Introduction
Esophageal cancer is the eighth most common
malignancy and the sixth most common cause of cancer-associated
mortality worldwide (1). In the
Chinese population, esophageal squamous cell carcinoma (ESCC) is
the predominant histological subtype, which is responsible for
~210,000 cases of mortality each year, or 52% of all cases of
ESCC-associated mortality worldwide (2). The characteristics of ESCC include
being prone to invasive local growth, early lymphatic spread and
vascular invasion (3,4). ESCC has a very poor survival rate,
despite significant progress in multimodal therapy. Numerous
genetic alterations involved in the carcinogenesis of ESCCs have
been reported (5); however, the
exact molecular mechanisms remain unclear. Characterizing the
precise mechanisms of esophageal carcinogenesis may help to
identify biomarkers that can improve early diagnosis and targeted
therapy of ESCC.
Kinetochore-associated protein 1 (KNTC1) is a key
component of mitotic checkpoints, which ensures proper chromosomal
segregation during cell division (6). Both chromosomal segregation and cell
division are critical, highly ordered biological processes that are
dependent on numerous evolutionarily conserved protein complexes.
Many of the proteins that modulate the process of mitosis are
overexpressed in human malignancies compared with their normal
counterparts, and some have been reported to serve as oncogenes
(7). KNTC2 is known to serve a
critical role in chromosomal segregation during M phase (8). KNTC2 is upregulated in the tumor
tissues of patients with various types of cancer, including gastric
cancer, colorectal cancer, pancreatic cancer, hepatocellular
carcinoma (HCC), breast cancer and non-small cell lung carcinoma
(9-14). In addition, small interfering RNA
(siRNA)-mediated silencing of KNTC2 has been reported to suppress
cell proliferation and to induce apoptotic cell death in these
tumor cells (9,12). Furthermore, repeated administration
of KNTC2 siRNAs has been revealed to specifically inhibit tumor
growth in an orthotopic transplantation tumor model of HCC in nude
mice (15). Targeted knockdown of
kinetochore scaffold 1, which is widely expressed in various types
of primary cancer, induces apoptosis in human transformed or tumor
cell lines in vitro, and markedly impedes the growth of
implanted tumors in vivo (16). These studies demonstrated that
kinetochore proteins may serve as potential biomarkers for the
early diagnosis of cancer, and the characterization of the role of
kinetochore proteins may further contribute to the development of
novel personalized treatments for human malignancies.
KNTC1 is an evolutionarily conserved subunit of the
kinetochore protein complex, which is vital for spindle assembly
and chromosomal segregation. It elicits an inhibitory signal to
prevent the onset of anaphase, until all chromosomes have been
properly arranged in the spindle. Cells produce lagging chromosomes
and chromatin bridges after being injected with KNTC1-blocking
antibody, thus suggesting that KNTC1 is a vital component of the
mitotic checkpoint (6). However,
to the best of our knowledge, there are no previous reports
describing the significance of KNTC1 in human malignancies, or in
ESCC in particular.
The present study analyzed the expression levels of
KNTC1 in ESCC cell lines. Subsequently, lentivirus-mediated short
hairpin RNAs (shRNAs) were employed to investigate the effects of
KNTC1 knockdown on human ESCC cell viability and apoptosis in
vitro. In addition, gene set enrichment analysis (GSEA) was
used to infer the possible mechanisms underlying the involvement of
KNTC1 in ESCC tumorigenesis.
Materials and methods
Cell culture and human tissues
The human ESCC cell lines Eca-109, EC9706 and TE-1
were provided by the Cancer Institute and Hospital, Chinese Academy
of Medical Sciences (Beijing, China). Authentication of cell lines
was performed using short tandem repeat analysis. The three ESCC
cell lines were cultured in RPMI-1640 medium (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10%
fetal bovine serum (HyClone; GE Healthcare Life Sciences, Logan,
UT, USA) and antibiotics (100 U/ml penicillin G and 100
µg/ml streptomycin) at 37°C and 95% humidity, in an
atmosphere containing 5% CO2. Normal esophageal tissues
were obtained via endoscopic biopsy from two healthy volunteers
(one male, 37 years old; one female, 44 years old; recruited on
October 21, 2017). Informed consent was obtained from the
volunteers. The present study was approved by the ethics committees
of the Capital Medical University Affiliated Beijing Friendship
Hospital (Beijing, China).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Cells were pelleted using a cell scraper. Total RNA
was isolated from cell pellets using TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. Total RNA concentration and purity were
measured with a UV spectrophotometer (Ultrospec 2100 Pro; GE
Healthcare, Chicago, IL, USA). RT was performed using the
PrimeScript RT reagent kit (Takara Biotechnology Co., Ltd., Dalian,
China), according to the manufacturer's protocol. qPCR was
performed using SYBR® Premix Ex Taq™ II (Takara
Biotechnology Co., Ltd.) and the CFX96™ Real-Time system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA), according to the
manufacturers' protocols. The specific primers used were as
follows: KNTC1, forward 5′-GCAACAACTTGTAGACGACGCT-3′, reverse
5′-TCAATCCAAGAACTGCCACTG-3′; and GAPDH, forward
5′-TGACTTCAACAGCGACACCCA-3′ and reverse
5′-CACCCTGTTGCTGTAGCCAAA-3′. RT-qPCR consisted of an initial
denaturation step at 95°C for 15 sec, followed by 45 cycles at 95°C
for 5 sec and 60°C for 30 sec. PCR reactions were optimized for the
number of cycles to ensure product intensity within the logarithmic
phase of amplification. The relative mRNA expression levels of
KNTC1 were normalized to the GAPDH internal control. The relative
mRNA expression levels were calculated using the 2−ΔΔCq
method (17). The PCR products of
KNTC1 and GAPDH were 270 and 121 bp, respectively.
Recombinant lentivirus vector production
and transduction
The KNTC1-targeted short hairpin RNA (shRNA)
sequence (5′-TGAGTTTATGGGATATTTA-3′) was designed by Shanghai
GeneChem Co. Ltd. (Shanghai, China). The shRNA sequences were
synthesized and subsequently cloned into the pGCSIL-green
fluorescent protein (GFP) lentiviral vector by Shanghai GeneChem
Co. Ltd. The nonsense shRNA (5′-TTCTCCGAACGTGTCACGT-3′), which
shared no homology with any known human genes, was generated as a
negative control (NC) (Shanghai GeneChem Co. Ltd.). For recombinant
lentiviral transduction, Eca-109 and TE-1 cells were seeded into
6-well plates until cell confluence reached 70%, and were then
infected with the KNTC1-shRNA-lentivirus or the NC lentivirus at a
multiplicity of infection of 50. Transduction effects were
determined by detecting GFP expression 24-72 h post-transduction
using fluorescence and light microscopy. Cells were harvested 72 h
post-transduction, and the knockdown effect was confirmed with
RT-qPCR and western blot analysis.
Western blot analysis
Total protein was isolated from cells using the
total protein extraction kit (Nanjing Keygen Biotech Co., Ltd.,
Nanjing, China), according to the manufacturer's protocol. Briefly,
the cells were collected and were lysed in lysis buffer containing
150 mM sodium chloride, 0.1 M Tris, 1% (v/v) Tween-20, 50 mM
diethyldithiocarbamic acid, 1 mM ethylenediaminetetraacetic acid
and protease inhibitors at pH 8.0. The lysates were kept on ice for
30 min and were centrifuged at 12,000 × g for 15 min at 4°C, and
the supernatants were collected. Protein concentrations were
measured using a Pierce bicinchoninic acid protein assay kit
(Pierce; Thermo Fisher Scientific, Inc.). The protein samples (50
µg/lane) were separated by 10% SDS-PAGE according to
Laemmli's method (18) and were
then electrophoretically transferred to polyvinylidene fluoride
membranes (EMD Millipore, Billerica, MA, USA). The membranes were
blocked with 5% non-fat dry milk in Tris-buffered saline containing
0.1% Tween-20 (TBST) buffer for 2 h at room temperature, followed
by incubation with primary antibodies at a dilution of 1:2,000 at
4°C overnight. After being washed with TBST, the membranes were
incubated with horseradish peroxidase-conjugated immunoglobulin G
(IgG) secondary antibody (anti-rabbit IgG, cat. no. sc-2004 and
anti-mouse IgG, cat. no. sc-2005; dilution 1:5,000; Santa Cruz
Biotechnology, Inc. Dallas, TX, USA) for 1 h at room temperature.
Immunoblots were developed using an enhanced chemiluminescence
(ECL) reagent (ECL Plus; GE Healthcare). KNTC1 antibody (cat. no.
ab85996) was purchased from Abcam (Cambridge, MA, USA) and GAPDH
antibody (cat. no. sc-32233) was purchased from Santa Cruz
Biotechnology, Inc.
Cell growth and viability assays
Cell growth was measured using multiparametric
high-content screening (HCS), according to a previously reported
method (19). Briefly, following
lentiviral transduction, the cells were seeded at 2,000 cells/well
in 96-well plates and were incubated at 37°C in an atmosphere
containing 5% CO2 for 5 days. The 96-well plates were
scanned using Celigo Image Cytometer (version 4.1.3.0; Nexcelom
Bioscience LLC, Lawrence, MA, USA) each day. The system includes a
computerized fluorescence-imaging microscope that automatically
identifies stained cells, and reports the intensity and
distribution of the stained cells in each well. Fluorescence images
were acquired for each channel (magnification, ×20) using suitable
filters.
Cell viability was measured using the MTT assay
post-lentiviral transduction. Briefly, cells in the logarithmic
growth phase were seeded in 96-well plates at a density of
1.0×104 cells/ml in 200 µl medium and were
incubated at 37°C in an atmosphere containing 5% CO2. Subsequently,
the medium was replaced with serum-free RPMI-1640 containing 0.2
mg/ml MTT at 24, 48, 72, 96 and 120 h, and cells were incubated for
an additional 4 h at 37°C. After removal of the supernatant, 150
µl dimethyl sulfoxide was added to each well, in order to
completely dissolve the formazan. The plates were lightly vortexed
and agitated for 10 min, and optical density values were detected
at 490 nm using a microplate reader (Bio-Rad Laboratories,
Inc.).
Analysis of cell apoptosis and
caspase-3/7 activities
Cell apoptosis was assessed by staining cells with
Annexin V-allophycocyanin (APC) (eBioscience; Thermo Fisher
Scientific, Inc.) and was detected using FACScan flow cytometry
(FCM; BD Biosciences, Franklin Lakes, NJ, USA). Briefly, cells were
plated in 6-well plates at a density of 2.0×104 cells/ml
in 3 ml medium. After 5 days of transduction with the
KNTC1-shRNA-lentivirus or NC-shRNA-lentivirus, the cells were
harvested and washed twice with ice-cold PBS. The cells were then
resuspended and adjusted to 1×106/ml with 1X staining
buffer. A 100-µl cell suspension was stained with 5
µl Annexin V-APC for 15 min at room temperature in the dark
and cell apoptosis was detected using FCM (Guava®
easyCyte HT; EMD Millipore) within 1 h.
Caspase-3 and caspase-7 activities of Eca-109 and
TE-1 cells following KNTC1-shRNA-lentiviral transduction were
detected using the Caspase-Glo® 3/7 assay (Promega
Corporation, Madison, WI, USA), according to a previously reported
method (20). Briefly, 72 h after
lentiviral transduction, cells were seeded in 96-well microplates
(Promega Corporation) at 10,000 cells/well. Briefly, the
Caspase-Glo® 3/7 reagent (100 µl/well) was added
to each well, and the plates were gently agitated for 2 h at
20-25°C. Finally, the luminescence was detected using a
plate-reading luminometer.
GSEA
An online ESCC dataset with supplementary clinical
information was downloaded from The Cancer Genome Atlas (TCGA) and
was used for GSEA. The dataset contained mRNA profiling data for 97
patients with ESCC from TCGA and was downloaded from the API of
cBioPortal (http://www.cbioportal.org/webAPI). A Pearson
correlation analysis was performed to examine gene-gene expression
correlations. Hierarchical clustering was performed to distinguish
different subgroups, according to the expression levels of the
given genes. Four gene sets were downloaded to perform the GSEA
analysis, including mitotic spindle, hypoxia, DNA repair and
mismatch repair, which were located in the Molecular Signatures
Database (MSigDB) and the Kyoto Encyclopedia of Genes and Genomes
(KEGG), respectively (21,22). The GSEA was performed to evaluate
the correlation between KNTC1 expression and these four gene
sets.
Statistical analysis
All of the results are presented as the mean values
of at least three independent experiments using cells from separate
cultures. Data are presented as the means ± standard deviation.
Differences between two groups were analyzed using Student's
t-test. For comparisons of more than two groups, repeated measures
analysis of variance, followed by Tukey's multiple comparison test,
was conducted. P<0.05 was considered to indicate a statistically
significant difference. Statistical analyses were performed using
SPSS standard version 16.0 (SPSS, Inc., Chicago, IL, USA) and R
3.1.2 software (www.r-project.org).
Results
Expression levels of KNTC1 in ESCC cell
lines and normal esophageal tissues
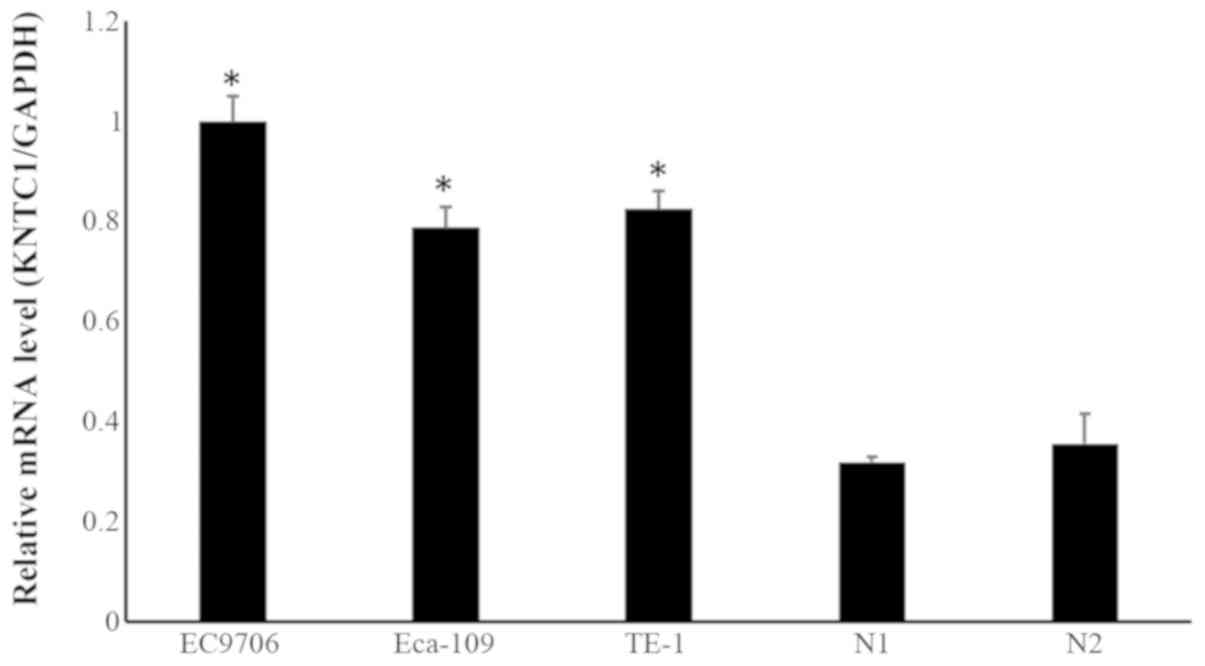
The mRNA expression levels of KNTC1 were assessed in
the ESCC cell lines Eca-109, EC9706 and TE-1, and in two normal
esophageal tissues using RT-qPCR. The results revealed that KNTC1
expression was significantly higher in ESCC cell lines compared
with in normal esophageal tissues (Fig. 1).
Lentivirus-mediated shRNA specifically
inhibits KNTC1 expression
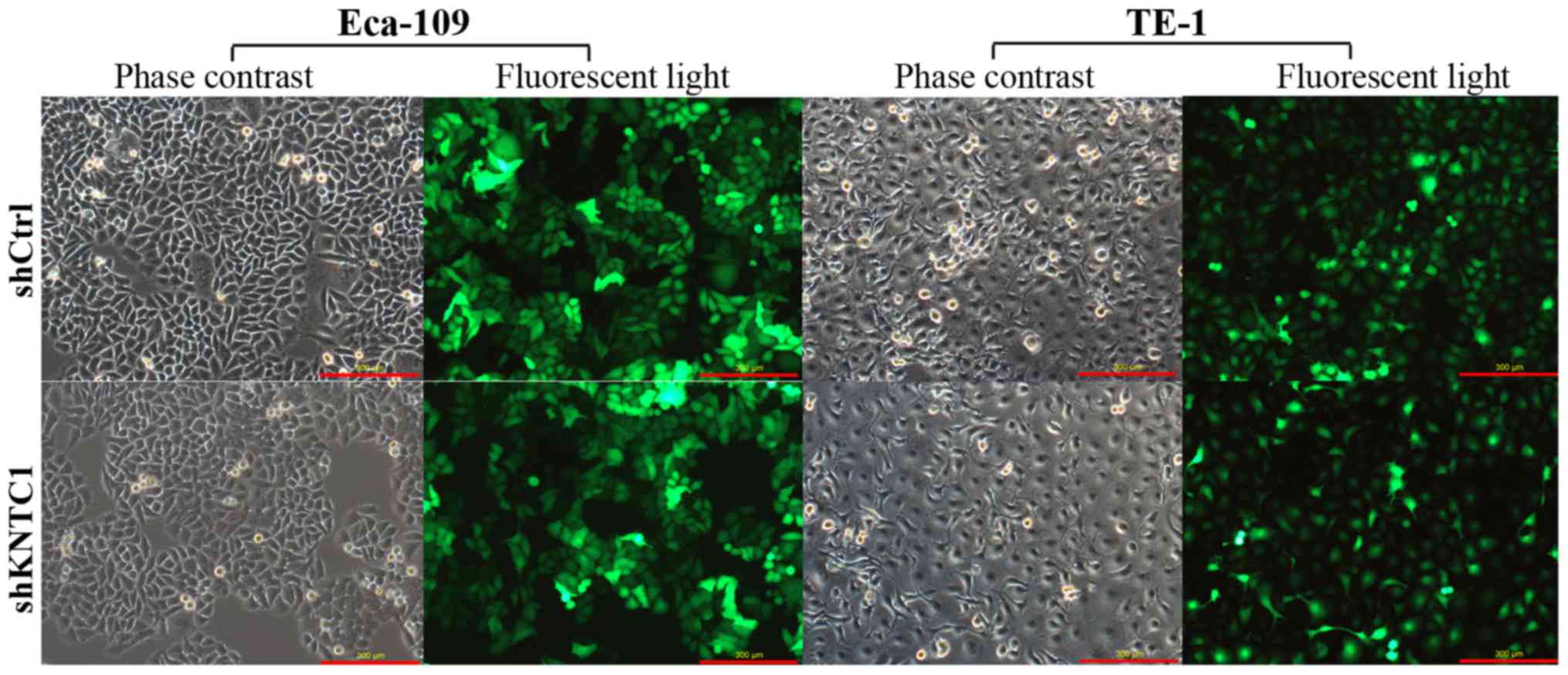
To explore the role of KNTC1 knockdown in the
biological behaviors of ESCC cells, a lentivirus-mediated shRNA
that targeted KNTC1 was introduced into Eca-109 and TE-1 cells,
both of which expressed high levels of KNTC1. A fluorescence
microscope was used to determine the transduction efficacy of the
recombinant lentivirus. Transduction efficiencies reached 80% in
both cell lines. As shown in Fig.
2, by day 3 post-transduction with KNTC1-shRNA or NC-shRNA
lentiviruses, the proportion of infected cells was >80%.
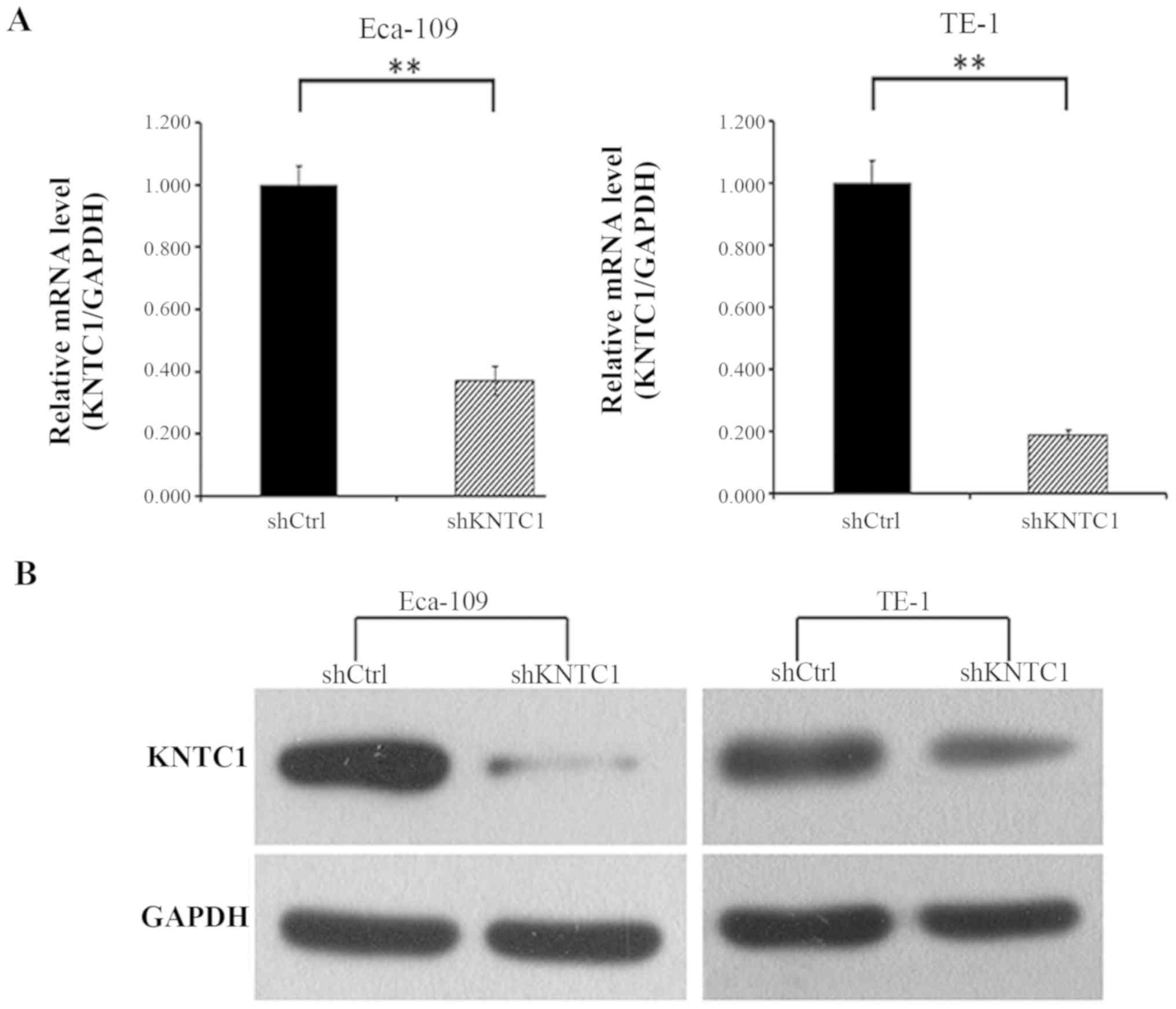
Subsequently, the knockdown efficacy of KNTC1-shRNA was confirmed
by RT-qPCR and western blotting. As shown in Fig. 3A, the mRNA expression levels of
KNTC1 in Eca-109 and TE-1 cells transduced with the KNTC1-shRNA
lentivirus were decreased by ~63 and 81%, respectively, compared
with in the cells transduced with the NC-shRNA lentivirus after 72
h (P<0.01). KNTC1 protein expression was also examined by
western blotting in these cells; the protein expression levels of
KNTC1 were significantly reduced in KNTC1-shRNA
lentivirus-transfected cells, thus indicating the effective
knockdown of KNTC1 (Fig. 3B).
Knockdown of KNTC1 inhibits growth and
viability of ESCC cells
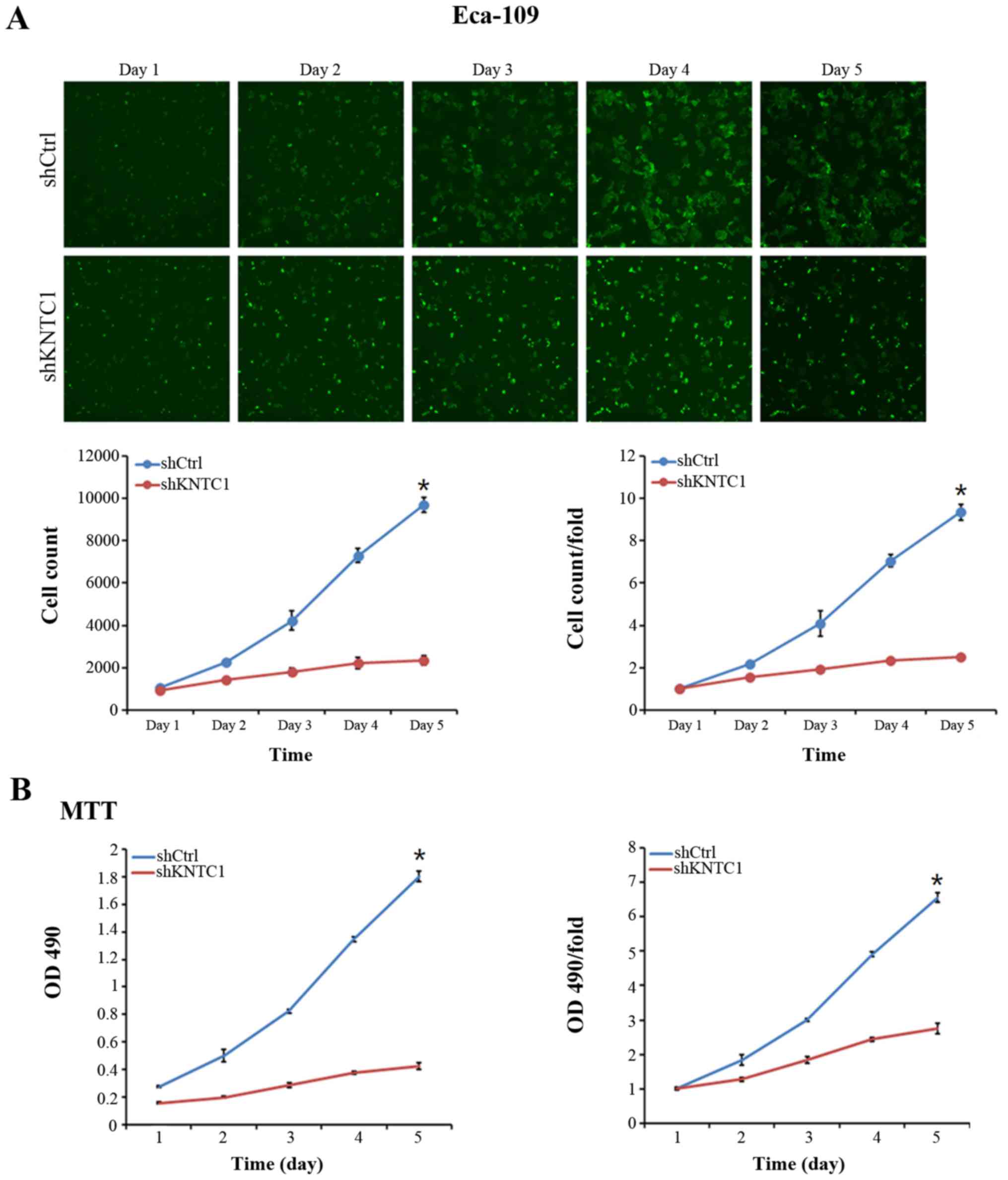
HCS was performed to assess the effects of KNTC1
knockdown on ESCC cell growth in vitro. Eca-109 and TE-1
cells expressing either KNTC1-shRNA or NC-shRNA lentiviruses were
plated in 96-well plates and were assessed using HCS every day for
5 days. Cell growth rate was defined as the ratio of the cell count
on a certain day to the cell count on the first day. As illustrated
in Fig. 4A, the growth rates of
Eca-109 cells transduced with the KNTC1-shRNA lentivirus were
significantly reduced compared with in the cells transduced with
the NC-shRNA lentivirus (P<0.01). As shown in Fig. 4B, similar results were observed
using the MTT assay, which further corroborated the negative effect
of KNTC1 knockdown on Eca-109 cell viability (P<0.01). Taken
together, these results suggested that KNTC1 knockdown
significantly inhibited the viability of Eca-109 and TE-1 cells,
thus indicating that KNTC1 may serve a critical role in the growth
and viability of ESCC cells.
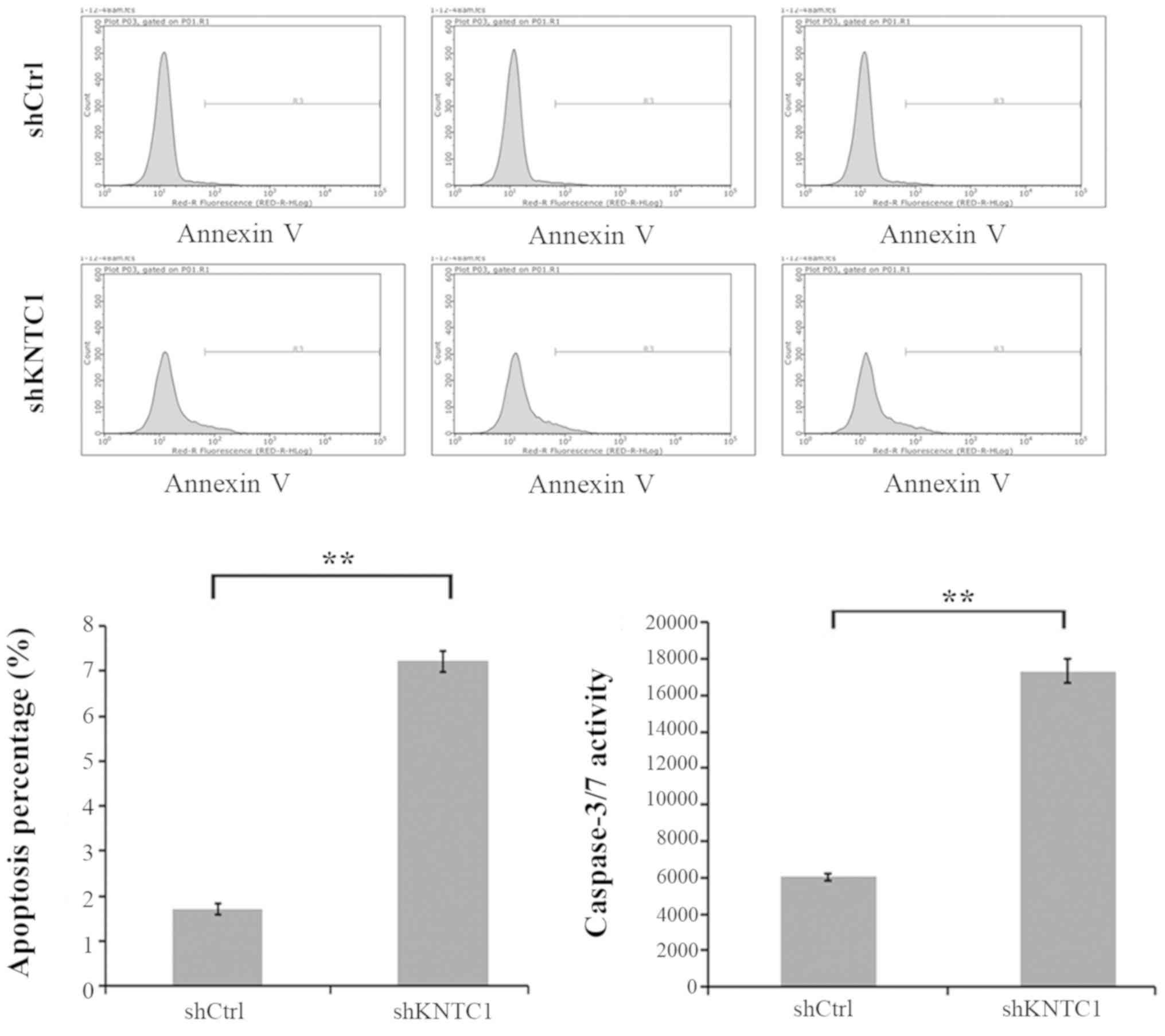
Knockdown of KNTC1 in ESCC cells
increases cell apoptosis
To investigate whether KNTC1 expression affects
apoptotic cell death in ESCC cells, KNTC1 was knocked down in
Eca-109 and TE-1 cells. Cell apoptosis was assessed with Annexin V
staining and FCM. As shown in Fig.
5, cell apoptosis of Eca-109 was significantly increased in the
KNTC1-shRNA lentivirus group compared with in the NC-shRNA
lentivirus group (P<0.01). TE-1 cell line exhibited similar
results (data not shown). To further confirm the ability of KNTC1
knockdown to induce apoptosis, caspase-3/7 activity analysis was
performed. In accordance with the aforementioned observations,
caspase-3/7 activity was markedly elevated in the KNTC1-shRNA
lentivirus group compared with in the NC-shRNA lentivirus group
(P<0.01). These results indicated that KNTC1 expression may be a
determinant of cell apoptosis in Eca-109 and TE-1 cells.
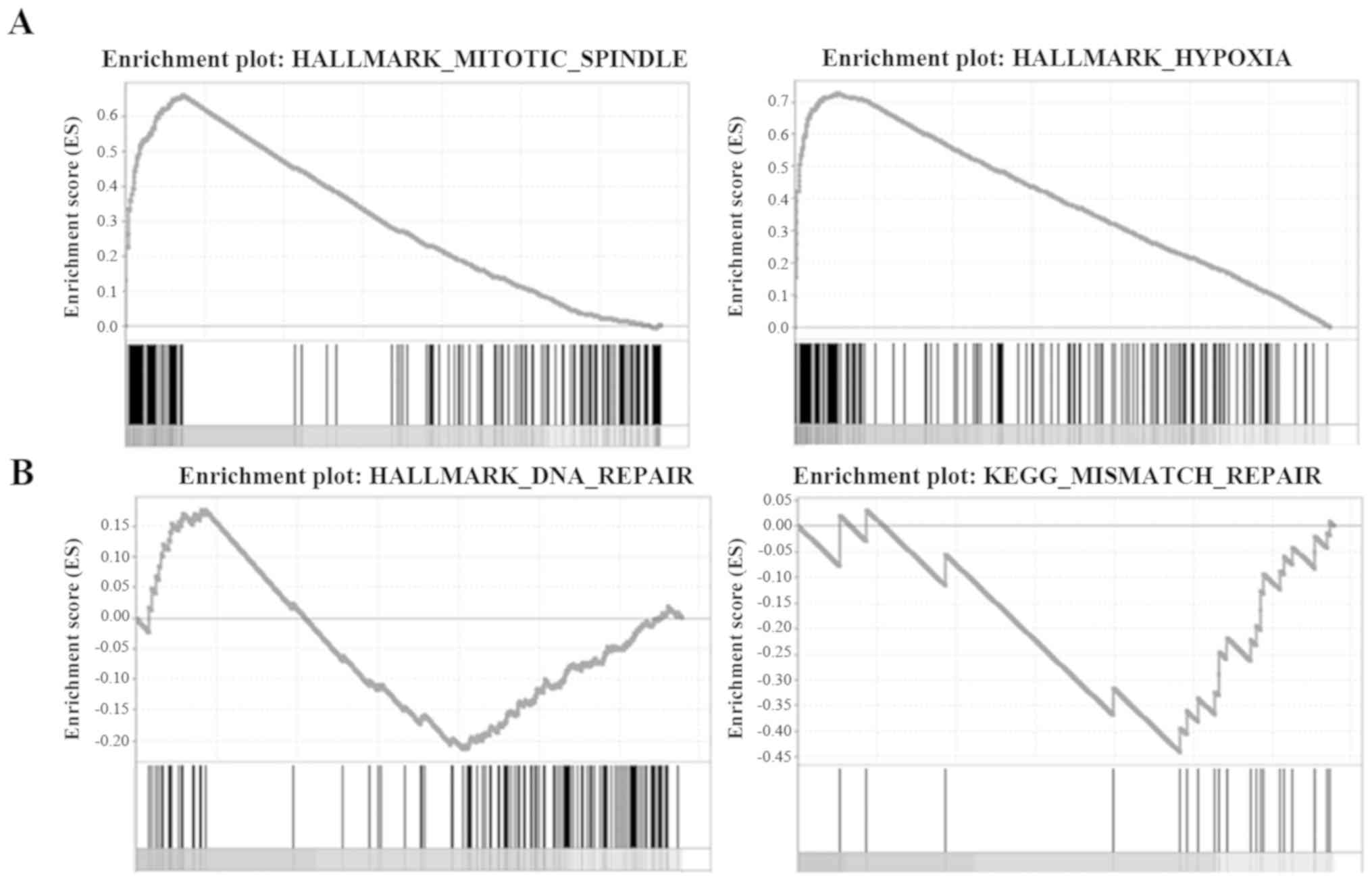
Association between KNTC1 expression and
mitotic spindle, hypoxia, DNA repair and mismatch repair gene
sets
Four gene sets were downloaded to perform the GSEA,
including the mitotic spindle, hypoxia, DNA repair and mismatch
repair gene sets. The normalized enrichment scores (NES) of the
mitotic spindle and hypoxia gene sets were 1.016 (P<0.001) and
1.004 (P=0.005), respectively (Fig.
6A), indicating that KNTC1 expression may be positively
correlated with the mitotic spindle and hypoxia pathways. For the
DNA repair and mismatch repair gene sets, the NES were −1.064
(P=0.021) and −1.093 (P=0.015), respectively (Fig. 6B), indicating that KNTC1 expression
was negatively correlated with the DNA repair and mismatch repair
pathways.
Discussion
Proper and accurate chromosomal segregation is
essential for maintaining genomic integrity during cell division. A
properly functioning centromere and its associated kinetochore are
crucial in determining the fidelity of chromosomal segregation
during mitosis. This step is under the control of numerous proteins
and protein complexes, which are localized to the kinetochores and
monitor the accuracy of this process (23). Defects in centromere- and
kinetochore-associated proteins have been proposed to contribute to
aneuploidy and chromosomal instability, which are associated with
cancer progression in several types of human tumor (24,25).
The presence of aneuploid cells is detected in ~90% of solid tumors
and ~50% of hematopoietic cancers (26). Chromosomal instability is believed
to be the driver of cancer predisposition, progression and
intratumoral heterogeneity, whereas aneuploidy and chromosomal
instability have been attributed to poor patient prognosis,
metastasis and chemotherapeutic resistance (27).
In a previous study, upon analyzing the publicly
available expression compendia of human normal tissues and cancer
samples, a group of kinetochore genes, which encode proteins
functioning exclusively at the kinetochores, has been revealed to
be concurrently upregulated in cancers (28). Their upregulation coincides with
the increased expression of genes associated with the cell cycle
and DNA replication, thus suggesting a widespread activation of the
cell division program potentially readying the cells for division
whenever the signal for cell cycle entry is received (28). These kinetochore proteins have been
noted for their involvement in increasing aneuploidy rates and the
risk of tumor progression. KNTC1 is a core kinetochore protein that
is required for faithful chromosomal segregation and spindle
assembly. Experimental evidence has indicated that KNTC1 functions
in a similar manner to that of the Drosophila rough deal
protein. The three activities that have so far been ascribed to
KNTC1 are as follows: i) Recruiting cytoplasmic dynein to the
kinetochore; ii) participating in the poleward movement of
chromosomes during mitosis; and iii) maintaining a functional
metaphase checkpoint (29).
However, to the best of our knowledge, no functional information is
available to date regarding the function of KNTC1 in human cancer,
particularly in ESCC.
The present study examined the mRNA expression
levels of KNTC1 in three ESCC cell lines and revealed that it was
expressed in all tested cell lines. To explore the role of KNTC1 in
ESCC, a KNTC1-shRNA lentiviral vector was constructed, which
efficiently silenced KNTC1 in Eca-109 and TE-1 cell lines. HCS and
MTT were performed to assess the effects of KNTC1 knockdown on ESCC
cell growth and viability. Compared with the NC-shRNA
lentivirus-transduced cells, KNTC1-shRNA-transduced cells exhibited
significantly reduced cell growth and viability. In addition,
knockdown of KNTC1 increased apoptosis of Eca-109 and TE-1 cells.
Taken together, these results strongly suggested that KNTC1 may
serve an onco-genic role in cell growth, viability and apoptosis in
ESCC.
To further explore the mechanisms underlying
KNTC1-mediated promotion of ESCC tumorigenesis, a GSEA was
conducted using online ESCC datasets from TCGA. Three hallmark
MSigDB gene sets and one KEGG gene set were adopted for analysis,
including the mitotic spindle, hypoxia, DNA repair and mismatch
repair gene sets. The results revealed that KNTC1 expression was
positively correlated with the mitotic spindle and hypoxia gene
sets, and negatively correlated with the DNA repair and mismatch
repair gene sets. Hypoxia signaling regulates diverse biological
processes, such as glycolysis, angiogenesis and invasion, and leads
to genomic instability, promotion of cancer progression, increased
metastatic potential, increased resistance to chemotherapy and
radiation, and poor prognosis (30). A previous study revealed that
hypoxia contributes to the metastasis and angiogenesis of ESCC
(31). Hypoxia is known to induce
phosphorylation of histone H2AX, which not only serves as a DNA
damage marker but also has a critical role in mediating homologous
DNA repair (32). Moderate hypoxia
can lead to replication stress and activation of the DNA damage
repair pathway proteins (33).
Conversely, hypoxia-reoxygenation induces DNA damage through
reactive oxygen species (34). DNA
mismatch repair (MMR) is a highly conserved biological pathway that
has a critical role in maintaining the fidelity of DNA replication,
mutation avoidance and genomic stability. MMR also suppresses
homologous recombination and was recently reported to serve a role
in DNA damage signaling. Defects in MMR are associated with
genome-wide instability, predisposition to several types of human
cancer, including ESCC, and resistance to certain chemotherapeutic
agents (35,36). The cellular response to DNA damage
is to limit viability, whilst initiating DNA repair. The present
GSEA results detected increases in the mitotic spindle and hypoxia
gene sets, and decreases in the DNA repair and mismatch repair gene
sets in response to KNTC1 overexpression in ESCC. Therefore, it was
inferred that KNTC1 overexpression may induce genomic instability
and DNA damage through increased mitotic spindle and hypoxia
pathways, which cannot be corrected by the decreased DNA repair and
mismatch repair pathways, which may eventually lead to
carcinogenesis and development of ESCC. However, the key molecular
mechanisms underlying the malignant functions of KNTC1 require
further investigation. We aim to further explore the molecular
mechanisms, particularly the effects of KNTC1 on hypoxia and DNA
repair.
In conclusion, the present study confirmed that
knockdown of KNTC1 expression by shRNA inhibited cell viability and
induced cell apoptosis in ESCC cell lines. In addition, GSEA of
online ESCC datasets revealed that KNTC1 overexpression was
associated with increases in the mitotic spindle and hypoxia
pathways, and decreases in the DNA repair and mismatch repair
pathways. However, further studies are required to reveal the
precise mechanism of KNTC1 in the carcinogenesis and development of
ESCC.
Funding
The present study was supported by the Beijing
Municipal Administration of Hospitals Youth Programme (grant no.
QML20150105) and the Beijing Excellent Talents Training Program
(grant no. 2015000021469G233).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CTL and STZ made substantial contributions to the
conception and design of the study. CTL performed the majority of
the study and wrote the manuscript. LM performed the GSEA. YJW, PL
and YDW were involved in designing the experiments, analyzing the
data and editing the manuscript. All authors read and approved the
final draft of the manuscript.
Ethics approval and consent to
participate
Informed consent was obtained from the volunteers.
The present study was approved by the ethics committees of the
Capital Medical University Affiliated Beijing Friendship
Hospital.
Patient consent for publication
Informed consent was obtained from all
participants.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar
|
|
2
|
He YT, Li DJ, Liang D, Jin J, Wen DG, Chen
WQ and He J: Estimated of esophageal cancer incidence and mortality
in China, 2013. Zhonghua Zhong Liu Za Zhi. 39:315–320. 2017.In
Chinese. PubMed/NCBI
|
|
3
|
Enzinger PC and Mayer RJ: Esophageal
cancer. N Engl J Med. 349:2241–2252. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li JY: Epidemiology of esophageal cancer
in China. Natl Cancer Inst Monogr. 62:113–120. 1982.PubMed/NCBI
|
|
5
|
Liu X, Zhang M, Ying S, Zhang C, Lin R,
Zheng J, Zhang G, Tian D, Guo Y, Du C, et al: Genetic Alterations
in Esophageal Tissues From Squamous Dysplasia to Carcinoma.
Gastroenterology. 153:166–177. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chan GK, Jablonski SA, Starr DA, Goldberg
ML and Yen TJ: Human Zw10 and ROD are mitotic checkpoint proteins
that bind to kinetochores. Nat Cell Biol. 2:944–947. 2000.
View Article : Google Scholar
|
|
7
|
Keen N and Taylor S: Aurora-kinase
inhibitors as anticancer agents. Nat Rev Cancer. 4:927–936. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen Y, Riley DJ, Chen PL and Lee WH: HEC,
a novel nuclear protein rich in leucine heptad repeats specifically
involved in mitosis. Mol Cell Biol. 17:6049–6056. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kaneko N, Miura K, Gu Z, Karasawa H,
Ohnuma S, Sasaki H, Tsukamoto N, Yokoyama S, Yamamura A, Nagase H,
et al: siRNA-mediated knockdown against CDCA1 and KNTC2, both
frequently overexpressed in colorectal and gastric cancers,
suppresses cell proliferation and induces apoptosis. Biochem
Biophys Res Commun. 390:1235–1240. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qu Y, Li J, Cai Q and Liu B: Hec1/Ndc80 is
overexpressed in human gastric cancer and regulates cell growth. J
Gastroenterol. 49:408–418. 2014. View Article : Google Scholar
|
|
11
|
Bièche I, Vacher S, Lallemand F,
Tozlu-Kara S, Bennani H, Beuzelin M, Driouch K, Rouleau E,
Lerebours F, Ripoche H, et al: Expression analysis of mitotic
spindle checkpoint genes in breast carcinoma: Role of NDC80/HEC1 in
early breast tumorigenicity, and a two-gene signature for
aneuploidy. Mol Cancer. 10:232011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hayama S, Daigo Y, Kato T, Ishikawa N,
Yamabuki T, Miyamoto M, Ito T, Tsuchiya E, Kondo S and Nakamura Y:
Activation of CDCA1-KNTC2, members of centromere protein complex,
involved in pulmonary carcinogenesis. Cancer Res. 66:10339–10348.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Meng QC, Wang HC, Song ZL, Shan ZZ, Yuan
Z, Zheng Q and Huang XY: Overexpression of NDC80 is correlated with
prognosis of pancreatic cancer and regulates cell proliferation. Am
J Cancer Res. 5:1730–1740. 2015.PubMed/NCBI
|
|
14
|
Huang LY, Chang CC, Lee YS, Huang JJ,
Chuang SH, Chang JM, Kao KJ, Lau GM, Tsai PY, Liu CW, et al:
Inhibition of Hec1 as a novel approach for treatment of primary
liver cancer. Cancer Chemother Pharmacol. 74:511–520. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Makita Y, Murata S, Katou Y, Kikuchi K,
Uejima H, Teratani M, Hoashi Y, Kenjo E, Matsumoto S, Nogami M, et
al: Anti-tumor activity of KNTC2 siRNA in orthotopic tumor model
mice of hepatocellular carcinoma. Biochem Biophys Res Commun.
493:800–806. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Urata YN, Takeshita F, Tanaka H, Ochiya T
and Takimoto M: Targeted Knockdown of the Kinetochore Protein
D40/Knl-1 Inhibits Human Cancer in a p53 Status-Independent Manner.
Sci Rep. 5:136762015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
18
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou Y, Su Z, Huang Y, Sun T, Chen S, Wu
T, Chen G, Xie X, Li B and Du Z: The Zfx gene is expressed in human
gliomas and is important in the proliferation and apoptosis of the
human malignant glioma cell line U251. J Exp Clin Cancer Res.
30:1142011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Abe W, Nasu K, Nakada C, Kawano Y,
Moriyama M and Narahara H: miR-196b targets c-myc and Bcl-2
expression, inhibits proliferation and induces apoptosis in
endometriotic stromal cells. Hum Reprod. 28:750–761. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liberzon A, Birger C, Thorvaldsdóttir H,
Ghandi M, Mesirov JP and Tamayo P: The Molecular Signatures
Database (MSigDB) hallmark gene set collection. Cell Syst.
1:417–425. 2015. View Article : Google Scholar
|
|
22
|
Du J, Yuan Z, Ma Z, Song J, Xie X and Chen
Y: KEGG-PATH: Kyoto encyclopedia of genes and genomes-based pathway
analysis using a path analysis model. Mol Biosyst. 10:2441–2447.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cleveland DW, Mao Y and Sullivan KF:
Centromeres and kinetochores: From epigenetics to mitotic
checkpoint signaling. Cell. 112:407–421. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Orr B, Godek KM and Compton D: Aneuploidy.
Curr Biol. 25:R538–R542. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hartwell LH and Kastan MB: Cell cycle
control and cancer. Science. 266:1821–1828. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Beh TT and Kalitsis P: The Role of
Centromere Defects in Cancer. Prog Mol Subcell Biol. 56:541–554.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Thompson SL and Compton DA: Chromosomes
and cancer cells. Chromosome Res. 19:433–444. 2011. View Article : Google Scholar
|
|
28
|
Thiru P, Kern DM, McKinley KL, Monda JK,
Rago F, Su KC, Tsinman T, Yarar D, Bell GW and Cheeseman IM:
Kinetochore genes are coordinately up-regulated in human tumors as
part of a FoxM1-related cell division program. Mol Biol Cell.
25:1983–1994. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Scaërou F, Starr DA, Piano F, Papoulas O,
Karess RE and Goldberg ML: The ZW10 and Rough Deal checkpoint
proteins function together in a large, evolutionarily conserved
complex targeted to the kinetochore. J Cell Sci. 114:3103–3114.
2001.PubMed/NCBI
|
|
30
|
Wilson WR and Hay MP: Targeting hypoxia in
cancer therapy. Nat Rev Cancer. 11:393–410. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li Y, Fu L, Li JB, Qin Y, Zeng TT, Zhou J,
Zeng ZL, Chen J, Cao TT, Ban X, et al: Increased expression of
EIF5A2, via hypoxia or gene amplification, contributes to
metastasis and angiogenesis of esophageal squamous cell carcinoma.
Gastroenterology. 146:1701–1713.e9. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sulli G, Di Micco R and d'Adda di Fagagna
F: Crosstalk between chromatin state and DNA damage response in
cellular senescence and cancer. Nat Rev Cancer. 12:709–720. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Olcina MM, Foskolou IP, Anbalagan S, Senra
JM, Pires IM, Jiang Y, Ryan AJ and Hammond EM: Replication stress
and chromatin context link ATM activation to a role in DNA
replication. Mol Cell. 52:758–766. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hammond EM, Dorie MJ and Giaccia AJ:
ATR/ATM targets are phosphorylated by ATR in response to hypoxia
and ATM in response to reoxygenation. J Biol Chem. 278:12207–12213.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu D, Keijzers G and Rasmussen LJ: DNA
mismatch repair and its many roles in eukaryotic cells. Mutat Res.
773:174–187. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ling ZQ, Li P, Ge MH, Hu FJ, Fang XH, Dong
ZM and Mao WM: Aberrant methylation of different DNA repair genes
demonstrates distinct prognostic value for esophageal cancer. Dig
Dis Sci. 56:2992–3004. 2011. View Article : Google Scholar : PubMed/NCBI
|