Introduction
Colorectal cancer (CRC) is one of the most frequent
malignant diseases in the world. The metastatic spread of CRC is a
crucial factor for the progression and therapeutic management of
the disease. Due to the lack of suitable biomarkers for early
diagnosis and specific targets for precise treatment, the general
prognosis of late-stage CRC remains poor. Currently, the 5-year
relative survival rate for metastatic CRC is only ~10% (1). Therefore, the pursuit of early
diagnosis markers and novel therapeutic targets is imperative for
improving the outcome of these patients.
A substantial body of evidence demonstrates that
translationally controlled tumor protein (TCTP), a highly conserved
and multifunctional protein, is implicated in the tumorigenesis of
various malignances (2–4). The expression levels of TCTP are
frequently elevated in human cancer, and a high TCTP status has
been associated with a poor outcome in mammary (5), hepatocellular (6) and colorectal cancer (7).
In our previous studies, it was demonstrated that
TCTP is involved in CRC progression and metastasis (7,8), and
a possible mechanism was suggested, in which TCTP may induce
activation of the cell division cycle 42 (Cdc42)/c-Jun N-terminal
kinase/matrix metalloproteinase (MMP)9 signaling pathway (7), which is closely associated with the
invasion and metastatic potential of tumors. However, tumor
metastasis is a complex system in which several factors are
involved and interconnected to form a large and complex network.
Our previous findings may explain certain aspects of the role of
TCTP in tumor metastasis, however, whether there is any relevant
influential factor that it is important in this metastatic process
remains to be elucidated and requires further investigation.
In our previous proteome experiment, it was
determined that several important proteins were markedly altered
following the knockdown of TCTP (8). Among them, high mobility group box 1
(HMGB1) was of particular interest, as accumulating evidence
suggests that HMGB1 is overexpressed in CRC tissue and also
contributes to tumor growth and metastasis (9–11).
Therefore, it was hypothesized that there may be an important
interaction between TCTP and HMGB1 in the tumor metastasis
process.
As is known at present, HMGB1 is one of the
non-histone chromosomal proteins and also an extracellular
damage-associated molecular pattern molecule. It locates in the
nucleus where it binds to minor groove DNA, and functions as a
regulator of gene transcription. During states of cellular stress
or damage, HMGB1 can be released from cells and become an
extracellular cytokine, being involved in inflammation,
angiogenesis, cell proliferation and migration (12–14).
Toll-like receptors (TLRs) and receptor for advanced glycation end
products (RAGE) are the main receptors of HMGB1 (15,16).
Although signaling pathways for the two receptor groups are
different, ultimately, they both activate nuclear factor-κB (NF-κB)
(12,17-19).
NF-κB is an inducible dimeric transcription factor, and its
prototype in the majority of cell types is the p65/p50 heterodimer.
It is usually retained in an inactive state in the cytoplasm by its
association with inhibitor of NF-κB (IκB). Upon stimulation, IκBα
is rapidly phosphorylated and subsequently degraded by proteasomes,
permitting translocation of the p65/p60 heterodimer into the
nucleus. This results in the activation of target genes associated
with cell proliferation, inflammation, and tumor development and
metastasis (20,21).
On the basis of the above knowledge, it was
hypothesized that TCTP may promote CRC metastasis through
regulating the expression and secretion of HMGB1 and consequent
activation of the NF-κB signaling pathway. The present study aimed
to provide experimental evidence to confirm this hypothesis.
Materials and methods
Materials
Primary antibodies specific for TCTP (sc-133131),
TLR4 (sc-293072), RAGE (sc-365154), NF-κB p65 (sc-8008), IκBα
(sc-1643), GAPDH (sc-32233) and Lamin B (sc-374015), secondary
anti-mouse (sc-2005) and anti-rabbit (sc-2030) IgG-conjugated
horseradish peroxidase, and the specific NF-κB inhibitor Bay117082
were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA). Antibody specific for HMGB1 (ab18256) was obtained from Abcam
(Cambridge, MA, USA). The recombinant human HMGB1 (rhHMGB1) and all
other chemicals were supplied by Sigma-Aldrich; EMD Millipore
(Billerica, MA, USA).
Clinical samples
The present study recruited 30 patients who were
endoscopically diagnosed and histologically confirmed with CRC and
then admitted to Nanfang Hospital, Southern Medical University
(Guangzhou, China) between September, 2016 and August, 2017. Among
these patients, 21 were men and nine were women, with a mean age of
58.6 years, and 12 patients (40%) were confirmed to have distant
metastases by histopathological examination of surgical specimens
or by imageological detection. Additionally, 10 individuals with
benign colorectal polyps were enrolled as controls (six men and
four women, mean age 52.5 years). Peripheral blood samples were
obtained from all participants prior to any treatment and stored at
−80°C until analysis. The colorectal specimens were obtained
following tumor excision or endoscopic polypectomy. The protocols
for the collection of human tissues were approved by the Ethics
Committee of Nanfang Hospital, Southern Medical University, and
written informed consent was signed by each participant prior to
their inclusion in the study.
Immunohistochemical examination
Tissue sections (3 μm thickness) were
deparaffinized in histolene and hydrated in graded ethanol.
Endogenous peroxidase activity was quenched by treating the
sections with 3% hydrogen peroxide in phosphate-buffered saline
(PBS) for 10 min. The primary antibody against TCTP or HMGB1 (both
at a dilution of 1:1,000) was applied overnight at 4°C. The
sections were then incubated for 30 min with the secondary antibody
(1:1,000) at room temperature. Finally, the sections were developed
by applying diaminobenzidine as a chromogen, followed by
counterstaining with hematoxylin. Sections incubated without
primary antibody served as negative controls. The slides were
semi-quantitatively scored using a Nikon Eclipse E100 microscope
(Nikon Instruments, Tokyo, Japan), according to the mean density
(the ratio of the integral optical density to the total area), as
previously described (7). The mean
percentage of positive cells was classified into four categories: 0
for negative, 1 (weak) for <10%, 2 (moderate) for 10–50%, and 3
(strong) for >50%.
Enzyme-linked immunosorbent assay
(ELISA)
The HMGB1 concentrations in serum samples were
quantified by means of an HMGB1 ELISA Kit II (Shino-Test, Tokyo,
Japan) according to the manufacturer's protocol. Briefly, the
standard, sample or control was added to 96-well microtiter plates
and incubated for 24 h at 37°C. Following washing, anti-HMGB1
peroxidase-conjugated monoclonal antibody was added and incubated
at room temperature for 2 h. The plates were washed again and a
substrate solution was added. The enzyme colorimetric reaction was
allowed to proceed for 30 min at room temperature and terminated by
the addition of stop solution. The absorbance was read at 450 nm,
and results were calculated using a calibration curve prepared from
the standards.
Cell culture
Human colon adenocarcinoma LoVo cells, obtained from
American Type Culture Collection (Rockville, MD, USA), were plated
on culture flasks and cultured in Dulbecco's modified Eagle's
medium (DMEM; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% inactivated fetal bovine serum (FBS), 2
mM L-glutamine (both from Invitrogen; Thermo Fisher Scientific,
Inc.), and 1% penicillin/streptomycin (Gibco; Thermo Fisher
Scientific, Inc.) in a humidified incubator at 37°C and 5%
CO2.
Protein preparation
Total proteins were extracted from the tissues with
RIPA lysis buffer (Beyotime Institute of Biotechnology, Jiangsu,
China) containing protease inhibitors, whereas nuclear and
cytoplasmic extracts from LoVo cells were prepared using a Nuclear
and Cytoplasmic Protein Extraction kit (Beyotime Institute of
Biotechnology). For preparation of the protein in the cell culture
medium, an ultrafiltration method was used, as described previously
(22). Briefly, the cells in a
100-mm plate were washed and incubated in serum-free medium for 24
h. Following the elimination of cellular debris by 10 min of
centrifugation at 3,000 × g at 4°C, the culture medium was
concentrated to 100 μl with an Amicon Ultra-15 centrifugal
filter (EMD Millipore, Burlington, MA, USA).
Immunoblotting
The protein concentration of samples was detected by
Pierce BCA Protein Assay Kit (Thermo Fisher Scientific, Inc.),
according to the manufacturer's instructions. Samples containing
the same quantity of protein (20 μg) were separated by
sodium dodecyl sulfate-polyacrylamide gel electrophoresis with 12%
gels and were electroblotted onto polyvinylidene difluoride
membranes. Following transfer, the membranes were blocked with 5%
non-fat milk in Tris-buffered saline-Tween and then incubated
overnight at 4°C with the different primary antibodies (specific
for TCTP, IκBα, NF-κB p65, GAPDH and Lamin B, respectively) at a
dilution of 1:200, with the exception of anti-HMGB1 used at a
dilution of 1:1,000, followed by incubation with the horseradish
peroxidase-conjugated secondary antibodies (1:1,000) at room
temperature for 1 h. The immunoreactive bands were detected with an
enhanced chemiluminescence system (Pierce; Thermo Fisher
Scientific, Inc.). The density of individual bands was quantified
by densitometric scanning of the blots using ImageJ software
version 1.45 (NIH, Bethesda, MD, USA).
Immunofluorescence assay
The cells (5×106/ml) were placed on
Matrigel-coated cover slides. After 24 h, cells on the cover slides
were fixed with paraformaldehyde and permeabilized with 0.2% Triton
X-100. The slides were blocked with 10% bovine serum albumin
(Sigma-Aldrich, St. Louis, MO, USA) in PBS for 1 h and then
incubated with rabbit anti-HMGB1 antibody (1:200) at 4°C for 24 h.
Following washing with PBS, the cells were incubated with
fluorescence-conjugated goat anti-rabbit IgG (A11008; Invitrogen;
Thermo Fisher Scientific, Inc.) at room temperature for 30 min and
then incubated with Hoechst 33258 (Beyotime Institute of
Biotechnology) for 10 min. The cells were then rinsed in PBS, and
viewed under a fluorescent microscope.
Construction of short hairpin (sh)RNA
plasmid targeting TCTP
The RNA interference experiment protocol was
performed as previously described (8). Nucleotides 228-246
(CTCGCTCATTGGTGGAAAT) of the TCTP (GenBank accession no. NM003295)
coding sequence was selected as the target for shRNA. The
shRNA-TCTP encoding sequence was created by using two complementary
oligonucleotides, each containing the 19-nucleotide target sequence
of TCTP (228-246), followed by a short spacer (TTCAAGAGA). The
shRNA TCTP-encoding sequence was cloned into the BamHI and
BbsI sites of the pGPU6/GFP/Neo siRNA expression vector
(GenePharma, Shanghai, China).
Construction of plasmid overexpressing
TCTP
Primers were designed to amplify the coding region
of TCTP as follows: Forward, GAAGATCTATGATTATCTACCGGGACCTCAT
(BglII) and reverse, GGAATTCTTAACATTTTTCCATTTCTAAACCA
(EcoRI). The pEGFP-C1-TCTP was constructed and verified by
sequencing the inserted gene. The plasmid was transfected into
cells with Lipofectamine 2000™ (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. Stable
transfectants were selected using G418 (Gibco; Thermo Fisher
Scientific, Inc.), and the GFP signal was visualized under a
laser-scanning confocal microscope.
Lactate dehydrogenase (LDH) assay
The levels of LDH released into the culture medium
were determined using an LDH assay kit (Sigma-Aldrich; EMD
Millipore) in accordance with the manufacturer's protocol. The cell
culture supernatants were collected by centrifugation at 300 × g
for 5 min at room temperature, followed by transfer onto 96-well
plates. The plates were treated with reaction solution and placed
in the dark for 30 min at room temperature. The absorbance was
measured at 490 nm in an automatic microplate reader, subtracting
the corresponding background values from all samples.
Cell invasion assay
Cell invasion assays were performed using
Costar® Transwell™ Permeable Supports (Corning
Incorporated, Corning, NY, USA). The cells (2×105) were
suspended in DMEM supplemented with 0.1% FBS and added to the upper
chamber. The lower chamber contained 500 μl of DMEM with 10%
FBS, and the polycarbonate filter was coated with 40 μl of
Matrigel (R&D Systems, Inc., Wiesbaden, Germany). Following 24
h of incubation at 37°C, the cells on the upper surface of the
filter were removed by wiping with a cotton swab. The filters were
fixed in 4% paraformaldehyde and were stained with crystal violet.
Those cells that migrated to the lower surface of the filter were
counted based on six fields randomly under an EVOS FL Auto
fluorescence microscope (Invitrogen; Thermo Fisher Scientific,
Inc.) at ×400 magnification.
Luciferase assay
The LoVo cells were seeded in 24-well plates at
1×105 cells per well. The cells were transiently
co-transfected with 400 ng of pNF-κB-Luc (Clontech Laboratories,
Inc., Mountain View, CA, USA) and 4 ng of pRL-SV40 (Promega,
Madison, WI, USA) using Lipofectamine 2000™ (Invitrogen). The
pRL-SV40 plasmid with a cDNA encoding Renilla luciferase was
used as an internal control. The cell extracts were prepared in
luciferase cell culture lysis buffer (Promega). The activities of
firefly and Renilla luciferases were measured sequentially
from a single sample with the Dual Luciferase Reporter Assay system
(Promega) using a Lumat LB 9507 luminometer (Bethold Technologies,
Bad Wildbad, Germany).
Tumor xenografts
Male BALB/c nude mice (n=6 each group), aged from 6
to 8 weeks and weighing approximately 18–22 g, were supplied by the
Laboratory Animal Center of Southern Medical University (Guangzhou,
China). All mice were bred under specific pathogen-free conditions,
at light periods of 12 h each day, and were fed water and mouse
chow ad libitum. The experiment was approved by the
Association for the Accreditation and Assessment of Laboratory
Animal Care (Guangzhou, China). To produce the experimental liver
metastasis model, LoVo cells (5×106) were injected into
the spleen of nude mice, as described previously (7,8).
Following 5 weeks of inoculation, the mice (weighing from 28 to 39
g) were sacrificed and the tumors were collected. The number of
macroscopically visible metastatic nodules on the surface of liver
was counted. The tumor specimens from primary or metastatic sites
were used for histological examinations and immunohistochemical
assays.
Statistical analysis
All data are expressed as the mean ± standard
deviation. Multiple comparisons were performed using one-way
analysis of variance (ANOVA) or nested ANOVA with the Bonferroni
test or Dunnett's T3 test, depending on whether equal variances
could be assumed. The non-parametric Kruskal-Wallis test was used
for the comparison of the immunohistochemical staining scores.
Pearson's correlation coefficient was used to measure the
association between detected values of TCTP and HMGB1. P<0.05
was considered to indicate a statistically significant difference.
All statistical analyses were performed using SPSS 23.0 for Windows
(IBM SPSS, Armonk, NY, USA).
Results
Expression of TCTP in CRC tissue is
closely correlated with HMGB1 serum concentration in patients with
CRC
To investigate whether TCTP and HMGB1 have
correlativity in the tumor progression and metastasis of CRC, the
present study measured the protein level of TCTP in CRC tissue and
the serum concentration of HMGB1 in patients with CRC, and assessed
the correlation between them. The representative result of
immunohistochemical staining of TCTP is shown in Fig. 1A. In contrast to the immunoreaction
of TCTP in adjacent normal colorectal mucosa or lymph nodes, where
it was either absent or barely detectable, the expression of TCTP
was markedly increased in carcinoma and metastatic foci in lymph
nodes. Significant differences were also found in the comparison of
TCTP-staining scores between samples from patients with CRC and
controls (P<0.001). As shown in Fig. 1B, the protein levels of TCTP were
detected in CRC samples from patients with CRC (with or without
metastasis) and from control subjects with benign polyps. As shown
by the immunoblotting analysis, the expression of TCTP was markedly
higher in the CRC samples than in the non-tumor samples, and was
further elevated in tumors that had undergone metastasis. The
average serum levels of HMGB1 detected by ELISA examination in
patients with CRC and control subjects are shown in Fig. 1C. The serum concentrations of HMGB1
were significantly higher in patients with non-metastatic CRC than
in the control group (35.443±3.873 vs. 14.007±3.348 ng/ml), and
this difference was more evident when the patients developed
distant metastasis (mean HMGB1 serum level, 41.682±4.689 ng/ml). As
shown in Fig. 1D, there was a
positive correlation between serum HMGB1 value and tumor tissue
TCTP expression (Pearson's correlation coefficient, 0.935;
P<0.001). All these data suggest there may be a potential
interaction between the two important tumor-promoting proteins.
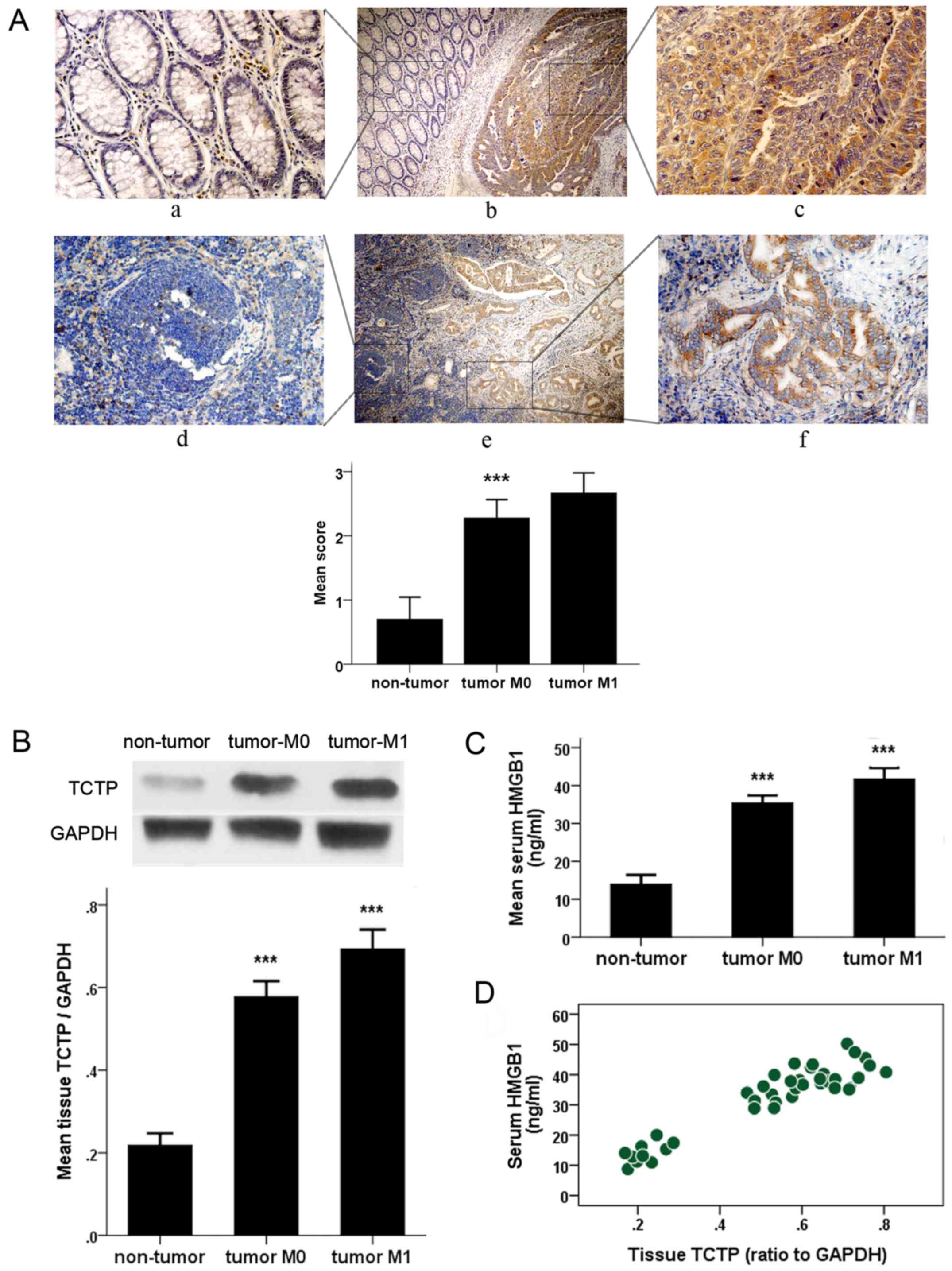 | Figure 1Detection of the histological
expression of TCTP and serum value of HMGB1 in patients with CRC.
(A) Immunohistochemical detection of expression of TCTP in CRC
tissues (upper panel) and metastatic lymph nodes (lower panel) and
the adjacent normal tissues. Specific TCTP staining is shown in
brown. Panels a, c, d and f, show a magnification of ×400; panels b
and e show a magnification of ×100. Panels a, c, d and f are
amplifications of the marked (with squares) areas shown in panels b
and e, respectively (***P<0.001 between CRC and
non-tumor groups). (B) Immunoblot analysis of expression of TCTP in
colorectal samples from patients with CRC or from control subjects
with benign polyps (non-tumor). Values were normalized to the
internal control GAPDH (***P<0.001 between each two
groups). (C) Enzyme-linked immunoassay examination of serum levels
of HMGB1 in patients with CRC and control subjects (non-tumor)
(***P<0.001 between each two groups). (D) Correlation
plot generated with serum HMGB1 values and tumor tissue TCTP
expression in 30 patients with CRC and 10 non-cancerous controls
(Pearson's correlation coefficient, 0.935; P<0.001). CRC,
colorectal cancer; TCTP, translationally controlled tumor protein;
HMGB1, high mobility group box 1; M0, no metastasis; M1,
metastasis. |
TCTP induces HMGB1 cytoplasmic
translocation in LoVo cells and extracellular release from
cells
To better understand the functions of TCTP in
carcinoma cells, vectors for overexpression or knockdown of the
TCTP gene were constructed and stably transfected into LoVo cells.
This was followed by investigation of whether alteration of the
expression of TCTP elicits variation in the expression and location
of HMGB1. The results of the immunofluorescence assay revealed the
transportation of HMGB1, as shown in Fig. 2A. The fluorescent staining of HMGB1
was largely confined to the nuclei in parental LoVo cells, and was
weak when TCTP was knocked down. By contrast, the signal was
redistributed to the cytoplasm following upregulation of the
expression of TCTP. The immunoblotting analysis (Fig. 2B) further revealed that the
cytoplasmic expression of HMGB1 was significantly increased with
the increased expression of TCTP, with a correlation rate of
0.984.
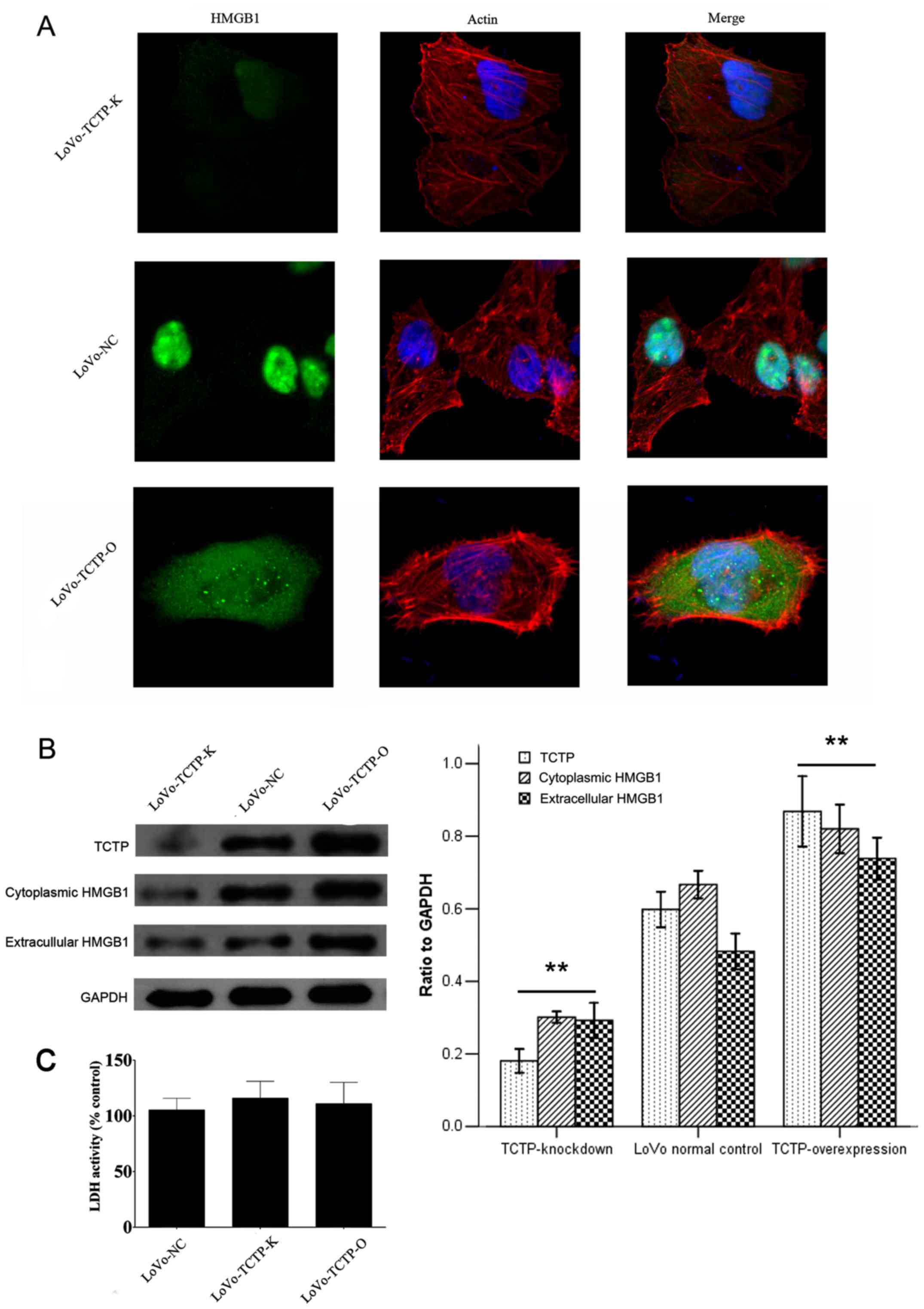 | Figure 2Effect of TCTP on the expression and
secretion of HMGB1 in LoVo cells. (A) Immunofluorescence analysis
of the expression of HMGB1 following modulation of the expression
of TCTP in LoVo cells (magnification, ×400). Green, HMGB1; blue,
nuclei; red, F-actin. (B) Immunoblotting analysis of the expression
of HMGB1 in the cytoplasm and the culture medium of LoVo cells
following modulation of the expression of TCTP. Values were
normalized to the internal control GAPDH. **P<0.01,
compared with LoVo-NC cells. Similar results were obtained in four
independent experiments. (C) Measurement of extracellular LDH
release from control LoVo cells or cells transfected with vectors.
The results are expressed as a percentage relative to the control
group. No significant difference was observed in LDH activity among
the three cell lines. LoVo-NC, normal control LoVo cells; TCTP-K,
LoVo cells transfected with TCTP-knockdown vector; TCTP-O, LoVo
cells transfected with TCTP-overexpression vector; TCTP,
translationally controlled tumor protein; HMGB1, high mobility
group box 1; LDH, lactate dehydrogenase. |
High cytoplasmic levels of HMGB1 usually occur in
the context of active HMGB1 release. To confirm this, the present
study measured the level of HMGB1 in the culture medium of LoVo
cells transfected with the vectors described above. As shown in the
immunoblotting (Fig. 2B), the
alteration in the protein level of HMGB1 in the medium was
approximately parallel with the change in the expression of TCTP in
LoVo cells, which followed a similar pattern to that observed for
the expression of HMGB1 in the cytoplasm. Taken together, these
results demonstrated that TCTP induced the release of HMGB1 from
the nucleus to the cytoplasm and into the extracellular space.
It is reported that LDH can leak into extracellular
fluids only when damage to the plasma membrane occurs (23). Therefore, the LDH assay was applied
to determine whether TCTP can destroy the integrity of the cellular
membrane, thus resulting in the release of HMGB1. As shown in
Fig. 2C, transfection with either
the TCTP-overexpression or TCTP-knockdown plasmids caused minimal
LDH leakage, which showed no significant differences when compared
with the control LoVo cells. This result indicated that the
TCTP-induced release of HMGB1 was the consequence of cell secretion
rather than cell membrane damage.
TCTP triggers NF-κB activation in LoVo
cells, which is mediated by HMGB1 and its receptors TLR4 and
RAGE
As NF-κB has been revealed as a downstream signaling
component of several receptors of HMGB1 (12,17–19),
and the behavior of HMGB1 in cancer cells appeared to be influenced
by TCTP according to the above experiments, the present study
examined whether NF-κB is also involved in the TCTP-induced
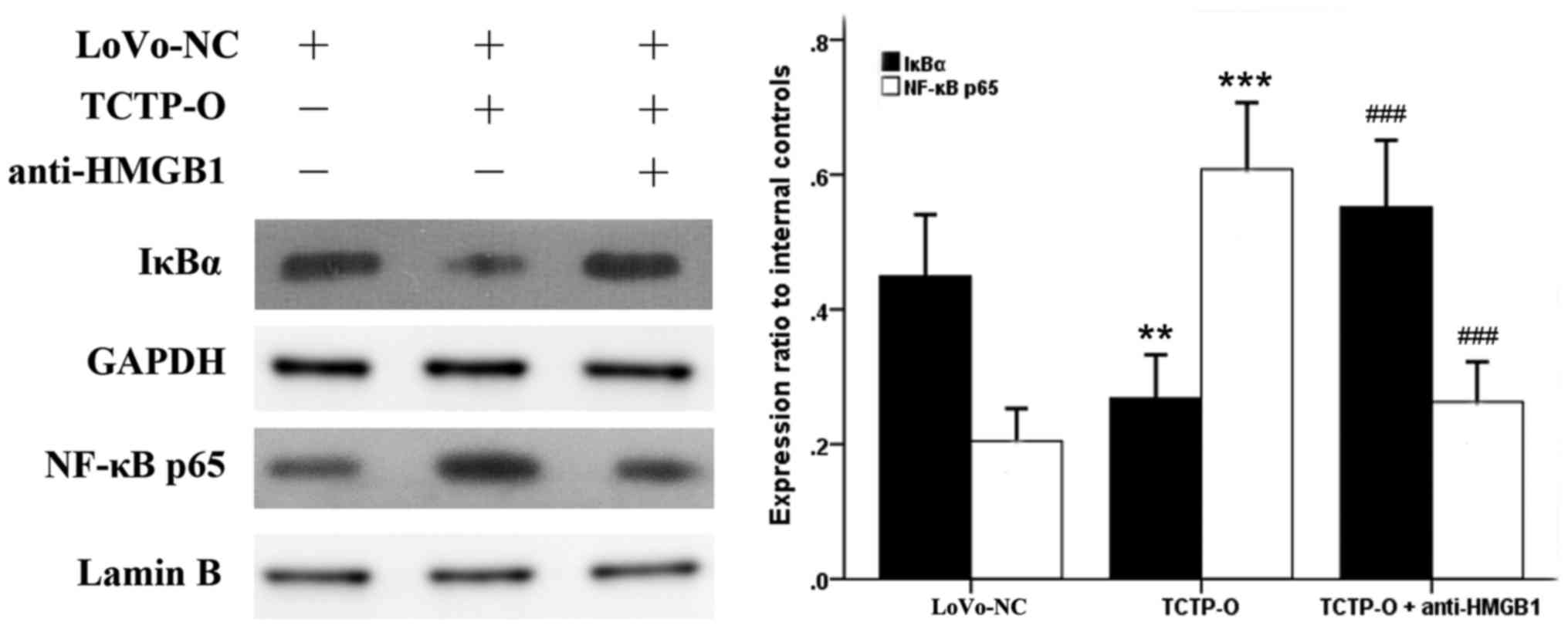
signaling pathway. The expression levels and the activity of NF-κB
were evaluated in LoVo cells following altering of the gene
expression of TCTP. As revealed by the immunoblotting analysis
(Fig. 3), compared with the
untreated normal control LoVo cells, the overexpression of TCTP
reduced the protein level of IκBα in the cytoplasm, and
simultaneously increased the expression of NF-κB p65 subunit in the
nucleus (P=0.003 and P<0.001, respectively), indicating a
process of proteolytic degradation of IκB and nuclear translocation
of NF-κB. By contrast, when the TCTP-overexpressing cells were
treated with antibody against HMGB1 (5 μg/ml for 24 h), the
responses of IκBα and NF-κB p65 elicited by TCTP were significantly
reversed (P<0.001). Further experiments revealed (Fig. 4A) that the inhibition of HMGB1 by
applying antibodies to its receptors TLR4 or RAGE (5 μg/ml
for 24 h) in these cells also led to a reduction of TCTP-induced
NF-κB activation. These findings were consistent with the results
of the luciferase assay (Fig. 4B),
which showed that the activity of NF-κB was enhanced by the
overexpression of TCTP or administration of rhHMGB1 (10
μg/ml for 24 h), but decreased by the antagonists of HMGB1.
Knockdown of the TCTP gene in LoVo cells led to a similar effect to
these antagonisms of HMGB1 on the activation of NF-κB. Taken
together, these results indicated that TCTP activated NF-κB in LoVo
cells through the mediation of HMGB1 and its receptors TLR4 and
RAGE.
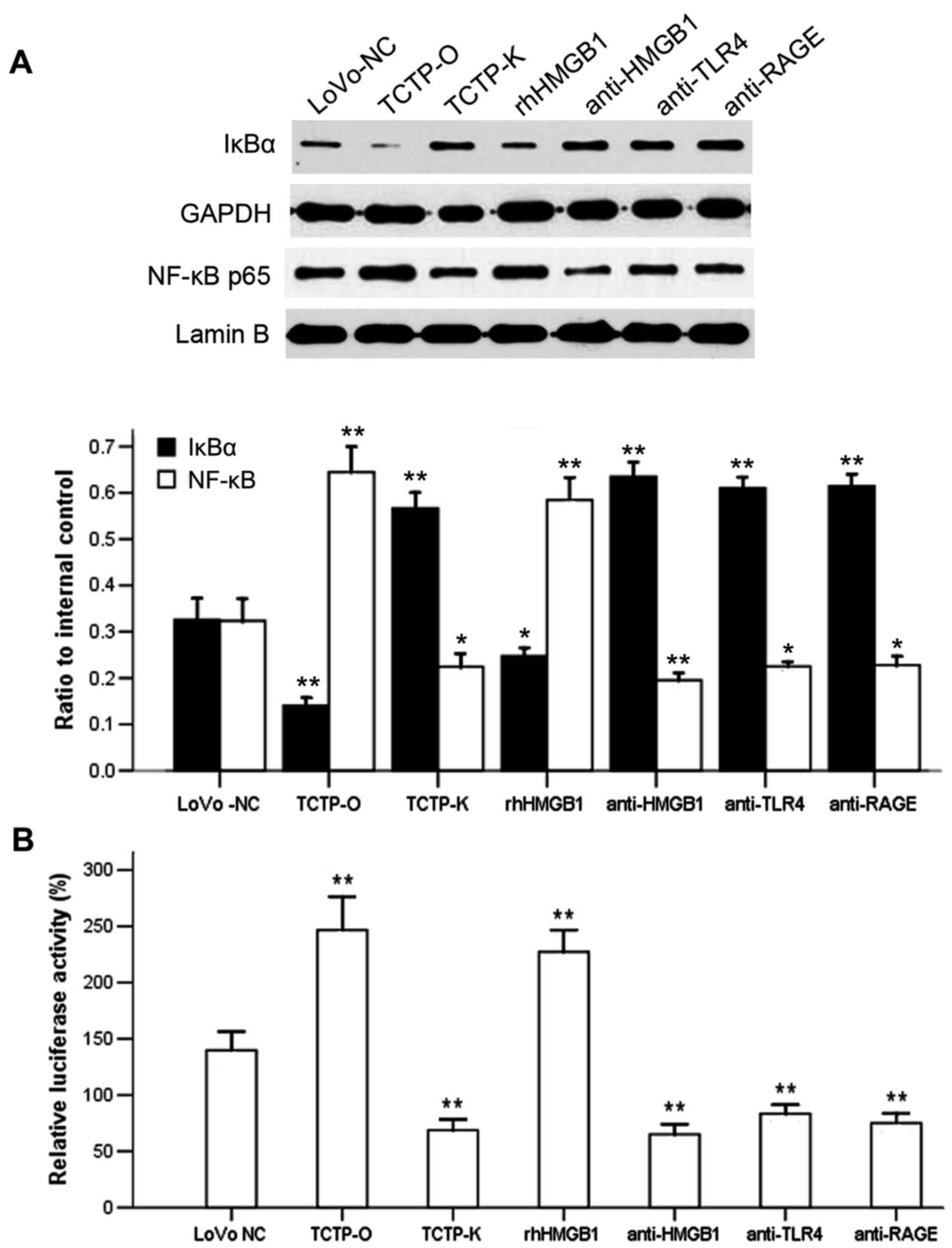 | Figure 4Effect of antibodies against HMGB1 or
its receptors TLR4 and RAGE on the TCTP-induced activation of NF-κB
in LoVo cells. (A) Immunoblotting analysis of the expression of
cytoplasmic IκBα and nuclear NF-κB p65. Lanes: 1, LoVo-NC; 2,
TCTP-O; 3, TCTP-K; 4, TCTP-O +10 μg/ml rhHMGB1; 5, TCTP-O +
5 μg/ml anti-HMGB1; 6, TCTP-O + 5 μg/ml anti-TLR4; 7,
TCTP-O + 5 μg/ml anti-RAGE. GAPDH and Lamin B were used as
internal controls of cytoplasmic and nuclear protein, respectively.
*P<0.05 and **P<0.01, compared with
LoVo-NC cells. Data are the representative of four experiments with
similar results. (B) Activity of NF-κB was determined by a
Luciferase reporter assay. **P<0.01, compared with
LoVo-NC cells. Each assay was performed in triplicate. LoVo-NC,
normal control LoVo cells; TCTP-K, LoVo cells transfected with
TCTP-knockdown vector; TCTP-O, LoVo cells transfected with
TCTP-overexpression vector; TCTP, translationally controlled tumor
protein; HMGB1, high mobility group box 1; rhHMGB1, recombinant
human HMGB1; NF-κB, nuclear factor-κB; TLR4, Toll-like receptor 4;
RAGE, receptor for advanced glycation end products. |
TCTP increases the invasion potential of
LoVo cells through activation of the HMGB1-TLR4/RAGE-NF-κB
pathway
The present study further investigated the invasive
capacity of LoVo cells following the treatments described above. As
shown in Fig. 5, compared with the
untreated parental LoVo cells, the downregulation of TCTP
significantly inhibited the invasion ability of the tumor cells,
whereas the upregulation of TCTP led to the opposite result (both
P<0.001). These three groups of cells were then treated with
rhHMGB1 or relevant neutralizing antibodies. The results
demonstrated that the increased cell invasiveness induced by TCTP
was further promoted by rhHMGB1, but was substantially attenuated
by administration of antibodies against HMGB1, TLR4 or RAGE. In
addition, treatment with the specific NF-κB inhibitor Bay117082 (5
μmol/l) significantly suppressed tumor cell invasion. These
results indicated that the effect of TCTP on the invasion of LoVo
cells was mediated by the HMGB1-TLR4/RAGE-NF-κB pathway.
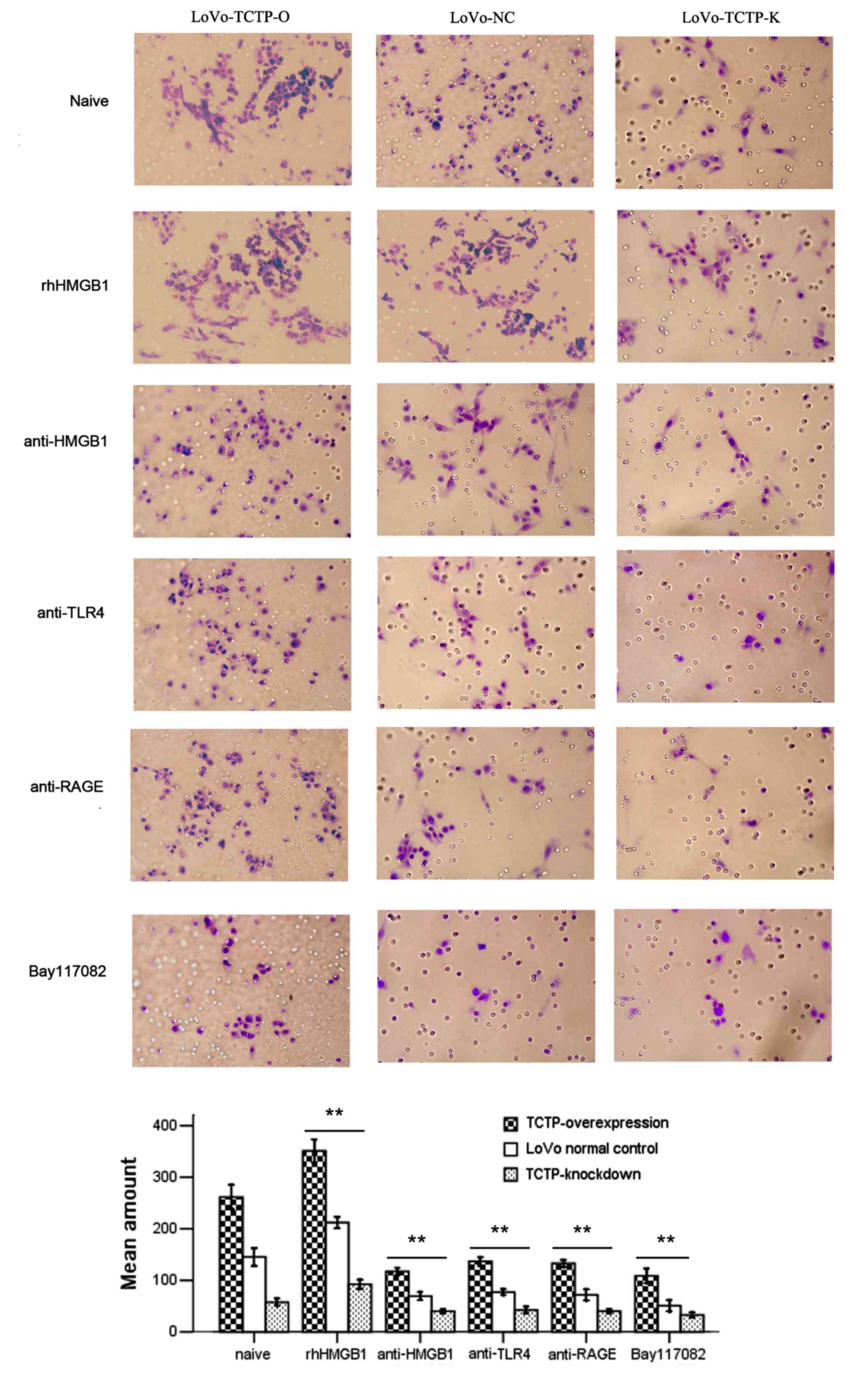 | Figure 5Effect of TCTP and the
HMGB1-TLR4/RAGE-NF-κB pathway on the invasion potential of LoVo
cells (magnification, ×400). The invasive capacity of LoVo NC cells
(middle), LoVo TCTP-O cells (left) or LoVo-TCTP-K (right) were
assessed by a cell invasion assay. The number of invasive cells was
quantified by visual counting of six randomly selected fields. The
groups comprised cells left untreated as controls, cells treated
with rhHMGB1 (10 μg/ml) or anti-HMGB1, anti-TLR4, anti-RAGE
(all 5 μg/ml) or the specific NF-κB inhibitor Bay117082 (5
μmol/l), respectively. **P<0.01, compared with
untreated cells. Similar results were obtained in four independent
experiments. LoVo-NC, normal control LoVo cells; TCTP-K, LoVo cells
transfected with TCTP-knockdown vector; TCTP-O, LoVo cells
transfected with TCTP-overexpression vector; TCTP, translationally
controlled tumor protein; HMGB1, high mobility group box 1;
rhHMGB1, recombinant human HMGB1; NF-κB, nuclear factor-κB; TLR4,
Toll-like receptor 4; RAGE, receptor for advanced glycation end
products. |
TCTP regulates the expression of HMGB1 in
xenograft tumors in nude mice
As it was found that the expression of HMGB1 in LoVo
cells was regulated by TCTP, a corresponding change in the
expression of HMGB1 was anticipated in xenograft tumors in nude
mice following modulation of TCTP gene expression. The visualized
observations were similar to our previous studies (7,8),
which showed that the weight of xenograft tumors and the number of
hepatic metastases were synchronously altered with the expression
level of TCTP in LoVo cells (data not shown). In addition, the
immunostaining density of HMGB1 was positively linked with that of
TCTP in primary tumor tissues (Fig.
6A) and in metastatic foci (Fig.
6B), both of which varied in accordance with the gene
expression levels of TCTP in the LoVo cells. Further microscopic
examinations revealed that the metastatic nodules in mouse livers
were significantly augmented with the increased expression of TCTP
(Fig. 6B, 'H&E’
panel). There were significant differences in the
immunohistochemical staining scores of TCTP and HMGB1 between
either the TCTP-overexpression or TCTP-knockdown groups and the
control (all P<0.05). These findings suggested that TCTP
promoted the progression of CRC and liver metastasis; it also
regulated the expression of HMGB1, which possibly produced
synergistic effects on the metastatic process of CRC.
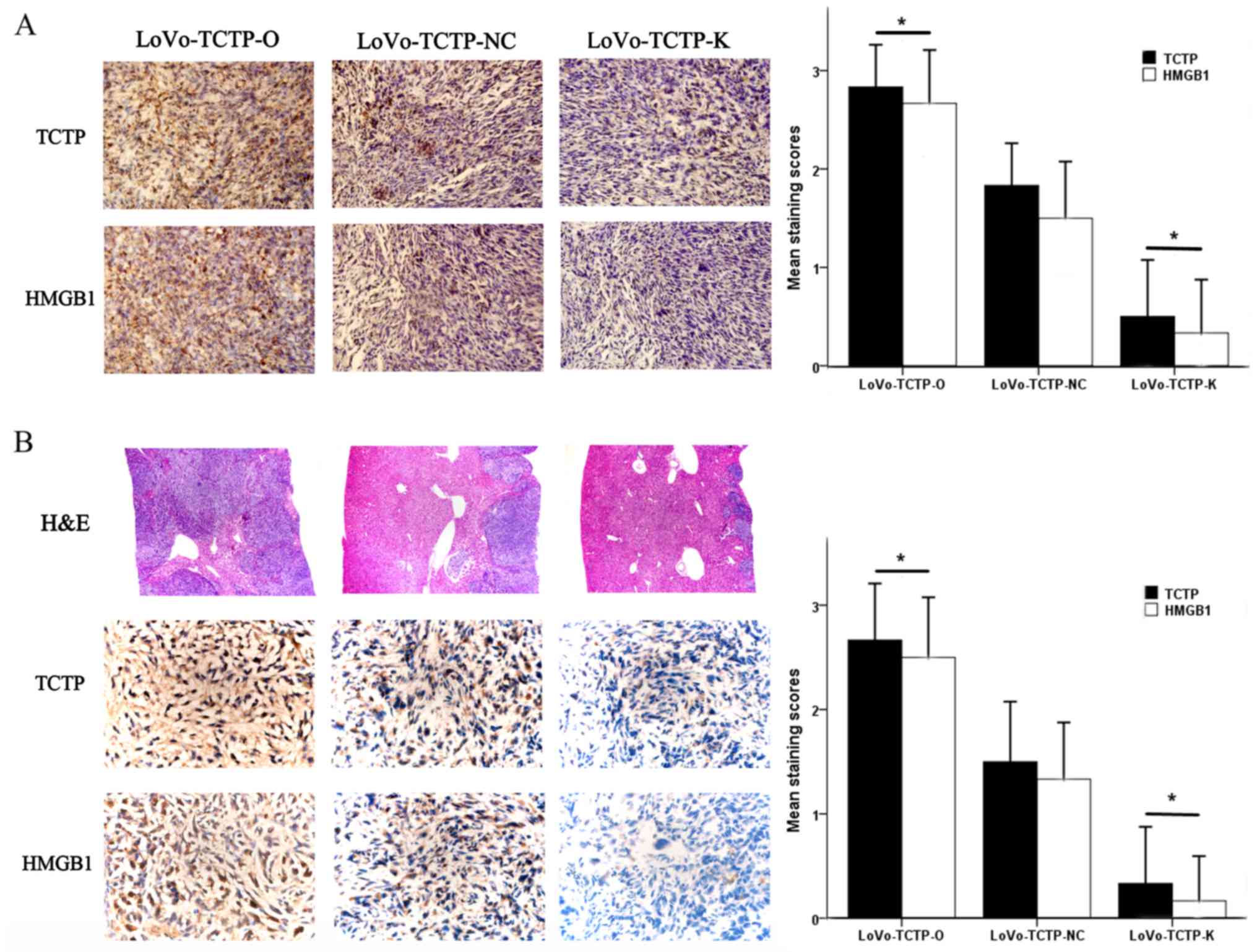 | Figure 6Effect of TCTP on the expression of
HMGB1 in xenograft tumors in nude mice. (A) Expression of TCTP and
HMGB1 in the developed tumors in the nude mouse spleen were
detected 5 weeks following inoculation with LoVo cells
(magnification, ×400). *P<0.05, compared with mice
receiving normal control LoVo cells. (B) Expression of TCTP and
HMGB1 in metastatic tumors in the nude mouse liver were detected 5
weeks following inoculation with LoVo cells (magnification, ×400).
*P<0.05, compared with mice receiving normal control
LoVo cells. The H&E panel shows sections of liver stained with
H&E (magnification, ×100). TCTP-NC, normal control LoVo cells;
TCTP-K, LoVo cells transfected with TCTP-knockdown vector; TCTP-O,
LoVo cells transfected with TCTP-overexpression vector; TCTP,
translationally controlled tumor protein; HMGB1, high mobility
group box 1; H&E, hematoxylin and eosin. |
Discussion
In the present study, it was demonstrated that the
expression level of TCTP was significantly increased in CRC
tissues, which supports others' and our previous reports (7,8,24).
It was found that the serum level of HMGB1 was markedly elevated in
patients with CRC, which is consistent with previous reports
(25,26). The results also showed that the
expression of TCTP and the concentration of HMGB1 were markedly
higher in the samples from patients with CRC with distant
metastasis, indicating that they are associated with the clinical
severity and prognosis of CRC. In addition, it was found that there
was a strong positive correlation between the two increased
proteins. This is the first time, to the best of our knowledge,
that TCTP and HMGB1 have been assessed together in an integrated
study to test the hypothesis that they may have an interconnection
in the process of tumor progression and metastasis. These
preliminary findings provide incentive for further investigation of
the interaction between the two tumor-promoting factors.
From the in vitro experiments, it was noted
that TCTP induced the cytoplasmic translocation of HMGB1 and its
further release into the extracellular environment. These findings
support the hypothesis that HMGB1 can be secreted from CRC cells,
which has been documented previously (14,27),
and provide important novel insight that the secretion of HMGB1 is
regulated by TCTP. Furthermore, in xenograft tumors in nude mice,
it was found that augmentation of expression of TCTP promoted liver
metastasis of CRC cells, which was accompanied by a marked increase
in the expression of HMGB1. This finding confirmed that TCTP
regulated the behavior of HMGB1, which may produce synergistic
effects on the formation and metastasis of CRC.
It appears that, when secreted from cells, HMGB1
becomes a multifunctional cytokine for regulating cell
proliferation, survival and migration (12–17).
Previous evidence indicates that extracellular HMGB1 is not only
involved in chronic inflammatory-reparative responses, which
contribute to tumor cell survival and metastasis (27–29),
but also induces apoptosis in immune cells, resulting in an
attenuation of anticancer immune responses (30). TLR and RAGE, the main receptors of
HMGB1, also contribute to the progression and metastasis of CRC
(31–35). It has been shown that the
HMGB1-TLR-RAGE tripod frequently activates the downstream NF-κB
signaling pathway (17–19), and it has subsequently been
demonstrated that dimerized TCTP, the biologically active form of
TCTP, can also activate the NF-κB pathway and induce inflammation
(36). However, there is no
literature concerning whether the NF-κB pathway is involved in
TCTP-promoted tumor cell invasion and metastasis. The present study
provided evidence that TCTP stimulated the activation of NF-κB in
colon adenocarcinoma cells, and that this was abrogated by
antibodies against HMGB1, TLR4 or RAGE. These results indicated
that TCTP can induce the activation of NF-κB through the mediation
of HMGB1 and its receptors TLR4 and RAGE. In addition, it was found
that TCTP and the successive activation of the
HMGB1-TLR4/RAGE-NF-κB pathway enhanced the invasion potential of
LoVo cells, whereas the specific NF-κB inhibitor Bay117082
attenuated the increased invasiveness of tumor cells. This
indicated that the activation of NF-κB signaling is essential for
TCTP-mediated tumor cell migration and invasion.
Although experimental studies have revealed the
importance of NF-κB in the initiation and propagation of CRC, the
mechanisms underlying how NF-κB promotes tumor metastasis remain to
be fully elucidated. There are several reasons that may account for
NF-κB-facilitated tumor metastasis. Firstly, NF-κB orchestrates a
variety of cellular effectors of inflammation to constitute a local
environment that may promote cancer cell invasiveness (37–39).
Secondly, NF-κB upregulates the expression of target genes that are
involved in tumor metastasis, for example MMPs (20,40,41).
Finally, increasing data suggest that NF-κB is implicated in the
progression of cancer epithelial-to-mesenchymal transition, which
is an essential process in the initiation of tumor spread (42–44).
Previous studies have indicated that HMGB1 and TCTP can induce
epithelial-to-mesenchymal transition and thus promote tumor cell
migration and invasion (31,42,45).
Therefore, NF-κB and its upstream and downstream network present a
rational target to curb the progression of CRC.
Of note, HMGB1 can be modified posttranslationally
in multiple ways, which may in turn determine the location and
secretion of HMGB1 and its interactions with DNA and other proteins
(46). The discrepancy in
bioactivities of HMGB1 may be associated with different cell types
or tissue sources, or its responses to different stimuli (47). In addition, TCTP has diverse
biological activities on account of its different forms (36), localizations (7) or extracellular environments (2). Therefore, future investigations
require that all these factors are taken into consideration, and
the design of further investigations requires the use of strategies
to inhibit the expression, release or activity of these dynamic
proteins for comprehensive assessment of their functions and
interactions in regulating tumor progression.
In conclusion, the present study indicated that TCTP
affected the behaviors of HMGB1, and facilitated CRC cell invasion
via HMGB1-mediated activation of the NF-κB pathway. These findings
provide novel clues for elucidating the mechanism of TCTP-induced
metastasis of CRC, and suggest that TCTP can be used as a potential
target for anticancer therapy.
Acknowledgments
The authors would like to thank the staff of
Digestive Endoscopy Center of Nanfang Hospital and Department of
Pathology, Nanfang Hospital for providing the tissue specimens.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81000952 and 81772133), the
Guangdong Natural Science Fund (grant no. S2013030013217), the
Guangdong Natural Science Fund (grant no. 2016A030313529) and the
Guangxi Natural Science Fund (grant nos. 2017GXNSFDA198051 and
2016GXNSFAA380313).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
MH, QM and YW were involved in the conception and
design of the study; MH, YG, QD, QM, RL, XS, ZZ and WX performed
the experiments; MH, YG and QM analyzed the data; MH drafted the
manuscript; QM and YW reviewed and edited the manuscript. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The protocols for the collection of human tissues
were approved by the Ethics Committee of Nanfang Hospital, Southern
Medical University, and written informed consent was signed by each
participant prior to their inclusion in the study. The animal
experiment was approved by the Association for the Accreditation
and Assessment of Laboratory Animal Care (Guangzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
CRC
|
colorectal cancer
|
|
TCTP
|
translationally controlled tumor
protein
|
|
HMGB1
|
high mobility group box 1
|
|
NF-κB
|
nuclear factor-κB
|
|
IκB
|
inhibitor of NF-κB
|
|
TLR4
|
Toll-like receptor 4
|
|
RAGE
|
receptor for advanced glycation end
products
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
LDH
|
lactate dehydrogenase
|
References
|
1
|
Siegel R, DeSantis C, Virgo K, Stein K,
Mariotto A, Smith T, Cooper D, Gansler T, Lerro C, Fedewa S, et al:
Cancer treatment and survivorship statistics, 2012. CA Cancer J
Clin. 62:220–241. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Acunzo J, Baylot V, So A and Rocchi P:
TCTP as therapeutic target in cancers. Cancer Treat Rev.
40:760–769. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Amson R, Pece S, Marine JC, Di Fiore PP
and Telerman A: TPT1/TCTP-regulated pathways in phenotypic
reprogramming. Trends Cell Biol. 23:37–46. 2013. View Article : Google Scholar
|
|
4
|
Chan TH, Chen L and Guan XY: Role of
translationally controlled tumor protein in cancer progression.
Biochem Res Int. 2012:3693842012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Amson R, Pece S, Lespagnol A, Vyas R,
Mazzarol G, Tosoni D, Colaluca I, Viale G, Rodrigues-Ferreira S,
Wynendaele J, et al: Reciprocal repression between P53 and TCTP.
Nat Med. 18:91–99. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chan TH, Chen L, Liu M, Hu L, Zheng BJ,
Poon VK, Huang P, Yuan YF, Huang JD, Yang J, et al: Translationally
controlled tumor protein induces mitotic defects and chromosome
missegregation in hepatocellular carcinoma development. Hepatology.
55:491–505. 2012. View Article : Google Scholar
|
|
7
|
Xiao B, Chen D, Luo S, Hao W, Jing F, Liu
T, Wang S, Geng Y, Li L, Xu W, et al: Extracellular translationally
controlled tumor protein promotes colorectal cancer invasion and
metastasis through Cdc42/JNK/MMP9 signaling. Oncotarget.
7:50057–50073. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ma Q, Geng Y, Xu W, Wu Y, He F, Shu W,
Huang M, Du H and Li M: The role of translationally controlled
tumor protein in tumor growth and metastasis of colon
adenocarcinoma cells. J Proteome Res. 9:40–49. 2010. View Article : Google Scholar
|
|
9
|
Ueda M, Takahashi Y, Shinden Y, Sakimura
S, Hirata H, Uchi R, Takano Y, Kurashige J, Iguchi T, Eguchi H, et
al: Prognostic significance of high mobility group box 1 (HMGB1)
expression in patients with colorectal cancer. Anticancer Res.
34:5357–5362. 2014.PubMed/NCBI
|
|
10
|
Zhang Z, Wang M, Zhou L, Feng X, Cheng J,
Yu Y, Gong Y, Zhu Y, Li C, Tian L, et al: Increased HMGB1 and
cleaved caspase-3 stimulate the proliferation of tumor cells and
are correlated with the poor prognosis in colorectal cancer. J Exp
Clin Cancer Res. 34:512015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Süren D, Yıldırım M, Demirpençe Ö, Kaya V,
Alikanoğlu AS, Bülbüller N, Yıldız M and Sezer C: The role of high
mobility group box 1 (HMGB1) in colorectal cancer. Med Sci Monit.
20:530–537. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
van Beijnum JR, Buurman WA and Griffioen
AW: Convergence and amplification of toll-like receptor (TLR) and
receptor for advanced glycation end products (RAGE) signaling
pathways via high mobility group B1 (HMGB1). Angiogenesis.
11:91–99. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dumitriu IE, Baruah P, Valentinis B, Voll
RE, Herrmann M, Nawroth PP, Arnold B, Bianchi ME, Manfredi AA and
Rovere-Querini P: Release of high mobility group box 1 by dendritic
cells controls T cell activation via the receptor for advanced
glycation end products. J Immunol. 174:7506–7515. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kang HJ, Lee H, Choi HJ, Youn JH, Shin JS,
Ahn YH, Yoo JS, Paik YK and Kim H: Non-histone nuclear factor HMGB1
is phosphorylated and secreted in colon cancers. Lab Invest.
89:948–959. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee W, Ku SK, Bae JW and Bae JS:
Inhibitory effects of lycopene on HMGB1-mediated pro-inflammatory
responses in both cellular and animal models. Food Chem Toxicol.
50:1826–1833. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sims GP, Rowe DC, Rietdijk ST, Herbst R
and Coyle AJ: HMGB1 and RAGE in inflammation and cancer. Annu Rev
Immunol. 28:367–388. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nogueira-Machado JA and de Oliveira Volpe
CM: HMGB-1 as a target for inflammation controlling. Recent Pat
Endocr Metab Immune Drug Discov. 6:201–209. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nogueira-Machado JA, Volpe CM, Veloso CA
and Chaves MM: HMGB1, TLR and RAGE: A functional tripod that leads
to diabetic inflammation. Expert Opin Ther Targets. 15:1023–1035.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li Y, He J, Zhong D, Li J and Liang H:
High-mobility group box 1 protein activating nuclear factor-κB to
upregulate vascular endothelial growth factor C is involved in
lymphangiogenesis and lymphatic node metastasis in colon cancer. J
Int Med Res. 43:494–505. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang S, Liu Z, Wang L and Zhang X:
NF-kappaB signaling pathway, inflammation and colorectal cancer.
Cell Mol Immunol. 6:327–334. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Raina K, Agarwal C and Agarwal R: Effect
of silibinin in human colorectal cancer cells: Targeting the
activation of NF-κB signaling. Mol Carcinog. 52:195–206. 2013.
View Article : Google Scholar
|
|
22
|
Chang MS, Kim DH, Roh JK, Middeldorp JM,
Kim YS, Kim S, Han S, Kim CW, Lee BL, Kim WH, et al: Epstein-Barr
virus-encoded BARF-1 promotes proliferation of gastric carcinoma
cells through regulation of NF-κB. J Virol. 87:10515–10523. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shafiu Kamba A and Zakaria ZA: Osteoblasts
growth behaviour on bio-based calcium carbonate aragonite
nanocrystal. BioMed Res Int. 2014:2150972014. View Article : Google Scholar :
|
|
24
|
Bommer UA, Vine KL, Puri P, Engel M,
Belfiore L, Fildes K, Batterham M, Lochhead A and Aghmesheh M:
Translationally controlled tumour protein TCTP is induced early in
human colorectal tumours and contributes to the resistance of
HCT116 colon cancer cells to 5-FU and oxaliplatin. Cell Commun
Signal. 15:92017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang X, Yu J, Li M, Zhu H, Sun X and Kong
L: The association of HMGB1 expression with clinicopathological
significance and prognosis in Asian patients with colorectal
carcinoma: A meta-analysis and literature review. OncoTargets Ther.
9:4901–4911. 2016. View Article : Google Scholar
|
|
26
|
Lee H, Song M, Shin N, Shin CH, Min BS,
Kim HS, Yoo JS and Kim H: Diagnostic significance of serum HMGB1 in
colorectal carcinomas. PLoS One. 7:e343182012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Luo Y, Chihara Y, Fujimoto K, Sasahira T,
Kuwada M, Fujiwara R, Fujii K, Ohmori H and Kuniyasu H: High
mobility group box 1 released from necrotic cells enhances regrowth
and metastasis of cancer cells that have survived chemotherapy. Eur
J Cancer. 49:741–751. 2013. View Article : Google Scholar
|
|
28
|
Tang D, Kang R, Zeh HJ III and Lotze MT:
High-mobility group box 1 and cancer. Biochim Biophys Acta.
1799:131–140. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen RC, Yi PP, Zhou RR, Xiao MF, Huang
ZB, Tang DL, Huang Y and Fan XG: The role of HMGB1-RAGE axis in
migration and invasion of hepatocellular carcinoma cell lines. Mol
Cell Biochem. 390:271–280. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Völp K, Brezniceanu ML, Bösser S, Brabletz
T, Kirchner T, Göttel D, Joos S and Zörnig M: Increased expression
of high mobility group box 1 (HMGB1) is associated with an elevated
level of the antiapoptotic c-IAP2 protein in human colon
carcinomas. Gut. 55:234–242. 2006. View Article : Google Scholar
|
|
31
|
Sharma S, Evans A and Hemers E:
Mesenchymal-epithelial signalling in tumour microenvironment: Role
of high-mobility group Box 1. Cell Tissue Res. 365:357–366. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Han M, Song Y and Zhang X: Quercetin
suppresses the migration and invasion in human colon cancer Caco-2
cells through regulating toll-like receptor 4/nuclear factor-kappa
B pathway. Pharmacogn Mag. 12(Suppl 2): S237–S244. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yesudhas D, Gosu V, Anwar MA and Choi S:
Multiple roles of toll-like receptor 4 in colorectal cancer. Front
Immunol. 5:3342014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Deng R, Wu H, Ran H, Kong X, Hu L, Wang X
and Su Q: Glucose-derived AGEs promote migration and invasion of
colorectal cancer by up-regulating Sp1 expression. Biochim Biophys
Acta. 1861:1065–1074. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dahlmann M, Okhrimenko A, Marcinkowski P,
Osterland M, Herrmann P, Smith J, Heizmann CW, Schlag PM and Stein
U: RAGE mediates S100A4-induced cell motility via MAPK/ERK and
hypoxia signaling and is a prognostic biomarker for human
colorectal cancer metastasis. Oncotarget. 5:3220–3233. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee H and Lee K: Dimerized translationally
controlled tumor protein increases interleukin-8 expression through
MAPK and NF-κB pathways in a human bronchial epithelial cell line.
Cell Biosci. 8:132018. View Article : Google Scholar
|
|
37
|
Karin M and Greten FR: NF-kappaB: Linking
inflammation and immunity to cancer development and progression.
Nat Rev Immunol. 5:749–759. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mantovani A, Allavena P, Sica A and
Balkwill F: Cancer-related inflammation. Nature. 454:436–444. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vaiopoulos AG, Athanasoula KC and
Papavassiliou AG: NF-κB in colorectal cancer. J Mol Med (Berl).
91:1029–1037. 2013. View Article : Google Scholar
|
|
40
|
Chen S, Chen W, Zhang X, Lin S and Chen Z:
Overexpression of KiSS-1 reduces colorectal cancer cell invasion by
downregulating MMP-9 via blocking PI3K/Akt/NF-κB signal pathway.
Int J Oncol. 48:1391–1398. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Qin M and Liu S, Li A, Xu C, Tan L, Huang
J and Liu S: NIK- and IKKβ-binding protein promotes colon cancer
metastasis by activating the classical NF-κB pathway and MMPs.
Tumour Biol. 37:5979–5990. 2016. View Article : Google Scholar
|
|
42
|
Zhu L, Li X, Chen Y, Fang J and Ge Z:
High-mobility group box 1: A novel inducer of the
epithelial-mesenchymal transition in colorectal carcinoma. Cancer
Lett. 357:527–534. 2015. View Article : Google Scholar
|
|
43
|
Ma J, Gao Q, Zeng S and Shen H: Knockdown
of NDRG1 promote epithelial-mesenchymal transition of colorectal
cancer via NF-κB signaling. J Surg Oncol. 114:520–527. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yan Z, Yin H, Wang R, Wu D, Sun W, Liu B
and Su Q: Overexpression of integrin-linked kinase (ILK) promotes
migration and invasion of colorectal cancer cells by inducing
epithelial-mesenchymal transition via NF-κB signaling. Acta
Histochem. 116:527–533. 2014. View Article : Google Scholar
|
|
45
|
Bae SY, Kim HJ, Lee KJ and Lee K:
Translationally controlled tumor protein induces epithelial to
mesenchymal transition and promotes cell migration, invasion and
metastasis. Sci Rep. 5:80612015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Richard SA, Jiang Y, Xiang LH, Zhou S,
Wang J, Su Z and Xu H: Post-translational modifications of high
mobility group box 1 and cancer. Am J Transl Res. 9:5181–5196.
2017.
|
|
47
|
Huebener P, Pradere JP, Hernandez C, Gwak
GY, Caviglia JM, Mu X, Loike JD, Jenkins RE, Antoine DJ and Schwabe
RF: The HMGB1/RAGE axis triggers neutrophil-mediated injury
amplification following necrosis. J Clin Invest. 125:539–550. 2015.
View Article : Google Scholar : PubMed/NCBI
|