Introduction
Von Hippel-Lindau (VHL) disease is an autosomal
dominant hereditary syndrome caused by germline mutations in the
VHL tumor suppressor gene, which is located on chromosome
3p25-26. The clinical phenotype of VHL disease is characterized by
the development of a panel of benign and malignant, highly
vascularized tumors in several organ systems. One of the major
clinical manifestations is clear-cell renal cell carcinoma (ccRCC).
The emergence of tumors follows the inactivation of the remaining
wild-type allele. Somatic VHL inactivation is also a
hallmark of the majority of cases of sporadic ccRCC, and insights
into VHL gene function have resulted in the use of
antiangiogenic targeted therapies, which are now first line in the
treatment of advanced renal tumors (1). Fuhrman's nuclear grade is the
cornerstone of the prognostic classification of ccRCC and is based
on increasing nuclear size, irregularity and nucleolar prominence
(2).
In the last few years, numerous pan-genomic studies
[comparative genomic hybridization-array, and gene and microRNA
(miRNA/miR) expression profiles] have well characterized sporadic
ccRCC (3–7). However, few studies have been
published in the field of VHL-associated ccRCC (8,9).
Fisher et al demonstrated that kidney tumors developing from
a germline VHL mutation exhibit complementarity of the evolutionary
principles of contingency and convergence. Notably, reduced
mutation burden and limited evidence of intra-tumor heterogeneity
were detected in these tumors (10). To the best of our knowledge, no
previous study has focused on miRNA profiles in VHL-associated
renal tumors compared with in sporadic ccRCC; these two entities
are considered similar but no transcriptomic comparison study has
been conducted to confirm this fact. The main function of miRNAs is
to suppress the translation of target genes; however, they can also
process mRNAs for cellular decay (11). Mature miRNAs have numerous targets,
are often members of the same regulatory networks, and operate in
regulatory feedback loops. These properties position them as
fine-tuning modulators of set points in homeostatic processes in
normal cells. It has previously been reported that discrete sets of
miRNAs are induced and repressed in various types of cancer, and
are specific to particular diagnoses and progression patterns, and
predictive of responses to treatment. In cancer, aberrations in the
expression of specific miRNAs may have well-defined
tumor-suppressing or oncogenic functions (12). Furthermore, since a given miRNA has
several targets, multiple pro-oncogenic or tumor-suppressing
pathways are affected, and these pathways, in turn, regulate the
miRNA expression in a feedback-loop mechanism (13). Studies regarding miRNA
dysregulation in cancer have risen rapidly recently, including
those in sporadic ccRCC (14,15).
The transcriptomic analysis of synchronous tumors
occurring within the kidney in one patient offers a rare
opportunity to investigate the evolution of tumors. In the present
study, to better understand the biological processes implicated in
the tumorigenesis of VHL-associated ccRCC, the transcriptomic
(miRNA and mRNA) signature of VHL-associated tumors was determined
using multiple tumor samples from two distinct patients who
possessed several different primary kidney tumors. For comparison,
the miRNA and mRNA profiles of 12 independent VHL-associated tumors
were determined and were compared with the profiles of 22 sporadic
renal tumors. The present study may provide information regarding
the molecular pathogenesis of ccRCC in patients with VHL disease
and offer possibilities for further molecular investigations.
Materials and methods
Patient samples and ethical consent
A total of 36 patients were recruited between 2002
and 2009. Their mean age at diagnosis was 54.9 years old. The
sample series, which comprised two sets of 13 (from 2 patients) and
34 (from 34 patients) human ccRCC samples were obtained from the
French Kidney Cancer Consortium coordinated by Professor Stéphane
Richard (French National Network for Rare Cancers in Adults PREDIR
Center, Bicêtre Hospital, Le Kremlin-Bicêtre, France). The present
study was approved by the ethical committee of Bicêtre Hospital (Le
Kremlin-Bicêtre, France). Primary tumors, and for some cases
adjacent non-tumor samples, were obtained from patients who
underwent surgical tumor resection. All patients provided written
informed consent prior to surgery for use of their tumors. Tumor
samples were frozen immediately in liquid nitrogen following
surgery and were classified according to the Fuhrman nuclear
grading system, after which they were grouped into low grade (grade
1+2) and high grade (grade 3+4) tumors (2).
The main clinical and genetic features of the
patients, and tumor characteristics, are described in Table I. Part of the tumor series was
previously reported (16).
Differences in the numbers of samples are due to the lack of
sufficient RNA quantity.
 | Table ICharacteristics of tumor samples. |
Table I
Characteristics of tumor samples.
| Tumor number | Hospital | Sex | Age (years) | Histology | Fuhrman's
grade | Grade class | VHL
status | microRNA microarray
analysis | Gene microarray
analysis |
|---|
| First set | | | | | | | | | |
| 2203_T1 | Necker | M | 61 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| 2203_T10 | Necker | M | 61 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| 2203_T3 | Necker | M | 61 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| 2203_T6 | Necker | M | 61 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| 2203_T7 | Necker | M | 61 | VHL-ccRCC | 3 | High | Mutated | Yes | Yes |
| 2203_T9 | Necker | M | 61 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| 1674_T1 | Necker | F | 39 | VHL-ccRCC | 3 | High | Mutated | Yes | Yes |
| 1674_T11 | Necker | F | 39 | VHL-ccRCC | 3 | High | Mutated | Yes | Yes |
| 1674_T13 | Necker | F | 39 | VHL-ccRCC | 2 | Low | Mutated | | Yes |
| 1674_T19 | Necker | F | 39 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| 1674_T2 | Necker | F | 39 | VHL-ccRCC | 3 | High | Mutated | Yes | Yes |
| 1674_T3 | Necker | F | 39 | VHL-ccRCC | 3 | High | Mutated | Yes | Yes |
| 1674_T4 | Necker | F | 39 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| Second set | | | | | | | | | |
| 1919 | St-Joseph | F | 75 | Sporadic ccRCC | 2 | Low | Wild-type | Yes | Yes |
| 2040 | Necker | F | 45 | Sporadic ccRCC | 3 | High | Wild-type | Yes | Yes |
| 3042 | St-Joseph | M | 83 | Sporadic ccRCC | 2 | Low | Mutated | Yes | Yes |
| 3503 | St-Joseph | M | 59 | Sporadic ccRCC | 4 | High | Wild-type | Yes | Yes |
| 3554 | St-Joseph | M | 47 | Sporadic ccRCC | 3 | High | Mutated | Yes | Yes |
| 3559 | St-Joseph | M | 61 | Sporadic ccRCC | 3 | High | Wild-type | Yes | Yes |
| 4320 | St-Joseph | F | 78 | Sporadic ccRCC | 3 | High | Mutated | Yes | Yes |
| 4667 | St-Joseph | F | 70 | Sporadic ccRCC | 3 | High | Mutated | Yes | Yes |
| 5290 | St-Joseph | F | 69 | Sporadic ccRCC | 2 | Low | Mutated | Yes | Yes |
| 5668 | St-Joseph | M | 69 | Sporadic ccRCC | 2 | Low | Mutated | Yes | Yes |
| 5835 | St-Joseph | M | 60 | Sporadic ccRCC | 2 | Low | Mutated | Yes | Yes |
| 5887 | St-Joseph | M | 85 | Sporadic ccRCC | 2 | Low | Mutated | Yes | Yes |
| 6517 | St-Joseph | M | 76 | Sporadic ccRCC | 2 | Low | Mutated | Yes | Yes |
| 6739 | St-Joseph | M | 53 | Sporadic ccRCC | 2 | Low | Mutated | Yes | Yes |
| 7294 | St-Joseph | F | 65 | Sporadic ccRCC | 4 | High | Mutated | Yes | Yes |
| 7896 | St-Joseph | F | 70 | Sporadic ccRCC | 2 | Low | Mutated | Yes | Yes |
| 8527 | St-Joseph | F | 77 | Sporadic ccRCC | 2 | Low | Mutated | Yes | Yes |
| 9490 | St-Joseph | M | 67 | Sporadic ccRCC | 4 | High | Mutated | Yes | Yes |
| 9671 | St-Joseph | F | 45 | Sporadic ccRCC | 4 | High | Mutated | Yes | Yes |
| 40442 | St-Joseph | M | 56 | Sporadic ccRCC | 3 | High | Wild-type | – | Yes |
| 40815 | St-Joseph | M | 71 | Sporadic ccRCC | 3 | High | Wild-type | Yes | Yes |
| 40842 | St-Joseph | F | 77 | Sporadic ccRCC | 1 | Low | Mutated | Yes | Yes |
| 2132 | Necker | M | 34 | VHL-ccRCC | 2 | Low | Mutated | – | Yes |
| 2920 | Bicêtre | M | 26 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| 4573 | Bicêtre | M | 24 | VHL-ccRCC | 1 | Low | Mutated | Yes | Yes |
| 4734 | Bicêtre | M | 65 | VHL-ccRCC | 3 | High | Mutated | Yes | Yes |
| 5205 | Necker | M | 27 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| 6315 | Bicêtre | M | 35 | VHL-ccRCC | 3 | High | Mutated | Yes | Yes |
| 6434 | Necker | F | 40 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| 6600 | Necker | M | 23 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| 7000 | Necker | F | 45 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| 8156 | Necker | F | 40 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| 8464 | Bicêtre | M | 28 | VHL-ccRCC | 2 | Low | Mutated | Yes | Yes |
| 50201 | Necker | M | 31 | VHL-ccRCC | 3 | High | Mutated | Yes | Yes |
Tumor cryosections and total RNA
extraction
All samples were frozen at −80°C prior to RNA
extraction. The percentage of malignant tumor cell content was
determined in the first and last sections obtained using a
cryostat, and sections with >60% malignant tumor cell content
were used for subsequent experimentation (mean of all the series:
83±12%). Total RNA was isolated using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
according to the manufacturer's protocol. Nucleic acid
concentration and purity were determined using NanoDrop ND-1000
(NanoDrop; Thermo Fisher Scientific, Inc., Wilmington, DE, USA),
and RNA quality was verified using a 2100 Bioanalyser (Agilent
Technologies, Inc., Santa Clara, CA, USA) using an RNA integrity
no. >6. The reference sample was based on a pool of RNA
extracted from all normal adjacent tissues available, which
consisted of 17 renal tissue samples from patients with sporadic
ccRCC, and was used for all analyses.
miRNA and gene microarray expression
analysis
Each sample was prepared according to the Agilents
miRNA Microarray system protocol (Agilent Technologies, Inc.).
Total RNA (100 ng) was labeled and hybridized to Agilent human
miRNA 8×15K microarrays v3 (AMADID 21827; Agilent Technologies,
Inc.) containing 851 human and 88 human viral miRNAs, each
replicated 16 times, and Agilent human genome 4×44K microarrays
(Agilent Technologies, Inc.) for gene expression, according to the
manufacturer's protocol. All processing methods used for miRNA
analyses were performed on the Cy3 Median Signal in Agilent Feature
software v10.7 (Agilent Technologies, Inc.). For gene expression,
analyses were performed on the Cy3 and Cy5 Median signal in Agilent
Feature software v10.7 (Agilent Technologies, Inc.). Raw data files
were extracted using functions in Bioconductor (17). Flagged spots, as well as control
spots, were systematically removed, and data were log2
transformed. Quantile normalization was performed using the
normalizeBetweenArray function from R package 'LIMMA' (version no.
3.34.9) (18). The median of each
probe for a given miRNA was computed and the corresponding value
was assigned to the miRNA. Data were then filtered according to the
maximum number of missing values allowed for each miRNA (30%).
Hierarchical clusters were computed using the 'dist'
function from R, using the 'Euclidian' method as a measure of
distance. Hierarchical clustering was performed using the 'hclust'
function from R using the distance matrix previously computed and
Ward's method.
To assess differentially expressed miRNAs, the
fold-changes and standard errors were initially estimated between
two groups of samples by fitting a linear model for each miRNA with
the 'lmFit' function of LIMMA package. Subsequently, empirical
Bayes smoothing was applied to the standard errors in the linear
model previously computed using the 'eBayes' function of LIMMA. To
extract a table of the top-ranked genes from the linear model fit,
the topTable function in LIMMA was utilized. The results were saved
in a table file format.
BRB analyses were performed to compare the tumor
groups and the reference group. miRNAs and genes that were
significantly differentially expressed between the groups at
P<0.05 using BRB ArrayTools v2 (https://brb.nci.nih.gov/BRB-ArrayTools/), as
determined using two-way analysis of variance and multiple
correction Benjamini-Hochberg test, were selected for further
analysis. Significantly differentially expressed miRNAs were used
to build a hierarchical cluster using Gene-E (https://software.broadinstitute.org/GENE-E/index.html).
For biological interpretation of significant genes, the Database
for Annotation, Visualization and Integrated Discovery (DAVID,
https://david.ncifcrf.gov/) was employed
to perform the Kyoto Encyclopedia of Genes and Genomes (KEGG)
pathway analysis. Significant KEGG pathways (P<0.05) were
selected in VHL-associated and sporadic ccRCC tumors.
Results
Patients and samples
The main characteristics of the two tumor sets used
in the present study and individual patient data are reported in
Table I. The first set consisted
of several samples from multiple tumors obtained from two patients
with VHL disease [Patient 2203, n=6; Patient 1674, n=7 (6 for miRNA
analysis and 7 for mRNA analysis)]. The second set consisted of 12
VHL-associated renal tumors (11 for miRNA analysis and 12 for mRNA
analysis) and 22 sporadic ccRCC tumors (21 for miRNA analysis and
22 for mRNA analysis) from 34 patients.
Lack of heterogeneity for miRNA and mRNA
expression profiles between multiple tumors for the same patient
with VHL disease
Patients affected with VHL disease can
simultaneously develop several multifocal and bilateral tumors in
the kidneys. The present study performed microarray gene and miRNA
expression analyses on several samples from different tumors
obtained from the same patient, for two independent patients
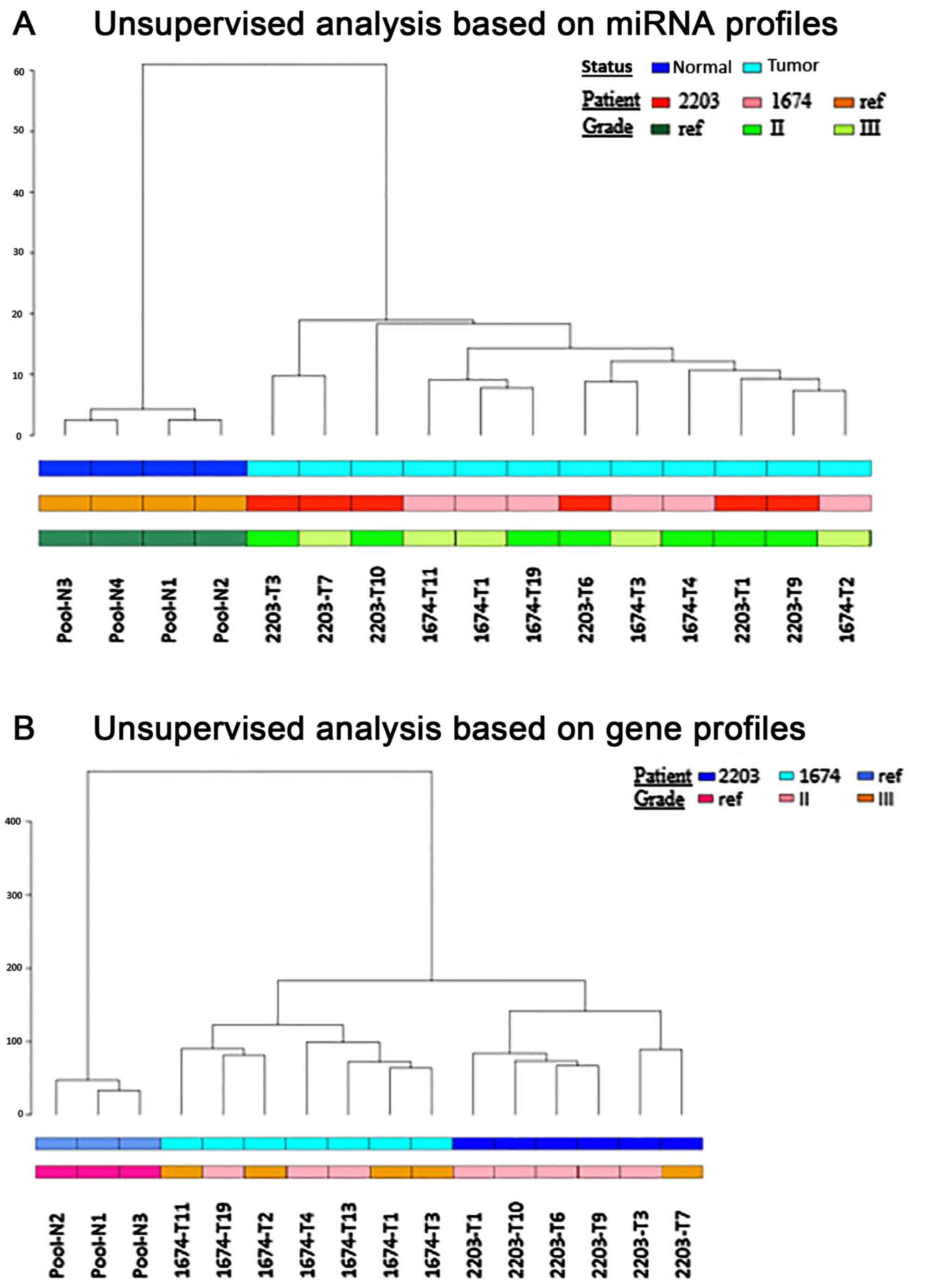
(Patients 2203 and 1674). Firstly, unsupervised hierarchical
clustering analyses were conducted on the miRNA and gene expression
profiles. In both analyses, tumor samples were well separated from
the normal renal tissue pool. In addition, unsupervised
hierarchical clustering of the miRNA expression profiles was not
able to discriminate between the two patients (Fig. 1A); however, unsupervised
hierarchical clustering of the mRNA expression profiles was able to
distinguish between the two patients; this may be explained by the
different genetic background (Fig.
1B).
The present study also explored the differences
between tumor samples and the normal reference samples, for each
patient. For patient 2203, a total of 1,377 genes and 51 miRNAs
were significantly differentially expressed. For patient 1674, a
total of 1,282 genes and 56 miRNAs were differentially expressed
(Table II). In addition, as shown
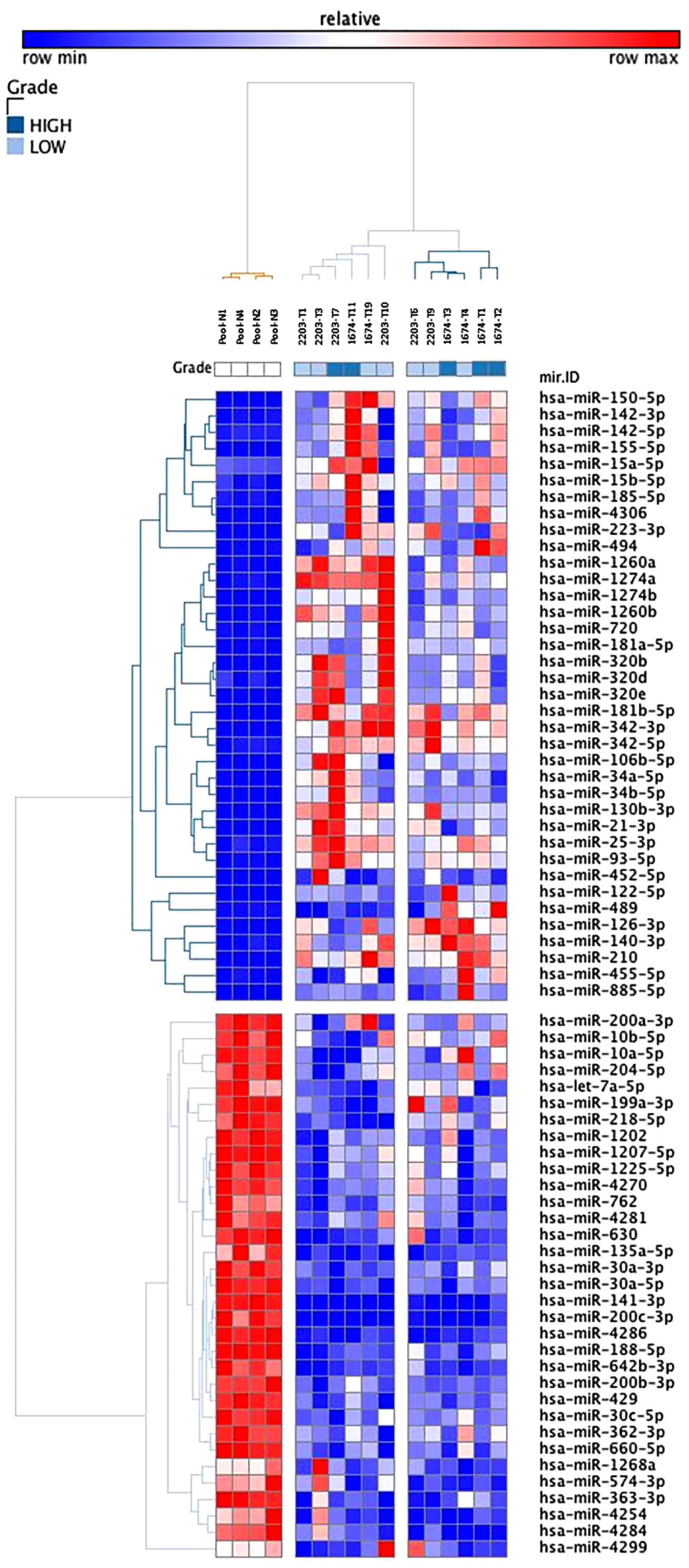
in Fig. 2, a hierarchical cluster
analysis built with 70 dysregulated miRNAs clearly indicated
separate tumor clusters from the normal reference group. No obvious
differences were detected between the samples, but the two
subclusters could be distinguished according to the nuclear grade
of these tumors. When tumors were compared two by two, no
significant difference was found, thus supporting the hypothesis of
a similar molecular evolution between independent tumors. KEGG
biological pathway analysis was conducted using DAVID software.
This analysis globally identified three classes of pathways that
were overrepresented with dysregulated genes. Notably, in the two
patients, the most significant pathways were similar, and were
implicated in 'immunity' and 'metabolism' (Table III).
| Table IIDysregulated miRNAs in patients 2203
and 1674. |
Table II
Dysregulated miRNAs in patients 2203
and 1674.
| miRNA ID | Patient 2203 vs.
normal pool
| Patient 1674 vs.
normal pool
|
|---|
| Fold-change | P-value | adj-P-value | Fold-change | P-value | adj-P-value |
|---|
| Common miRNAs | | | | | | |
| Upregulated | | | | | | |
| hsa-miR-210 | 13.694 |
4.119×10−09 |
1.110×10−06 | 18.204 |
1.344×10−09 |
9.051×10−07 |
| hsa-miR-155-5p | 6.695 |
7.973×10−06 |
4.130×10−04 | 7.965 |
2.615×10−04 |
2.887×10−03 |
| hsa-miR-342-3p | 3.389 |
1.927×10−06 |
1.527×10−04 | 3.132 |
1.483×10−07 |
1.537×10−05 |
| hsa-miR-34a-5p | 3.373 |
2.187×10−04 |
3.983×10−03 | 2.519 |
7.045×10−04 |
5.858×10−03 |
| hsa-miR-1274a | 2.853 |
1.377×10−04 |
2.995×10−03 | 2.441 |
3.774×10−05 |
7.159×10−04 |
| hsa-miR-720 | 2.773 |
9.353×10−05 |
2.348×10−03 | 2.265 |
9.233×10−05 |
1.353×10−03 |
| hsa-miR-122-5p | 2.696 |
9.032×10−06 |
4.345×10−04 | 3.808 |
4.595×10−05 |
8.253×10−04 |
| hsa-miR-150-5p | 2.645 |
1.554×10−04 |
3.271×10−03 | 3.333 |
7.686×10−05 |
1.204×10−03 |
| hsa-miR-1274b | 2.531 |
6.167×10−04 |
7.416×10−03 | 2.235 |
1.829×10−05 |
4.663×10−04 |
| hsa-miR-34b-5p | 2.472 |
1.131×10−03 |
1.199×10−02 | 2.239 |
7.917×10−05 |
1.226×10−03 |
|
hsa-miR-106b-5p | 2.463 |
8.830×10−03 |
4.815×10−02 | 2.191 |
1.354×10−06 |
7.601×10−05 |
| hsa-miR-25-3p | 2.361 |
5.440×10−05 |
1.629×10−03 | 2.343 |
3.438×10−07 |
2.724×10−05 |
| hsa-miR-93-5p | 2.342 |
3.627×10−05 |
1.164×10−03 | 2.209 |
1.910×10−07 |
1.838×10−05 |
| hsa-miR-885-5p | 2.059 |
7.217×10−05 |
1.984×10−03 | 2.749 |
4.733×10−04 |
4.438×10−03 |
| Downregulated | | | | | | |
| hsa-miR-762 | −2.148 |
6.373×10−04 |
7.597×10−03 | −2.456 |
3.143×10−06 |
1.520×10−04 |
| hsa-miR-4270 | −2.167 |
1.655×10−03 |
1.538×10−02 | −2.318 |
7.754×10−06 |
2.611×10−04 |
| hsa-miR-218-5p | −2.259 |
1.106×10−04 |
2.701×10−03 | −2.030 |
3.164×10−03 |
1.661×10−02 |
| hsa-miR-30a-3p | −2.371 |
3.591×10−05 |
1.164×10−03 | −2.185 |
6.318×10−05 |
1.051×10−03 |
|
hsa-miR-1207-5p | −2.380 |
2.682×10−03 |
2.101×10−02 | −2.425 |
7.777×10−04 |
6.349×10−03 |
|
hsa-miR-199a-3p | −2.382 |
5.068×10−03 |
3.354×10−02 | −3.405 |
6.913×10−03 |
3.053×10−02 |
| hsa-miR-188-5p | −2.474 |
3.029×10−05 |
1.046×10−03 | −2.172 |
3.194×10−06 |
1.520×10−04 |
| hsa-miR-30c-5p | −2.477 |
2.188×10−04 |
3.983×10−03 | −2.108 |
3.468×10−05 |
6.970×10−04 |
|
hsa-miR-1225-5p | −2.485 |
3.166×10−03 |
2.331×10−02 | −2.403 |
7.302×10−04 |
6.034×10−03 |
| hsa-miR-4286 | −2.487 |
1.325×10−09 |
7.936×10−07 | −2.279 |
6.848×10−08 |
1.004×10−05 |
| hsa-miR-30a-5p | −2.490 |
8.286×10−06 |
4.134×10−04 | −2.253 |
5.463×10−06 |
2.230×10−04 |
| hsa-miR-4284 | −2.718 |
3.360×10−03 |
2.446×10−02 | −4.295 |
7.493×10−06 |
2.611×10−04 |
| hsa-miR-660-5p | −2.813 |
4.062×10−05 |
1.244×10−03 | −2.239 |
4.978×10−04 |
4.530×10−03 |
|
hsa-miR-642b-3p | −2.863 |
3.629×10−05 |
1.164×10−03 | −2.775 |
4.850×10−08 |
1.004×10−05 |
|
hsa-miR-135a-5p | −2.965 |
1.202×10−06 |
1.233×10−04 | −2.413 |
3.352×10−06 |
1.520×10−04 |
| hsa-miR-630 | −3.208 |
2.354×10−03 |
1.880×10−02 | −3.347 |
3.092×10−08 |
8.330×10−06 |
| hsa-miR-363-3p | −3.319 |
2.369×10−04 |
4.200×10−03 | −2.378 |
1.989×10−05 |
4.871×10−04 |
|
hsa-miR-200b-3p | −3.577 |
5.841×10−05 |
1.684×10−03 | −2.609 |
2.844×10−05 |
5.918×10−04 |
| hsa-miR-429 | −3.657 |
1.300×10−04 |
2.917×10−03 | −2.893 |
2.199×10−05 |
5.197×10−04 |
| hsa-miR-1202 | −3.772 |
6.780×10−04 |
7.613×10−03 | −2.993 |
1.971×10−03 |
1.149×10−02 |
| hsa-miR-141-3p | −9.901 |
1.767×10−09 |
7.936×10−07 | −9.272 |
9.951×10−07 |
5.828×10−05 |
|
hsa-miR-200c-3p | −12.485 |
3.817×10−09 |
1.110×10−06 | −11.832 |
9.017×10−07 |
5.784×10−05 |
| Specific miRNAs of
patient 2203 | | | | | | |
| Upregulated | | | | | | |
| hsa-miR-21-3p | 2.705 |
1.544×10−05 |
6.710×10−04 | 1.845 |
1.092×10−03 |
7.619×10−03 |
| hsa-miR-1260b | 2.461 |
2.786×10−04 |
4.750×10−03 | 1.968 |
1.898×10−04 |
2.265×10−03 |
| hsa-miR-320d | 2.359 |
1.634×10−04 |
1.538×10−02 | 1.756 |
3.132×10−03 |
1.654×10−02 |
|
hsa-miR-181b-5p | 2.339 |
4.412×10−08 |
9.083×10−06 | 1.983 |
3.519×10−05 |
6.970×10−04 |
| hsa-miR-1260a | 2.261 |
6.489×10−04 |
7.613×10−03 | 1.922 |
8.246×10−04 |
6.573×10−03 |
| hsa-miR-452-5p | 2.245 |
4.431×10−03 |
3.045×10−02 | 1.382 |
1.265×10−02 |
4.801×10−02 |
|
hsa-miR-130b-3p | 2.243 |
1.656×10−06 |
1.394×10−04 | 1.742 |
2.452×10−06 |
1.270×10−04 |
| hsa-miR-320e | 2.178 |
5.735×10−04 |
7.087×10−03 | 1.837 |
2.525×10−05 |
5.755×10−04 |
| hsa-miR-320b | 2.168 |
7.956×10−04 |
8.856×10−03 | 1.714 |
2.918×10−04 |
3.144×10−03 |
| hsa-miR-342-5p | 2.129 |
1.601×10−05 |
6.741×10−04 | 1.971 |
1.339×10−07 |
1.503×10−05 |
| hsa-miR-223-3p | 2.056 |
2.665×10−04 |
4.602×10−03 | 1.867 |
9.326×10−03 |
3.830×10−02 |
|
hsa-miR-181a-5p | 2.019 |
9.307×10−04 |
1.011×10−02 | 1.932 |
1.444×10−05 |
4.052×10−04 |
| Downregulated | | | | | | |
| hsa-miR-362-3p | −2.317 |
5.680×10−04 |
7.087×10−03 | −1.826 |
2.457×10−03 |
1.385×10−02 |
|
hsa-miR-200a-3p | −2.552 |
1.657×10−04 |
3.433×10−03 | −1.575 |
3.224×10−02 | ns |
| hsa-miR-10a-5p | −3.326 |
4.392×10−04 |
5.915×10−03 | −1.965 |
4.617×10−02 | ns |
| Specific miRNAs of
patient 1674 | | | | | | |
| Upregulated | | | | | | |
| hsa-miR-142-3p | 2.941 |
2.613×10−02 | ns | 4.372 |
1.588×10−03 |
9.811×10−03 |
| hsa-miR-489 | 1.231 | ns | ns | 3.434 |
1.139×10−02 |
4.447×10−02 |
| hsa-miR-142-5p | 2.116 | ns | ns | 3.205 |
1.006×10−03 |
7.348×10−03 |
| hsa-miR-494 | 1.975 |
1.863×10−03 |
1.647×10−02 | 2.621 |
1.116×10−03 |
7.709×10−03 |
| hsa-miR-126-3p | 1.940 |
1.606×10−03 |
1.538×10−02 | 2.191 |
1.047×10−04 |
1.454×10−03 |
| hsa-miR-15a-5p | 1.569 | ns | ns | 2.187 |
1.493×10−05 |
4.103×10−04 |
| hsa-miR-15b-5p | 1.045 |
1.237×10−02 | ns | 2.181 |
1.112×10−03 |
1.488×10−03 |
| hsa-miR-455-5p | 1.230 | ns | ns | 2.096 |
4.226×10−05 |
7.798×10−04 |
| hsa-miR-140-3p | 1.840 |
5.298×10−04 |
6.796×10−03 | 2.032 |
1.710×10−04 |
2.105×10−03 |
| hsa-miR-4306 | 1.368 |
3.099×10−03 |
2.306×10−02 | 2.005 |
3.613×10−04 |
3.632×10−03 |
| hsa-miR-185-5p | 1.387 |
2.402×10−02 | ns | 2.005 |
9.862×10−04 |
7.339×10−03 |
| Downregulated | | | | | | |
| hsa-miR-4299 | −1.489 | ns | ns | −2.035 |
1.415×10−05 |
4.052×10−04 |
| hsa-miR-4254 | −1.879 |
3.745×10−03 |
2.660×10−02 | −2.081 |
3.636×10−06 |
1.531×10−04 |
| hsa-miR-204-5p | −5.917 |
2.029×10−02 | ns | −2.12 |
8.344×10−03 |
3.491×10−02 |
| hsa-miR-1268a | −1.486 | ns | ns | −2.264 |
1.260×10−05 |
3.689×10−04 |
| hsa-miR-574-3p | −1.526 |
3.078×10−02 | ns | −2.314 |
9.952×10−07 |
5.828×10−05 |
| hsa-miR-10b-5p | −1.881 |
1.377×10−02 | ns | −2.338 |
8.231×10−03 |
3.463×10−02 |
| hsa-let-7a-5p | −1.933 |
1.263×10−03 |
1.288×10−02 | −2.377 |
1.679×10−03 |
1.014×10−02 |
| hsa-miR-4281 | −1.917 |
7.692×10−03 |
4.525×10−02 | −2.529 |
8.819×10−05 |
1.320×10−03 |
| Table IIISummary of major implicated pathways
in VHL-associated and sporadic renal tumors. |
Table III
Summary of major implicated pathways
in VHL-associated and sporadic renal tumors.
| KEGG ID | KEGG
description | KEGG subclass | KEGG class |
|---|
| Common pathways in
both tumor groups | | | |
| hsa04115 | p53 signaling
pathway | Cell growth and
death | Cellular
processes |
| hsa04510 | Focal adhesion | Cellular
community | |
| hsa04540 | Gap junction | | |
| hsa04060 | Cytokine-cytokine
receptor interaction | Signaling molecules
and interaction | Environmental
information processing |
| hsa04514 | Cell adhesion
molecules (CAMs) | | |
| hsa04512 | ECM-receptor
interaction | | |
| hsa05200 | Pathways in
cancer | Cancers:
Overview | Human diseases |
| hsa03320 | PPAR signaling
pathway | Endocrine
system | Organismal
systems |
| hsa04960 |
Aldosterone-regulated sodium
reabsorption | Excretory
system | |
| hsa04610 | Complement and
coagulation cascades | Immune system | |
| hsa04650 | Natural killer cell
mediated cytotoxicity | | |
| hsa04640 | Hematopoietic cell
lineage | | |
| hsa04672 | Intestinal immune
network for IgA production | | |
| hsa04610 | Complement and
coagulation cascades | | |
| hsa04062 | Chemokine signaling
pathway | | |
| hsa04612 | Antigen processing
and presentation | | |
| hsa04621 | NOD-like receptor
signaling pathway | | |
| hsa04660 | T cell receptor
signaling pathway | | |
| hsa04670 | Leukocyte
transendothelial migration | | |
| hsa00532 | Chondroitin sulfate
biosynthesis | Glycan biosynthesis
and metabolism | Metabolism |
| hsa00280 | Valine, leucine and
isoleucine degradation | Amino acid
metabolism | |
| hsa00380 | Tryptophan
metabolism | | |
| hsa00330 | Arginine and
proline metabolism | | |
| hsa00260 | Glycine, serine and
threonine metabolism | | |
| hsa00250 | Alanine, aspartate
and glutamate metabolism | | |
| hsa00340 | Histidine
metabolism | | |
| hsa00310 | Lysine
degradation | | |
| hsa00270 | Cysteine and
methionine metabolism | | |
| hsa00350 | Tyrosine
metabolism | | |
| hsa00640 | Propanoate
metabolism | Carbohydrate
metabolism | |
| hsa00650 | Butanoate
metabolism | | |
| hsa00620 | Pyruvate
metabolism | | |
| hsa00020 | Citrate cycle (TCA
cycle) | | |
| hsa00053 | Ascorbate and
aldarate metabolism | | |
| hsa00010 |
Glycolysis/gluconeogenesis | | |
| hsa00040 | Pentose and
glucuronate interconversions | | |
| hsa00500 | Starch and sucrose
metabolism | | |
| hsa00630 | Glyoxylate and
dicarboxylate metabolism | | |
| hsa00190 | Oxidative
phosphorylation | Energy
metabolism | |
| hsa00910 | Nitrogen
metabolism | | |
| hsa00071 | Fatty acid
metabolism | Global and overview
maps | |
| hsa00072 | Synthesis and
degradation of ketone bodies | Lipid
metabolism | |
| hsa00140 | Steroid hormone
biosynthesis | | |
| hsa00120 | Primary bile acid
biosynthesis | | |
| hsa00590 | Arachidonic acid
metabolism | | |
| hsa00062 | Fatty acid
elongation in mitochondria | | |
| hsa00830 | Retinol
metabolism | Metabolism of
cofactors and vitamins | |
| hsa00410 | β-Alanine
metabolism | Metabolism of other
amino acids | |
| hsa00480 | Glutathione
metabolism | | |
| hsa00903 | Limonene and pinene
degradation | Metabolism of
terpenoids and polyketides | |
| hsa00982 | Drug
metabolism-cytochrome P450 | Xenobiotics
biodegradation and metabolism | |
| hsa00980 | Metabolism of
xenobiotics | | |
| by cytochrome
P450 | | |
| hsa00983 | Drug
metabolism-other enzymes | | |
| Specific to
VHL-associated tumors | | | |
| hsa04330 | Notch signaling
pathway | Signal
transduction | Environmental
information processing |
| hsa00900 | Terpenoid backbone
biosynthesis | Metabolism of
terpenoids and polyketides | Metabolism |
| Specific to
sporadic tumors | | | |
| hsa04110 | Cell cycle | Cell growth and
death | Cellular
processes |
| hsa04630 | Jak-STAT signaling
pathway | Signal
transduction | Environmental
information processing |
| hsa04666 | FcγR-mediated
phagocytosis | Immune system | Organismal
systems |
| hsa04662 | B cell receptor
signaling pathway | | |
| hsa04620 | Toll-like receptor
signaling pathway | | |
| hsa01040 | Biosynthesis of
unsaturated fatty acids | Lipid
metabolism | Metabolism |
| hsa00100 | Steroid
biosynthesis | | |
| hsa00360 | Phenylalanine
metabolism | Amino acid
metabolism | |
miRNA expression levels distinguish
VHL-associated tumors from sporadic ccRCC
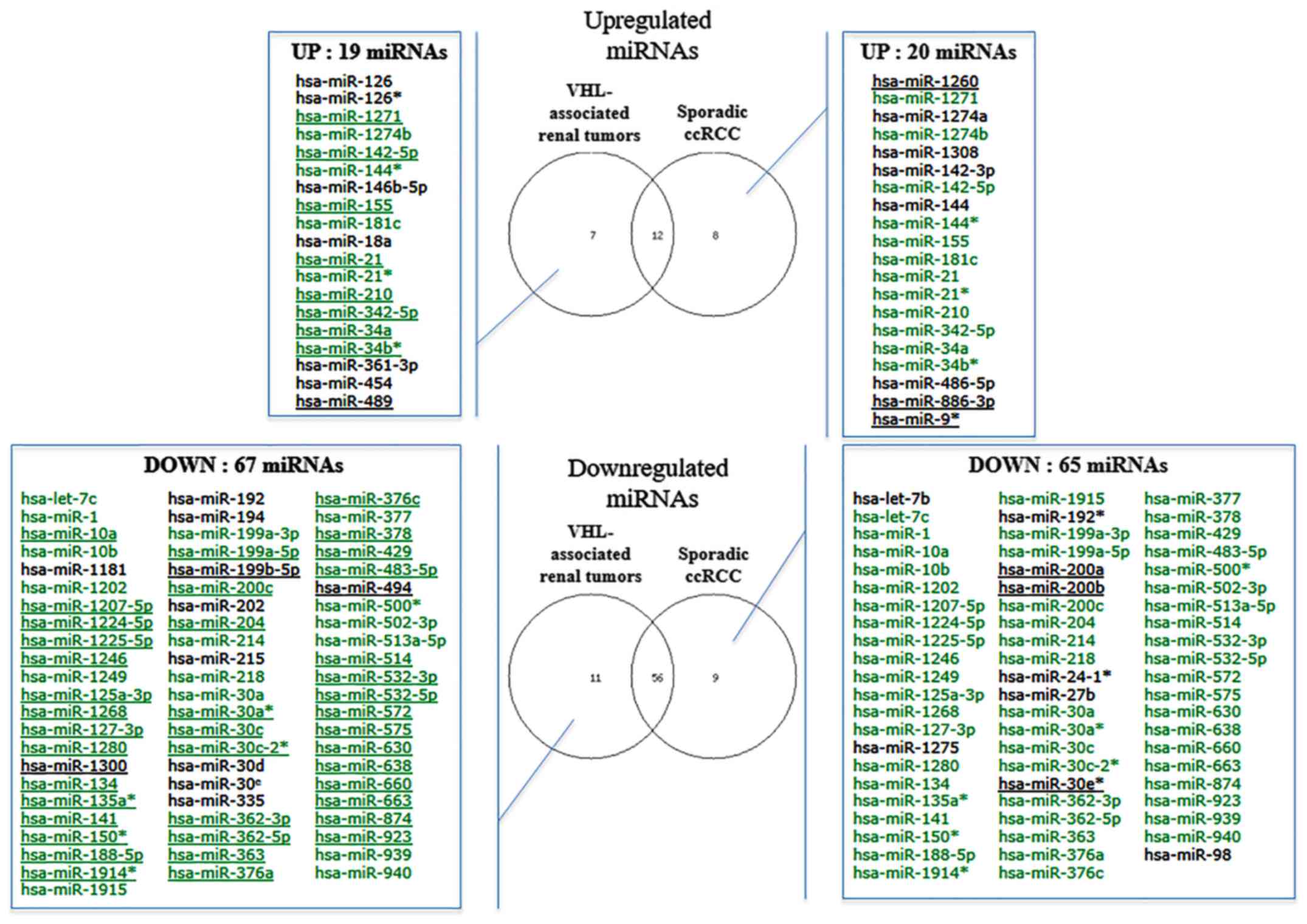
Using microarray analysis, a total of 103 miRNAs
were identified as differentially expressed among the
VHL-associated and sporadic ccRCC samples (fold-change <−2 or
>2) compared with in the normal reference group (Fig. 3). These differentially expressed
miRNAs are similar to those described in the first set of samples.
Two thirds of miRNAs (12 upregulated and 56 downregulated) were
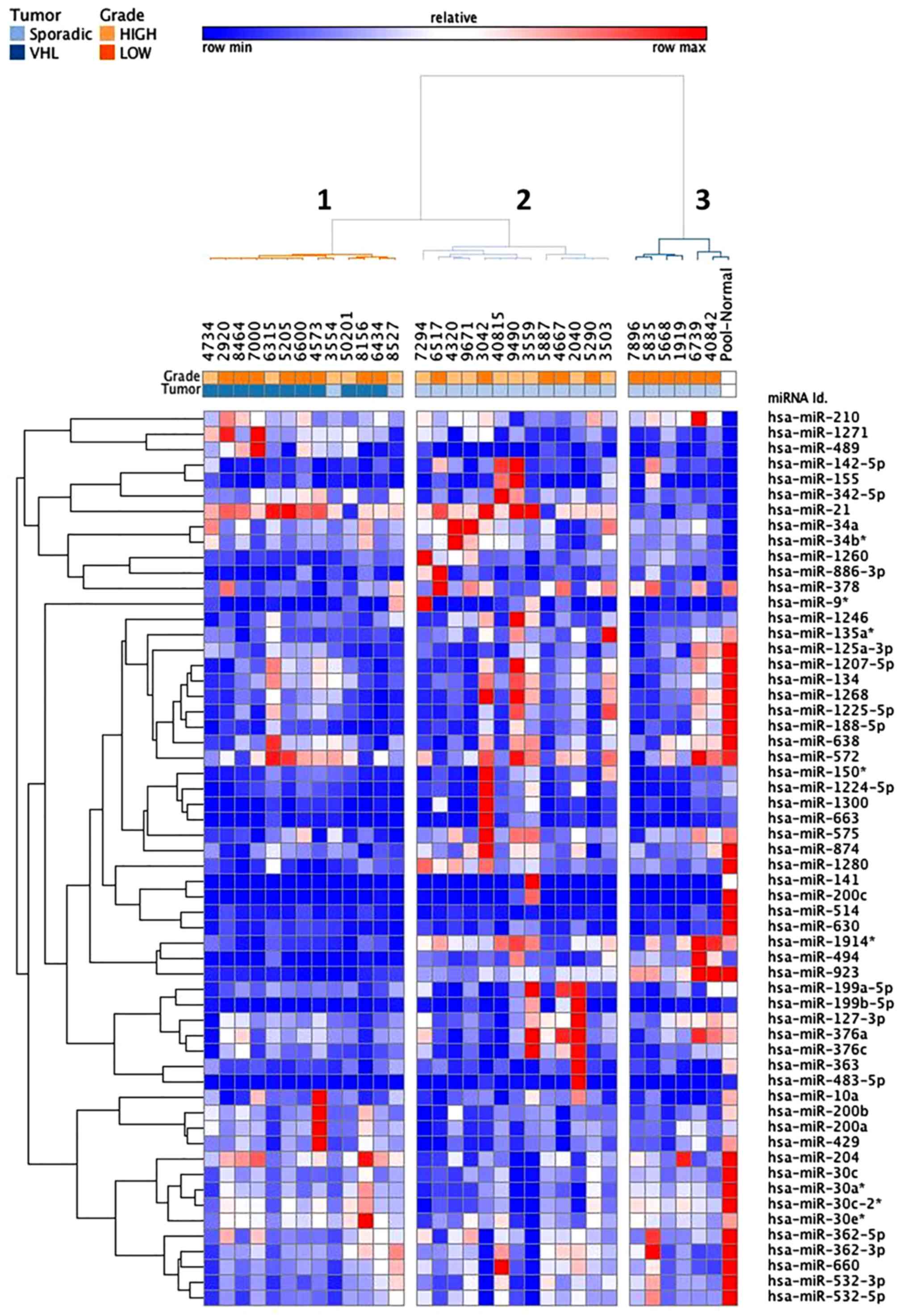
common to both groups. Hierarchical cluster analysis, based on the
58 most differentially expressed miRNAs (fold-change <-3 or
>3), revealed three clusters that mainly define VHL-associated
(cluster 1) and sporadic (clusters 2 and 3) specimen profiles
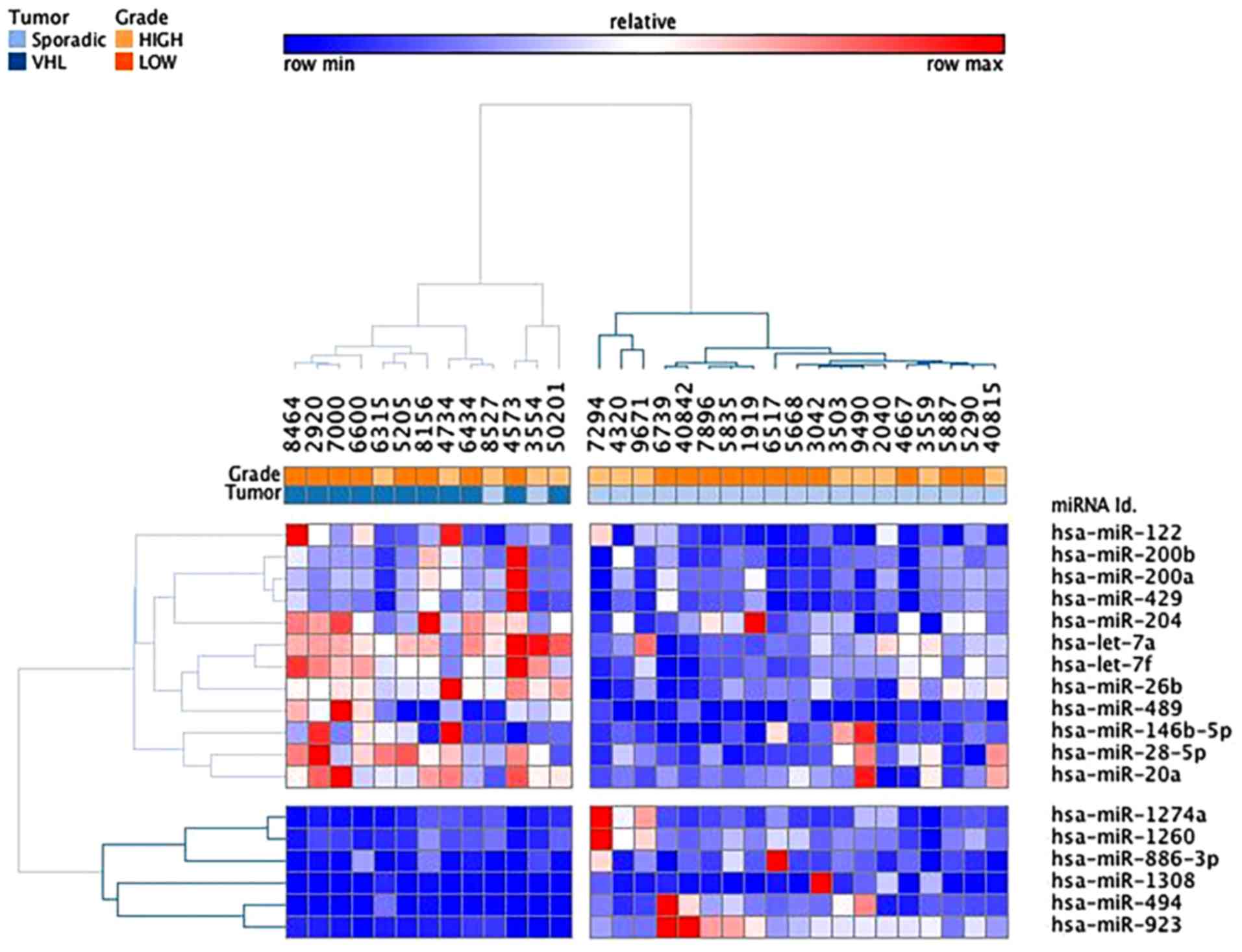
(Fig. 4). Of the 21 sporadic ccRCC
samples, only two samples were allocated to the VHL-related branch.
Branches 2 and 3 were able to distinguish high and low grades of
sporadic ccRCC, respectively. The VHL-associated tumors formed a
separate subcluster (cluster 1), thus indicating that their miRNA
expression levels were different from those of sporadic tumor
samples. In addition, supervised analysis directly comparing the
VHL-associated and sporadic tumors (fold-change <−1.5 or
>1.5; raw P<0.05) identified 18 differentially expressed
miRNAs (Table IV and Fig. 5). Taken together, these analyses
indicated that, even though some differentially expressed miRNAs
were similar between the two tumor groups, it was possible to
distinguish between these two groups.
| Table IVDysregulated miRNAs between
VHL-associated and sporadic ccRCC samples. |
Table IV
Dysregulated miRNAs between
VHL-associated and sporadic ccRCC samples.
| miRNA ID | Fold-change
(VHL/sporadic) | Raw P-value | adj-P-value |
|---|
| Upregulated
miRNAs | | | |
| hsa-miR-489 | 2.267 | 0.0103 | ns |
| hsa-miR-204 | 2.266 | 0.0078 | ns |
| hsa-let-7f | 1.946 | 0.0004 | 0.0123 |
| hsa-miR-200b | 1.914 | 0.0012 | 0.0216 |
| hsa-let-7a | 1.821 |
5.8836×10−05 | 0.0036 |
| hsa-miR-200a | 1.767 | 0.0035 | 0.0454 |
|
hsa-miR-146b-5p | 1.684 | 0.0283 | ns |
| hsa-miR-429 | 1.611 | 0.0066 | ns |
| hsa-miR-26b | 1.579 | 0.0004 | 0.0121 |
| hsa-miR-28-5p | 1.542 | 0.0006 | 0.0121 |
| hsa-miR-122 | 1.527 | 0.0347 | ns |
| hsa-miR-20a | 1.521 | 0.0002 | 0.0092 |
| Downregulated
miRNAs | | | |
| hsa-miR-1274a | −1.580 | 0.0114 | ns |
| hsa-miR-1260 | −1.727 | 0.0027 | 0.0386 |
| hsa-miR-886-3p | −1.764 | 0.0399 | ns |
| hsa-miR-1308 | −1.812 | 0.0136 | ns |
| hsa-miR-494 | −2.882 |
6.0369×10−05 | 0.0036 |
| hsa-miR-923 | −4.149 |
2.0833×10−06 | 0.0006 |
Transcriptomic analyses identify
biological pathways involved in VHL-associated tumors
Transcriptomic analysis was performed to identify
genes exhibiting altered expression in renal VHL-associated tumors.
mRNA profiles from VHL-associated and sporadic ccRCC groups were
compared with the normal renal tissue pool. Similar to the findings
of the miRNA analysis, few differences in mRNA expression profiles
were detected between the two series of ccRCC. Notably, 3,489
(1,563 up- and 1,926 downregulated) and 3,059 (1,218 up- and 1,841
downregulated) genes were dysregulated in VHL-related tumors and
sporadic ccRCC, respectively, compared with in the normal renal
tissue pool. A total of 2,474 genes (959 up- and 1515
downregulated) were found in common between the groups,
representing 71 and 81% of each signature, respectively (Fig. 6). Dysregulated pathways were
similar to those previously described for the first set of samples
(Table III).
Discussion
The present study detected differentially expressed
mRNAs and miRNAs in VHL-associated ccRCC, in order to investigate
the molecular mechanisms underlying the pathogenesis of these
hereditary tumors. The results of the miRNA and mRNA integrative
analysis indicated that synchronous tumors occurring in the same
organ in one individual, and developing from an identical germline
background, are molecularly similar even if these renal tumors are
of independent clonal origin. These results suggested that the
final molecular evolution is not random, and confirmed the
'contingency and convergency' hypothesis previously described by
Fisher et al (10).
Therefore, it may be hypothesized that VHL-associated tumors harbor
the same expression pattern due to loss of the VHL gene;
however, the difference in global genetic background between the
two patients used in the present study allows for the distinction
between the different samples of these patients.
The present study demonstrated that miRNA and mRNA
expression levels may be used to distinguish VHL-associated renal
tumors from sporadic ccRCC. The sporadic ccRCC miRNA signature was
similar to ones previously described (14,19,20).
The identified miRNAs were also able to distinguish between
high-grade and low-grade ccRCC, as previously reported (21).
In the present study, miR-210 was markedly
overexpressed in VHL-associated and sporadic tumors. Overexpression
of miR-210 has previously been described in sporadic ccRCC and in
several hypoxic tumors (22–24);
therefore, its overexpression in VHL-associated RCC is not
surprising due to its pseudohypoxic gene signature (25). miR-210 modulates the cellular
hypoxic response via a wide range of actions. In particular, miRNA
and mRNA profiles of renal VHL-associated tumors are very similar
to hypoxic signatures previously reported (26). Another pro-oncogenic pathway was
identified through miR-155, which promotes the growth of tumors by
targeting VHL mRNA (27,28)
and the activity of hypoxia-inducible factor 1 (HIF1) during
prolonged hypoxia (29). Other
miRNAs identified in the present study have also been described in
the literature, including miR-28-5p, which promotes chromosomal
instability in ccRCC by inhibiting mitotic arrest deficient 2
translation (30), or miR-30c-3p
(previous ID: miR-30c-2*) and miR-30a-3p, which inhibit HIF2
activity in ccRCC (31).
An altered metabolic pattern has previously been
identified in ccRCC studies (32,33).
VHL, MET proto-oncogene, receptor tyrosine kinase,
folliculin, TSC complex subunit 1 (TSC1), TSC2,
fumarate hydratase and succinate dehydrogenase are known as renal
cancer-predisposing genes, which are all involved in pathways that
respond to metabolic stress or nutrient stimulation. It has
previously been suggested that RCC may be regarded as a metabolic
disease (34). It may be
interesting to perform a metabolic analysis to assess metabolic
alterations in ccRCC. It is possible that dysregulated metabolism
is fundamental for the occurrence of ccRCC and may provide the
basis for the development of novel forms of therapy. In addition,
the present study identified numerous upregulated pathways that
were associated with the immune system. ccRCC has previously been
demonstrated to be immunogenic. Notably, a number of immune cells
have been isolated from ccRCC, including natural killer cells,
cytotoxic T cells, helper T cells and dendritic cells (35–38).
When ccRCC appears as an antigen in the human body, immune activity
is induced, leading to a series of enhanced immunization
activities. Several genes implicated in these pathways were
regulated by the VHL/HIF pathway, including C-X-C motif
chemokine receptor 4 and stromal cell-derived factor-1α (39). Further studies, in order to obtain
an in-depth understanding of the mechanisms implicating the
VHL gene may be beneficial for the development of novel
treatments.
Within the genome, clustering of miRNA genes is
common, with 38% of known miRNA genes residing in clusters
(40). The present profiling data
revealed dysregulation of several miRNA clusters, notably the
δ-like non-canonical Notch ligand 1-maternally expressed 3 miRNA
cluster (on chromosome 14q32) in all ccRCC samples. Evolutionary
conservation of clustered miRNA genes suggests an important common
biological function, co-regulating identical targets or components
in the same pathway (41).
Notably, several miRNAs mapping to 14q32 have been predicted to
regulate the same target genes. Loss of expression of this miRNA
cluster or other genes in close proximity has previously been
reported in ccRCC (42), as well
as in other types of cancer (43–45).
For example, in osteosarcoma, down-regulation of 14q32 miRNAs
stabilizes c-MYC, facilitates apoptotic escape, and sustains
tumorigenesis (43). In addition,
the MYC pathway is activated in ccRCC and essential for
proliferation of ccRCC cells (46). These findings suggested that loss
of expression of miRNAs clustered at 14q32 further dysregulates the
MYC network and likely contributes to ccRCC development. The 14q32
miRNA cluster members, miR-134 and miR-494, were generally
downregulated in nearly all tumors and are described as tumor
suppressors in ccRCC cells. miR-134 downregulates cell
proliferation and epithelial-to-mesenchymal transition by targeting
KRAS proto-oncogene, GTPase (47).
A miRNA regulatory balance of oncogenic, metabolic and immune
pathways must be struck in ccRCC tumor cells to permit tumor
progression. Functional assays and global proteomic analysis are
required to better characterize these interaction networks.
In conclusion, from various clonal tumors within the
kidney of the same patient, a functional convergence on hypoxic,
immune response and metabolism pathways was evidenced, thus
contributing to the synchronous oncogenesis of these tumors.
Several miRNAs significantly differentially expressed between
VHL-related renal tumors and sporadic ccRCC were identified through
global miRNA expression profiling, thus suggesting a role for these
miRNAs in these tumors. Although further cellular and histological
studies are required to determine the precise roles played by these
miRNA-mRNA pathways in VHL-associated ccRCC, the present results
may help provide a better understanding of these hereditary
tumors.
Acknowledgments
The authors of the present study would like to thank
Dr Vladimir Lazar and Mrs Véronique Roux (Genomic Platform, Gustave
Roussy, Villejuif, France) for helping to design the micro-array
experiments. The authors are also grateful to Centre de Ressources
Biologiques from Necker and Saint Joseph Hospitals (Paris, France)
and Bicêtre Hospital (Le Kremlin-Bicêtre, France) for the frozen
samples used to perform the present study.
Funding
The present study was supported by grants from the
'Ligue Nationale contre le Cancer' (Comités du Cher et de l'Indre),
the French National Cancer Institute (INCa, PNES Kidney Cancer),
the 'Association VHL France' and by 'Taxes d'Apprentissage Gustave
Roussy' (P18_SG, P20_VPT et P24_MACH).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding authors on request.
Authors' contributions
CHG participated in the design of the study,
analyzed the data, performed the statistical analysis and drafted
the manuscript. CO and ND performed the microarray experiments. GM
and PD performed the microarray analyses. MC helped to analyze the
data. AM, SGi and SR recruited the patients, conducted follow-up
appointments and obtained their consent for the study. SF, VVe, VVa
and VM collected ccRCC tissues stored in official structures at
Bicêtre, Necker and St Joseph hospitals. SC, BG, BB, BTT and SR
helped to analyze and interpret the data, and critically revised
the manuscript. SGa conceived the study, and participated in its
design and coordination, supervised the experiments, and drafted
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the ethical
committee of Bicêtre Hospital (Le Kremlin-Bicêtre, France). All
patients provided written informed consent prior to surgery for the
use of their tumors.
Consent for publication
All patients had provided written informed consent
prior to surgery.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Richard S, Gardie B, Couvé S and Gad S:
Von Hippel-Lindau: How a rare disease illuminates cancer biology.
Semin Cancer Biol. 23:26–37. 2013. View Article : Google Scholar
|
|
2
|
Fuhrman SA, Lasky LC and Limas C:
Prognostic significance of morphologic parameters in renal cell
carcinoma. Am J Surg Pathol. 6:655–663. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sato Y, Yoshizato T, Shiraishi Y, Maekawa
S, Okuno Y, Kamura T, Shimamura T, Sato-Otsubo A, Nagae G, Suzuki
H, et al: Integrated molecular analysis of clear-cell renal cell
carcinoma. Nat Genet. 45:860–867. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu P, Liu JL, Pei SM, Wu CP, Yang K, Wang
SP and Wu S: Integrated genomic analysis identifies clinically
relevant subtypes of renal clear cell carcinoma. BMC Cancer.
18:2872018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thiesen HJ, Steinbeck F, Maruschke M,
Koczan D, Ziems B and Hakenberg OW: Stratification of clear cell
renal cell carcinoma (ccRCC) genomes by gene-directed copy number
alteration (CNA) analysis. PLoS One. 12:e01766592017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gerlinger M, Horswell S, Larkin J, Rowan
AJ, Salm MP, Varela I, Fisher R, McGranahan N, Matthews N, Santos
CR, et al: Genomic architecture and evolution of clear cell renal
cell carcinomas defined by multiregion sequencing. Nat Genet.
46:225–233. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cancer Genome Atlas and Research Network:
Comprehensive molecular characterization of clear cell renal cell
carcinoma. Nature. 499:43–49. 2013. View Article : Google Scholar
|
|
8
|
Beroukhim R, Brunet JP, Di Napoli A, Mertz
KD, Seeley A, Pires MM, Linhart D, Worrell RA, Moch H, Rubin MA, et
al: Patterns of gene expression and copy-number alterations in
von-hippel lindau disease-associated and sporadic clear cell
carcinoma of the kidney. Cancer Res. 69:4674–4681. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shuib S, Wei W, Sur H, Morris MR, McMullan
D, Rattenberry E, Meyer E, Maxwell PH, Kishida T, Yao M, et al:
Copy number profiling in von Hippel-Lindau disease renal cell
carcinoma. Genes Chromosomes Cancer. 50:479–488. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fisher R, Horswell S, Rowan A, Salm MP, de
Bruin EC, Gulati S, McGranahan N, Stares M, Gerlinger M, Varela I,
et al: Development of synchronous VHL syndrome tumors reveals
contingencies and constraints to tumor evolution. Genome Biol.
15:4332014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Iorio MV and Croce CM: Causes and
consequences of microRNA dysregulation. Cancer J. 18:215–222. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Juan D, Alexe G, Antes T, Liu H,
Madabhushi A, Delisi C, Ganesan S, Bhanot G and Liou LS:
Identification of a microRNA panel for clear-cell kidney cancer.
Urology. 75:835–841. 2010. View Article : Google Scholar
|
|
15
|
Neal CS, Michael MZ, Rawlings LH, Van der
Hoek MB and Gleadle JM: The VHL-dependent regulation of microRNAs
in renal cancer. BMC Med. 8:642010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Messai Y, Gad S, Noman MZ, Le Teuff G,
Couve S, Janji B, Kammerer SF, Rioux-Leclerc N, Hasmim M, Ferlicot
S, et al: Renal cell carcinoma programmed death-ligand 1, a new
direct target of hypoxia-inducible factor-2 alpha, is regulated by
von Hippel-Lindau gene mutation status. Eur Urol. 70:623–632. 2016.
View Article : Google Scholar
|
|
17
|
Gentleman RC, Carey VJ, Bates DM, Bolstad
B, Dettling M, Dudoit S, Ellis B, Gautier L, Ge Y, Gentry J, et al:
Bioconductor: Open software development for computational biology
and bioinformatics. Genome Biol. 5:R802004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Silva-Santos RM, Costa-Pinheiro P, Luis A,
Antunes L, Lobo F, Oliveira J, Henrique R and Jerónimo C: MicroRNA
profile: A promising ancillary tool for accurate renal cell tumour
diagnosis. Br J Cancer. 109:2646–2653. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Christinat Y and Krek W: Integrated
genomic analysis identifies subclasses and prognosis signatures of
kidney cancer. Oncotarget. 6:10521–10531. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gowrishankar B, Ibragimova I, Zhou Y,
Slifker MJ, Devarajan K, Al-Saleem T, Uzzo RG and Cairns P:
MicroRNA expression signatures of stage, grade, and progression in
clear cell RCC. Cancer Biol Ther. 15:329–341. 2014. View Article : Google Scholar :
|
|
22
|
Miko E, Czimmerer Z, Csánky E, Boros G,
Buslig J, Dezso B and Scholtz B: Differentially expressed microRNAs
in small cell lung cancer. Exp Lung Res. 35:646–664. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chan SY and Loscalzo J: MicroRNA-210: A
unique and pleiotropic hypoxamir. Cell Cycle. 9:1072–1083. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Puisségur MP, Mazure NM, Bertero T,
Pradelli L, Grosso S, Robbe-Sermesant K, Maurin T, Lebrigand K,
Cardinaud B, Hofman V, et al: miR-210 is overexpressed in late
stages of lung cancer and mediates mitochondrial alterations
associated with modulation of HIF-1 activity. Cell Death Differ.
18:465–478. 2011. View Article : Google Scholar :
|
|
25
|
Couvé S, Ladroue C, Laine E, Mahtouk K,
Guégan J, Gad S, Le Jeune H, Le Gentil M, Nuel G, Kim WY, et al:
Genetic evidence of a precisely tuned dysregulation in the hypoxia
signaling pathway during oncogenesis. Cancer Res. 74:6554–6564.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kulshreshtha R, Davuluri RV, Calin GA and
Ivan M: A microRNA component of the hypoxic response. Cell Death
Differ. 15:667–671. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen D, Cabay RJ, Jin Y, Wang A, Lu Y,
Shah-Khan M and Zhou X: MicroRNA deregulations in head and neck
squamous cell carcinomas. J Oral Maxillofac Res. 4:e22013.
View Article : Google Scholar
|
|
28
|
Kong W, He L, Richards EJ, Challa S, Xu
CX, Permuth-Wey J, Lancaster JM, Coppola D, Sellers TA, Djeu JY, et
al: Upregulation of miRNA-155 promotes tumour angiogenesis by
targeting VHL and is associated with poor prognosis and
triple-negative breast cancer. Oncogene. 33:679–689. 2014.
View Article : Google Scholar :
|
|
29
|
Bruning U, Cerone L, Neufeld Z,
Fitzpatrick SF, Cheong A, Scholz CC, Simpson DA, Leonard MO,
Tambuwala MM, Cummins EP, et al: MicroRNA-155 promotes resolution
of hypoxia-inducible factor 1alpha activity during prolonged
hypoxia. Mol Cell Biol. 31:4087–4096. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hell MP, Thoma CR, Fankhauser N,
Christinat Y, Weber TC and Krek W: miR-28-5p promotes chromosomal
instability in VHL-associated cancers by inhibiting Mad2
translation. Cancer Res. 74:2432–2443. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mathew LK, Lee SS, Skuli N, Rao S, Keith
B, Nathanson KL, Lal P and Simon MC: Restricted expression of
miR-30c-2-3p and miR-30a-3p in clear cell renal cell carcinomas
enhances HIF2α activity. Cancer Discov. 4:53–60. 2014. View Article : Google Scholar
|
|
32
|
Favier J, Brière JJ, Burnichon N, Rivière
J, Vescovo L, Benit P, Giscos-Douriez I, De Reyniès A, Bertherat J,
Badoual C, et al: The Warburg effect is genetically determined in
inherited pheochromocytomas. PLoS One. 4:e70942009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mullen AR, Wheaton WW, Jin ES, Chen PH,
Sullivan LB, Cheng T, Yang Y, Linehan WM, Chandel NS and
DeBerardinis RJ: Reductive carboxylation supports growth in tumour
cells with defective mitochondria. Nature. 481:385–388. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Schmidt LS and Linehan WM: Genetic
predisposition to kidney cancer. Semin Oncol. 43:566–574. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Elsässer-Beile U, Grussenmeyer T,
Gierschner D, Schmoll B, Schultze-Seemann W, Wetterauer U and
Schulte Mönting J: Semiquantitative analysis of Th1 and Th2
cytokine expression in CD3+, CD4+, and
CD8+ renal-cell-carcinoma-infiltrating lymphocytes.
Cancer Immunol Immunother. 48:204–208. 1999. View Article : Google Scholar
|
|
36
|
Finke JH, Rayman P, Edinger M, Tubbs RR,
Stanley J, Klein E and Bukowski R: Characterization of a human
renal cell carcinoma specific cytotoxic CD8+ T cell
line. J Immunother. 1991(11): 1–11. 1992. View Article : Google Scholar
|
|
37
|
Gaudin C, Dietrich PY, Robache S, Guillard
M, Escudier B, Lacombe MJ, Kumar A, Triebel F and Caignard A: In
vivo local expansion of clonal T cell subpopulations in renal cell
carcinoma. Cancer Res. 55:685–690. 1995.PubMed/NCBI
|
|
38
|
Schwaab T, Schned AR, Heaney JA, Cole BF,
Atzpodien J, Wittke F and Ernstoff MS: In vivo description of
dendritic cells in human renal cell carcinoma. J Urol. 162:567–573.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zagzag D, Krishnamachary B, Yee H, Okuyama
H, Chiriboga L, Ali MA, Melamed J and Semenza GL: Stromal
cell-derived factor-1alpha and CXCR4 expression in hemangioblastoma
and clear cell-renal cell carcinoma: Von Hippel-Lindau
loss-of-function induces expression of a ligand and its receptor.
Cancer Res. 65:6178–6188. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Altuvia Y, Landgraf P, Lithwick G, Elefant
N, Pfeffer S, Aravin A, Brownstein MJ, Tuschl T and Margalit H:
Clustering and conservation patterns of human microRNAs. Nucleic
Acids Res. 33:2697–2706. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yuan X, Liu C, Yang P, He S, Liao Q, Kang
S and Zhao Y: Clustered microRNAs' coordination in regulating
protein-protein interaction network. BMC Syst Biol. 3:652009.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kawakami T, Chano T, Minami K, Okabe H,
Okada Y and Okamoto K: Imprinted DLK1 is a putative tumor
suppressor gene and inactivated by epimutation at the region
upstream of GTL2 in human renal cell carcinoma. Hum Mol Genet.
15:821–830. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Thayanithy V, Sarver AL, Kartha RV, Li L,
Angstadt AY, Breen M, Steer CJ, Modiano JF and Subramanian S:
Perturbation of 14q32 miRNAs-cMYC gene network in osteosarcoma.
Bone. 50:171–181. 2012. View Article : Google Scholar
|
|
44
|
Gattolliat CH, Thomas L, Ciafrè SA,
Meurice G, Le Teuff G, Job B, Richon C, Combaret V, Dessen P,
Valteau-Couanet D, et al: Expression of miR-487b and miR-410
encoded by 14q32.31 locus is a prognostic marker in neuroblastoma.
Br J Cancer. 105:1352–1361. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gattolliat CH, Le Teuff G, Combaret V,
Mussard E, Valteau-Couanet D, Busson P, Bénard J and Douc-Rasy S:
Expression of two parental imprinted miRNAs improves the risk
stratification of neuroblastoma patients. Cancer Med. 3:998–1009.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tang SW, Chang WH, Su YC, Chen YC, Lai YH,
Wu PT, Hsu CI, Lin WC, Lai MK and Lin JY: MYC pathway is activated
in clear cell renal cell carcinoma and essential for proliferation
of clear cell renal cell carcinoma cells. Cancer Lett. 273:35–43.
2009. View Article : Google Scholar
|
|
47
|
Liu Y, Zhang M, Qian J, Bao M, Meng X,
Zhang S, Zhang L, Zhao R, Li S, Cao Q, et al: miR-134 functions as
a tumor suppressor in cell proliferation and
epithelial-to-mesenchymal Transition by targeting KRAS in renal
cell carcinoma cells. DNA Cell Biol. 34:429–436. 2015. View Article : Google Scholar : PubMed/NCBI
|