NC, negative control; YAP1, Yes-associated protein
1; TAZ, transcriptional co-activator with PDZ-binding motif. |
Dysregulation of MORC2 disrupts p53 and
Hippo pathways
The p53 tumor-suppressor gene regulates apoptosis
through the transcriptional activation of its target genes, and the
mitochondrial apoptotic pathway is regulated by several p53-target
genes, including Bax, Bcl-2, p53AIP1, Noxa and Puma (23–25).
Therefore, the present study examined the possible association
between p53 and MORC2 in liver cancer. By comparing the expression
profile of L02MORC2 and L02Vector, we found
that the overexpression of MORC2 significantly enriched genes
involved in the p53 pathway (P<0.001) (Fig. 12B), which was consistent with our
hypothesis. To confirm this result, the effect of MORC2-knockdown
on the expression level of p53 and its target genes, including Bax,
Bcl-2 and PUMAα, were examined. It was found that the knockdown of
MORC2 increased the expression of p53, Bax and PUMAα, and reduced
the expression of Bcl-2, compared with the control (Fig. 12C). The overexpression of MORC2
also notably enriched genes involved in the Hippo pathway (Fig. 12B). Further validating these
results, the results of the western blot analysis showed that
MORC2-knockdown increased phosphorylated Yes-associated protein
(YAP) and TAZ (Fig. 12D). These
results suggested that MORC2 may promote Hippo pathway activation
in tumorigenesis. Collectively, the dysregulation of MORC2 in liver
cancer disrupted the p53 and Hippo pathways during cancer
progression.
Discussion
The DNA damage response (DDR), is associated with
oncogenesis (26,27). The dysregulation of DNA damage
repair-related genes results in the genomic instability, promoting
the accumulation of DNA mutations and chromosomal aberrations of
the cancer genome (20,26). Increasing evidence indicates that a
high expression of certain DDR-related genes is crucial in causing
chemoresistance, which eventually results in unsatisfactory
treatment in cancer patients (28–31).
It has been reported that p21 protein (Cdc42/Rac)-activated kinase
1 phosphorylation of MORC2 on serine 739 modulates ATPase-dependent
chromatin remodeling following double-strand break damage, and
facilitates efficient DNA damage repair (6). On finding that MORC2 was upregulated
in liver cancer tissue, it was hypothesized that liver cancer cells
with high expression levels of MORC2 are able to elicit more
effective homologous recombination DNA repair, and may be less
sensitive to apoptotic signals, leading to aberrant cell cycle
progression, and higher survival ability and chemoresistance.
Accordingly, in patients with liver cancer, the present study found
that a higher fraction of copy number alterations in the genome was
detected with higher expression of MORC2. It was also found that
MORC2-knockdown induced cell cycle arrest and endogenous apoptotic
pathways. The knockdown of MORC2 also sensitized liver cancer cells
to doxorubicin, 5-fluorouracil and cisplatin, and markedly
increased IC50 values, which suggested that MORC2 may be
involved in the chemoresistance of liver cancer.
The p53 and Hippo pathways are two crucial pathways
in cancer progression. p53 has been investigated intensively as a
major tumor suppressor (23–25).
p53 can be activated and transcriptionally induces several target
genes in response to various stress signals. Its downstream genes
include modulators controlling cell proliferation, apoptosis, DNA
repair, autophagy, migration and metabolism (23). In the majority of types of human
cancer, p53 is inactivated or missing through multiple mechanisms,
resulting in tumorigenesis, cancer progression and metastasis
(23–25). It has been demonstrated that the
Hippo signaling pathway is pivotal in the regulation of tissue and
organ size during development (32). The dysregulation of Hippo signaling
leads to the inhibition of apoptosis and uncontrolled cellular
proliferation (32,33). Despite diverse upstream mechanisms
that regulate the Hippo pathway in cancer, the result is the common
activation of YAP and TAZ (34).
Previous studies have revealed that reorganization of the cell
skeleton is a crucial factor affecting the phosphorylation levels
of YAP and TAZ, and disrupting the structure of F-actin results in
the activation of LATS1, which in turn inactivates YAP and TAZ
(35–37). It is reported that ArgBP2 is a
crucial protein involved in the formation of F-actin, and its
upregulation inhibits the normal polymerization of actin (38,39).
It has been demonstrated that MORC2 can inhibit the transcription
of ArgBP2 via H2K27 trimethylation (40); therefore, MORC2 may be involved in
maintaining the normal structure of F-actin and promoting the
dephosphorylation of YAP/TAZ via the suppression of ArgBP2. This
hypothesis will be tested in future investigations. When YAP and
TAZ are activated, the proteins remain unphosphorylated,
interacting with transcriptional factors TEA domain family member
1–4, translocating into the nucleus and activating the
transcription of target genes, including connective tissue growth
factor, insulin-like growth factor binding protein 3, integrin
subunit β2, survivin, GLI family zinc finger 2 and AXL receptor
tyrosine kinase, promoting cancer progression (34). In the present study, it was
demonstrated that the dysregulation of MORC2 in liver cancer
disrupted p53 and Hippo pathways, clarifying why p53 and Hippo
pathways are aberrant in liver cancer.
The prognosis of patients with liver cancer is poor
with high incidences of early metastasis and postoperative
recurrence (22,41,42).
Radical surgery, radiotherapy and liver transplantation are
effective mainly for primary tumors, therefore, molecular-targeted
therapy has been considered as a potential treatment tool (22,42).
Clarifying the molecular mechanisms involved in the distant
metastasis in liver cancer will enable promotion of the advancement
of molecular-targeted therapies. In the present study, another
novel finding was that MORC2 promoted the migration and invasion of
cancer cells, which has not, to the best of our knowledge, been
reported previously. It was also found that MORC2-knockdown
sensitized liver cancer cells to chemotherapeutic drugs. These
results indicated that MORC2-targeted therapy may be a potentially
promising regimen to inhibit liver cancer metastasis and to improve
the effect of chemotherapeutics for patients with advanced liver
cancer and metastasis.
The present study had a number of limitations. It
was found that MORC2-knockdown affected the proliferation and
metastasis of liver cancer cells in vitro, however the
present study did not investigate the effects of inhibiting MORC2
on the proliferation and lung metastasis of liver cancer cells
in vivo. In addition, the study did not provide data on
whether MORC2 modulated the p53, hippo and apoptotic pathways in
vivo. The comprehensive underlying mechanism involved in MORC2
promoting liver cancer proliferation remains to be fully
elucidated. Sánchez-Solana et al demonstrated that MORC2
promoted the activity of adenosine triphosphate citrate lyase
(ACLY) by phosphorylation, and in turn activated acetyl-CoA
carboxylase (ACC) and fatty acid synthase (FASN) (7). Of note, de novo lipogenesis is
considered to be crucial in oncogenesis, and enhanced lipogenesis
in cancer cells is reflected by improved activities of lipogenic
enzymes, including ACLY, ACC and FASN (7,43–45).
It is appropriate to suggest that MORC2 may promote cancer
progression partly via this route. Additionally, by containing a CW
zinc finger motif, which is predicted to be involved in DNA
binding, MORC2 is considered to function as a transcriptional
regulator (5). The high expression
of MORC2 may alter the expression levels of certain crucial
downstream tumor suppressors or oncogenes, in turn promoting the
progression of cancer. These potential mechanisms require
investigation and confirmation in the future. Additionally, it has
been reported that the mutations of MORC2 may be involved in the
development of Charcot-Marie-Tooth disease (46,47).
The present study determined the mutation frequencies of MORC2 in
tumor tissues using TCGA data, and it was demonstrated that the
mutation frequencies of MORC2 in liver cancer and other types of
cancer were low (data not shown). Therefore, it was hypothesized
that mutation of MORC2 is not an important factor in tumor biology,
which also requires validation in future investigations.
Acknowledgments
The authors would like to thank Professor Dongfeng
Chen and Mr. Tao Wang (Daping Hospital, Chongqing, China) for
providing access to the expression profile data of
MORC2-overexpressing cells prior to publication.
Funding
This study was supported by the Natural Science
Foundation of China (grant nos. 81172129, 81472798 and
81703030).
Availability of data and materials
The original expression profile or RNA sequencing
data used in this study can be obtained from the following
websites: TCGA database (http://www.cbioportal.org/) and GEO datasets (GSE14520
and GSE22058) (https://www.ncbi.nlm.nih.gov/gds). The authors also
declare that the datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
QD, HY, YG and FZ conceived and designed the
experiments; QD, ZP and LW performed the experiments; QD, ZP,QG, LW
and MT conducted the statistical analysis; ZP, QG, QD, LW and MT
wrote the paper.
Ethics approval and consent to
participate
The inclusion of human samples was approved by the
Ethics Review Board of the Second People's Hospital of Guangdong
Province (approval no. 2015-KYLL-023). All patients provided
informed written consent prior to the investigation. All animal
experiments were approved by the Ethics Review Board of the Second
People's Hospital of Guangdong Province (approval no.
2015-KYLL-063).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li DQ, Nair SS and Kumar R: The MORC
family: New epigenetic regulators of transcription and DNA damage
response. Epigenetics. 8:685–693. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang GL, Wang CY, Cai XZ, Chen W, Wang XH
and Li F: Identification and expression analysis of a novel CW-type
zinc finger protein MORC2 in cancer cells. Anat Rec (Hoboken).
293:1002–1009. 2010. View
Article : Google Scholar
|
|
4
|
Moissiard G, Cokus SJ, Cary J, Feng S,
Billi AC, Stroud H, Husmann D, Zhan Y, Lajoie BR, McCord RP, et al:
MORC family ATPases required for heterochromatin condensation and
gene silencing. Science. 336:1448–1451. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shao Y, Li Y, Zhang J, Liu D, Liu F, Zhao
Y, Shen T and Li F: Involvement of histone deacetylation in
MORC2-mediated down-regulation of carbonic anhydrase IX. Nucleic
Acids Res. 38:2813–2824. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li DQ, Nair SS, Ohshiro K, Kumar A, Nair
VS, Pakala SB, Reddy SD, Gajula RP, Eswaran J, Aravind L, et al:
MORC2 signaling integrates phosphorylation-dependent,
ATPase-coupled chromatin remodeling during the DNA damage response.
Cell Rep. 2:1657–1669. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sánchez-Solana B, Li DQ and Kumar R:
Cytosolic functions of MORC2 in lipogenesis and adipogenesis.
Biochimica et Biophysica Acta (BBA). Mol Cell Res. 1843:316–326.
2014.
|
|
8
|
Chen LH, Kuo W-H, Tsai M-H, Chen P-C,
Hsiao CK, Chuang EY, Chang LY, Hsieh FJ, Lai LC and Chang KJ:
Identification of prognostic genes for recurrent risk prediction in
triple negative breast cancer patients in Taiwan. PLoS One.
6:e282222011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liao XH, Zhang Y, Dong WJ, Shao ZM and Li
DQ: Chromatin remodeling protein MORC2 promotes breast cancer
invasion and metastasis through a PRD domain-mediated interaction
with CTNND1. Oncotarget. 8:97941–97954. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tong Y, Li Y, Gu H, Wang C, Liu F, Shao Y
and Li F: HSF1, in association with MORC2, downregulates ArgBP2 via
the RC2 family in gastric cancer cells. Biochim Biophys Acta.
1864:1104–1114. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang G, Song Y, Liu T, Wang C, Zhang Q,
Liu F, Cai X, Miao Z, Xu H, Xu H, et al: PAK1-mediated MORC2
phosphorylation promotes gastric tumorigenesis. Oncotarget.
6:9877–9886. 2015.PubMed/NCBI
|
|
12
|
Zhang Q, Song Y, Chen W, Wang X, Miao Z,
Cao L, Li F and Wang G: By recruiting HDAC1, MORC2 suppresses p21
Waf1/Cip1 in gastric cancer. Oncotarget. 6:16461–16470.
2015.PubMed/NCBI
|
|
13
|
Roessler S, Jia HL, Budhu A, Forgues M, Ye
QH, Lee JS, Thorgeirsson SS, Sun Z, Tang ZY, Qin LX, et al: A
unique metastasis gene signature enables prediction of tumor
relapse in early-stage hepatocellular carcinoma patients. Cancer
Res. 70:10202–10212. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Burchard J, Zhang C, Liu AM, Poon RT, Lee
NP, Wong KF, Sham PC, Lam BY, Ferguson MD, Tokiwa G, et al:
microRNA-122 as a regulator of mitochondrial metabolic gene network
in hepatocellular carcinoma. Mol Syst Biol. 6:4022010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu AM, Yao TJ, Wang W, Wong KF, Lee NP,
Fan ST, Poon RT, Gao C and Luk JM: Circulating miR-15b and miR-130b
in serum as potential markers for detecting hepatocellular
carcinoma: A retrospective cohort study. BMJ Open. 2:e0008252012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gao J, Aksoy BA, Dogrusoz U, Dresdner G,
Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, et al:
Integrative analysis of complex cancer genomics and clinical
profiles using the cBio-Portal. Sci Signal. 6:pl12013. View Article : Google Scholar
|
|
17
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: an open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
19
|
Ding Q, He K, Luo T, Deng Y, Wang H, Liu
H, Zhang J, Chen K, Xiao J, Duan X, et al: SSRP1 Contributes to the
Malignancy of Hepatocellular Carcinoma and Is Negatively Regulated
by miR-497. Mol Ther. 24:903–914. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tao ZH, Wan JL, Zeng LY, Xie L, Sun HC,
Qin LX, Wang L, Zhou J, Ren ZG, Li YX, et al: miR-612 suppresses
the invasive-metastatic cascade in hepatocellular carcinoma. J Exp
Med. 210:789–803. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li P, Lin Y, Zhang Y, Zhu Z and Huo K:
SSX2IP promotes metastasis and chemotherapeutic resistance of
hepatocellular carcinoma. J Transl Med. 11:522013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Arakawa H: p53, apoptosis and
axon-guidance molecules. Cell Death Differ. 12:1057–1065. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu B, Chu X, Feng C, Hou J, Fan H, Liu N,
Li C, Kong X, Ye X and Meng S: Heat shock protein gp96 decreases
p53 stability by regulating Mdm2 E3 ligase activity in liver
cancer. Cancer Lett. 359:325–334. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cui X, Choi HK, Choi YS, Park SY, Sung GJ,
Lee YH, Lee J, Jun WJ, Kim K, Choi KC, et al: DNAJB1 destabilizes
PDCD5 to suppress p53-mediated apoptosis. Cancer Lett. 357:307–315.
2015. View Article : Google Scholar
|
|
26
|
Kumar Y, Yang J, Hu T, Chen L, Xu Z, Xu L,
Hu XX, Tang G, Wang JM, Li Y, et al: Massive interstitial
copy-neutral loss-of-heterozygosity as evidence for cancer being a
disease of the DNA-damage response. BMC Med Genomics. 8:422015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Stuhldreier F, Kassel S, Schumacher L,
Wesselborg S, Proksch P and Fritz G: Pleiotropic effects of
spongean alkaloids on mechanisms of cell death, cell cycle
progression and DNA damage response (DDR) of acute myeloid leukemia
(AML) cells. Cancer Lett. 361:39–48. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ho H, Aruri J, Kapadia R, Mehr H, White MA
and Ganesan AK: RhoJ regulates melanoma chemoresistance by
suppressing pathways that sense DNA damage. Cancer Res.
72:5516–5528. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jia H, Cong Q, Chua JF, Liu H, Xia X,
Zhang X, Lin J, Habib SL, Ao J, Zuo Q, et al: p57Kip2 is an
unrecognized DNA damage response effector molecule that functions
in tumor suppression and chemoresistance. Oncogene. 34:3568–3581.
2015. View Article : Google Scholar
|
|
30
|
Wang L, Mosel AJ, Oakley GG and Peng A:
Deficient DNA damage signaling leads to chemoresistance to
cisplatin in oral cancer. Mol Cancer Ther. 11:2401–2409. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang SF, Chang CW, Wei RJ, Shiue YL, Wang
SN and Yeh YT: Involvement of DNA damage response pathways in
hepatocellular carcinoma. Biomed Res Int.
2014:1538672014.PubMed/NCBI
|
|
32
|
Hong L, Cai Y, Jiang M, Zhou D and Chen L:
The Hippo signaling pathway in liver regeneration and
tumorigenesis. Acta Biochim Biophys Sin (Shanghai). 47:46–52. 2015.
View Article : Google Scholar
|
|
33
|
Zheng T, Wang J, Jiang H and Liu L: Hippo
signaling in oval cells and hepatocarcinogenesis. Cancer Lett.
302:91–99. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hong W and Guan KL: The YAP and TAZ
transcription co-activators: Key downstream effectors of the
mammalian Hippo pathway. Semin Cell Dev Biol. 23:785–793. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Aragona M, Panciera T, Manfrin A, Giulitti
S, Michielin F, Elvassore N, Dupont S and Piccolo S: A mechanical
checkpoint controls multicellular growth through YAP/TAZ regulation
by actin-processing factors. Cell. 154:1047–1059. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bae JS, Kim SM and Lee H: The Hippo
signaling pathway provides novel anti-cancer drug targets.
Oncotarget. 8:16084–16098. 2017. View Article : Google Scholar :
|
|
37
|
Zanconato F, Cordenonsi M and Piccolo S:
YAP/TAZ at the roots of cancer. Cancer Cell. 29:783–803. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Anekal PV, Yong J and Manser E: Arg
kinase-binding protein 2 (ArgBP2) interaction with α-actinin and
actin stress fibers inhibits cell migration. J Biol Chem.
290:2112–2125. 2015. View Article : Google Scholar
|
|
39
|
Cestra G, Toomre D, Chang S and De Camilli
P: The Abl/Arg substrate ArgBP2/nArgBP2 coordinates the function of
multiple regulatory mechanisms converging on the actin
cytoskeleton. Proc Natl Acad Sci USA. 102:1731–1736. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tong Y, Li Y, Gu H, Wang C, Liu F, Shao Y,
Li J, Cao L and Li F: Microchidia protein 2, MORC2, downregulates
the cytoskeleton adapter protein, ArgBP2, via histone methylation
in gastric cancer cells. Biochem Biophys Res Commun. 467:821–827.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ni F, Zhao H, Cui H, Wu Z, Chen L, Hu Z,
Guo C, Liu Y, Chen Z, Wang X, et al: MicroRNA-362-5p promotes tumor
growth and metastasis by targeting CYLD in hepatocellular
carcinoma. Cancer Lett. 356:809–818. 2015. View Article : Google Scholar
|
|
42
|
Liu J, Han P, Li M, Yan W, Liu J, Liu J,
He J, Tu W, Xia Y, Zhou Z, et al: The histidine-rich calcium
binding protein (HRC) promotes tumor metastasis in hepatocellular
carcinoma and is upregulated by SATB1. Oncotarget. 6:6811–6824.
2015.PubMed/NCBI
|
|
43
|
Calvisi DF, Wang C, Ho C, Ladu S, Lee SA,
Mattu S, Destefanis G, Delogu S, Zimmermann A, Ericsson J, et al:
Increased lipogenesis, induced by AKT-mTORC1-RPS6 signaling,
promotes development of human hepatocellular carcinoma.
Gastroenterology. 140:1071–1083. 2011. View Article : Google Scholar :
|
|
44
|
Wang C, Rajput S, Watabe K, Liao DF and
Cao D: Acetyl-CoA carboxylase-a as a novel target for cancer
therapy. Front Biosci (Schol Ed). 2:515–526. 2010.
|
|
45
|
Wang Y and He D, Yang L, Wen B, Dai J,
Zhang Q, Kang J, He W, Ding Q and He D: TRIM26 functions as a novel
tumor suppressor of hepatocellular carcinoma and its downregulation
contributes to worse prognosis. Biochem Biophys Res Commun.
463:458–465. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Laššuthová P, Šafka Brožková D, Krůtová M,
Mazanec R, Züchner S, Gonzalez MA and Seeman P: Severe axonal
Charcot-Marie-Tooth disease with proximal weakness caused by de
novo mutation in the MORC2 gene. Brain. 139:e262016. View Article : Google Scholar
|
|
47
|
Zhao X, Li X, Hu Z, Liu L, Xie Y, Tian T,
Man J, Wang J, Zi X, Xia K, et al: MORC2 mutations in a cohort of
Chinese patients with Charcot-Marie-Tooth disease type 2. Brain.
139:e562016. View Article : Google Scholar : PubMed/NCBI
|