Introduction
Lung cancer is the leading cause of cancer-related
mortality worldwide (1). Non-small
cell lung cancer (NSCLC) accounts for ~85% of lung cancer cases,
with adenocarcinoma being the most common subtype, followed by
squamous cell carcinoma (2). The
majority of patients with NSCLC are not diagnosed within the
optimal time frame and thus cannot undergo successful radical
resection due to tumor invasion and distant metastasis (3). The identification of novel potential
therapeutic targets which may be used to prevent the invasion of
lung adenocarcinoma is urgently required.
Tumor growth and metastasis are closely associated
with the properties of tumor cells, as well as with the tumor
stroma (4,5). Fibroblasts in the tumor stroma are
permanently activated and serve as important precursors of tumor
growth, invasion and metastasis (6-9).
Cancer-associated fibroblasts (CAFs) express α-smooth muscle actin
(α-SMA) and vimentin (VIM), and exhibit a high degree of
heterogeneity (4,5,10).
However, cytokeratin (CK) derived from epithelium-originated cancer
is not expressed in CAFs (11).
CAFs promote tumor progression by secreting chemokines, and other
factors in various types of cancer (4,12-14).
A recent study suggested that CAFs enhance the invasive ability of
lung adenocarcinoma cells (15).
However, the underlying mechanisms remain unclear.
Metastasis-associated in colon cancer-1 (MACC1) was
originally identified in primary and metastatic colon carcinomas
(16). MACC1 expression has been
demonstrated to be upregulated in different types of cancer
(17). It has also been indicated
that MACC1 is involved in tumor invasion and metastasis in gastric
cancer (18), breast cancer
(19) and hepatocellular cancer
(20,21). It was previously shown that MACC1
was overexpressed in lung adenocarcinoma tissue, and was associated
with the rate of tumor metastasis and invasion (22,23).
However, the role of MACC1 in CAFs, particularly as regards the
promoting effects on cancer cell invasive ability, remains poorly
characterized. Therefore, in the present study, the effects and
potential mechanisms of action of MACC1 as regards tumor invasion
in lung adenocarcinoma were investigated.
Materials and methods
Cell lines
The human lung cancer cell line (A549), the normal
human fetal lung fibroblast cell line (HFL-1) and human embryonic
kidney 293T cell line were purchased from the American Type Culture
Collection (ATCC, Manassas, VA, USA).
Patients and tissue specimens
The present study was approved by the Ethics
Committee of the First Affiliated Hospital of Dalian Medical
University (Ethics no. YJ-KY-FB-20). Tissue samples were collected
from 7 patients with lung adenocarcinoma who underwent radical
surgery of the primary tumor and systematic nodal dissection
without any adjuvant therapy at the Department of Thoracic Surgery
of the First Affiliated Hospital of Dalian Medical University from
January, 2017 to March, 2017. All the patients had signed an
informed consent form. The included patients (Table I) were aged between 58 to 76 years
(median age, 64 years).
 | Table ICharacteristics of the patients with
pancreatic adenocarcinoma. |
Table I
Characteristics of the patients with
pancreatic adenocarcinoma.
| Patient no. | Age (years) | Sex | Metastasis
status | Tumor stage | Grade | Pathological
subtype |
|---|
| 1 (CAF A) | 58 | F | – | T1N0M0 | 1 | Adenocarcinoma |
| 2 (CAF B) | 59 | F | – | T1N0M0 | 1 | Adenocarcinoma |
| 3 (CAF 1) | 65 | M | – | T1N0M0 | 1 | Adenocarcinoma |
| 4 (CAF 2) | 63 | M | – | T1N0M0 | 1 | Adenocarcinoma |
| 5 (CAF 3) | 59 | F | – | T1N0M0 | 1 | Adenocarcinoma |
| 6 (CAF 4) | 68 | M | – | T1N0M0 | 1 | Adenocarcinoma |
| 7 (CAF 5) | 76 | M | – | T1N0M0 | 1 | Adenocarcinoma |
Primary cell culture
The fresh specimens sectioned into a smooth paste
were digested with 5 mg/ml collagenase I and 0.2 mg/ml hyaluronic
acid dissolved into Dulbecco’s modified Eagle’s medium (DMEM;
Thermo Fisher Scientific, Waltham, MA, USA) plus 10% fetal bovine
serum (FBS; Thermo Fisher Scientific) in a 37°C incubator for 5 h.
Tissue debris was removed by a filter and cells were collected and
cultured in a 37°C humidified atmosphere containing 95% air and 5%
CO2. Once the cells reached 80-90% confluence, they were
harvested and reseeded in complete medium (DMEM plus 10% fetal
bovine serum).
Immunohistochemical staining (IHC)
The lung tissue specimens were first formalin-fixed
and paraffin-embedded, rehydrated and incubated in 3% (v/v)
hydrogen peroxide (Sigma-Aldrich, St. Louis, MO, USA) for 10 min.
Antigenic retrieval was processed with sodium citrate. IHC staining
was performed using the Streptavidin-Peroxidase IHC assay kit
(ZSGB-Bio, Beijing, China) following the manufacturer’s
instructions. The antibody to α-SMA (rabbit; cat. no. ab5694;
Abcam, Cambridge, MA, USA) was diluted 1:200 proportionally in PBS
containing 2% goat bovine serum. Immunostaining was evaluated by
two pulmonary pathologists using a blinded protocol design.
Immunofluorescence
The grown CAFs isolated from the tumor tissues were
seeded on coverslips within 6-well plates. were fixed with 4%
paraformaldehyde, permeabilized with 0.2% Triton X-100, and then
blocked with 10% bovine serum albumin. The primary antibodies
against pan-ck (mouse; ZSGB-Bio; diluted 1:100), α-SMA (rabbit;
cat. no. ab5694; diluted 1:100; Abcam) and Vimentin (rabbit; cat.
no. D21H3; diluted 1:200; Cell Signaling Technology, Danvers, MA,
USA) and diluted in blocking solution. Immune complexes were
stained with the secondary antibody conjugated to Alexa-488 (cat.
no. A-11008; diluted 1:100; Thermo Fisher Scientific) or Alexa-546
(cat. no. A10040; diluted 1:100; Thermo Fisher Scientific). The
nuclei were stained with DAPI at 25°C for 4 min (Sigma-Aldrich).
Each experimental group contained 6 wells each time, and 5 fields
of each well were imaged using a fluorescence microscope (Olympus
IX71; Olympus Corporation, Tokyo, Japan).
Western blot analysis
The cells were collected at the optimum cell culture
time in a microcentrifuge tube. Total protein was extracted using
RIPA lysing buffer, and lysates were centrifuged at 12,000 x g for
10 min at 4°C. The concentration of the total, nuclei or cytoplasm
cell protein lysate was quantified using a BCA protein assay kit
(Thermo Fisher Scientific). Subsequently, 30-50 μg of
protein lysates were separated by 10% SDS-PAGE and transferred onto
a polyvinylidene difluoride (PVDF) membrane. The membrane was
incubated with the specific antibodies (appropriate concentration
referred to specification) and protein bands were detected with an
enhanced chemiluminescence system (MilliporeSigma, Burlington, MA,
USA). The following antibodies were used: Anti-Vimentin (rabbit;
cat. no. D21H3; diluted 1:1,000; Cell Signaling Technology),
anti-GAPDH (mouse; diluted 1:5,000; cat. no. SAB1405848;
Sigma-Aldrich), anti-MACC1 (rabbit; diluted 1:1,000; cat. no.
ab106579; Abcam), anti-hepatocyte growth factor (HGF; diluted
1:1,000; rabbit; cat. no. ab83760; Abcam), anti-leukemia inhibitory
factor (LIF; diluted 1:1,000; rabbit; cat. no. 26757-1-AP;
Proteintech, Rosemont, IL, USA), anti-CXCL12 (rabbit; diluted
1:1,000; cat. no. 17402-1-AP; Proteintech), anti-insulin-like
growth factor 2 (IGF2; diluted 1:1,000; mouse; cat. no. SAB1406010;
Sigma-Aldrich), goat-antirabbit IgG conjugated to horseradish
peroxidase (HRP; cat. no. 31460; Thermo Fisher Scientific) and
goat-anti-mouse IgG conjugated to HRP (cat. no. 31430; Thermo
Fisher Scientific) which was used as the secondary antibody. The
densitometry of the blots was calculated using Image lab 3.5 and
the bar graph was generated using the Graphpad Prism6.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the cells using TRIzol
reagent (Invitrogen/Thermo Fisher Scientific), which was used to
generate cDNA using the PrimeScript RT reagent kit (Takara Bio USA,
Inc., Mountain View, CA, USA). The resulting cDNA was analyzed in
triplicate using reaction 2 steps qPCR (95°C for 30 sec, followed
by 40 cycles of 95°C for 10 sec, annealing at 60°C for 30 sec and
followed by 1 cycle of 95°C for 15 sec, at 60°C for 1 min and 95°C
for 15 sec) with SYBR Select Master Mix (Life Technologies/Thermo
Fisher Scientific). The qPCR sequences were listed as follows: ACTB
sense, 5′-CATGTACGTTGCTATCCAGGC-3′ and antisense,
5′-CTCCTTAATGTCACGCACGAT-3′); MACC1 sense,
5′-TTCTTTTGATTCCTCCGGTGA-3′ and antisense, 5′-ACT
CTGATGGGCATGTGCTG-3′; HGF sense, 5′-TGCACG ACAGTGTTTCCCTT-3′ and
antisense, 5′-CACATCCAC GACCAGGAACA-3′; LIF sense, 5′-GTCTTGGCGGCA
GTACACAG-3′ and antisense, 5′-CGACTATGCGGTACA GCTCC-3′; CXCL12
sense, 5′-CAGATGCCCATGCCGATT CT-3′ and antisense,
5′-TTCTTCAGCCGGGCTACAAT-3′; IGF2 sense, 5′-AACTTCCCCAGATACCCCGT-3′
and antisense, 5′-GGGTGGGTAGAGCAATCAGG-3′.
Plasmid constructs
The plasmid encoding human MACC1 was generated by
PCR amplification and inserted into the pLVX-DsRed-N1-Monomer
vector (Clontech/Takara Bio USA, Inc.). The primers for gene
cloning were as follows: MACC1 5′XhoI,
5′-CACTCGAGGCCACCATGCTAATC ACTGAAAGAAAACATTTTCGG-3′; 3′XmaI,
5′-CACCCG GGCTACTTCCTCAGAAGTGGAGAATGCAGTTACTC-3′.
Lentivirus preparation and
transfection
The targeting MACC1 and the non-target shRNA
(shMACC1 and shNC) were purchased from GenePharma (Shanghai,
China). The target siRNA sequences of MACC1 were as follows: 5′-GUG
ACGAGGAAUUGACAAUTT-3′, and that of the negative control siRNA was
5′-UUCUCCGAACGUGUCACGUTT-3′). 293T cells were co-transfected with
the psPAX2 and pMD2.G using Lipofectamine 2000 (Life
Technologies/Thermo Fisher Scientific). Lentiviruses were harvested
and concentrated by ultracentrifugation (1,500 rpm at room
temperature for 5 min) at 48 h following transfection, and viral
titer determined by serial dilutions. Infected cells (A549 and
CAFs) were selected with puromycin (2 mg/ml; Sigma-Aldrich).
Cell invasion assay based on the
microfluidic model
To estimate the invasive potential of the CAFs, a
microfluidic device developed in our previous study was used
(24). Matrigel, as the substitute
for ECM, was loaded into the middle channel of the microfluidic
device. The cells (A549, CAF#I, CAF#II, CAF-2, CAF-3, CAF-2-VECTOR,
CAF-2-MACC1-OE, CAF-shNC and CAF-3-shMACC1) were seeded into the
upper channel in serum-free medium for cell culture and medium with
10% FBS was loaded into the under channel for stimulation. The
device was incubated for 48 h at 37°C. Cell invasion area was
determined as the area invaded by cells in the matrix channel and
measured by Image-Pro Plus6.0. For observation, A549 with green
fluorescence by infected by pEZ-vector (GeneCopoeia USA, Inc.) and
CAF#I CAF#II were marked as CAF-A and CAF-B after
immunofluorescence assay by infected pLVX-DsRed-Nl-Monomer vector
(Clontech/Takara Bio USA, Inc.).
Statistical analysis
The Student’s t-test and analysis of variance
(one-way ANOVA) followed by appropriate post hoc tests (Fishers’
Least Significant Difference) were used to compare the values of
the test and control samples in vitro. Values of P<0.05 were
considered to indicate statistically significant differences.
Results
Highly active CAFs are detected in
patients with lung adenocarcinoma
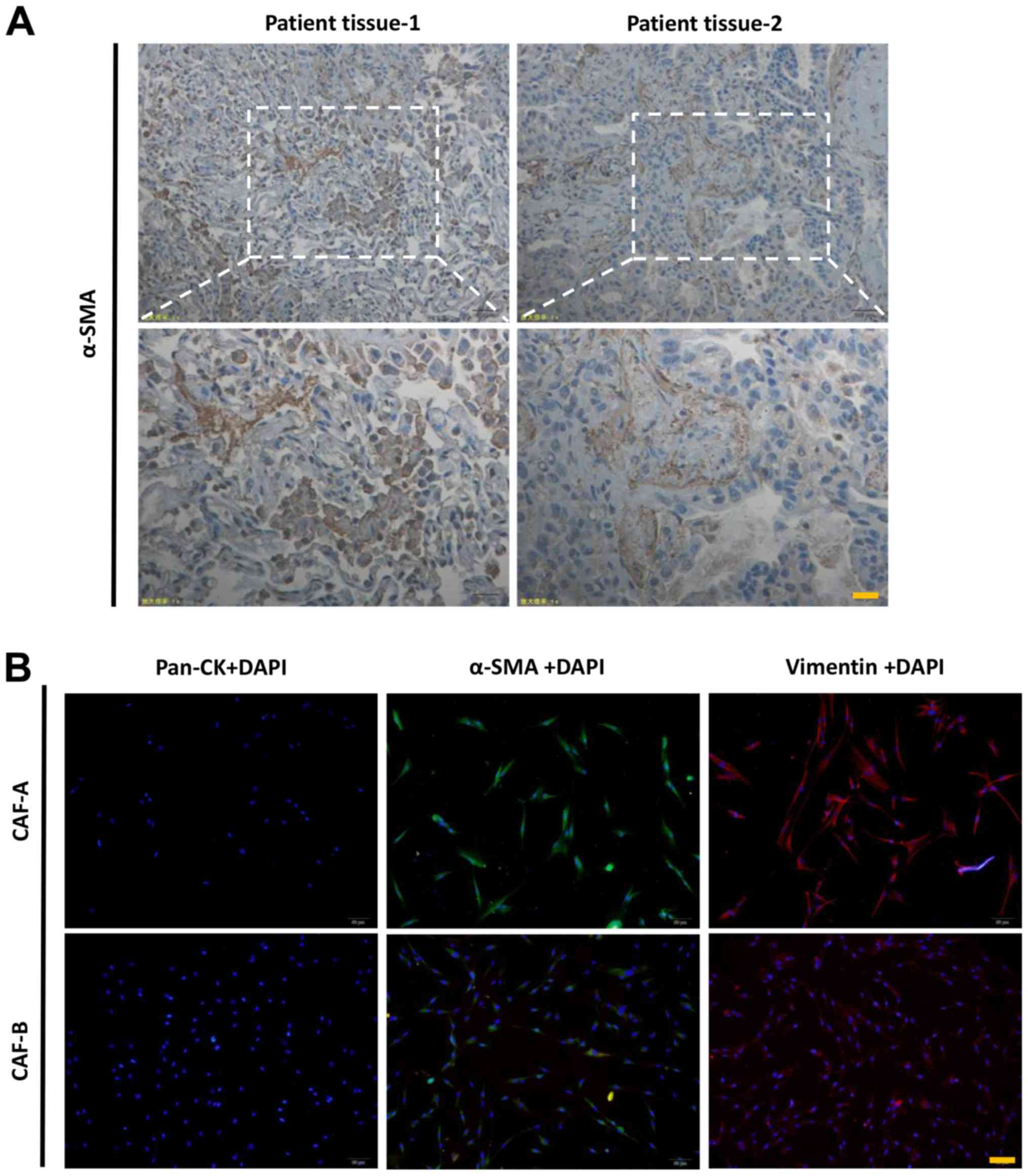
A number of α-SMA-positive cells were observed in
the pulmonary mesenchyme by immunohistochemical staining (Fig. 1A), indicating the CAFs in two lung
adenocarcinoma cases. CAF-A and CAF-B were extracted from the two
lung adenocarcinoma patient tissues, and both cell types exhibited
a typical fibroblastic morphology with cytoplasmic processes on a
monolayer. They were negative for pan-ck and positive for α-SMA and
Vimentin. The activated status of the CAFs were detected in the
primary cells from the patients with lung adenocarcinoma (Fig. 1B).
CAFs promote the invasion of lung
adenocarcinoma cells
Accumulating evidence suggests that the migration
and invasion process of tumor cells may be regulated by the
function of CAFs (5). In this
study, to confirm whether CAFs can promote lung adenocarcinoma
cells lateral invasion process beyond co-culture condition, a
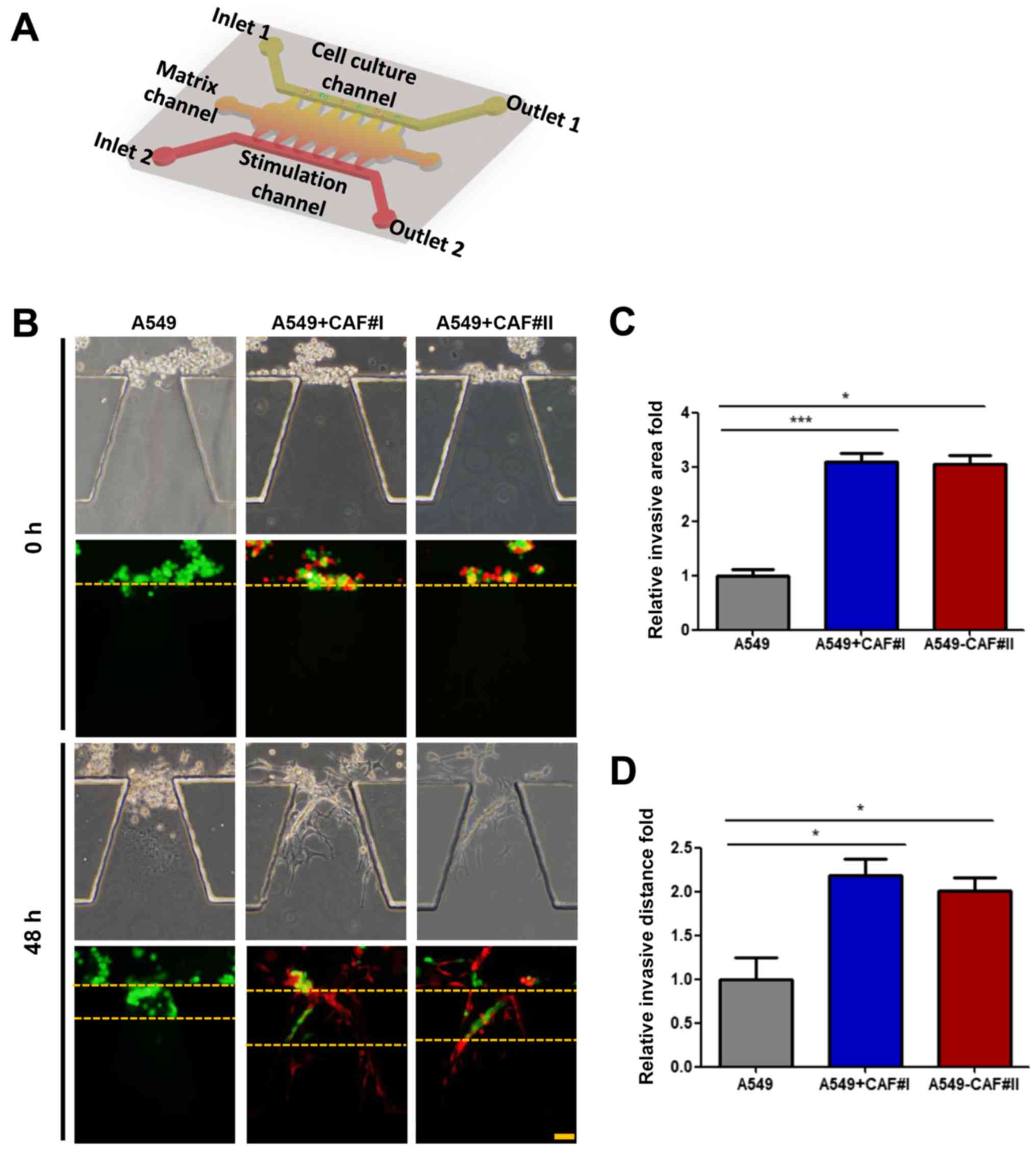
microfluidic model was employed, as previously described (24). The cell culture, stimulation
channel and matrix channels were independent. The device,
consisting of a glass slide and translucent PDMS, was sterilized
and bonded to a 10-cm dish prior to use (Fig. 2A). The A549 cells were cultured
alone as a standard control, or co-cultured with CAF#I or CAF#II in
a microfluidic chip (Fig. 2B).
Matrigel and medium containing 10% serum was loaded into the matrix
channels prior to addition of the separate groups into the cell
culture channel into the cell culture channel, and cultured in
serum-free medium for 48 h. The invasive distance and area were
increased in the co-culture condition, compared with the A549 cells
cultured alone (Fig. 2C and
D).
MACC1 promotes the invasive ability of
CAFs
MACC1 has been demonstrated to promote the
metastasis and invasion of lung adenocarcinoma cells (25). In this study, to investigate the
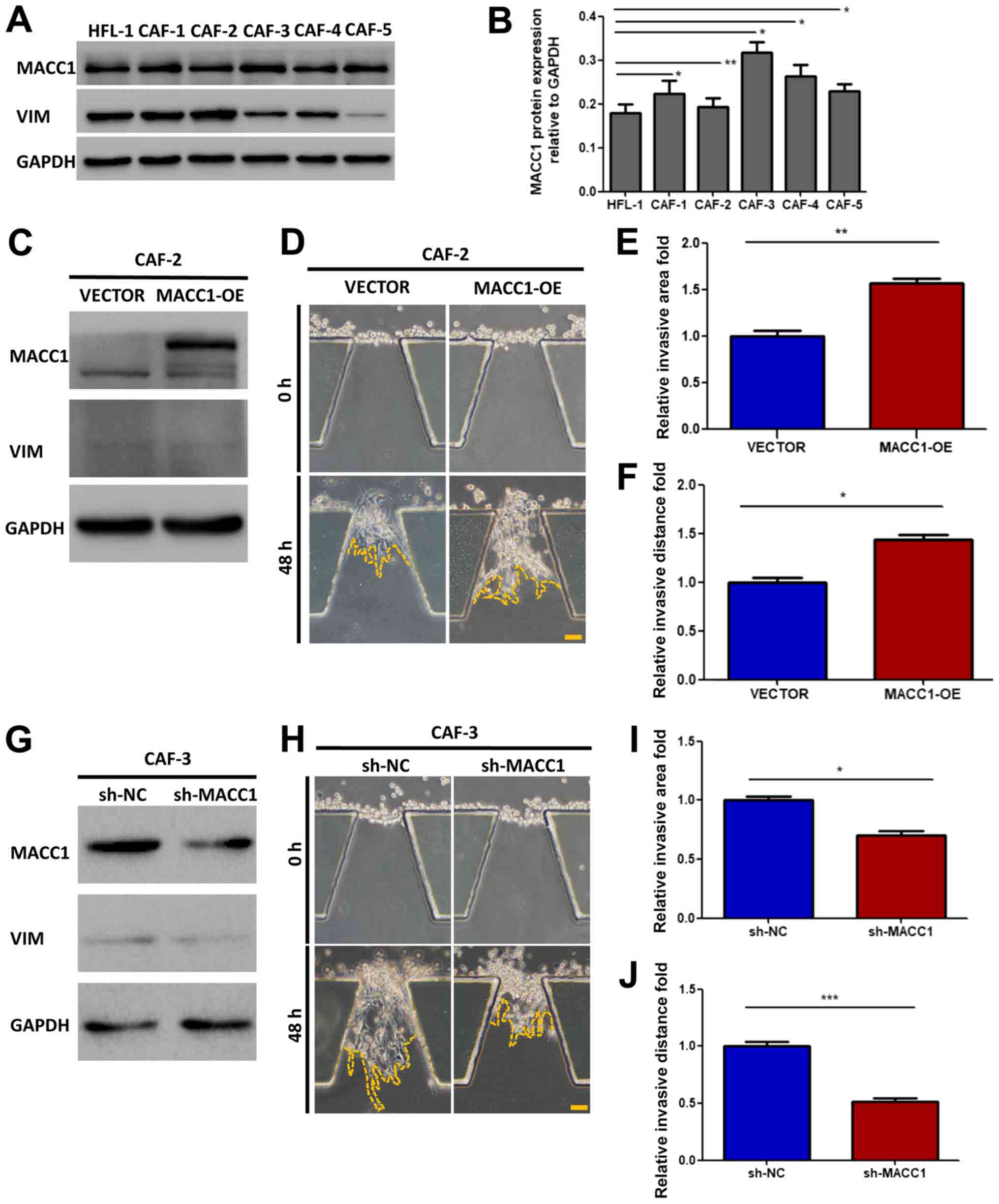
role of MACC1 in the invasive process of CAFs, we detected the
protein expression level of MACC1 in CAFs isolated from 5 patients
with lung adenocarcinoma, and compared the result with normal
fibroblast cells (HFL-1) (Fig. 3A and
B). MACC1 was expressed in all the CAFs, and the cells
exhibiting the lowest and highest expression were selected for use
in further experiments: CAF-2 and CAF-3, respectively.
Subsequently, the overexpression of MACC1 was induced in the CAF-2
cells (Fig. 3C) and it was knocked
down in the CAF-3 cells (Fig. 3G).
The stably transfected cells (CAF-2-VECTOR, CAF-2-MACC1 or
CAF-3-shNC, CAF-3 - shMACC 1) were injected into microfluidic
chips. After 48 h, the cell invasive ability was examined and it
was found to be enhanced in the CAF-2 cells (Fig. 3D), as indicated by the increase in
both the invasive distance and area, compared with the controls
(Fig. 3E and F). By contrast, the
invasive ability was markedly reduced in the CAF-3 cells in which
MACC1 was knocked down (Fig. 3H),
as indicated by a decrease in both invasive the distance and area
(Fig. 3I and J).
Expression of MACC1 by CAFs stimulates
the invasion of lung adenocarcinoma cells
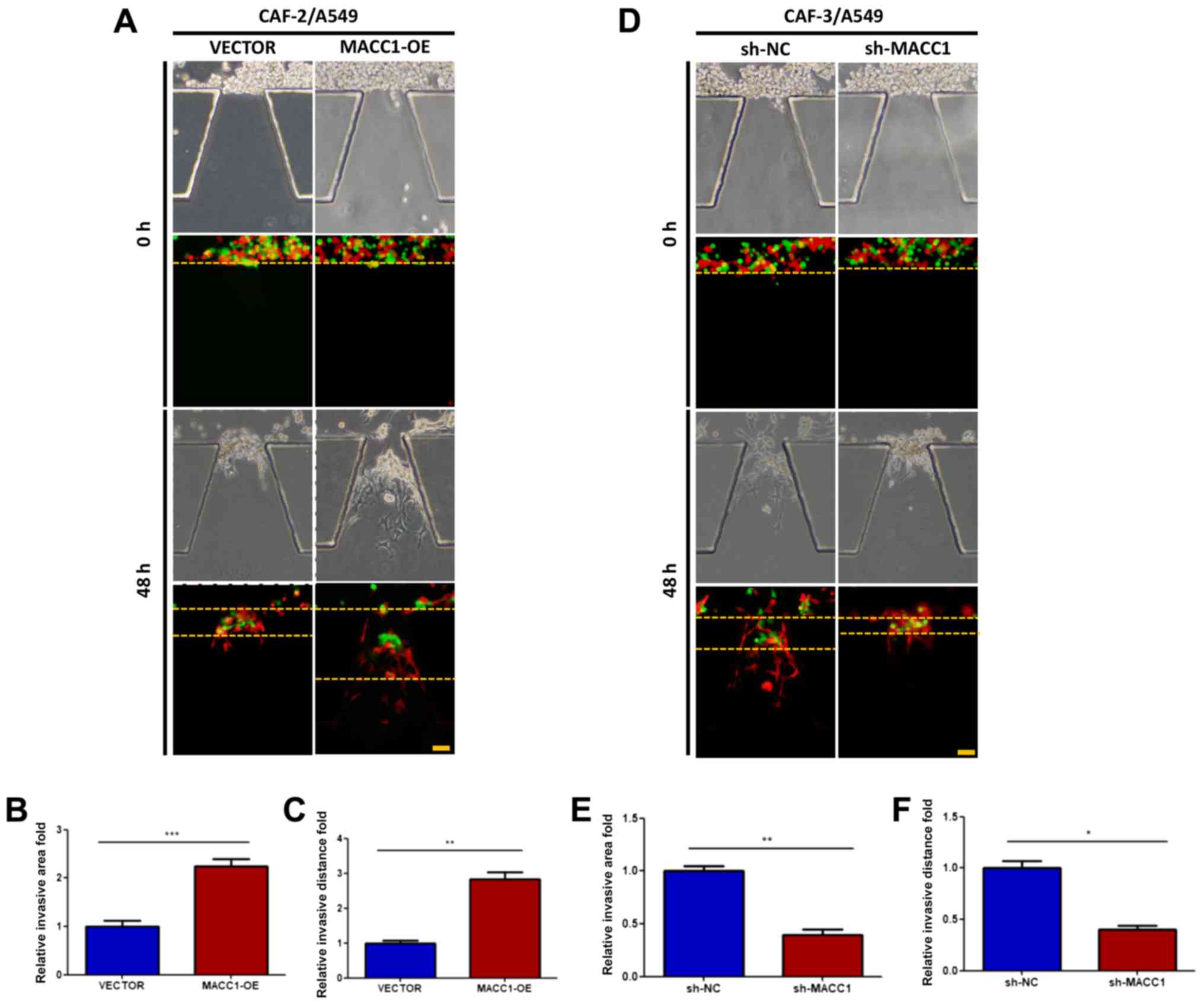
As suggested by the above-mentioned results, MACC1
promotes the invasive ability of the CAFs. We investigated whether
interfering with MACC1 expression in CAFs can influence the
invasive activity of lung adenocarcinoma cells in co-culture
conditions. Stably transfected cell lines, CAF-2-VECTOR,
CAF-2-MACC1, CAF-3-shNC, CAF-3-shMACC1 were injected with A549
cells into microfluidic chips. After 48 h, the invasive ability of
the A549 cells was enhanced by co-culture with CAF-2-MACC1 cells
(Fig. 4A-C). However, the invasive
ability of the A549 cells was impaired by co-culture with
CAF-3-shMACC1 cells (Fig. 4D), as
indicated by the changes in the invasive distance and area
(Fig. 4E and F).
Overexpression of MACC1 induces CAFs to
secrete chemokines through paracrine signaling
Stromal-derived IGF2, HGF, LIF and CXCL12 have been
demonstrated to promote cancer progression and invasion via
paracrine signaling (4,12-14).
In this study, we aimed to explore the mechanisms responsible for
the enhancement of lung adenocarcinoma cell motility by MACC1, by
co-culture with CAFs. In stable MACC1-overexpressing CAF-2 cells,
the expression levels of CXCL12, LIF (P<0.05) and HGF
(P<0.001) were increased at the transcriptional and proteinic
level compared with the CAF-2-VECTOR cells (Fig. 5A and B). In stable CAF-3-shMACC1
cells, the expression levels of HGF, LIF (P<0.05) and CXCL12
(P<0.01) were decreased at the transcriptional and protein level
compared with the CAF-3-shNC cells (Fig. 5C and D). It can thus be
hypothesized that the overexpression of MACC1 may stimulate CAFs to
produce and secrete these chemokines via paracrine signaling, thus
promoting lung adenocarcinoma cell invasion.
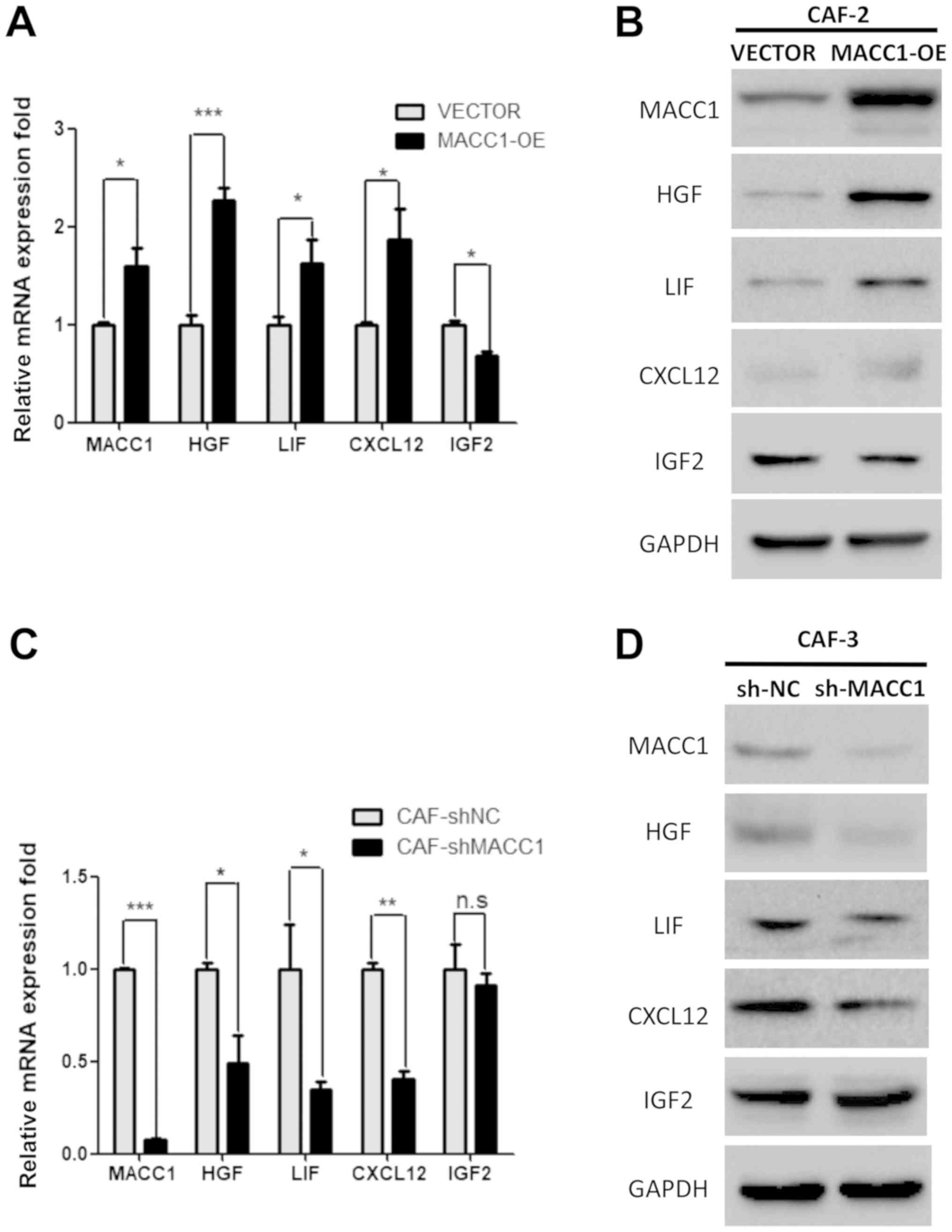 | Figure 5Overexpression of MACC1 induces CAFs
to secrete chemokines through paracrine signaling. RT-qPCR analysis
revealed the expression levels of HFG, LIF, CXCL12 and IGF2 in (A)
MACC1-overexpressing CAF-2 cells and (B) shMACC1 CAF-3 cells.
Western blot analysis revealed the expression level of HFG, LIF,
CXCL12 and IGF2 in (C) MACC1-overexpressing CAF-2 cells and (D)
shMACC1 CAF-3. Error bars represent the means ± SD derived from 3
independent experiments. *P<0.05, **P<0.01 and ***P<0.005.
MACC1, metastasis-associated in colon cancer-1; CAFs,
carcinoma-associated fibroblasts; HFG, hepatocyte growth factor;
LIF, leukemia inhibitory factor; IGF2, insulin-like growth factor
2. |
Discussion
The present study indicates a previously
unrecognized role of MACC1 in CAFs, namely the promotion of the
invasive ability of lung adenocarcinoma cells based on the
following findings: i) CAFs existed in lung adenocarcinoma tissue;
ii) CAFs induced the invasion of lung adenocarcinoma cells; iii)
MACC1 overexpression promoted the invasive ability of CAFs; and iv)
MACC1 overexpression in CAFs stimulated the invasion of lung
adenocarcinoma cells, and MACC1 overexpression induced CAFs to
secrete chemokines through paracrine signaling.
Previous researchhas identified that the invasion
andmetastasis of various types of tumor are induced by CAFs,
including lung adenocarcinoma (15,26).
It has become increasingly clear that CAFs promote tumor
progression (27). Activated
fibroblasts induce epithelial cells, causing tumor invasion and
metastasis, this effect is termed ‘collective invasion’ (28,29).
In this study, we constructed a microfluidic device to confirm that
CAFs promoted the invasive ability of A549 cells (lung
adenocarcinoma) in the 3D co-culture conditions.
It has been reported that MACC1 promotes the
invasion of various types of cancer cells, proliferation and
migration in cell culture and distant metastasis in mouse models
(16,30,31).
The overexpression of MACC1 has been associated with a poor
prognosis in various types of cancer (32-34).
The inhibition of MACC1 expression has been demonstrated to repress
tumor cell invasion and migration (35). Recently, we found that the
activated MACC1/c-Met pathway, regulated by YB-1, promoted tumor
proliferation and invasion in lung adenocarcinoma (36). CAFs and MACC1 both function in
tumor progression, and we hypothesized that MACC1 may modulated CAF
activity. In this study, we demonstrated that the local invasive
capacity of CAFs was enhanced by the overexpression MACC1.
Furthermore, the overexpression of MACC1 in CAFs increased the area
and distance of lung adenocarcinoma cell invasion. MACC1 knockdown
resulted in the reduced ability of CAFs to induce lung
adenocarcinoma invasion.
Some researchers have demonstrated that CAFs can
secrete extracellular matrix components, growth factors and
chemokines which promote tumor invasion and metastasis (4,37).
The crosstalk between tumor cells and stroma CAFs has also been
indicated to contribute to tumor progression via HGF signaling
(38). MACC1 was previously
reported to regulate invasion and metastasis through regulating the
HGF/c-Met pathway (16). Of note,
in this study, we found that HGF, among other chemokines, was
upregulated in MACC1-overexpressing CAFs. We thus hypothesized that
MACC1 enhances paracrine signaling in CAFs and promotes lung
adenocarcinoma cell invasion. However, further studies are required
to focus on the effect of paracrine signaling inhibition on
crosstalk between CAFs and lung adenocarcinoma.
In conclusion, in this study, we demonstrated that
the overexpression of MACC1 in CAFs promoted the invasive ability
of lung adenocarcinoma cells via paracrine signaling. The targeting
of MACC1 may thus prove to be a potential therapeutic strategy for
overcoming the CAF-induced invasion in lung adenocarcinoma.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (nos. 81173453 and 81774078),
and the Natural Science Foundation of Liaoning Province, China
(nos. 201602227 and 20170540300).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors’ contributions
TG, ZL, JL and CG conceived and designed the study.
TG and ZL were involved in research administration. ZL, TG, LF, XW,
NL, PW, SZ, FL, YC, XS and LZ performed the experiments and
analyzed the data. TG, ZL, JL and CG wrote and edited the
manuscript. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
All experiments using human samples were approved by
the Medical Ethical Committees of the First Affiliated Hospital of
Dalian ethics committee Medical University (Ethics no.
YJ-KY-FB-20). All patients signed written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors are thankful to Miss Qianqian Bi, Miss
Mengying Yang, Mr. Zhe Sun, Mr. Qi Yang and Mr. Chao Li from Dalian
Medical University for preparing the pathological specimens.
References
|
1
|
Allemani C, Weir HK, Carreira H, Harewood
R, Spika D, Wang XS, Bannon F, Ahn JV, Johnson CJ, Bonaventure A,
et al CONCORD Working Group: Global surveillance of cancer survival
1995-2009: Analysis of individual data for 25,676,887 patients from
279 population-based registries in 67 countries (CONCORD-2).
Lancet. 385:977–1010. 2015. View Article : Google Scholar
|
|
2
|
Langer CJ, Besse B, Gualberto A, Brambilla
E and Soria JC: The evolving role of histology in the management of
advanced non-small-cell lung cancer. J Clin Oncol. 28:5311–5320.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Perlikos F, Harrington KJ and Syrigos KN:
Key molecular mechanisms in lung cancer invasion and metastasis: A
comprehensive review. Crit Rev Oncol Hematol. 87:1–11. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kalluri R and Zeisberg M: Fibroblasts in
cancer. Nat Rev Cancer. 6:392–401. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gupta GP and Massagué J: Cancer
metastasis: Building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shekhar MP, Pauley R and Heppner G: Host
microenvironment in breast cancer development: Extracellular
matrix-stromal cell contribution to neoplastic phenotype of
epithelial cells in the breast. Breast Cancer Res. 5:130–135. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Olaso E, Salado C, Egilegor E, Gutierrez
V, Santisteban A, Sancho-Bru P, Friedman SL and Vidal-Vanaclocha F:
Proangiogenic role of tumor-activated hepatic stellate cells in
experimental melanoma metastasis. Hepatology. 37:674–685. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Orimo A, Gupta PB, Sgroi DC,
Arenzana-Seisdedos F, Delaunay T, Naeem R, Carey VJ, Richardson AL
and Weinberg RA: Stromal fibroblasts present in invasive human
breast carcinomas promote tumor growth and angiogenesis through
elevated SDF-1/CXCL12 secretion. Cell. 121:335–348. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
De Wever O and Mareel M: Role of tissue
stroma in cancer cell invasion. J Pathol. 200:429–447. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ostman A and Augsten M: Cancer-associated
fibroblasts and tumor growth–bystanders turning into key players.
Curr Opin Genet Dev. 19:67–73. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xing F, Saidou J and Watabe K: Cancer
associated fibroblasts (CAFs) in tumor microenvironment. Front
Biosci. 15:166–179. 2010. View
Article : Google Scholar :
|
|
12
|
Unger C, Kramer N, Unterleuthner D,
Scherzer M, Burian A, Rudisch A, Stadler M, Schlederer M, Lenhardt
D, Riedl A, et al: Stromal-derived IGF2 promotes colon cancer
progression via paracrine and autocrine mechanisms. Oncogene.
36:5341–5355. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Teng F, Tian WY, Wang YM, Zhang YF, Guo F,
Zhao J, Gao C and Xue FX: Cancer-associated fibroblasts promote the
progression of endometrial cancer via the SDF-1/CXCR4 axis. J
Hematol Oncol. 9:82016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kuzet SE and Gaggioli C: Fibroblast
activation in cancer: When seed fertilizes soil. Cell Tissue Res.
365:607–619. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Neri S, Hashimoto H, Kii H, Watanabe H,
Masutomi K, Kuwata T, Date H, Tsuboi M, Goto K, Ochiai A, et al:
Cancer cell invasion driven by extracellular matrix remodeling is
dependent on the properties of cancer-associated fibroblasts. J
Cancer Res Clin Oncol. 142:437–446. 2016. View Article : Google Scholar
|
|
16
|
Stein U, Walther W, Arlt F, Schwabe H,
Smith J, Fichtner I, Birchmeier W and Schlag PM: MACC1, a newly
identified key regulator of HGF-MET signaling, predicts colon
cancer metastasis. Nat Med. 15:59–67. 2009. View Article : Google Scholar
|
|
17
|
Arlt F and Stein U: Colon cancer
metastasis: MACC1 and Met as metastatic pacemakers. Int J Biochem
Cell Biol. 41:2356–2359. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang L, Wu Y, Lin L, Liu P, Huang H, Liao
W, Zheng D, Zuo Q, Sun L, Huang N, et al: Metastasis-associated in
colon cancer-1 upregulation predicts a poor prognosis of gastric
cancer, and promotes tumor cell proliferation and invasion. Int J
Cancer. 133:1419–1430. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang Y, Zhang H, Cai J, Fang L, Wu J, Ye
C, Zhu X and Li M: Overexpression of MACC1 and Its significance in
human Breast Cancer Progression. Cell Biosci. 3:162013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ji D, Lu ZT, Li YQ, Liang ZY, Zhang PF, Li
C, Zhang JL, Zheng X and Yao YM: MACC1 expression correlates with
PFKFB2 and survival in hepatocellular carcinoma. Asian Pac J Cancer
Prev. 15:999–1003. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gao J, Ding F, Liu Q and Yao Y: Knockdown
of MACC1 expression suppressed hepatocellular carcinoma cell
migration and invasion and inhibited expression of MMP2 and MMP9.
Mol Cell Biochem. 376:21–32. 2013. View Article : Google Scholar
|
|
22
|
Shimokawa H, Uramoto H, Onitsuka T,
Chundong G, Hanagiri T, Oyama T and Yasumoto K: Overexpression of
MACC1 mRNA in lung adenocarcinoma is associated with postoperative
recurrence. J Thorac Cardiovasc Surg. 141:895–898. 2011. View Article : Google Scholar
|
|
23
|
Chundong G, Uramoto H, Onitsuka T,
Shimokawa H, Iwanami T, Nakagawa M, Oyama T and Tanaka F: Molecular
diagnosis of MACC1 status in lung adenocarcinoma by
immunohistochemical analysis. Anticancer Res. 31:1141–1145.
2011.PubMed/NCBI
|
|
24
|
Guo T, Kong J, Liu Y, Li Z, Xia J, Zhang
Y, Zhao S, Li F, Li J and Gu C: Transcriptional activation of NANOG
by YBX1 promotes lung cancer stem-like properties and metastasis.
Biochem Biophys Res Commun. 487:153–159. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang Z, Cai M, Weng Y, Zhang F, Meng D,
Song J, Zhou H and Xie Z: Circulating MACC1 as a novel diagnostic
and prognostic biomarker for nonsmall cell lung cancer. J Cancer
Res Clin Oncol. 141:1353–1361. 2015. View Article : Google Scholar
|
|
26
|
Yu T, Guo Z, Fan H, Song J, Liu Y, Gao Z
and Wang Q: Cancer-associated fibroblasts promote non-small cell
lung cancer cell invasion by upregulation of glucose-regulated
protein 78 (GRP78) expression in an integrated bionic microfluidic
device. Oncotarget. 7:25593–25603. 2016.PubMed/NCBI
|
|
27
|
Mueller MM and Fusenig NE: Friends or foes
- bipolar effects of the tumour stroma in cancer. Nat Rev Cancer.
4:839–849. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Friedl P, Noble PB, Walton PA, Laird DW,
Chauvin PJ, Tabah RJ, Black M and Zanker KS: Migration of
coordinated cell clusters in mesenchymal and epithelial cancer
explants in vitro. Cancer Res. 55:4557–4560. 1995.PubMed/NCBI
|
|
29
|
Friedl P, Locker J, Sahai E and Segall JE:
Classifying collective cancer cell invasion. Nat Cell Biol.
14:777–783. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Migliore C, Martin V, Leoni VP, Restivo A,
Atzori L, Petrelli A, Isella C, Zorcolo L, Sarotto I, Casula G, et
al: MiR-1 downregulation cooperates with MACC1 in promoting MET
overexpression in human colon cancer. Clin Cancer Res. 18:737–747.
2012. View Article : Google Scholar
|
|
31
|
Galimi F, Torti D, Sassi F, Isella C, Corà
D, Gastaldi S, Ribero D, Muratore A, Massucco P, Siatis D, et al:
Genetic and expression analysis of MET, MACC1, and HGF in
metastatic colorectal cancer: Response to met inhibition in patient
xenografts and pathologic correlations. Clin Cancer Res.
17:3146–3156. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen S, Zong ZH, Wu DD, Sun KX, Liu BL and
Zhao Y: The role of metastasis-associated in colon cancer 1 (MACC1)
in endometrial carcinoma tumorigenesis and progression. Mol
Carcinog. 56:1361–1371. 2017. View
Article : Google Scholar
|
|
33
|
Zhou L, Yu L, Zhu B, Wu S, Song W, Gong X
and Wang D: Metastasis-associated in colon cancer-1 and aldehyde
dehydrogenase 1 are metastatic and prognostic biomarker for
non-small cell lung cancer. BMC Cancer. 16:8762016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tan W, Xie X, Li L, Tang H, Ye X, Chen L,
Tang W, Gao J, Pan L, Zhang X, et al: Diagnostic and prognostic
value of serum MACC1 in breast cancer patients. Oncotarget.
7:84408–84415. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Y, Wang Z, Chen M, Peng L, Wang X,
Ma Q, Ma F and Jiang B: MicroRNA-143 targets MACC1 to inhibit cell
invasion and migration in colorectal cancer. Mol Cancer. 11:232012.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Guo T, Zhao S, Wang P, Xue X, Zhang Y,
Yang M, Li N, Li Z, Xu L, Jiang L, et al: YB-1 regulates tumor
growth by promoting MACC1/c-Met pathway in human lung
adenocarcinoma. Oncotarget. 8:48110–48125. 2017.PubMed/NCBI
|
|
37
|
Gaggioli C, Hooper S, Hidalgo-Carcedo C,
Grosse R, Marshall JF, Harrington K and Sahai E: Fibroblast-led
collective invasion of carcinoma cells with differing roles for
RhoGTPases in leading and following cells. Nat Cell Biol.
9:1392–1400. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wu X, Chen X, Zhou Q, Li P, Yu B, Li J, Qu
Y, Yan J, Yu Y, Yan M, et al: Hepatocyte growth factor activates
tumor stromal fibroblasts to promote tumorigenesis in gastric
cancer. Cancer Lett. 335:128–135. 2013. View Article : Google Scholar : PubMed/NCBI
|