Introduction
Melanoma is the most severe form of skin cancer,
with ~232,000 new cases diagnosed and 55,000 cases of
melanoma-associated mortality estimated annually worldwide
(1). The current treatment of
melanoma is based on surgery plus chemotherapy or radiation
therapy; however, even dacarba-zine, the standard first-line
treatment for melanoma, does not improve the overall survival
benefit (2). Previous studies have
reported that numerous Food and Drug Administration (FDA)-approved
drugs may be useful as novel pharmacotherapies in the treatment of
malignant tumours (3–6). Consequently, additional novel drugs
need to be identified to suppress cancer cell growth, diversify
options and enhance the effectiveness of antitumour therapy.
As an FDA-approved neuroleptic drug, pimozide
belongs to the diphenylpiperidine class of drugs, and is commonly
used in the treatment of Tourette syndrome and schizophrenia
(7). Previous studies have
revealed that pimozide is efficacious in the treatment of various
types of leukaemia and carcinoma, including acute or chronic
myelogenous leukaemia (8,9), breast cancer (10), liver carcinoma (11) and prostate cancer (12). However, the effects of pimozide on
melanoma are not sufficient, and the molecular mechanism has yet to
be fully elucidated (13,14). Therefore, it is necessary to
determine the mechanism underlying the progression of melanoma, in
order to identify a novel strategy to enhance pimozide
treatment.
It has previously been reported that a major
impediment to cancer immunotherapy is the induction of indoleamine
2, 3-dioxygenase (IDO), which is an enzyme that may contribute to
tumour immune tolerance (15). IDO
is an immunosuppressive, intracellular rate-limiting enzyme, which
initiates the catabolism of essential amino acids along the
kynurenine pathway (16). IDO
expression has been detected in various stromal and immune cells,
and is best characterized in dendritic cells (17). In cancer, IDO is expressed in
various tumour cells, and tumours expressing IDO can resist immune
reaction by tumour-associated antigen-specific host cytotoxic T
lymphocyte cells in mouse models (18). Numerous studies and clinical trials
have evaluated the role of IDO and its inhibitors in animal models
and in patients with cancer, and have revealed the benefit of IDO
inhibitors in cancer treatment (19–22).
Therefore, the suppression of IDO expression may be considered a
novel, effective therapeutic strategy (18,23).
The present study aimed to investigate the mechanism
underlying the antitumour effects of pimozide on melanoma cells
in vitro. Since the promising effects of combination
therapies on melanoma have been verified in previous basic and
clinical studies (24–28), the present study combined an IDO
inhibitor, L-methyl-tryptophan (L-MT), with pimozide and
investigated the antitumour effects of the combination in
vivo.
Materials and methods
Cell lines, mice and reagents
The B16 mouse melanoma cell line was provided by
Professor Liying Wang (Department of Molecular Biology, Jilin
University, Changchun, China). The cells were cultured in RPMI-1640
medium (HyClone; GE Healthcare Life Sciences, Logan, UT, USA)
supplemented with 10% foetal bovine serum (MP Biomedicals, LLC,
Santa Ana, CA, USA) at 37°C in an atmosphere containing 5%
CO2. A total of 120 male C57BL/6 mice (weight, 18–22 g;
age, 6 weeks) were purchased from Beijing Vital River Laboratory
Animal Technology Co., Ltd. (Beijing, China) and were maintained at
22±2°C with a 12-h light/dark cycle under pathogen-free conditions.
All mice had free access to food and water, and the animal studies
were approved by the Ethics Committee of Xinxiang Medical
University (Xinxiang, China). Pimozide was obtained from Shanghai
ZZBIO Co., Ltd. (Shanghai, China) and L-MT was purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
Cell cytotoxicity and cell viability
assays
Cell cytotoxicity following treatment with various
concentrations of pimozide was monitored according to morphological
observation, whereas cell viability was detected using the Cell
Counting kit (CCK)-8 assay (Beyotime Institute of Biotechnology,
Shanghai, China), according to the manufacturer's protocol.
Briefly, 1×104 B16 cells were seeded onto 96-well plates
(Corning Incorporation, Corning, NY, USA) and incubated at 37°C in
a 5% CO2 humidified incubator for 24 h. Subsequently,
the cells were treated with dimethyl sulfoxide or 0–40 µg/ml
pimozide for 24 and 48 h. Cells in culture medium alone served as
the untreated control group. For the cell cytotoxicity assay,
images of the cells were captured under an inverted microscope at
24 or 48 h. For cell viability assays, CCK-8 regent (10 µl)
was added to each well, and cells were incubated for 2 h at 37°C,
after which the cells were analysed by measuring absorbance at 450
nm using a plate reader. Cell viability was exhibited as the
optical density value.
Wound-healing assay
A total of 3×105 B16 cells were seeded
onto 6-well plates (Corning Incorporated) and were incubated at
37°C in a 5% CO2 humidified incubator for 24 h.
Subsequently, each well was scratched using a thin, disposable
pipette tip, in order to generate a wound in the cell monolayer.
B16 cells were then treated with 0–5 µg/ml pimozide and
incubated for a further 24 or 48 h. Images of cell migration were
captured and analysed under a light microscope (TI-S, Nikon
Corporation, Tokyo, Japan).
Tumour challenge and animal
treatment
Melanoma-bearing mice were established via the
subcutaneous (s.c.) inoculation of B16 cells. Briefly,
5×105 B16 cells (s.c.) were injected into the right side
of the back of C57BL/6 mice. A total of 7 days after tumour
inoculation, mice were randomly assigned into four groups and
received daily intraperitoneal injections with PBS, pimozide (200
µg/mouse), L-MT (2 mg/mouse) or pimozide (200
µg/mouse) plus L-MT (2 mg/mouse) for 1 week (n=30/group).
Tumour incidence and tumour weight were measured daily, and the
experiment was terminated on day 30.
Haematoxylin and eosin (HE) staining,
immunohistochemistry (IHC) and terminal deoxynucleotidyl
transferase dUTP nick end-labelling (TUNEL) staining
A total of 21 days after inoculation, nine of the
mice in each group were sacrificed, and the tumours were excised
and fixed in 4% formalin at room temperature for >24 h.
Formalin-fixed and paraffin-embedded specimens were cut into
5-µm sections, which were mounted on glass slides. The
sections underwent standard HE staining (29). Images were captured under a light
microscope (TI-S; Nikon Corporation). For IHC, sections were
deparaffinized and dehydrated in a series of xylene and alcohol
washes. Following quenching of endogenous peroxidase activity with
3% (vol/vol) H2O2 in methanol for 15 min,
sections were microwaved (10 min) in citrate buffer for antigen
retrieval. The tissues were then blocked with 1% (wt/vol) bovine
serum albumin (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) at room temperature for 15 min and incubated with monoclonal
antibodies directed against CD4 (1:100) and CD8 (1:100) (cat. nos.
25229 and 98941; both Cell Signaling Technology, Inc., Danvers, MA,
USA) overnight at 4°C. After washing with PBS, the sections were
incubated with a horseradish peroxidase-conjugated immunoglobulin G
secondary antibody (1:1,000; cat. no. BS13278; Bioworld Technology,
Inc., St. Louis Park, MN, USA) for 30 min at room temperature.
Immunostaining for CD4 and CD8 was evaluated by light microscopy
(TI-S; Nikon Corporation) in a blinded fashion. The intensity of
CD4 or CD8 cell staining was scored on a 0 to 3+ scale: 0, no
staining identified; 1+, <25% of positive cells; 2+, 25–75%
positive cells; and 3+, >75% positive cells. A TUNEL assay was
used to detect tumour cell apoptosis and was performed according to
the manufacturer's protocol (Beyotime Institute of Biotechnology).
TUNEL-positive cells were detected under light microscopy.
Western blotting
Cells were harvested at the 24 or 48 h following
pimozide treatment. For animal experiments, nine mice were
sacrificed from each group, and tumours were harvested and
immediately frozen in liquid nitrogen at day 14 after treatment.
Proteins were extracted from the samples using
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology) and the protein concentrations were determined using
a bicinchoninic acid protein assay (Beyotime Institute of
Biotechnology). Protein samples (50 µg/lane) were
subsequently separated by SDS-PAGE on 10–15% resolving gels, and
were transferred onto poly-vinylidene fluoride membranes (EMD
Millipore, Billerica, MA, USA). After blocking with 5% non-fat milk
for 1 h at room temperature, the membranes were incubated with the
following primary antibodies: IDO (1:1,000; cat. no. 68572), cyclin
D1 (1:1,000; cat. no. 2978), proliferating cell nuclear antigen
(PCNA) (1:1,000; cat. no. 13110), signal transducer and activator
of transcription (Stat)3 (1:1,000; cat. no. 9139), phosphorylated
(p)-Stat3 (1:500; cat. no. 9145), Stat5 (1:1,000; cat. no. 9363),
p-Stat5 (1:500; cat. no. 4322), cleaved caspase 3 (1:500; cat. no.
9664s), cleaved caspase 7 (1:500; cat. no. 9491), matrix
metalloproteinase 2 (MMP2) (1:1,000; cat. no. 87809) and
α/β-tubulin (1:1,000; cat. no. 2148) (all from Cell Signaling
Technology, Inc.) overnight at 4°C. Appropriate, horseradish
peroxidase-conjugated anti-rabbit or anti-mouse immunoglobulin G
secondary antibodies (1:2,000; cat. nos. 7074 and 7076; Cell
Signaling Technology, Inc.) were subsequently used for 1 h at room
temperature. Specific immune complexes were visualized using
enhanced chemiluminescence (Beyotime Institute of Biotechnology).
Blots were semi-quantifed using Quantity One software (version
4.62; Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Flow cytometry
A total of 14 days post-treatment, spleens were
excised from nine mice in each group, lysed with Red Blood Cell
Lysis Buffer (Beyotime Institute of Biotechnology), centrifuged at
204 × g for 5 min at 4°C and washed with PBS. Cell suspension was
prepared at a concentration of 1×107/ml. Subsequently,
100 µl cell suspension was incubated with appropriate
fluorochrome-labelled CD3 (2 µl; cat. no. 100204), CD4 (1.5
µl; cat. no. 100412), CD8 (1.5 µl; cat. no. 100708)
and natural killer (NK)1.1 (0.7 µl; cat. no. 108706)
antibodies (all from BioLegend, Inc., San Diego, CA, USA) in the
dark for 30 min at 4°C. Regulatory T cell (Treg) detection was
performed using the Mouse Regulatory T Cell Staining kit
(eBioscience; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The fluorescence intensity of cells was
measured using a flow cytometer (Guava easyCyte; EMD Millipore)
with a minimum of 10,000 cells collected.
Statistical analysis
All values are expressed as the means ± standard
deviation of three independent experiments. Statistical analyses
were performed using SPSS software 21.0 (SPSS Inc., Chicago, IL,
USA). Differences of measurement data were compared using one-way
analysis of variance (ANOVA) followed by the least significant
difference post hoc test. For survival analysis, the Kaplan-Meier
method with log-rank test was used. P<0.05 was considered to
indicate a statistically significant difference.
Results
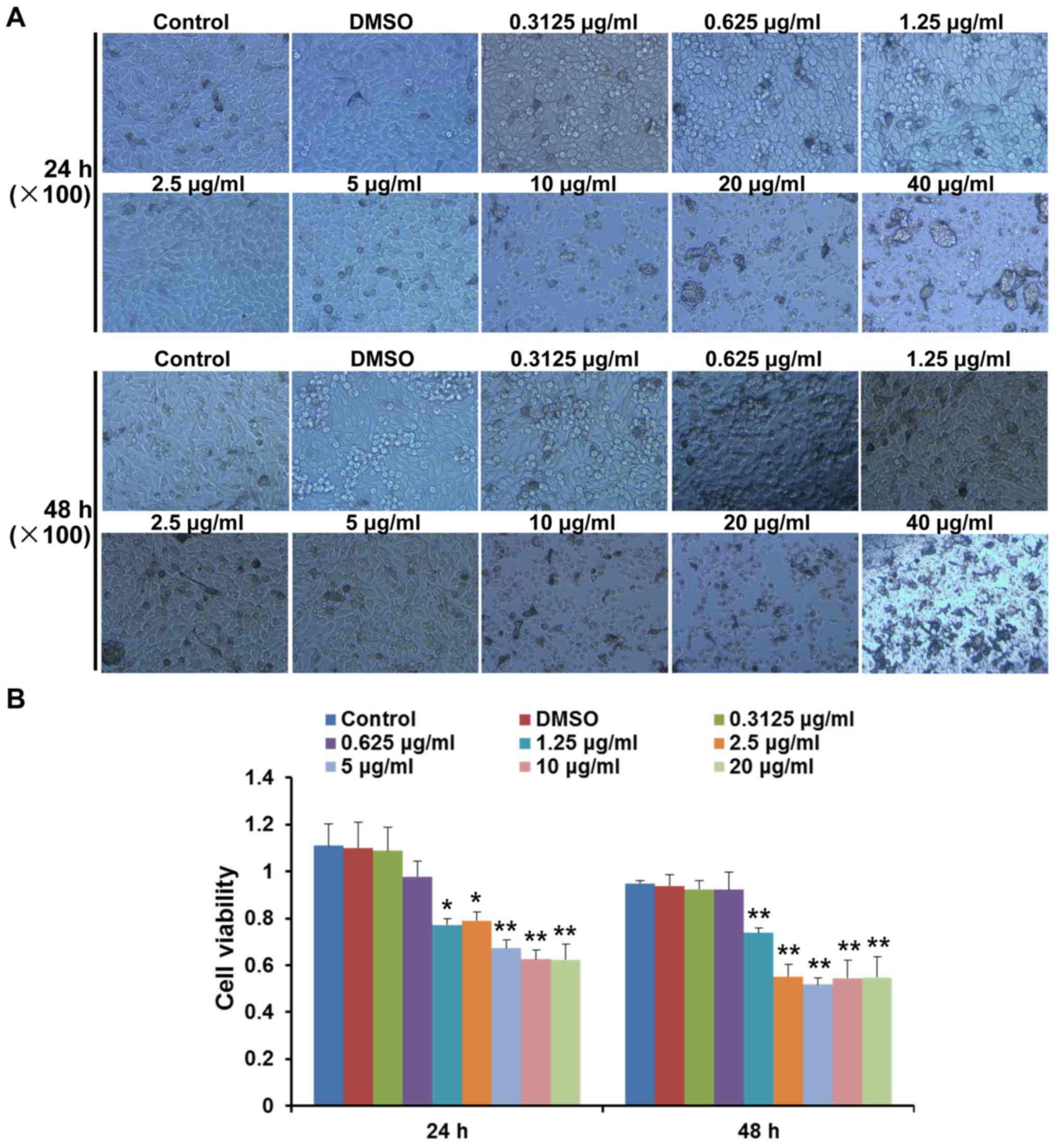
Pimozide inhibits B16 cell growth in
vitro without toxicity when the concentration is <10 µg/ml
The present study analysed the cytotoxic effects of
various concentrations of pimozide on B16 cell growth at 24 and 48
h. Morphological alterations and cell detachment were observed at
concentrations >10 µg/ml (Fig. 1A). To further determine whether
pimozide exerted direct effects on B16 cell growth, the viability
of B16 cells was detected following pimozide treatment for 24 and
48 h using the CCK-8 assay. Pimozide, at a concentration between
1.25 and 20 µg/ml, decreased the viability of B16 cells
following treatment for 24 and 48 h (P<0.05), whereas pimozide
at concentrations between 0.3125 and 0.625 µg/ml did not
exert antitumour effects (P>0.05; Fig. 1B). These results suggested that the
1.25–5 µg/ml pimozide did not exhibit toxicity but inhibited
B16 cell viability.
Apoptosis- and proliferation-associated
proteins are regulated following treatment with pimozide
To determine the effects of pimozide on
proliferation and apoptosis following 24 and 48-h treatment, the
expression levels of related proteins, including Stat5, p-Stat5,
cyclin D1, PCNA, cleaved caspase 3 and cleaved caspase 7 were
detected. As shown in Fig. 2A–D,
treatment with pimozide at various concentrations altered protein
expression; proliferation-associated proteins (cyclin D1, PCNA,
Stat5 and p-Stat5) were suppressed and the expression of
apoptosis-associated proteins (cleaved caspase 3 and cleaved
caspase 7) were increased. In addition, the present study detected
whether pimozide could affect the expression levels of IDO. The
results demonstrated that 1.25 and 2.5 µg/ml pimozide
inhibited the expression of IDO (P<0.01; Fig. 2C and D). Conversely, when the
concentration of pimozide reached 5 µg/ml, IDO expression
was increased (Fig. 2C and D).
These results suggested that apoptotic and proliferative proteins
were regulated when treated with the appropriate concentration of
pimozide.
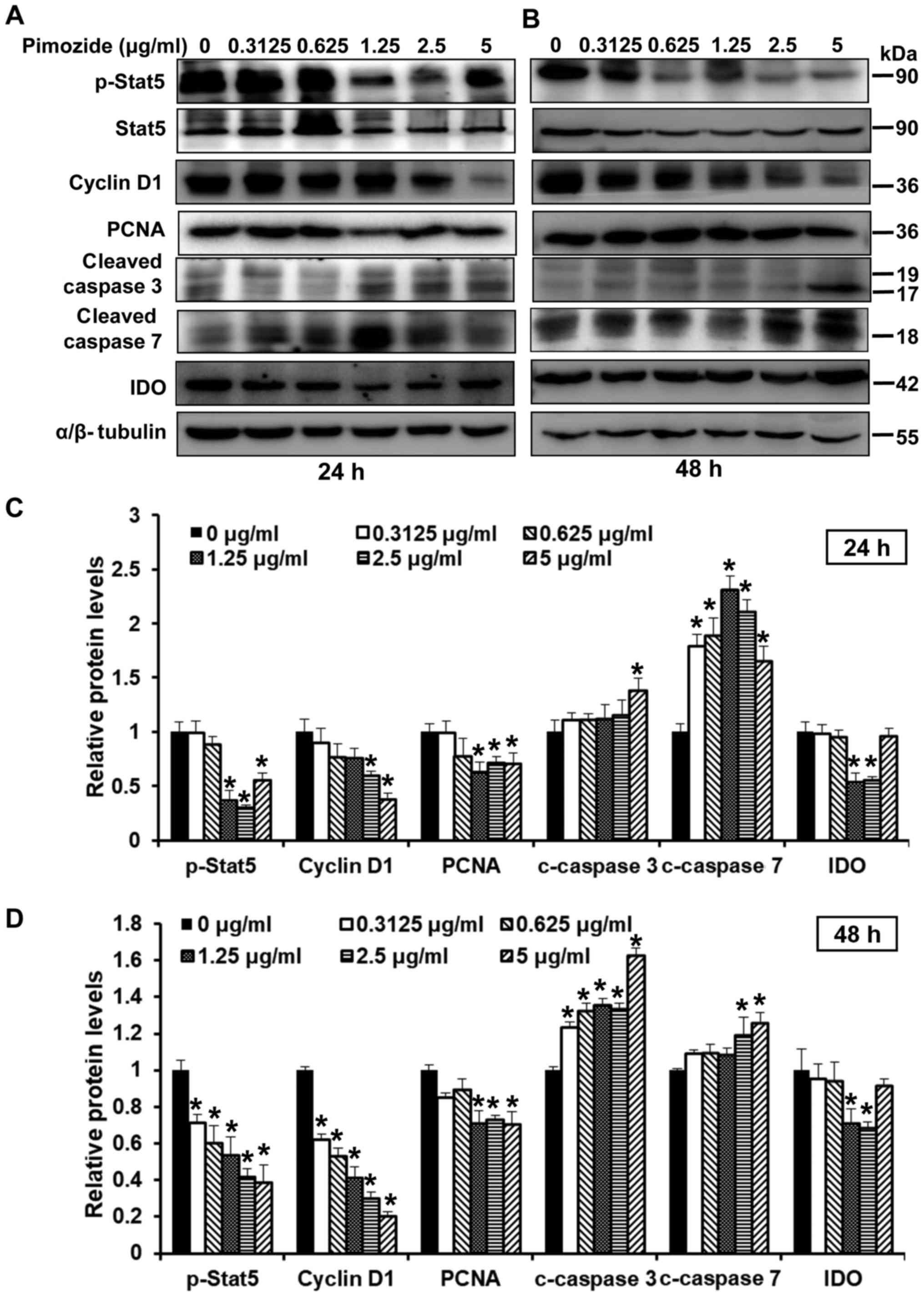 | Figure 2Effects of pimozide on the expression
of proliferation- and apoptosis-associated proteins in B16 cells.
B16 cells were treated with pimozide at the indicated doses.
Western blotting was conducted to detect the expression of
proliferation- and apoptosis-associated proteins, including
p-Stat5, Stat5, cyclin D1, PCNA, cleaved caspase 3, cleaved caspase
7 and IDO, following (A) 24 and (B) 48-h treatment. α/β-tubulin was
used as a standard. Semi-quantitative results of the relative
protein levels at (C) 24 and (D) 48 h. Values are presented as the
means ± standard deviation (n=3, in triplicate).
*P<0.01 vs. the control group. IDO, indoleamine 2,
3-dioxygenase; p-, phosphorylated; PCNA, proliferating cell nuclear
antigen; Stat, signal transducer and activator of
transcription. |
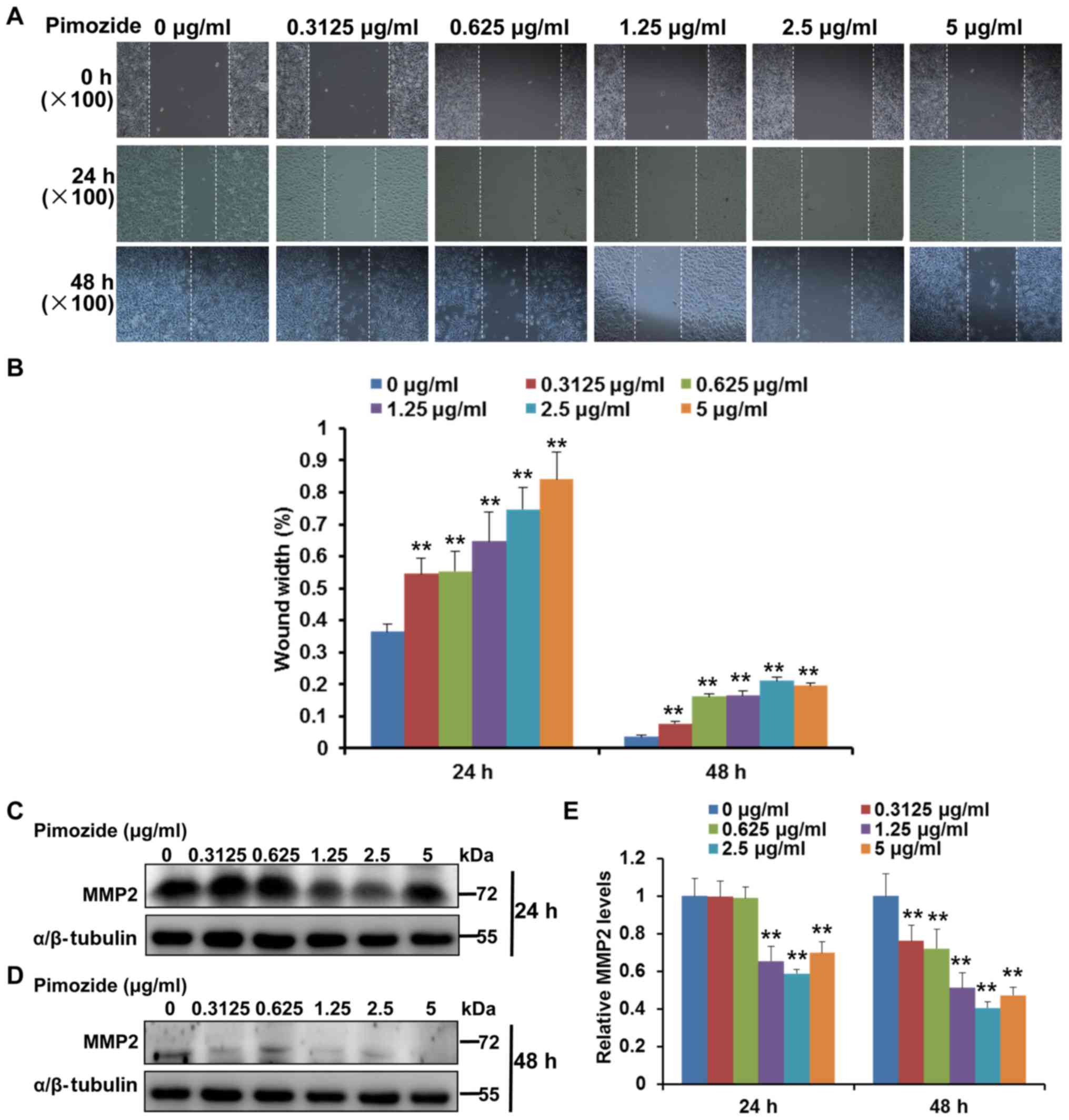
Pimozide inhibits the migration of B16
cells
Wound-healing assays were used to analyse the
effects of pimozide on cell migration. As shown in Fig. 3A and B, the wound-healing assay
indicated that treatment with >0.3125 µg/ml pimozide
significantly inhibited the migration of B16 cells at 24 and 48 h
(P<0.01), whereas in the control group, the cells migrated into
the wound area in 48 h. Furthermore, the present study investigated
whether MMP2, which has been reported to be associated with cell
migration (30), was involved in
the inhibitory effects of pimozide on migration. The results of the
western blot analysis indicated that treatment with pimozide (≥1.25
µg/ml) significantly inhibited the expression levels of MMP2
in B16 cells (P<0.01; Fig.
3C–E). Taken together, these results indicated that pimozide
may inhibit the migration of B16 cells.
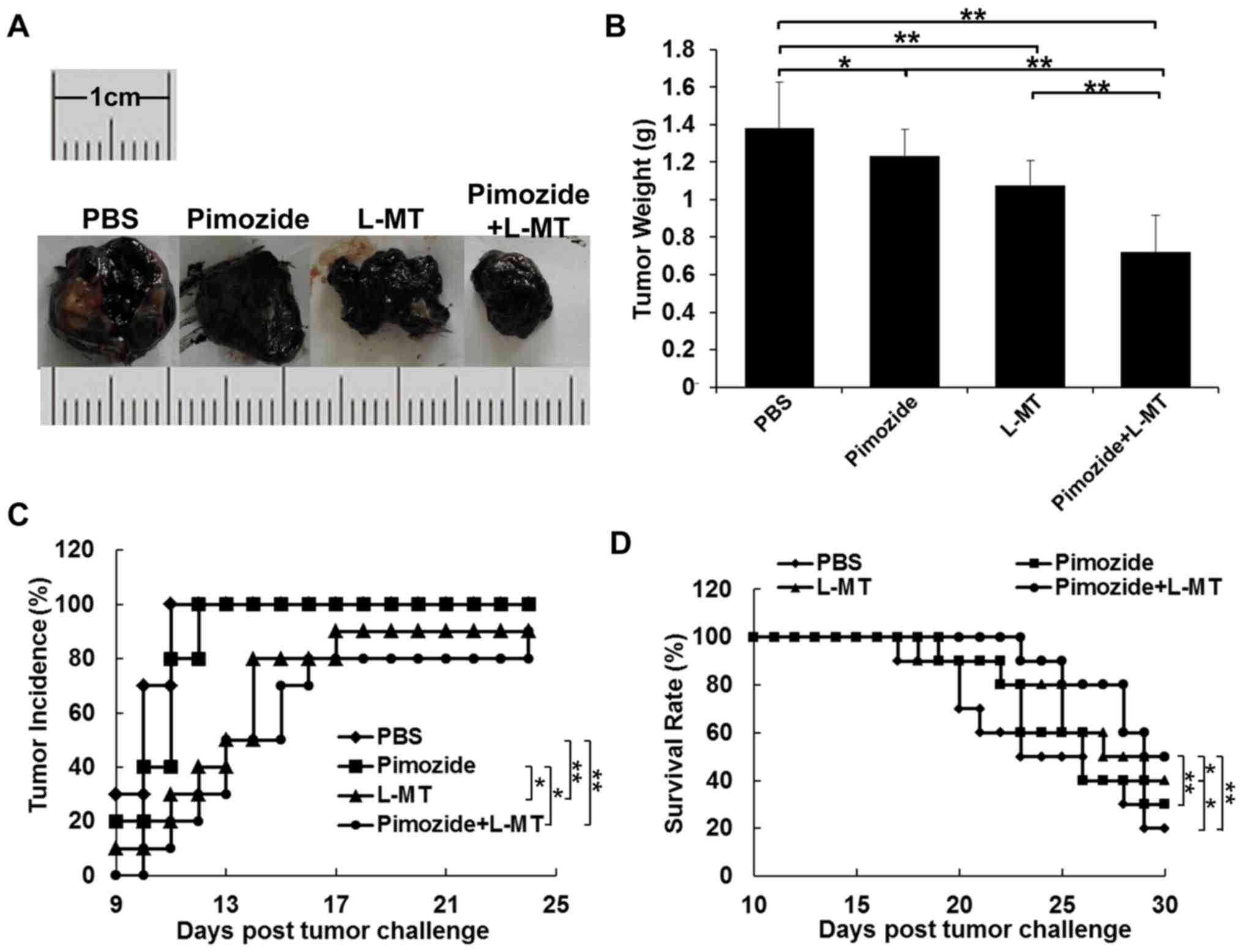
Antitumour efficacy of pimozide and L-MT
combined treatment in a mouse B16 xenograft model
To explore whether inhibiting IDO expression could
enhance the antitumour effects of pimozide in mice, the present
study administered pimozide or L-MT single therapy, and pimozide
and L-MT combined treatment. According to the results of previous
studies (9,12,31,32)
and a preliminary experiment, the optimal dosage of pimozide and
L-MT was 200 µg/mouse and 2 mg/mouse, respectively. As shown
in Fig. 4A and B, the average
tumour weight of the L-MT and pimozide groups was less than that of
the PBS group after 2 weeks of treatment; tumour weight was
decreased by 22 (P<0.01) and 11% (P<0.05), respectively. In
addition, the average tumour weight following combination therapy
was decreased by 33, 41 and 48% compared with that of the L-MT,
pimozide and PBS groups (P<0.01), respectively, thus indicating
that combination therapy exhibited the most substantial antitumour
effect. As shown in Fig. 4C,
tumour incidence in the PBS and pimozide groups was highest (100%)
on days 11 and 12, respectively, compared with in the L-MT and
combination groups, where tumour incidence rate was 90 and 80% on
day 24, respectively; this finding was consistent with tumour
weight alterations. Finally, treatment with L-MT significantly
prolonged the survival rate of B16 tumour-bearing mice compared
with in the PBS group (P<0.05). Notably, combined treatment with
pimozide and L-MT significantly prolonged the survival rate when
compared with the three other treatment groups (Fig. 4D).
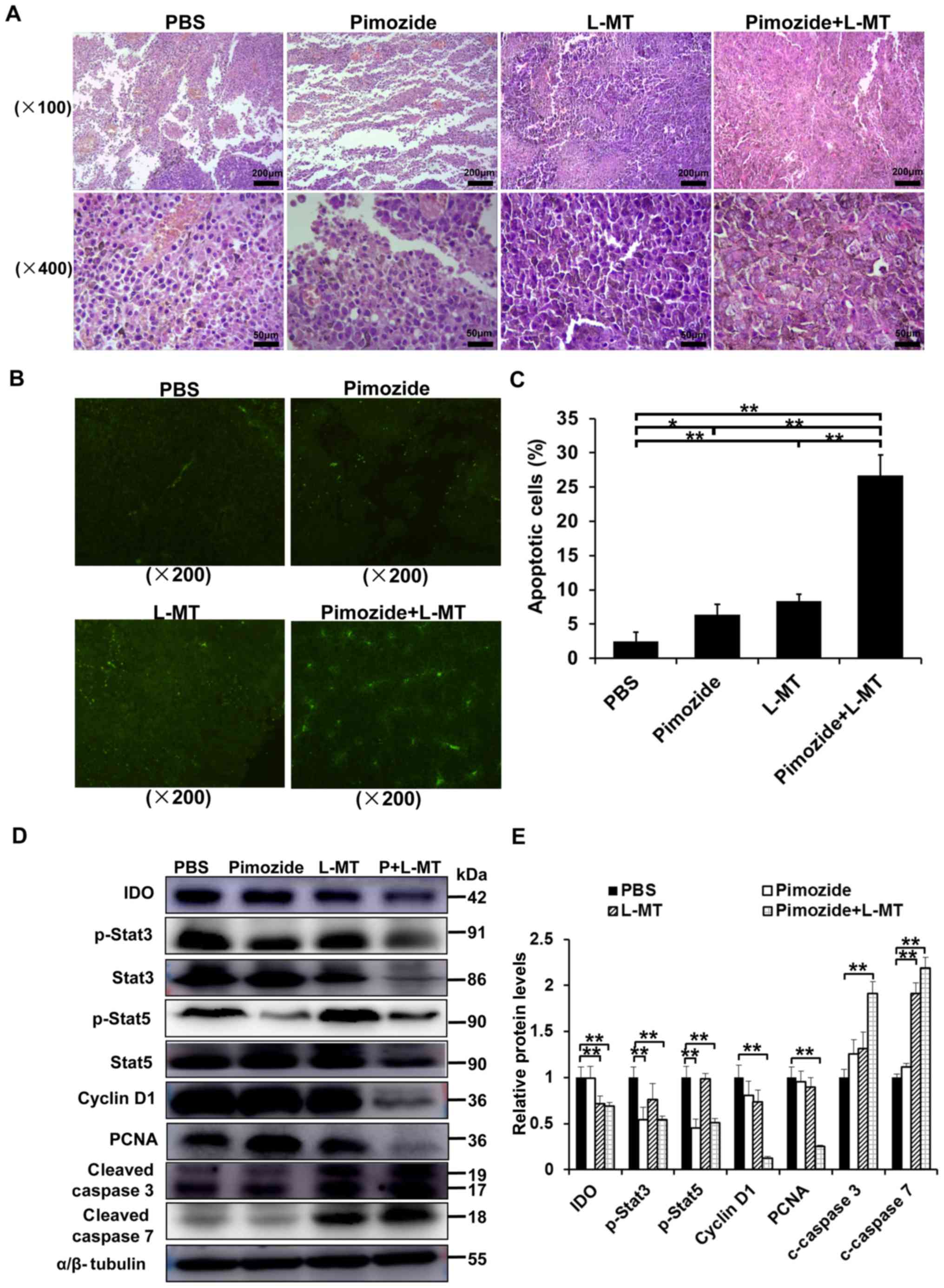
Pimozide + L-MT treatment significantly
inhibits melanoma cell proliferation and induces apoptosis in
vivo
The marked inhibitory effects of pimozide on cell
growth in vitro suggested that pimozide may suppress tumour
cell proliferation or induce B16 cell apoptosis in vivo. To
verify this hypothesis, and to detect the effects of combination
therapy, the underlying mechanisms were assessed. HE staining
detected obvious morphological differences in the tumour cells in
both monotherapy groups and particularly in the combination group
(Fig. 5A). TUNEL staining detected
an increase in the amount of tumour cells undergoing apoptosis in
the combination group (Fig. 5B and
C). To further characterize pimozide-induced tumour growth
inhibition and apoptosis, the expression levels of some associated
proteins were detected by western blotting. IDO, p-Stat3, Stat3,
p-Stat5, Stat5, cyclin D1, PCNA, cleaved caspase 3 and cleaved
caspase 7 expression levels were detected in tumour tissues
following various treatments for 2 weeks. As shown in Fig. 5D and E, IDO expression was
significantly inhibited in the L-MT and combination therapy groups,
thus indicating that the IDO inhibitor L-MT could efficiently
suppress IDO expression. Furthermore, the proliferation-associated
proteins, including p-Stat3, p-Stat5, cyclin D1 and PCNA, were
markedly decreased, whereas the apoptosis-associated proteins,
cleaved caspase 3 and cleaved caspase 7, were significantly
increased in the combination therapy group (P<0.01; Fig. 5D and E), suggesting that
combination therapy-induced apoptosis might occur via the
mitochondrial apoptotic pathway. Collectively, these results
indicated that combination therapy with pimozide and L-MT could
delay tumour growth in vivo, potentially via inhibiting
tumour cell proliferation and inducing apoptosis.
Effects of combination therapy on
T-lymphocyte infiltration into tumour tissue
To determine the effects of combination therapy on T
lymphocytes, an IHC assay was performed on tumour tissue samples.
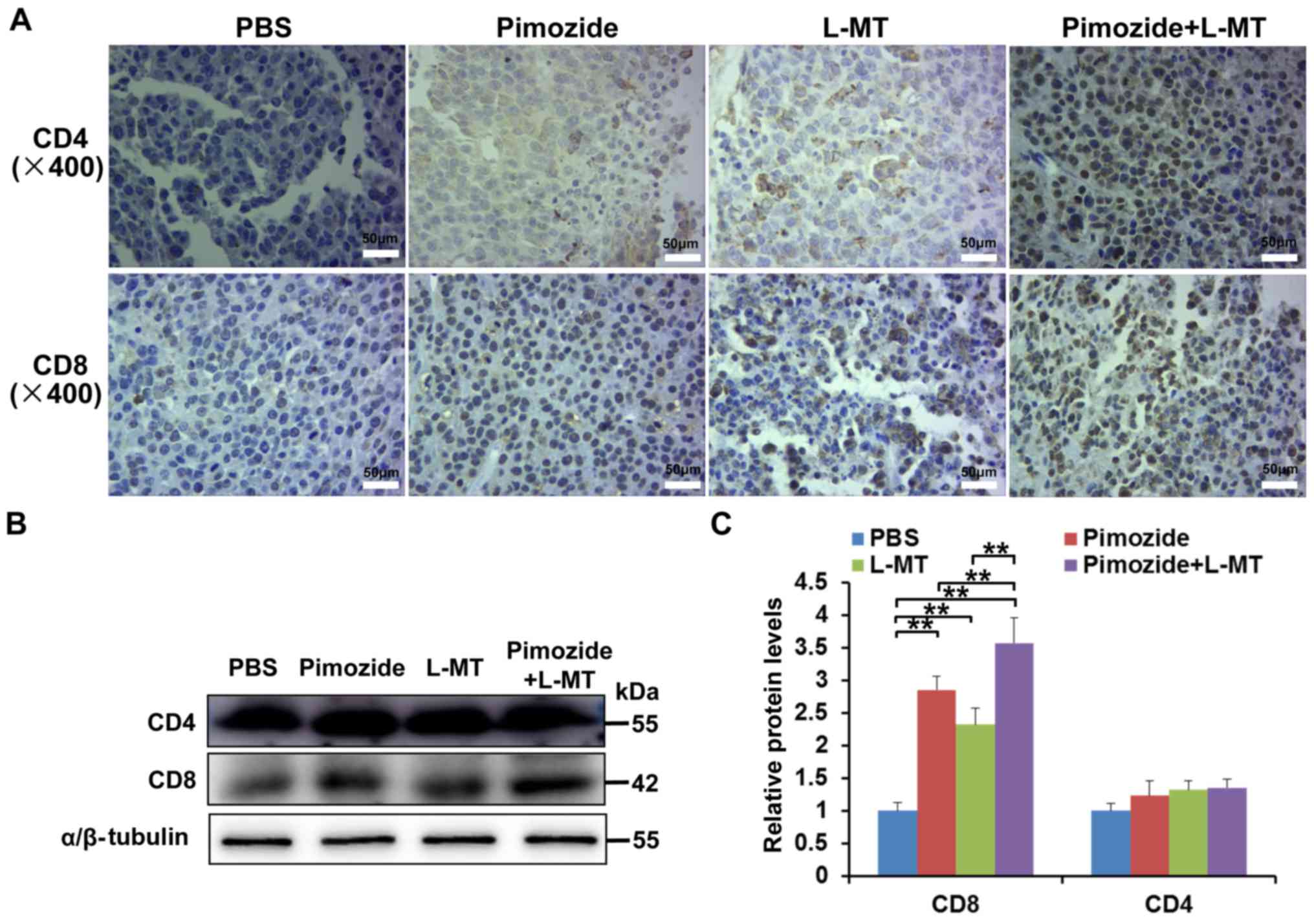
The results demonstrated that following treatment with pimozide and
L-MT, the number of CD4+ and CD8+ T cells in
tumour tissues increased, particularly CD8+ T cells
(Fig. 6A). In addition, the
results of western blotting confirmed that the expression levels of
CD8 were markedly increased in the three treatment groups,
particularly in the pimozide + L-MT group (P<0.01, Fig. 6B and C). These results suggested
that the therapeutic effects of combination therapy on melanoma may
be enhanced by promoting the T-lymphocyte response.
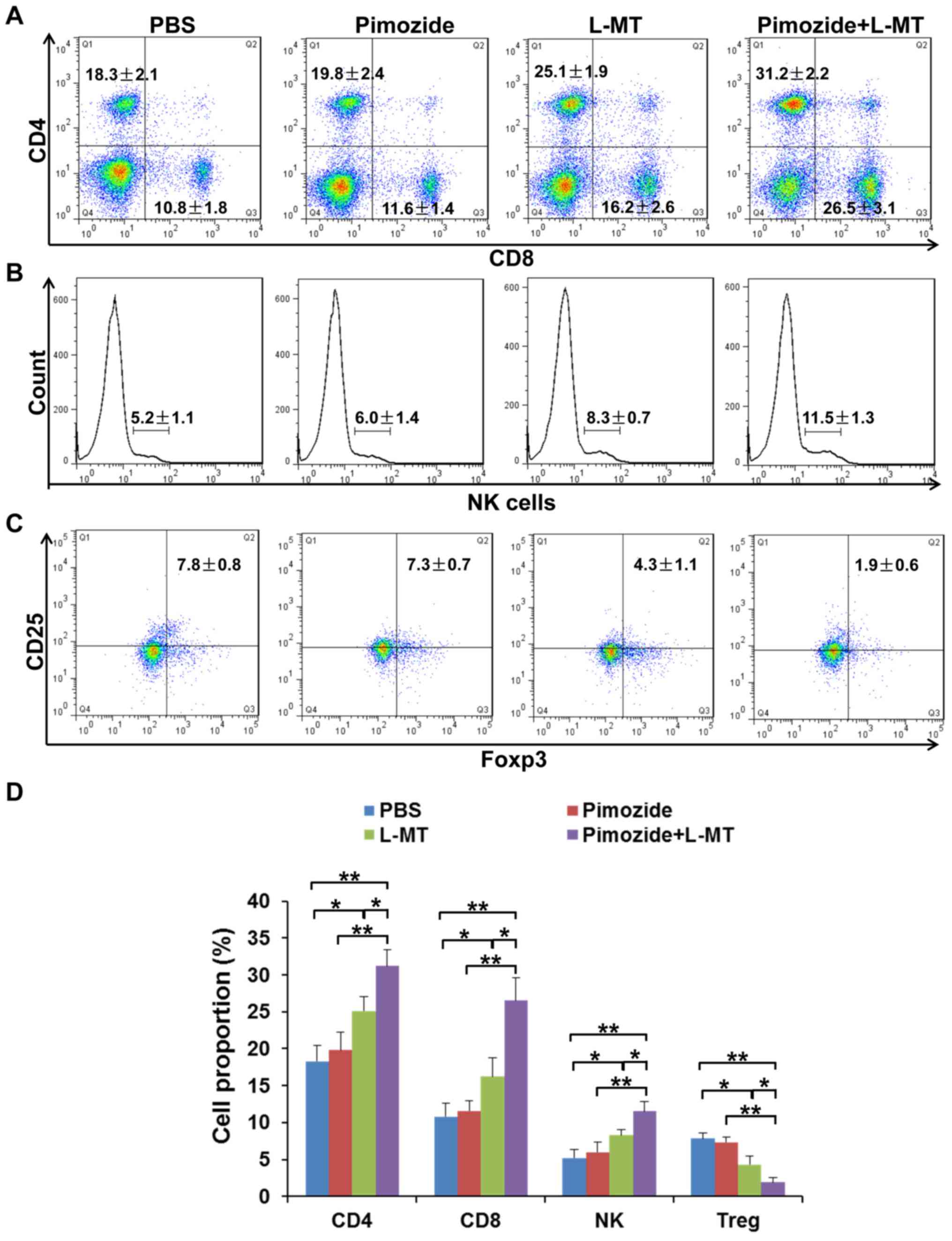
Effects of combination therapy on the
proportion of immune cells in the spleen
The spleen is the centre of cellular immunity; in
order to analyse whether tumour-infiltrating T cells multiplied
following treatment, the proportion of CD4+ and
CD8+ T cells, NK cells and
CD4+CD25+Foxp3+ Treg cells was
detected in the spleens of mice. As shown in Fig. 7, after combination therapy, the
proportion of CD4+ and CD8+ T cells, and NK
cells, was increased; among these cells, the proportion of
CD8+ T cells was most significantly increased. In
addition, Treg cells, as representatives of
CD25+Foxp3+ T cells, were significantly
inhibited following combination therapy. These results suggested
that L-MT enhanced the pimozide-induced therapeutic effect partly
through regulation of the immune response.
Discussion
Melanoma is a drug-resistant malignancy that is
found worldwide, the incidence of which is rapidly increasing
(33). This neoplasm can be
surgically removed at the early stages of disease (34); however, when the tumour is detected
at the advanced stage, there are few therapeutically effective
options that arrest or prevent tumour progression (35). Therefore, there is an urgent
requirement for a low-cost novel drug that is more effective
against melanoma (36,37). The present study provided evidence
to suggest that treatment with a combination of pimozide and an IDO
inhibitor exerted a significant therapeutic effect against
malignant melanoma in vivo.
In the present study, the antipsychotic agent
pimozide was investigated for its potential action against
melanoma. The results of a CCK-8 assay demonstrated that pimozide
(1.25–5 µg/ml) inhibited the growth of B16 cells with little
cytotoxicity. To evaluate the mechanism underlying the anti-growth
effects of pimozide, the expression levels of some key proteins
associated with proliferation and apoptosis were detected by
western blotting. Generally, as the target protein for pimozide,
Stat5 phosphorylation serves an important role in melanoma cell
survival (38). In the present
study, Stat5 phosphorylation was significantly suppressed in B16
cells following treatment with pimozide at low micromolar
concentrations. Cell cycle dysregulation has a vital role in
modulating cell proliferation regulated by cyclins and
cyclin-dependent kinases (39,40).
Additionally, the nuclear protein, PCNA, is expressed during the
G1-M phases of the cell cycle and promotes cell cycle
progression (41). The present
data indicated that treatment of B16 cells with pimozide decreased
the protein expression levels of cyclin D1 and PCNA. As a
fundamental component of cancer pathogenesis, apoptosis has a major
role in melanoma progression (42). Therefore, inducing apoptosis is an
important therapeutic approach to treat cancer (43). In the present study,
pimozide-induced apoptotic death of B16 cells was confirmed by
upregulation of cleaved caspase 3 and cleaved caspase 7. Melanoma
is a malignancy that can spread to the lungs, lymph nodes and other
organs (44), and the poor
effectiveness of melanoma treatment might be ascribed to its high
metastatic potential (45). The
occurrence of cancer cell metastasis is associated the activation
of MMP (46). In the present
study, pimozide inhibited cell migration and significantly reduced
MMP2 expression. Therefore, the present study indicated that
pimozide treatment may inhibit B16 cell growth and migration.
Drugs with potent anticancer activity in
vitro do not necessarily exhibit antitumour activity in
vivo. To date, the elimination of large tumours in the advanced
stages of cancer has been reliably achieved using a combination of
different therapies in clinical trials and animal models (25,47–50).
In addition, the IDO pathway has been reported to serve a central
role in regulating the immunological tolerance of tumours (51–54).
Numerous preclinical studies examined the application of
IDO-targeted therapy (55–57). Among the inhibitors of IDO, L-MT is
the most widely studied compound with a small molecular weight
(18,23,58).
In the present study, the results of western blotting demonstrated
that pimozide, at a concentration of 5 µg/ml, increased IDO
expression, which may be the cause of the insufficient effects of
pimozide against melanoma in the clinic. Therefore, an IDO
inhibitor combined with pimozide was used to generate an improved
antitumour effect. The results suggested that tumour growth was
markedly inhibited, and survival rate was prolonged in the
combination therapy group compared with in the PBS or other
monotherapy groups. Additionally, significantly reduced
proliferation and induced apoptosis were observed following
treatment with pimozide and L-MT, as detected by the alternative
levels of associated proteins. Unexpectedly, regardless of Stat5
suppression, activation of Stat3 was also reduced following
pimozide single treatment or pimozide + L-MT combination therapy,
thus suggesting that pimozide was not only an inhibitor of Stat5
but also a potential inhibitor of Stat3, which is consistent with
the results of a similar study in prostate cancer (12).
Tumours can escape immune attack through various
mechanisms of immunosuppression (59,60).
CD8+ T cells have an important role in antitumour
immunity (61,62), and the importance of innate immune
effector cells, such as NK cell, has also been described. A large
body of evidence has indicated that the tumour microenvironment may
prompt tumour development, progression and immune evasion (63,64).
In the present study, CD4+ and CD8+ T cells
were increased, not only in the spleen, but also in the tumour
microenvironment following combination therapy; in particular, the
proportion of CD8+ T cells was increased. Consistent
with the importance of the innate immune response, this enhanced
tumour immunity in response to combination therapy was attributed
not only to T lymphocytes, but also to NK cells. Unexpectedly,
immune stimulation was detected, to some extent, in the pimozide
monotherapy group; the underlying mechanism may be attributed to
the suppression of Stat3 activation, which is relevant to immune
escape (58). In addition, Tregs
have been identified as key components to induce the immune
tolerance of cancer cells (65).
As expected, the number of Tregs was significantly decreased
following treatment with pimozide and L-MT.
In conclusion, the present study provided evidence
to suggest that pimozide and L-MT have a combined effectiveness
against melanoma. Notably, L-MT enhanced the antitumour immunity of
pimozide against melanoma in vivo through regulating tumour
proliferation, apoptosis, migration and immunity. Therefore, these
results suggested that combination therapy with pimozide and L-MT
may be considered a novel treatment strategy for melanoma.
Acknowledgments
This study has been presented as an abstract at The
12th National Academic Congress of Immunology (303; 2017.10.26-29;
Tianjin, China).
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar
|
|
2
|
Eggermont AM and Kirkwood JM:
Re-evaluating the role of dacarbazine in metastatic melanoma: What
have we learned in 30 years. Eur J Cancer. 40:1825–1836. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhao T, Jia H, Cheng Q, Xiao Y, Li M, Ren
W, Li C, Feng Y, Feng Z, Wang H, et al: Nifuroxazide prompts
antitumor immune response of TCL-loaded DC in mice with
orthotopically-implanted hepatocarcinoma. Oncol Rep. 37:3405–3414.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Del Barco S, Vazquez-Martin A, Cufí S,
Oliveras-Ferraros C, Bosch-Barrera J, Joven J, Martin-Castillo B
and Menendez JA: Metformin: Multi-faceted protection against
cancer. Oncotarget. 2:896–917. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hossain MA, Kim DH, Jang JY, Kang YJ, Yoon
JH, Moon JO, Chung HY, Kim GY, Choi YH, Copple BL, et al: Aspirin
induces apoptosis in vitro and inhibits tumor growth of human
hepatocellular carcinoma cells in a nude mouse xenograft model. Int
J Oncol. 40:1298–1304. 2012. View Article : Google Scholar
|
|
6
|
Triscott J, Lee C, Hu K, Fotovati A, Berns
R, Pambid M, Luk M, Kast RE, Kong E, Toyota E, et al: Disulfiram, a
drug widely used to control alcoholism, suppresses the self-renewal
of glioblastoma and over-rides resistance to temozolomide.
Oncotarget. 3:1112–1123. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Egolf A and Coffey BJ: Current
pharmacotherapeutic approaches for the treatment of Tourette
syndrome. Drugs Today (Barc). 50:159–179. 2014. View Article : Google Scholar
|
|
8
|
Nelson EA, Walker SR, Xiang M, Weisberg E,
Bar-Natan M, Barrett R, Liu S, Kharbanda S, Christie AL, Nicolais
M, et al: The STAT5 inhibitor pimozide displays efficacy in models
of acute myelogenous leukemia driven by FLT3 mutations. Genes
Cancer. 3:503–511. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nelson EA, Walker SR, Weisberg E,
Bar-Natan M, Barrett R, Gashin LB, Terrell S, Klitgaard JL, Santo
L, Addorio MR, et al: The STAT5 inhibitor pimozide decreases
survival of chronic myelogenous leukemia cells resistant to kinase
inhibitors. Blood. 117:3421–3429. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Strobl JS, Kirkwood KL, Lantz TK, Lewine
MA, Peterson VA and Worley JF III: Inhibition of human breast
cancer cell proliferation in tissue culture by the neuroleptic
agents pimozide and thioridazine. Cancer Res. 50:5399–5405.
1990.PubMed/NCBI
|
|
11
|
Sun J, Jiang J, Lu K, Chen Q, Tao D and
Chen Z: Therapeutic potential of ADAM17 modulation in gastric
cancer through regulation of the EGFR and TNF-α signalling
pathways. Mol Cell Biochem. 426:17–26. 2017. View Article : Google Scholar
|
|
12
|
Zhou W, Chen MK, Yu HT, Zhong ZH, Cai N,
Chen GZ, Zhang P and Chen JJ: The antipsychotic drug pimozide
inhibits cell growth in prostate cancer through suppression of
STAT3 activation. Int J Oncol. 48:322–328. 2016. View Article : Google Scholar
|
|
13
|
Taub RN and Baker MA: Treatment of
metastatic malignant melanoma with pimozide. Lancet. 1:6051979.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Neifeld JP, Tormey DC, Baker MA, Meyskens
FL Jr and Taub RN: Phase II trial of the dopaminergic inhibitor
pimozide in previously treated melanoma patients. Cancer Treat Rep.
67:155–157. 1983.PubMed/NCBI
|
|
15
|
Mellor AL and Munn DH: IDO expression by
dendritic cells: Tolerance and tryptophan catabolism. Nat Rev
Immunol. 4:762–774. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Taylor MW and Feng GS: Relationship
between interferon-gamma, indoleamine 2.3-dioxygenase, and
tryptophan catabolism. FASEB J. 5:2516–2522. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Munn DH, Sharma MD, Lee JR, Jhaver KG,
Johnson TS, Keskin DB, Marshall B, Chandler P, Antonia SJ, Burgess
R, et al: Potential regulatory function of human dendritic cells
expressing indoleamine 2,3-dioxygenase. Science. 297:1867–1870.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Uyttenhove C, Pilotte L, Théate I,
Stroobant V, Colau D, Parmentier N, Boon T and Van den Eynde BJ:
Evidence for a tumoral immune resistance mechanism based on
tryptophan degradation by indoleamine 2,3-dioxygenase. Nat Med.
9:1269–1274. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Löb S, Königsrainer A, Rammensee HG, Opelz
G and Terness P: Inhibitors of indoleamine-2,3-dioxygenase for
cancer therapy: Can we see the wood for the trees. Nat Rev Cancer.
9:445–452. 2009. View Article : Google Scholar
|
|
20
|
Liu X, Shin N, Koblish HK, Yang G, Wang Q,
Wang K, Leffet L, Hansbury MJ, Thomas B, Rupar M, et al: Selective
inhibition of IDO1 effectively regulates mediators of antitumor
immunity. Blood. 115:3520–3530. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Su C, Zhang P, Liu J and Cao Y: Erianin
inhibits indoleamine 2,3-dioxygenase-induced tumor angiogenesis.
Biomed Pharmacother. 88:521–528. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang GM, Wang HS, Du J, Ma WF, Wang H,
Qiu Y, Zhang QG, Xu W, Liu HF and Liang JP: Bortezomib relieves
immune tolerance in nasopharyngeal carcinoma via STAT1 suppression
and indoleamine 2,3-dioxygenase downregulation. Cancer Immunol Res.
5:42–51. 2017. View Article : Google Scholar
|
|
23
|
Hou DY, Muller AJ, Sharma MD, DuHadaway J,
Banerjee T, Johnson M, Mellor AL, Prendergast GC and Munn DH:
Inhibition of indoleamine 2,3-dioxygenase in dendritic cells by
stereoisomers of 1-methyl-tryptophan correlates with antitumor
responses. Cancer Res. 67:792–801. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rogiers A, Wolter P and Bechter O:
Dabrafenib plus trametinib rechallenge in four melanoma patients
who previously progressed on this combination. Melanoma Res.
27:164–167. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dummer R, Hoeller C, Gruter IP and
Michielin O: Combining talimogene laherparepvec with
immunotherapies in melanoma and other solid tumors. Cancer Immunol
Immunother. 66:683–695. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zimmer L, Apuri S, Eroglu Z, Kottschade
LA, Forschner A, Gutzmer R, Schlaak M, Heinzerling L, Krackhardt
AM, Loquai C, et al: Ipilimumab alone or in combination with
nivolumab after progression on anti-PD-1 therapy in advanced
melanoma. Eur J Cancer. 75:47–55. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hermel DJ and Ott PA: Combining forces:
The promise and peril of synergistic immune checkpoint blockade and
targeted therapy in metastatic melanoma. Cancer Metastasis Rev.
36:43–50. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Deniger DC, Kwong ML, Pasetto A, Dudley
ME, Wunderlich JR, Langhan MM, Lee CR and Rosenberg SA: A pilot
trial of the combination of vemurafenib with adoptive cell therapy
in patients with metastatic melanoma. Clin Cancer Res. 23:351–362.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jia H, Cui J, Jia X, Zhao J, Feng Y, Zhao
P, Zang D, Yu J, Zhao T, Wang H, et al: Therapeutic effects of
STAT3 inhibition by nifuroxazide on murine acute graft
graft-vs.-host disease: Old drug, new use. Mol Med Rep.
16:9480–9486. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bauvois B: New facets of matrix
metalloproteinases MMP-2 and MMP-9 as cell surface transducers:
Outside-in signaling and relationship to tumor progression. Biochim
Biophys Acta. 1825:29–36. 2012.
|
|
31
|
Lim JY, Lee SE, Park G, Choi EY and Min
CK: Inhibition of indoleamine 2,3-dioxygenase by stereoisomers of
1-methyltryptophan in an experimental graft-versus-tumor model. Exp
Hematol. 42:862–866 e863. 2014. View Article : Google Scholar
|
|
32
|
Qian F, Liao J, Villella J, et al: Effects
of 1-methyltryptophan stereoisomers on IDO2 enzyme activity and
IDO2-mediated arrest of human T cell proliferation. Cancer Immunol
Immunother. 61:2013–2020. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cho YR and Chiang MP: Epidemiology,
staging (new system), and prognosis of cutaneous melanoma. Clin
Plast Surg. 37:47–53. 2010. View Article : Google Scholar
|
|
35
|
Damsky WE, Theodosakis N and Bosenberg M:
Melanoma metastasis: New concepts and evolving paradigms. Oncogene.
33:2413–2422. 2014. View Article : Google Scholar
|
|
36
|
Holmes D: The cancer that rises with the
sun. Nature. 515:S110–S111. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Schadendorf D, Fisher DE, Garbe C,
Gershenwald JE, Grob JJ, Halpern A, Herlyn M, Marchetti MA,
McArthur G, Ribas A, et al: Melanoma. Nat Rev Dis Primers.
1:150032015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mirmohammadsadegh A, Hassan M, Bardenheuer
W, Marini A, Gustrau A, Nambiar S, Tannapfel A, Bojar H, Ruzicka T
and Hengge UR: STAT5 phosphorylation in malignant melanoma is
important for survival and is mediated through SRC and JAK1
kinases. J Invest Dermatol. 126:2272–2280. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Malumbres M and Barbacid M: Cell cycle,
CDKs and cancer: A changing paradigm. Nat Rev Cancer. 9:153–166.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Schwartz GK and Shah MA: Targeting the
cell cycle: A new approach to cancer therapy. J Clin Oncol.
23:9408–9421. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kelman Z and Hurwitz J: Protein-PCNA
interactions: A DNA-scanning mechanism. Trends Biochem Sci.
23:236–238. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hersey P and Zhang XD: How melanoma cells
evade trail-induced apoptosis. Nat Rev Cancer. 1:142–150. 2001.
View Article : Google Scholar
|
|
43
|
Xia Y, Song X, Li D, Ye T, Xu Y, Lin H,
Meng N, Li G, Deng S, Zhang S, et al: YLT192, a novel, orally
active bioavailable inhibitor of VEGFR2 signaling with potent
antiangiogenic activity and antitumor efficacy in preclinical
models. Sci Rep. 4:60312014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fidler IJ: The pathogenesis of cancer
metastasis: The 'seed and soil' hypothesis revisited. Nat Rev
Cancer. 3:453–458. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hodis E, Watson IR, Kryukov GV, Arold ST,
Imielinski M, Theurillat JP, Nickerson E, Auclair D, Li L, Place C,
et al: A landscape of driver mutations in melanoma. Cell.
150:251–263. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Roy R, Yang J and Moses MA: Matrix
metalloproteinases as novel biomarkers and potential therapeutic
targets in human cancer. J Clin Oncol. 27:5287–5297. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bauer D, Werth F, Nguyen HA, Kiecker F and
Eberle J: Critical role of reactive oxygen species (ROS) for
synergistic enhancement of apoptosis by vemurafenib and the
potassium channel inhibitor TRAM-34 in melanoma cells. Cell Death
Dis. 8:e25942017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Booth L, Roberts JL, Sander C, Lee J,
Kirkwood JM, Poklepovic A and Dent P: The HDAC inhibitor AR42
interacts with pazopanib to kill trametinib/dabrafenib-resistant
melanoma cells in vitro and in vivo. Oncotarget. 8:16367–16386.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Thakur V, Lu J, Roscilli G, Aurisicchio L,
Cappelletti M, Pavoni E, White WL and Bedogni B: The natural
compound fucoidan from New Zealand Undaria pinnatifida synergizes
with the ERBB inhibitor lapatinib enhancing melanoma growth
inhibition. Oncotarget. 8:17887–17896. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lugini L, Sciamanna I, Federici C, Iessi
E, Spugnini EP and Fais S: Antitumor effect of combination of the
inhibitors of two new oncotargets: Proton pumps and reverse
transcriptase. Oncotarget. 8:4147–4155. 2017. View Article : Google Scholar :
|
|
51
|
Munn DH and Mellor AL: Indoleamine
2,3-dioxygenase and tumor-induced tolerance. J Clin Invest.
117:1147–1154. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Munn DH, Sharma MD, Hou D, Baban B, Lee
JR, Antonia SJ, Messina JL, Chandler P, Koni PA and Mellor AL:
Expression of indoleamine 2,3-dioxygenase by plasmacytoid dendritic
cells in tumor-draining lymph nodes. J Clin Invest. 114:280–290.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Frumento G, Rotondo R, Tonetti M and
Ferrara GB: T cell proliferation is blocked by indoleamine
2,3-dioxygenase. Transplant Proc. 33:428–430. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Munn DH and Mellor AL: IDO in the tumor
microenvironment: Inflammation, counter-regulation, and tolerance.
Trends Immunol. 37:193–207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Solms M: A previously-untranslated report
by Freud of a lecture on the mechanism of obsessional ideas and
phobias. Int J Psychoanal. 70:91–94. 1989.PubMed/NCBI
|
|
56
|
Holtzhausen A, Zhao F, Evans KS, Tsutsui
M, Orabona C, Tyler DS and Hanks BA: Melanoma-derived Wnt5a
promotes local dendritic-cell expression of IDO and
immunotolerance: Opportunities for pharmacologic enhancement of
immunotherapy. Cancer Immunol Res. 3:1082–1095. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chen D, Koropatnick J, Jiang N, Zheng X,
Zhang X, Wang H, Yuan K, Siu KS, Shunnar A, Way C, et al: Targeted
siRNA silencing of indoleamine 2, 3-dioxygenase in
antigen-presenting cells using mannose-conjugated liposomes: A
novel strategy for treatment of melanoma. J Immunother. 37:123–134.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Muller AJ, DuHadaway JB, Donover PS,
Sutanto-Ward E and Prendergast GC: Inhibition of indoleamine
2,3-dioxygenase, an immunoregulatory target of the cancer
suppression gene Bin1, potentiates cancer chemotherapy. Nat Med.
11:312–319. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
59
|
Mellman I, Coukos G and Dranoff G: Cancer
immunotherapy comes of age. Nature. 480:480–489. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Joyce JA and Fearon DT: T cell exclusion,
immune privilege, and the tumor microenvironment. Science.
348:74–80. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Fridman WH, Pagès F, Sautès-Fridman C and
Galon J: The immune contexture in human tumours: Impact on clinical
outcome. Nat Rev Cancer. 12:298–306. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Tumeh PC, Harview CL, Yearley JH, Shintaku
IP, Taylor EJ, Robert L, Chmielowski B, Spasic M, Henry G, Ciobanu
V, et al: PD-1 blockade induces responses by inhibiting adaptive
immune resistance. Nature. 515:568–571. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Kortylewski M, Swiderski P, Herrmann A,
Wang L, Kowolik C, Kujawski M, Lee H, Scuto A, Liu Y, Yang C, et
al: In vivo delivery of siRNA to immune cells by conjugation to a
TLR9 agonist enhances antitumor immune responses. Nat Biotechnol.
27:925–932. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Barker HE, Paget JT, Khan AA and
Harrington KJ: The tumour microenvironment after radiotherapy:
Mechanisms of resistance and recurrence. Nat Rev Cancer.
15:409–425. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Jia H, Li Y, Zhao T, Li X, Hu J, Yin D,
Guo B, Kopecko DJ, Zhao X, Zhang L, et al: Antitumor effects of
Stat3-siRNA and endostatin combined therapies, delivered by
attenuated Salmonella, on orthotopically implanted hepatocarcinoma.
Cancer Immunol Immunother. 61:1977–1987. 2012. View Article : Google Scholar : PubMed/NCBI
|